Structure and Neuroprotective Effect of Polysaccharide from Viscera Autolysates of Squid Ommastrephes bartrami

 ,
, 
Abstract
:1. Introduction
2. Results
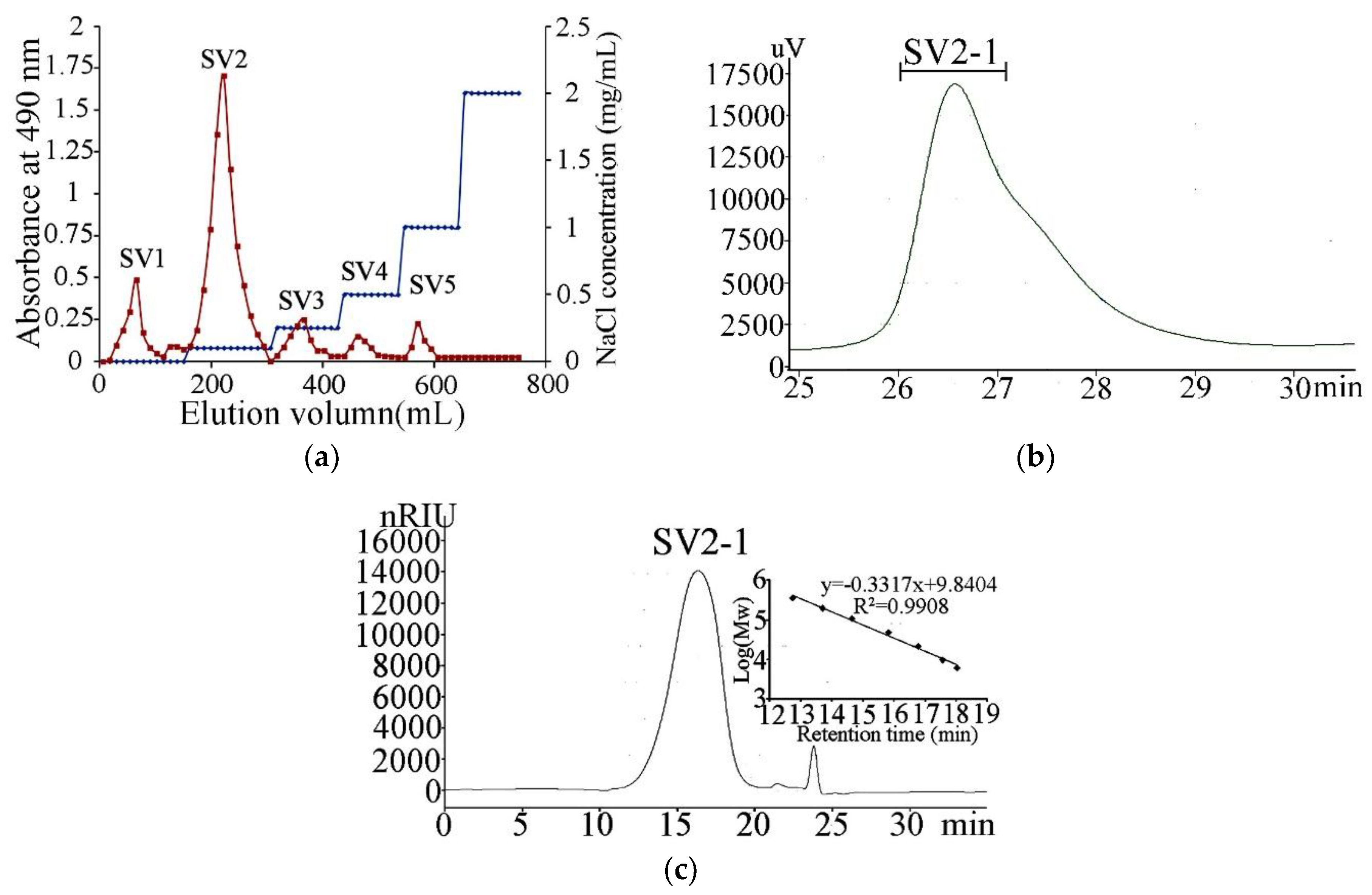
2.1. Extraction, Purification and Composition of the Polysaccharide
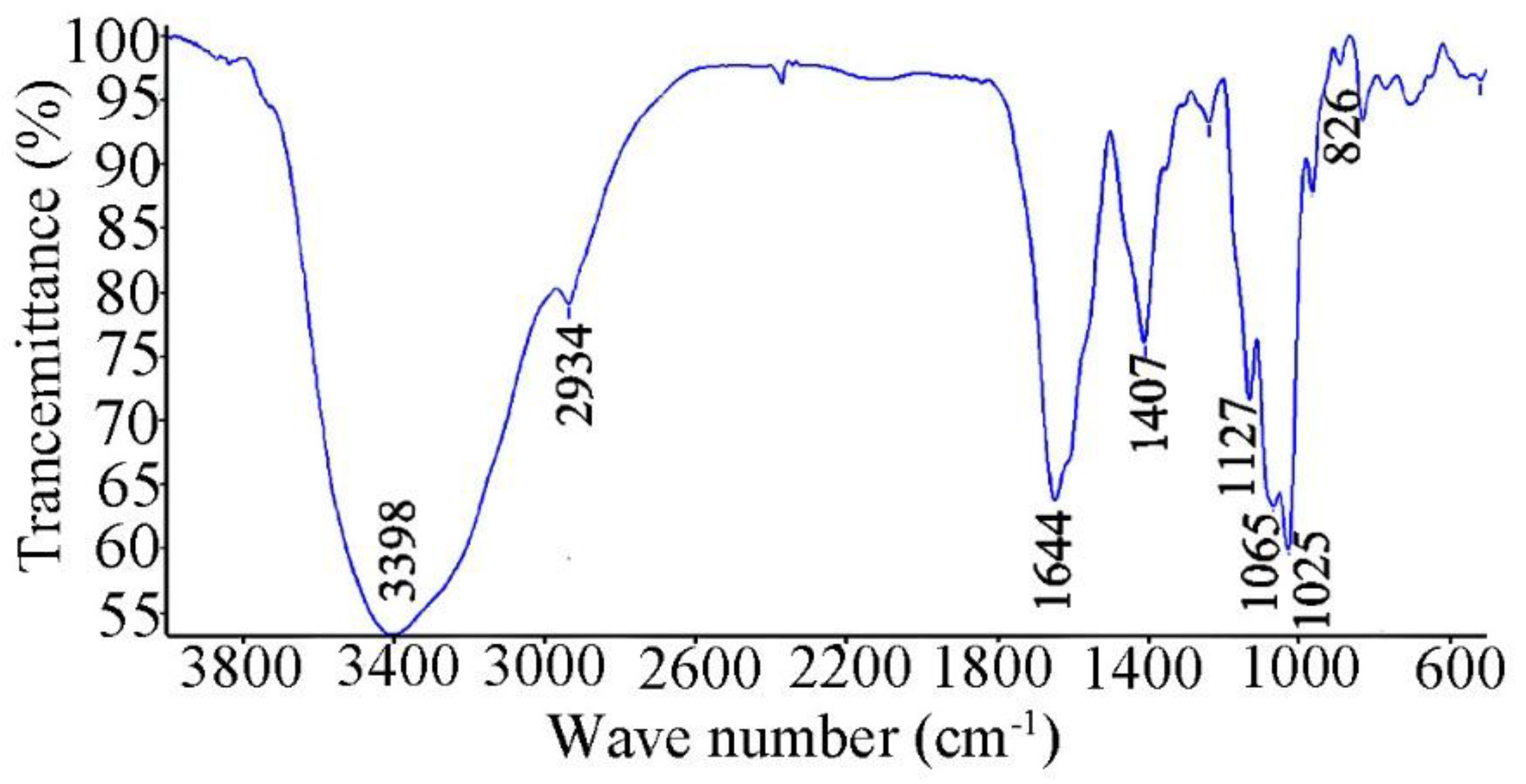
2.2. Infrared (IR)
2.3. Gas Chromatography-Mass Spectrometry (GC-MS) Analysis of the Methylated Products
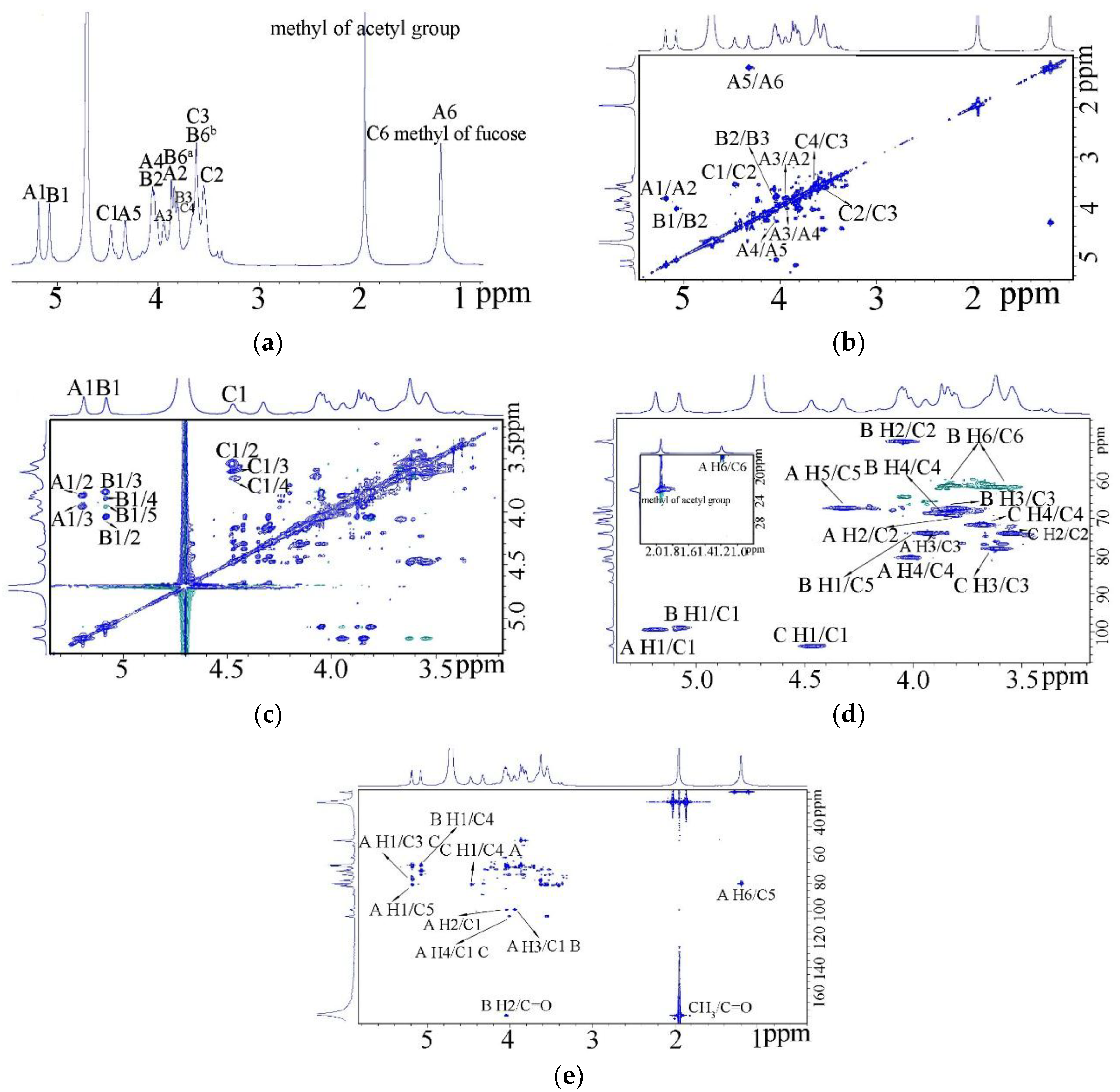
2.4. Nuclear Magnetic Resonance (NMR)
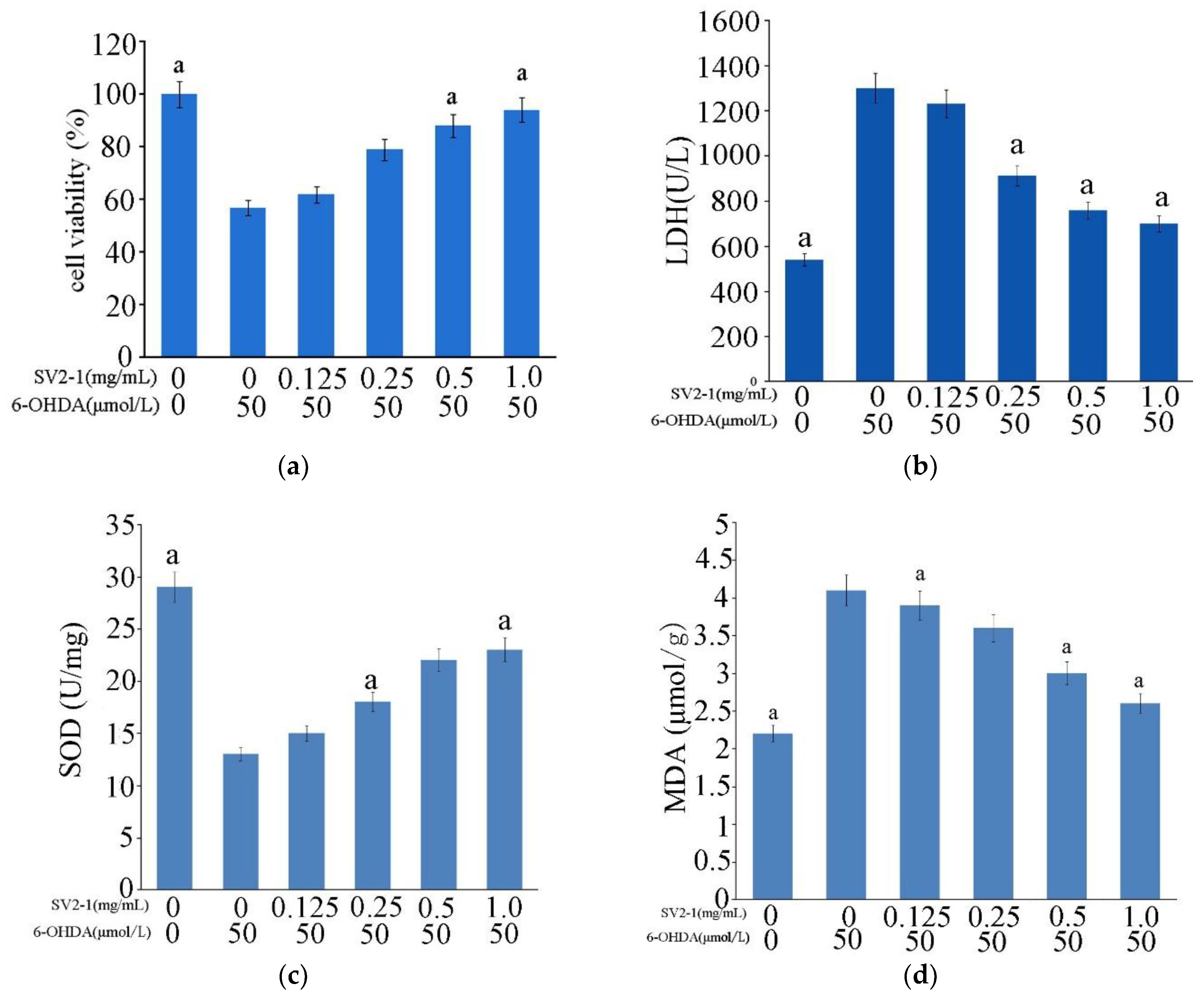
2.5. Neuroprotective Activity on PC12 Cell Line
3. Materials and Methods
3.1. Preparation of Polysaccharides from the Squid Viscera Autolysates
3.2. Purification of Polysaccharides from the Squid Viscera Autolysates
3.3. Physicochemical Characteristics and Composition Analysis of the Polysaccharide
3.4. IR Spectroscopy Analysis
3.5. Carboxyl Group Reduction and Methylation Analysis
3.6. NMR Spectroscopy
3.7. Neuroprotective Activity on PC12 Cell Line
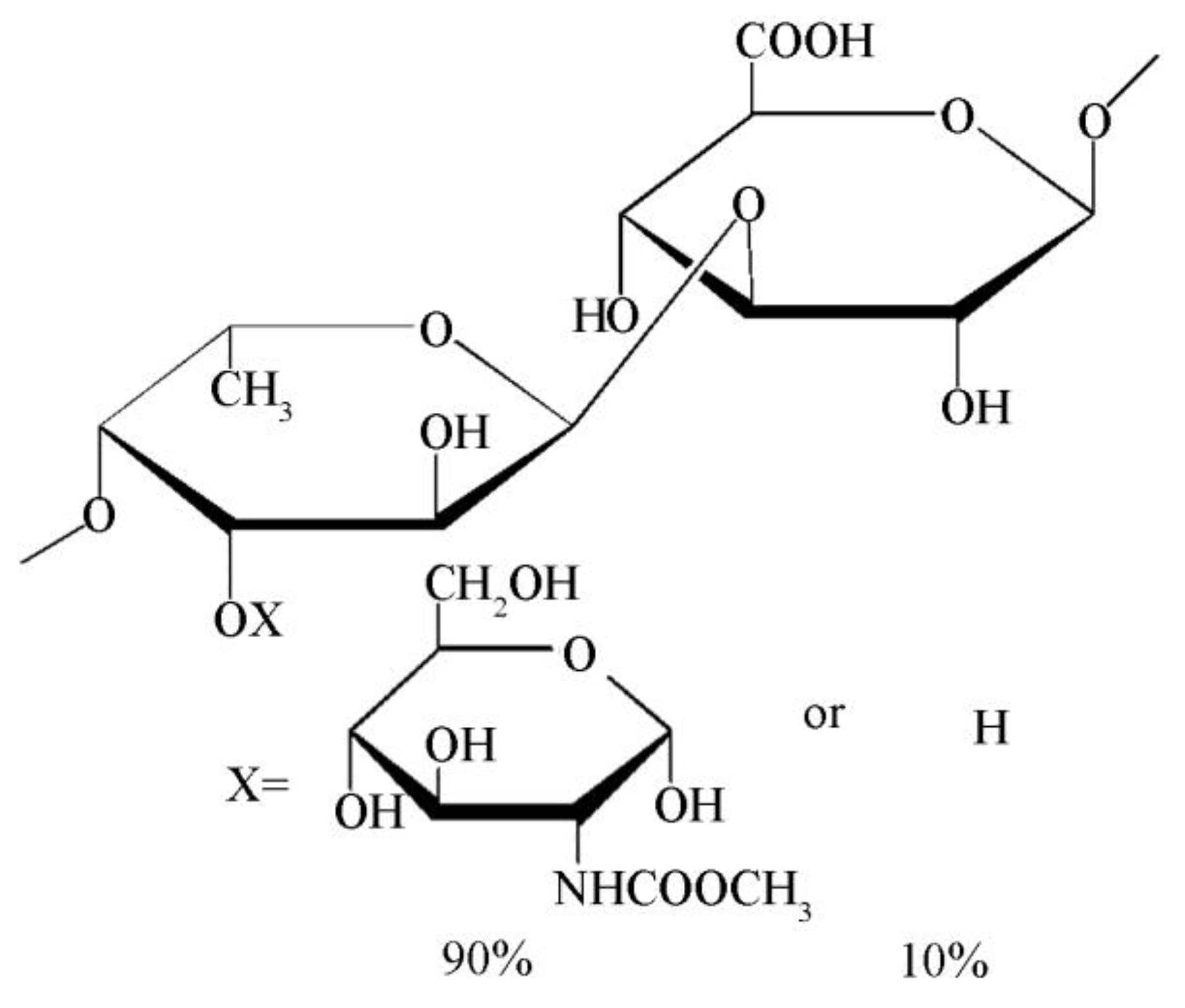
4. Conclusions and Discussion
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Apostolidis, E.; Karayannakidis, P.D.; Lee, C.M. Recovery of bioactive peptides and omega-3 fatty acids-containing phospholipids from squid processing by-product hydrolysate. J. Aquat. Food. Prod. T. 2015, 25, 496–506. [Google Scholar] [CrossRef]
- Arias-Moscoso, J.L.; Maldonado-Arce, A.; Rouzaud-Sandez, O.; Márquez-Ríos, E.; Torres-Arreola, W.; Santacruz-Ortega, H.; Gaxiola-Cortés, M.G.; Ezquerra-Brauer, J.M. Physicochemical characterization of protein hydrolysates Produced by autolysis of jumbo squid (Dosidicus gigas) byproducts. Food Biophys. 2014, 10, 145–154. [Google Scholar] [CrossRef]
- González-Félix, M.L.; Perez-Velazquez, M.; Ezquerra-Brauer, J.M.; Bringas-Alvarado, L.; Sánchez-Sánchez, A.; Torres-Arreola, W. Evaluation of jumbo squid (Dosidicus gigas) byproduct hydrolysates obtained by acid-enzymatic hydrolysis and by autohydrolysis in practical diets for Pacific white shrimp (Litopenaeus vannamei). Food Sci. Technol. (Campinas) 2014, 34, 552–558. [Google Scholar] [CrossRef]
- Wako, Y.; Ishikawa, S.; Muramoto, K. Angiotensin I-converting enzyme inhibitors in autolysates of squid liver and mantle muscle. Biosci. Biotech. Biochem. 1996, 60, 1353–1355. [Google Scholar] [CrossRef]
- Ru, S.; Zhang, K.Q.; Wei, R.B. In vitro antioxidative activities of squid (Ommastrephes bartrami) viscera autolysates and identification of active peptides. Process. Biochem. 2016, 51, 1674–1682. [Google Scholar] [CrossRef]
- Liu, C.; Li, X.; Li, Y.; Feng, Y.; Zhou, S.; Wang, F. Structural characterisation and antimutagenic activity of a novel polysaccharide isolated from Sepiella maindroni ink. Food Chem. 2008, 110, 807–813. [Google Scholar] [CrossRef]
- Chen, S.; Xu, J.; Xue, C.; Dong, P.; Sheng, W.; Yu, G.; Chai, W. Sequence determination of a non-sulfated glycosaminoglycan-like polysaccharide from melanin-free ink of the squid Ommastrephes bartrami by negative-ion electrospray tandem mass spectrometry and NMR spectroscopy. Glycoconj. J. 2008, 25, 481–492. [Google Scholar] [CrossRef]
- Shavandi, A.; Bekhit, E.D.A.; Sun, Z.; Ali, M.A. Injectable gel from squid pen chitosan for bone tissue engineering applications. J. Sol-Gel. Sci. Technol. 2016, 77, 675–687. [Google Scholar] [CrossRef]
- Uddin, M.S.; Hyangmin, A.; Kishimura, H.; Byungsoo, C. Production of valued materials from squid viscera by subcritical water hydrolysis. J. Environ. Biol. 2010, 31, 675. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhang, S.W.; Niu, Q.F.; Jin, X. The development andutilization of polysaccharide resources from marine cephalopods. Chin. J. Mar. Drugs 2014, 33, 87–92. [Google Scholar]
- Chen, S.; Xue, C.; Yin, L.A.; Tang, Q.; Guangli, Y.U.; Chai, W. Comparison of structures and anticoagulant activities of fucosylated chondroitin sulfates from different sea cucumbers. Carbohyd. Polym. 2011, 83, 688–696. [Google Scholar] [CrossRef]
- Wen, S.S.; Zhao, X.; Gao, Y.X.; Guang-Li, Y.U. Extraction, separation and physicochemical properties analyses of Octopus ocellatus polysaccharides. Chin. J. Mar. Drugs 2010, 1, 24–31. [Google Scholar]
- Li, P.; Wen, S.; Sun, K.; Zhao, Y.; Chen, Y. Structure and bioactivity screening of a low molecular weight ulvan from the green alga ulothrix flacca. Mar. Drugs 2018, 16, 281. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Lin, L.; Agyekum, I.; Zhang, X.; Ange, K.S.; Yu, Y.; Zhang, F.; Liu, J.; Amster, I.J.; Linhardt, R.J. Structural analysis of heparin-derived 3-O-sulfated tetrasaccharides: Antithrombin binding site variants. J. Pharm. Sci. 2016, 106, 973–981. [Google Scholar] [CrossRef]
- Pham Duc, T.; Menshova, R.V.; Ermakova, S.P.; Anastyuk, S.D.; Bui Minh, L.; Zvyagintseva, T.N. Structural characteristics and anticancer activity of fucoidan from the brown alga Sargassum mcclurei. Mar. Drugs 2013, 11, 1456–1476. [Google Scholar] [CrossRef]
- Chen, S.; Wang, J.; Xue, C.; Hui, L.; Sun, B.; Yong, X.; Chai, W. Sulfation of a squid ink polysaccharide and its inhibitory effect on tumor cell metastasis. Carbohydr. Polym. 2010, 81, 560–566. [Google Scholar] [CrossRef]
- Drouillard, S.; Jeacomine, I.; Buon, L.; Boisset, C.; Courtois, A.; Thollas, B.; Morvan, P.Y.; Vallée, R.; Helbert, W. Structure of the exopolysaccharide secreted by a marine strain Vibrio alginolyticus. Mar. Drugs 2018, 16, 164. [Google Scholar] [CrossRef]
- Shearman, M.S.; Ragan, C.I.; Iversen, L.L. Inhibition of PC12 cell redox activity is a specific, early indicator of the mechanism of beta-amyloid-mediated cell death. Proc. Natl. Acad. Sci. USA 1994, 91, 1470–1474. [Google Scholar] [CrossRef]
- Blum, D.; Wu, Y.; Nissou, M.F.; Arnaud, S.; Benabid, A.L.; Verna, J.M. p53 and Bax activation in 6-hydroxydopamine-induced apoptosis in PC12 cells. Brain Res. 1997, 751, 139–142. [Google Scholar] [CrossRef]
- Cui, C.; Song, S.L.; Liang, H.; Wang, P. Sulfated polysaccharide isolated from the sea Cucumber stichopus japonicus against PC12 Hypoxia/Reoxygenation injury by inhibition of the MAPK signaling pathway. Cell Mol. Neurobiol. 2015, 35, 1081–1092. [Google Scholar] [CrossRef]
- Ciancarelli, I.; Massimo, C.D.; Amicis, D.D.; Pistarini, C.; Ciancarelli, M.G. Uric acid and Cu/Zn superoxide dismutase: Potential strategies and biomarkers in functional recovery of post-acute ischemic stroke patients after intensive neurorehabilitation. Curr. Neurovasc. Res. 2015, 12, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Naziroğlu, M.; Kutluhan, S.; Uğuz, A.C.; Celik, O.; Bal, R.; Butterworth, P.J. Topiramate and vitamin e modulate the electroencephalographic records, brain microsomal and blood antioxidant redox system in pentylentetrazol-induced seizure of rats. J. Membr. Biol. 2009, 229, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Zhao, J.; Yu, Y.; Liu, X.; Lin, L.; Zhang, F.; Linhardt, R.J. Antithrombin III binding site analysis of low molecular weight heparin fractions. J. Pharm. Sci. 2018, 107, 1290–1295. [Google Scholar] [CrossRef] [PubMed]
- Sun, K.; Chen, Y.; Niu, Q.; Zhu, W.; Wang, B.; Li, P.; Ge, X. An exopolysaccharide isolated from a coral-associated fungus and its sulfated derivative activates macrophages. Int. J. Biol. Macromol. 2016, 82, 387–394. [Google Scholar] [CrossRef]
- Malson, T. Material of Polysaccharides Containing Carboxyl Groups, and a Process for Producing such Polysaccharides. U.S. Patent 4,963,666A, 16 October 1987. [Google Scholar]
- Chen, Y.; Mao, W.J.; Wang, H.; Zhu, W.M.; Niu, Q.F.; Fang, X.B.; Li, T. Structure elucidation of a galactofuranose-rich heteropolysaccharide from aciduric fungus penicillium purpurogenum JS03-21. Glycoconj. J. 2015, 32, 29–37. [Google Scholar] [CrossRef] [PubMed]
- Sims, I.M.; Carnachan, S.M.; Bell, T.J.; Hinkley, S.F.R. Methylation analysis of polysaccharides: Technical advice. Carbohydr. Polym. 2018, 188, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Mao, W.J.; Yan, M.X.; Liu, X.; Wang, S.Y.; Xia, Z.; Xiao, B.; Cao, S.J.; Yang, B.F.; Li, J. Purification, chemical characterization, and bioactivity of an extracellular polysaccharide produced by the marine sponge endogenous fungus Alternaria sp. SP-32. Mar. Biotechnol. 2016, 18, 301–313. [Google Scholar] [CrossRef] [PubMed]
- Myrona, P.; Siddiquee, S.; Azada, S. Fucosylated chondroitin sulfate diversity in sea cucumbers: A review. Carbohydr. Polym. 2014, 112, 173–178. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Methylation Product | Main MS (m/z) | Molar Ratio (%) | Linkage Pattern | |
|---|---|---|---|---|
| SV2-1 | SV2-1P | |||
| 1,4,5-tri-O-acetyl-2,3,6-tri-O-methyl-l-fucitol | 101,117,143,203 | 1 | 67 | →4)Fuc(1→ |
| 1,3,4,5-tetra-O-acetyl-2,6-di-O-methyl-l-fucitol | 113,117,129,173,275 | 6 | 15 | →3,4)Fuc(1→ |
| 1,3,5-tri-O-acetyl-2,4,6-tri-O-methyl-d-gluctitol | 101,117,129,161,233 | - | 8 | →3)Glc(1→ |
| Residue | H1/C1 | H2/C2 | H3/C3 | H4/C4 | H5/C5 | H6/C6 | ||
|---|---|---|---|---|---|---|---|---|
| A Fuc | 5.19 | 3.84 | 3.95 | 4.03 | 4.33 | 1.19 | ||
| 99.2 | 68.4 | 75.1 | 82 | 66.5 | 15.9 | |||
| B GalNAc | 5.08 | 4.03 | 3.80 | 3.87 | 3.95 | 3.80, 3.61 | CH3 | C=O |
| 1.96 | 174 | |||||||
| 98.8 | 50.2 | 67.5 | 68.8 | 74.5 | 61.2 | 22.0 | ||
| C GlcUA | 4.47 | 3.55 | 3.62 | 3.66 | -/- | -/174 | ||
| 103.9 | 74 | 78 | 70.5 | |||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ye, P.; Li, P.; Yang, W.; Zhao, Y.; Zhao, Y.; Sun, K.; Wang, B.; Chen, Y. Structure and Neuroprotective Effect of Polysaccharide from Viscera Autolysates of Squid Ommastrephes bartrami. Mar. Drugs 2019, 17, 188. https://doi.org/10.3390/md17030188
Ye P, Li P, Yang W, Zhao Y, Zhao Y, Sun K, Wang B, Chen Y. Structure and Neuroprotective Effect of Polysaccharide from Viscera Autolysates of Squid Ommastrephes bartrami. Marine Drugs. 2019; 17(3):188. https://doi.org/10.3390/md17030188
Chicago/Turabian StyleYe, Peng, Peipei Li, Wenge Yang, Yue Zhao, Yuqin Zhao, Kunlai Sun, Bin Wang, and Yin Chen. 2019. "Structure and Neuroprotective Effect of Polysaccharide from Viscera Autolysates of Squid Ommastrephes bartrami" Marine Drugs 17, no. 3: 188. https://doi.org/10.3390/md17030188
APA StyleYe, P., Li, P., Yang, W., Zhao, Y., Zhao, Y., Sun, K., Wang, B., & Chen, Y. (2019). Structure and Neuroprotective Effect of Polysaccharide from Viscera Autolysates of Squid Ommastrephes bartrami. Marine Drugs, 17(3), 188. https://doi.org/10.3390/md17030188