Metabolites of Seaweeds as Potential Agents for the Prevention and Therapy of Influenza Infection


 ,
,
Abstract
:1. Introduction
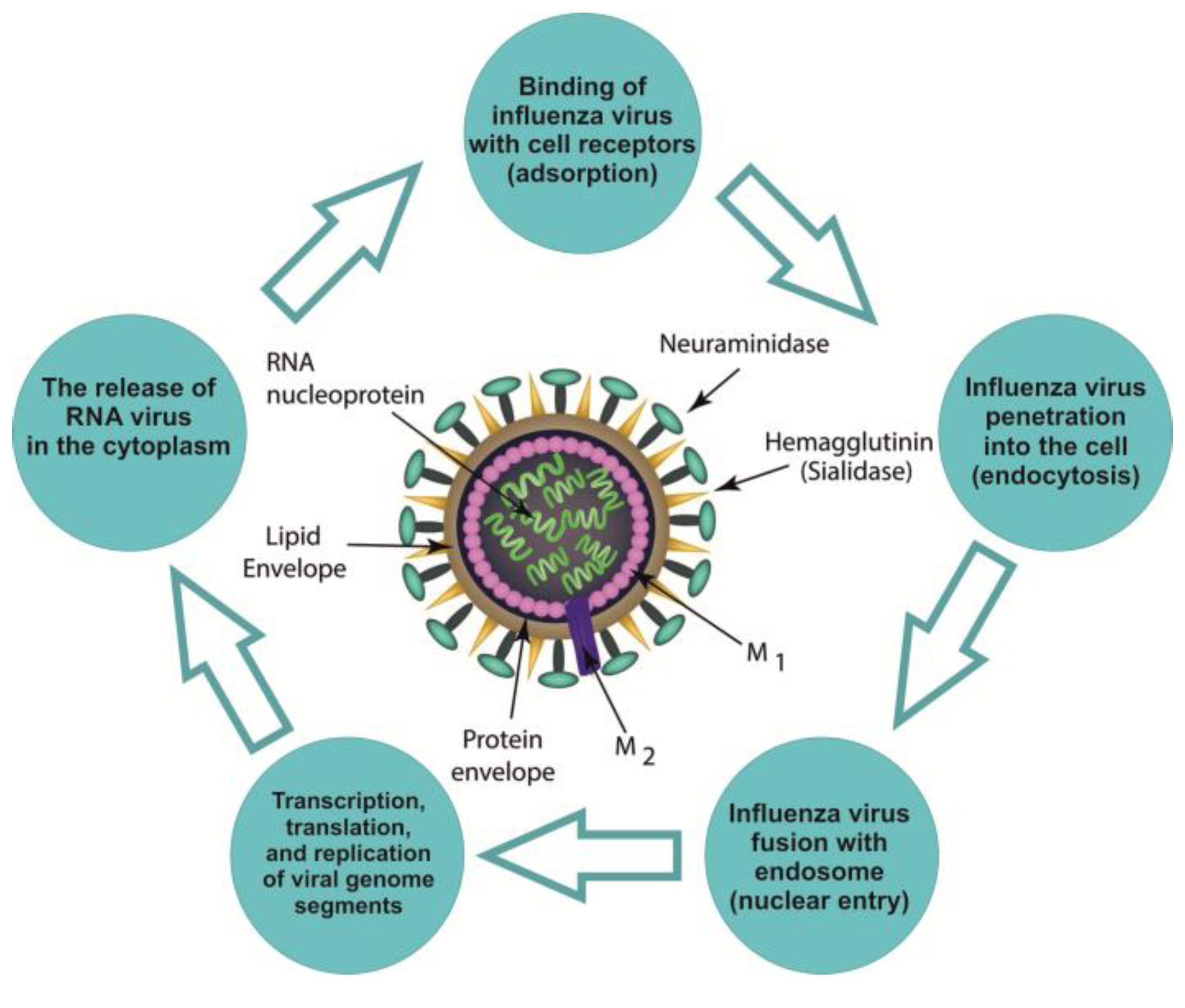
2. Influenza Virion
3. Pathogenic Targets of Influenza Viruses in Humans
4. Recent Anti-Influenza Drugs
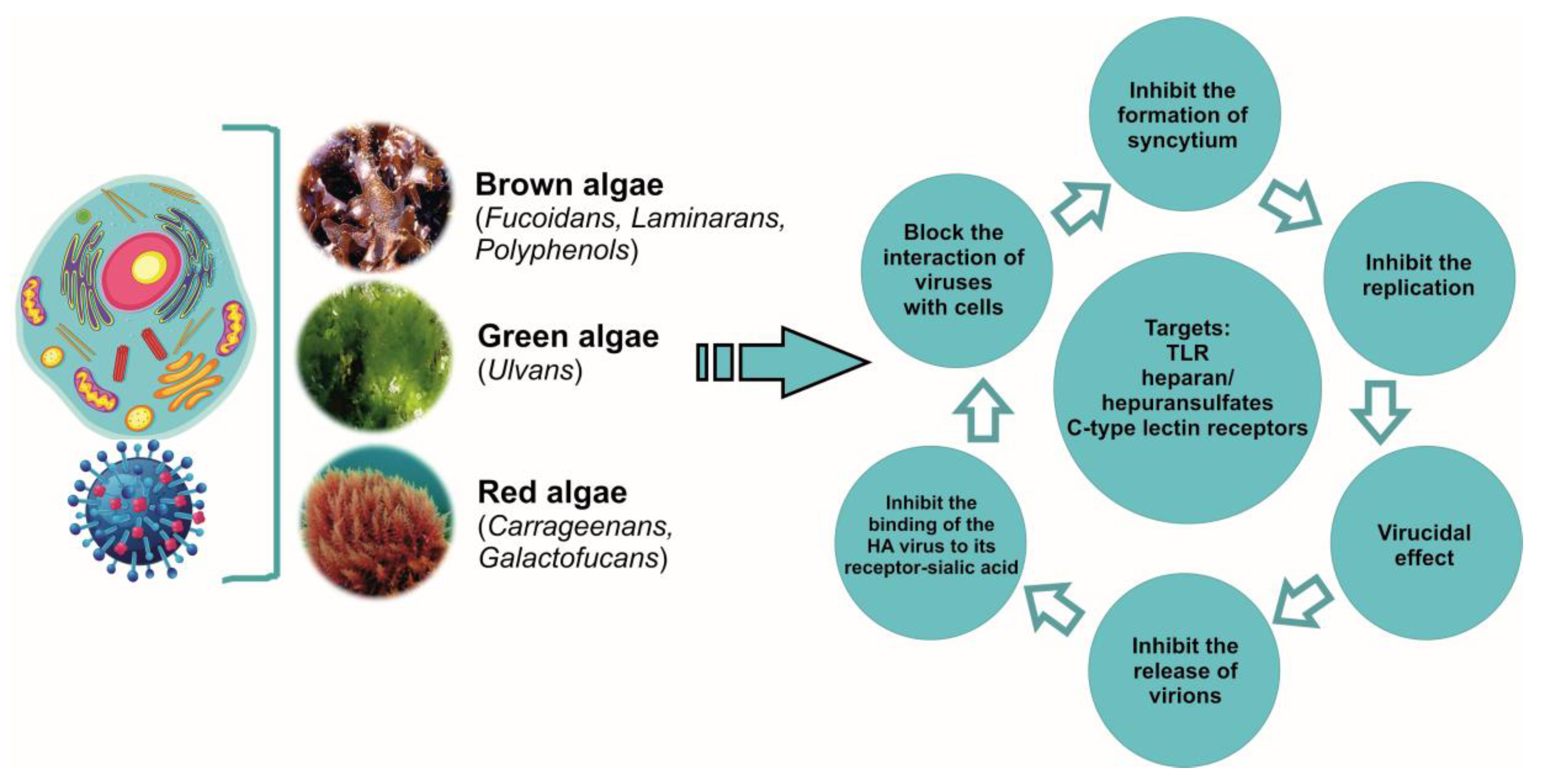
5. Polysaccharides
5.1. Carrageenans
5.2. Fucoidans
6. Lectins
7. Polyphenols of Algae
8. Biopolymers of Algae Are Adjuvants for the Influenza Vaccines
9. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Lvov, N.I.; Likhopoenko, V.P. Acute respiratory infections. Guide to infectious diseases. StP: Foliant 2011, 2, 7–122. [Google Scholar]
- Osidak, L.V.; Drinevskiy, V.P.; Erofeeva, M.K.; Eropkin, M.Y.; Konovalov, N.I.; Smororintsev, E.A.; Golovacheva, E.G.; Donluray, E.A.; Baybus, A.M.; Wojciechowska, E.M.; et al. Influenza A (H1N1) 2009 in Russia. Terra Medica Nova. Infect. Dis. 2009, 4–5, 6–9. (In Rissian) [Google Scholar]
- Henrickson, K.J.; Hoover, S.; Kehl, K.S.; Hua, W. National disease burden of respiratory viruses detected in children by polymerase chain reaction. Pediatr. Infect. Dis. J. 2004, 23 (Suppl. 1), S11–S18. [Google Scholar] [CrossRef]
- Vasin, A.V.; Sologub, T.V.; Tsybalova, L.M.; Tokin, I.I.; Tsvetkov, V.V. Influenza in the practice of a clinician, an epidemiologist and a virologist. Med. Inf. Agency Moscow 2017, 272. (In Russian) [Google Scholar]
- Sologub, T.V. Influenza in modern conditions. Med. Council 2015, 4, 36–45. (In Russian) [Google Scholar]
- Bilichenko, T.N.; Bystritskaya, E.V.; Chuchalin, A.G.; Belevsky, A.S.; Batyn, S.Z. Mortality from respiratory diseases in years 2014-2015 and ways to reduce it. Pulmonologiya 2016, 26, 389–397. (In Russian) [Google Scholar] [CrossRef]
- Arias, C.F.; Escalera-Zamudio, M.; Soto-Del Rio Mde, L.; Cobian-Guemes, A.G.; Isa, P.; Lopez, S. Molecular anatomy of 2009 influenza virus A (H1N1). Arch. Med. Res. 2009, 40, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Gill, J.R.; Sheng, Z.M.; Ely, S.F.; Guinee, D.G.; Beasley, M.B.; Suh, J.; Deshpande, C.; Mollura, D.J.; Morens, D.M.; Bray, M.; et al. Pulmonary pathologic findings of fatal 2009 pandemic influenza A/H1N1 viral infections. Arch. Pathol. Lab. Med. 2010, 134, 235–243. [Google Scholar]
- Bilichenko, T.N. Influence of year 2016. Med. Council 2016, 15, 52–57. (In Russian) [Google Scholar] [CrossRef]
- Mei, L.; Song, P.; Tang, Q.; Shan, K.; Tobe, R.G.; Selotlegeng, L.; Ali, A.H.; Cheng, Y.; Xu, L. Changes in and shortcomings of control strategies, drug stockpiles, and vaccine development during outbreaks of avian influenza A H5N1, H1N1, and H7N9 among humans. Biosci. Trends 2013, 7, 64–76. [Google Scholar] [CrossRef] [PubMed]
- Medina, R.A.; Garcia-Sastre, A. Influenza A viruses: New research developments. Nat. Rev. Microbiol. 2011, 9, 590–603. [Google Scholar] [CrossRef]
- Imbs, T.I.; Schevchenko, N.M.; Zvyagintseva, T.N. Structure, enzymatic transformation and biological properties. In Fucoidans - Sulfated Polysaccharides of Brown Algae; Dal’nauka FED RAS: Vladivostok, Russia, 2014; pp. 95–110. [Google Scholar]
- Norkin, L.C. Orthomyxoviruses. In Virology, Molecular Biology and Pathogenesis; Asm Press American Society Microbiol: Washington, DC, USA, 2010; pp. 296–345. [Google Scholar]
- Matusevich, O.V.; Gluzdikov, I.A.; Titov, M.I. Synthesis of PB1 RNA polymerase subunit fragments. Vestnik St. Petersburg State Univ. 2011, 2, 150–159. (In Russian) [Google Scholar]
- Zarubaev, V.V.; Smirnov, V.S. Impact on Cellular Targets as a Means of Combating Influenza Infection. 2014. Available online: https://doi.org/10.15789/2220-7619-2014-1-15-26 (accessed on 22 June 2019).
- Nobusawa, E.; Sato, K. Comparison of the mutation rates of human influenza A and B viruses. J. Virol. 2006, 80, 3675–3678. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.B.; Inglis, S.C. The mutation rate and variability of eukaryotic viruses: An analytical review. J. Gen. Virol. 1987, 68 (Pt 11), 2729–2740. [Google Scholar] [CrossRef]
- Drake, J.W.; Charlesworth, B.; Charlesworth, D.; Crow, J.F. Rates of spontaneous mutation. Genetics 1998, 148, 1667–1686. [Google Scholar] [PubMed]
- Hay, A.J.; Gregory, V.; Douglas, A.R.; Lin, Y.P. The evolution of human influenza viruses. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2001, 356, 1861–1870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tate, M.D.; Job, E.R.; Deng, Y.M.; Gunalan, V.; Maurer-Stroh, S.; Reading, P.C. Playing hide and seek: How glycosylation of the influenza virus hemagglutinin can modulate the immune response to infection. Viruses 2014, 6, 1294–1316. [Google Scholar] [CrossRef]
- Tsvetkov, V.V.; Golobokov, G.S. Neuraminidase inhibitors - the gold standard for antiviral therapy for influenza A. Med. Adv. 2017, 4, 25–30. (In Russian) [Google Scholar] [CrossRef]
- Kovalev, N.A.; Krasochko, P.A. Viruses and Prions in the Pathology of Animals and Humans. 2012. Available online: https://www.litres.ru/n-a-kovalev/virusy-i-priony-v-patologii-zhivotnyh-i-cheloveka-7063392 (accessed on 22 June 2019).
- Sergeeva, I.V.; Demko, I.V. Features of the Flu and Viral-Bacterial Pneumonia. 2017. Available online: https://www.monographies.ru/ru/book/view?id=686 (accessed on 22 June 2019).
- Kaverin, N.V.; Rudneva, I.A.; Timofeeva, T.A. Antigenic structure of influenza A hemagglutinin. Vopr. Virol. 2012, 1, 148–158. [Google Scholar]
- Desheva, Y.A.; Smolonogina, T.A.; Rudenko, L.G. Biological and protective properties of serum antibodies to influenza neuraminidase virus. Med. Acad. J. 2015, 15, 35–44. (In Russian) [Google Scholar]
- Glezen, W.P. Emerging infections: Pandemic influenza. Epidemiol. Rev. 1996, 18, 64–76. [Google Scholar] [CrossRef]
- Ruigrok, R.W. Structure of Influenza A, B and C Viruses. In Textbook of Influenza; Webster, R., Monto, A., Braciale, T., Lamb, R., Eds.; Blackwell Scientific Publications: Hoboken, NJ, USA, 2007. [Google Scholar] [CrossRef]
- Nayak, D.P.; Balogun, R.A.; Yamada, H.; Zhou, Z.H.; Barman, S. Influenza virus morphogenesis and budding. Virus Res. 2009, 143, 147–161. [Google Scholar] [CrossRef] [PubMed]
- Lapidus, N.I. School of therapist. SARS: Rational Pharmacotherapy. Medi. Sov. 2014, 16, 48–54. (In Russian) [Google Scholar]
- Sologub, T.V.; Ledvanov, M.Y.; Malyi, V.P.; Stukova, N.Y.; Romantsov, M.G.; Bizenkova, M.N.; Polyakova, T.D. Flu. Clinical symptoms. 2009. Available online: http://www.natural-sciences.ru/ru/article/view?id=14060 (accessed on 22 June 2019).
- Lee, N.; Wong, C.K.; Hui, D.S.; Lee, S.K.; Wong, R.Y.; Ngai, K.L.; Chan, M.C.; Chu, Y.J.; Ho, A.W.; Lui, G.C.; et al. Role of human Toll-like receptors in naturally occurring influenza A infections. Influenza Other Respir. Viruses 2013, 7, 666–675. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Londrigan, S.L.; Tate, M.D.; Brooks, A.G.; Reading, P.C. Cell-surface receptors on macrophages and dendritic cells for attachment and entry of influenza virus. J. Leukoc. Biol. 2012, 92, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Kok, K.H.; Jaume, M.; Cheung, T.K.; Yip, T.F.; Lai, J.C.; Guan, Y.; Webster, R.G.; Jin, D.Y.; Peiris, J.S. Toll-like receptor 10 is involved in induction of innate immune responses to influenza virus infection. Proc. Natl. Acad. Sci. USA 2014, 111, 3793–3798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weber, M.; Gawanbacht, A.; Habjan, M.; Rang, A.; Borner, C.; Schmidt, A.M.; Veitinger, S.; Jacob, R.; Devignot, S.; Kochs, G.; et al. Incoming RNA virus nucleocapsids containing a 5'-triphosphorylated genome activate RIG-I and antiviral signaling. Cell Host Microbe 2013, 13, 336–346. [Google Scholar] [CrossRef] [PubMed]
- Davidson, S. Treating Influenza Infection, From Now and Into the Future. Front. Immunol. 2018, 9, 1946. [Google Scholar] [CrossRef]
- Marty, F.M.; Vidal-Puigserver, J.; Clark, C.; Gupta, S.K.; Merino, E.; Garot, D.; Chapman, M.J.; Jacobs, F.; Rodriguez-Noriega, E.; Husa, P.; et al. Intravenous zanamivir or oral oseltamivir for hospitalised patients with influenza: An international, randomised, double-blind, double-dummy, phase 3 trial. Lancet Respir. Med. 2017, 5, 135–146. [Google Scholar] [CrossRef]
- Leibbrandt, A.; Meier, C.; Konig-Schuster, M.; Weinmullner, R.; Kalthoff, D.; Pflugfelder, B.; Graf, P.; Frank-Gehrke, B.; Beer, M.; Fazekas, T.; et al. Iota-carrageenan is a potent inhibitor of influenza A virus infection. PLoS ONE 2010, 5, e14320. [Google Scholar] [CrossRef]
- Jefferson, T.; Jones, M.; Doshi, P.; Del Mar, C. Neuraminidase inhibitors for preventing and treating influenza in healthy adults: systematic review and meta-analysis. BMJ 2009, 339, b5106. [Google Scholar] [CrossRef]
- Nelson, M.I.; Simonsen, L.; Viboud, C.; Miller, M.A.; Holmes, E.C. The origin and global emergence of adamantane resistant A/H3N2 influenza viruses. Virology 2009, 388, 270–278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sologub, T.V.; Tokin, I.I. Tactics of flu patients at the present stage. Effect. Pharm. 2017, 10, 14–19. (In Russian) [Google Scholar]
- Imbs, T.I.; Kharlamenko, V.I.; Zvyagintseva, T.N. Optimization of the extraction process of fucoidan from the brown alga Fucus evanescens. Chem. Plant Raw Mater. 2012, 1, 143–147. (In Russian) [Google Scholar]
- Hook, M.; Kjellen, L.; Johansson, S. Cell-surface glycosaminoglycans. Annu. Rev. Biochem. 1984, 53, 847–869. [Google Scholar] [CrossRef] [PubMed]
- Sheng, G.J.; Oh, Y.I.; Chang, S.K.; Hsieh-Wilson, L.C. Tunable heparan sulfate mimetics for modulating chemokine activity. J. Am. Chem. Soc. 2013, 135, 10898–10901. [Google Scholar] [CrossRef]
- Wang, W.; Wang, S.X.; Guan, H.S. The antiviral activities and mechanisms of marine polysaccharides: An overview. Mar. Drugs 2012, 10, 2795–2816. [Google Scholar] [CrossRef]
- Kusaykin, M.I.; Zvyagintseva, T.N. Structure, enzymatic transformation and biological properties. In Fucoidans - Sulfated Polysaccharides of Brown Algae; Dal’nauka FED RAS: Vladivostok, Russia, 2014; pp. 35–60. [Google Scholar]
- Yermak, I.M.; Barabanova, A.O.; Sokolova, E.V. Structural features and biological activity of carrageenans - sulfated polysaccharides of red algae of the Far Eastern seas. Vestn. FEB RAS 2014, 1, 25–36. (In Russian) [Google Scholar]
- Knutsen, S.H.; Myslabodsky, D.E.; Larsen, B.; Usov, A.I. A modified system of nomenclature for red algal galactans. Botanica Mar. 1994, 37, 163–169. [Google Scholar] [CrossRef]
- McKim, J.M., Jr.; Baas, H.; Rice, G.P.; Willoughby, J.A., Sr.; Weiner, M.L.; Blakemore, W. Effects of carrageenan on cell permeability, cytotoxicity, and cytokine gene expression in human intestinal and hepatic cell lines. Food Chem. Toxicol. 2016, 96, 1–10. [Google Scholar] [CrossRef]
- Yermak, I.M.; Davydova, V.N.; Aminin, D.L.; Barabanova, A.O.; Sokolova, E.V.; Bogdanovich, R.N.; Polyakova, A.M.; Solovyova, T.F. Immunomodulating activity of carrageenans from red algae of the Far Eastern seas. Pac. Med. J. 2009, 3, 40–45. (In Russian) [Google Scholar]
- Holdt, S.L.; Kraan, S. Bioactive compounds in seaweed: Functional food applications and legislation. J. Appl. Phycol. 2011, 23, 543–597. [Google Scholar] [CrossRef]
- Maksema, I.G.; Kompanets, G.G.; Barabanova, A.O.; Ermak, I.M.; Slonova, R.A. Antiviral effect of carrageenans from red algae during experimental hantavirus infection. Pac. Med. J. 2011, 1, 32–34. (In Russian) [Google Scholar]
- Necas, J.; Bartosikova, L. Carrageenan: A review. Vet. Med. 2013, 58, 187–205. [Google Scholar] [CrossRef]
- Bhattacharyya, S.; Liu, H.; Zhang, Z.; Jam, M.; Dudeja, P.K.; Michel, G.; Linhardt, R.J.; Tobacman, J.K. Carrageenan-induced innate immune response is modified by enzymes that hydrolyze distinct galactosidic bonds. J. Nutr. Biochem. 2010, 21, 906–913. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, L.S.; Fidelis, G.P.; Cordeiro, S.L.; Oliveira, R.M.; Sabry, D.A.; Camara, R.B.; Nobre, L.T.; Costa, M.S.; Almeida-Lima, J.; Farias, E.H.; et al. Biological activities of sulfated polysaccharides from tropical seaweeds. Biomed. Pharmacother. 2010, 64, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Damonte, E.B.; Matulewicz, M.C.; Cerezo, A.S. Sulfated seaweed polysaccharides as antiviral agents. Curr. Med. Chem. 2004, 11, 2399–2419. [Google Scholar] [CrossRef]
- Eccles, R.; Meier, C.; Jawad, M.; Weinmullner, R.; Grassauer, A.; Prieschl-Grassauer, E. Efficacy and safety of an antiviral Iota-Carrageenan nasal spray: A randomized, double-blind, placebo-controlled exploratory study in volunteers with early symptoms of the common cold. Respir. Res. 2010, 11, 108. [Google Scholar] [CrossRef]
- Ludwig, M.; Enzenhofer, E.; Schneider, S.; Rauch, M.; Bodenteich, A.; Neumann, K.; Prieschl-Grassauer, E.; Grassauer, A.; Lion, T.; Mueller, C.A. Efficacy of a carrageenan nasal spray in patients with common cold: A randomized controlled trial. Respir. Res. 2013, 14, 124. [Google Scholar] [CrossRef]
- Onari, Y.; Yokoyama, A.; Haruta, Y.; Nakashima, T.; Iwamoto, H.; Hattori, N.; Kohno, N. IL-12p40 is essential for the down-regulation of airway hyperresponsiveness in a mouse model of bronchial asthma with prolonged antigen exposure. Clin. Exp. Allergy 2009, 39, 290–298. [Google Scholar] [CrossRef]
- Rohde, G. Drug targets in rhinoviral infections. Infect. Disord. Drug Targets 2009, 9, 126–132. [Google Scholar] [CrossRef]
- Tregoning, J.S.; Schwarze, J. Respiratory viral infections in infants: causes, clinical symptoms, virology, and immunology. Clin. Microbiol. Rev. 2010, 23, 74–98. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhang, P.; Hao, C.; Zhang, X.E.; Cui, Z.Q.; Guan, H.S. In vitro inhibitory effect of carrageenan oligosaccharide on influenza A H1N1 virus. Antivir. Res. 2011, 92, 237–246. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wu, J.; Zhang, X.; Hao, C.; Zhao, X.; Jiao, G.; Shan, X.; Tai, W.; Yu, G. Inhibition of influenza A virus infection by fucoidan targeting viral neuraminidase and cellular EGFR pathway. Sci. Rep. 2017, 7, 40760. [Google Scholar] [CrossRef] [PubMed]
- Shao, Q.; Guo, Q.; Xu, W.; Li, Z.; Zhao, T. Specific Inhibitory Effect of kappa-Carrageenan Polysaccharide on Swine Pandemic 2009 H1N1 Influenza Virus. PLoS ONE 2015, 10, e0126577. [Google Scholar]
- Kim, M.; Yim, J.H.; Kim, S.Y.; Kim, H.S.; Lee, W.G.; Kim, S.J.; Kang, P.S.; Lee, C.K. In vitro inhibition of influenza A virus infection by marine microalga-derived sulfated polysaccharide p-KG03. Antivir. Res. 2012, 93, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Yu, G.; Li, M.; Wang, W.; Li, M.; Zhao, X.; Youjing, L.V.; Li, G.; Jiao, J.; Zhao, X. Structure and anti-Influenza A (H1N1) virus activity of three polysaccharides from Eucheuma denticulatum. J. Ocean Univ. China 2012, 11, 527–532. [Google Scholar] [CrossRef]
- Fazekas, T.; Eickhoff, P.; Pruckner, N.; Vollnhofer, G.; Fischmeister, G.; Diakos, C.; Rauch, M.; Verdianz, M.; Zoubek, A.; Gadner, H.; et al. Lessons learned from a double-blind randomised placebo-controlled study with a iota-carrageenan nasal spray as medical device in children with acute symptoms of common cold. BMC Complement. Altern. Med. 2012, 12, 147. [Google Scholar] [CrossRef] [PubMed]
- Koenighofer, M.; Lion, T.; Bodenteich, A.; Prieschl-Grassauer, E.; Grassauer, A.; Unger, H.; Mueller, C.A.; Fazekas, T. Carrageenan nasal spray in virus confirmed common cold: Individual patient data analysis of two randomized controlled trials. Multidiscip. Respir. Med. 2014, 9, 57. [Google Scholar] [CrossRef]
- Morokutti-Kurz, M.; Konig-Schuster, M.; Koller, C.; Graf, C.; Graf, P.; Kirchoff, N.; Reutterer, B.; Seifert, J.M.; Unger, H.; Grassauer, A.; et al. The Intranasal Application of Zanamivir and Carrageenan Is Synergistically Active against Influenza A Virus in the Murine Model. PLoS ONE 2015, 10, e0128794. [Google Scholar] [CrossRef]
- Graf, C.; Bernkop-Schnurch, A.; Eqyed, A.; Coller, C.; Prieschl-Grassauer, A.; Morokutti-Kurz, M. Development of a nasal spray containing xylometazoline hydrochloride and iota-carrageenan for the symptomatic relief of nasal congestion caused by rhinitis and sinusitis. Int. J. Gen. Med. 2018, 11, 275–283. [Google Scholar] [CrossRef]
- Gubina-Vakyulyk, G.I.; Gorbach, T.V.; Tkachenko, A.S.; Tkachenko, M.O. Damage and regeneration of small intestinal enterocytes under the influence of carrageenan induces chronic enteritis. Comparative Clin. Pathol. 2015, 24, 1473–1477. [Google Scholar] [CrossRef]
- Weiner, M.L. Parameters and pitfalls to consider in the conduct of food additive research, Carrageenan as a case study. Food Chem. Toxicol. 2016, 87, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Safety Evaluation of Certain Food Additives. In WHO Food Additives Series: 70: Prepared by the Seventy-Ninth Meeting of the Joint FAO/WHO Expert Committee on Food Additives (JECFA); World Health Organization: Geneva, Switzerland, 2015; Available online: http://apps.who.int/iris/bitstream/10665/171781/3/9789240693982_eng.pdf (accessed on 22 June 2019).
- Irhimeh, M.R.; Fitton, J.H.; Lowenthal, R.M.; Kongtawelert, P. A quantitative method to detect fucoidan in human plasma using a novel antibody. Methods Find Exp. Clin. Pharmacol. 2005, 27, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Tokita, Y.; Nakajima, K.; Mochida, H.; Iha, M.; Nagamine, T. Development of a fucoidan-specific antibody and measurement of fucoidan in serum and urine by sandwich ELISA. Biosci. Biotechnol. Biochem. 2010, 74, 350–357. [Google Scholar] [CrossRef] [PubMed]
- Fitton, J.H.; Stringer, D.N.; Karpiniec, S.S. Therapies from Fucoidan: An Update. Mar. Drugs 2015, 13, 5920–5946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nagamine, T.; Nakazato, K.; Tomioka, S.; Iha, M.; Nakajima, K. Intestinal absorption of fucoidan extracted from the brown seaweed Cladosiphon okamuranus. Mar. Drugs 2015, 13, 48–64. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, A.C.; Bourbon, A.I.; Cerqueira, M.A.; Maricato, E.; Nunes, C.; Coimbra, M.A.; Vicente, A.A. Chitosan/fucoidan multilayer nanocapsules as a vehicle for controlled release of bioactive compounds. Carbohydr. Polym. 2015, 115, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Synytsya, A.; Bleha, R.; Pohl, R.; Hayashi, K.; Yoshinaga, K.; Nakano, T.; Hayashi, T. Mekabu fucoidan: structural complexity and defensive effects against avian influenza A viruses. Carbohydr. Polym. 2014, 111, 633–644. [Google Scholar] [CrossRef]
- Hayashi, K.; Lee, J.B.; Nakano, T.; Hayashi, T. Anti-influenza A virus characteristics of a fucoidan from sporophyll of Undaria pinnatifida in mice with normal and compromised immunity. Microbes Infect. 2013, 15, 302–309. [Google Scholar] [CrossRef]
- Makarenkova, I.D.; Deryabin, P.G.; Lvov, D.K.; Zvyagintseva, T.N.; Besednova, N.N. Antiviral activity of sulfated polysaccharide from the brown algae Laminaria japonica against avian influenza A (H5N1) virus infection in the cultured cells. Probl. Virol. 2010, 1, 41–45. (In Russian) [Google Scholar]
- Jiao, G.; Yu, G.; Wang, W.; Zhao, X.; Zhang, J.; Ewart, S. Properties of polysaccharides in several seaweeds from Atlantic Canada and their potential anti-influenza viral activities. J. Ocean Univ. China 2012, 11, 205–212. [Google Scholar] [CrossRef] [Green Version]
- Song, L.; Chen, X.; Liu, X.; Zhang, F.; Hu, L.; Yue, Y.; Li, K.; Li, P. Characterization and comparison of the structural features, immunomodulatory and anti-avian influenza virus activities conferred by three algal sulfated polysaccharides. Mar. Drugs 2016, 14, 4. [Google Scholar] [CrossRef]
- Jin, Y.; Zhang, Y.; Wan, C.; Wang, H.; Hou, L.; Chang, J.; Fan, K.; Xie, X. Immunomodulatory Activity and Protective Effects of Polysaccharide from Eupatorium adenophorum Leaf Extract on Highly Pathogenic H5N1 Influenza Infection. Evid Based Complement. Alternat. Med. 2013, 2013, 194976. [Google Scholar] [CrossRef] [PubMed]
- Bouhlal, R.; Haslin, C.; Chermann, J.C.; Colliec-Jouault, S.; Sinquin, C.; Simon, G.; Cerantola, S.; Riadi, H.; Bourgougnon, N. Antiviral activities of sulfated polysaccharides isolated from Sphaerococcus coronopifolius (Rhodophytha, Gigartinales) and Boergeseniella thuyoides (Rhodophyta, Ceramiales). Mar. Drugs 2011, 9, 1187–1209. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L. Biological and therapeutic properties of the seaweed polysaccharides. Int. Biol. Rev. 2018, 2. [Google Scholar] [CrossRef] [Green Version]
- Bobrovnitsky, I.P.; Mikhailov, V.I.; Odinets, A.G.; Neretina, T.V.; Dobrynina, T.V.; Klinov, D.V. Study of the structure of fucoidan (isolated from Laminaria japonica) and the mechanism of its antiviral activity by atomic force microscopy. New Med. Technol. 2010, 2, 24–28. (In Russian) [Google Scholar]
- Zaporozhets, T.S.; Besednova, N.N. The effect of sulfated algae polysaccharides on factors of adaptive immunity and cytokine production. In Fucoidans - Sulfated Polysaccharides of Brown Algae. Structure and Biological Properties; Dal–nauka FED RAS: Vladivostok, Russia, 2014; pp. 217–229. [Google Scholar]
- Makarenkova, I.D.; Zaporozhets, T.S.; Besednova, N.N. Sulfated polysaccharides of brown algae are agonists of the functions of innate immunity. In Fucoidans - Sulfated Polysaccharides of Brown Algae. Structure and Biological Properties; Dal’nauka FED RAS: Vladivostok, Russia, 2014; pp. 178–217. [Google Scholar]
- Antonyuk, V.A. The role of lectins as biologically active substances in pharmaceutical preparations. Pharmacokinet. Pharmacodyn. 2014, 1, 14–20. (In Russian) [Google Scholar]
- Stonik, V.A. Biomolecules. 2018. Available online: http://www.piboc.dvo.ru/tmp/contents_Biomolecules.pdf (accessed on 22 June 2019).
- Kamiya, H. Possible multiple functions of the invertebrate humoral lectins. Fish Pathol. 1995, 30, 129–139. [Google Scholar] [CrossRef]
- Ferir, G.; Huskens, D.; Noppen, S.; Koharudin, L.M.; Gronenborn, A.M.; Schols, D. Broad anti-HIV activity of the Oscillatoria agardhii agglutinin homologue lectin family. J. Antimicrob. Chemother. 2014, 69, 2746–2758. [Google Scholar] [CrossRef] [Green Version]
- Koharudin, L.M.; Furey, W.; Gronenborn, A.M. Novel fold and carbohydrate specificity of the potent anti-HIV cyanobacterial lectin from Oscillatoria agardhii. J. Biol. Chem. 2011, 286, 1588–1597. [Google Scholar] [CrossRef]
- Koharudin, L.M.; Kollipara, S.; Aiken, C.; Gronenborn, A.M. Structural insights into the anti-HIV activity of the Oscillatoria agardhii agglutinin homolog lectin family. J. Biol. Chem. 2012, 287, 33796–33811. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Okuyama, S.; Hori, K. Primary structure and carbohydrate binding specificity of a potent anti-HIV lectin isolated from the filamentous cyanobacterium Oscillatoria agardhii. J. Biol. Chem. 2007, 282, 11021–11029. [Google Scholar] [CrossRef] [PubMed]
- Hori, K.; Miyazawa, K.; Ito, K. Some common properties of lectins from marine algae. Hydrobiologia 1990, 204, 561–566. [Google Scholar] [CrossRef]
- Mu, J.; Hirajama, M.; Sato, Y.; Morimoto, K.; Hori, K. A novel high-mannose specific lectin from the green alga Halimeda renshii exhibits potent anti-influenza virus activity through high-affinity binding to the viral hemagglutinin. Mar. Drugs 2017, 15, E255. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Morimoto, K.; Hirayama, M.; Hori, K. High mannose-specific lectin (KAA-2) from the red alga Kappaphycus alvarezii potently inhibits influenza virus infection in a strain-independent manner. Biochem. Biophys. Res. Commun. 2011, 405, 291–296. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y. Structure and Function of a Novel Class of High Mannose-binding Proteins with Anti-viral or Anti-tumor Activity. Yakugaku zasshi: J. Pharmac. Soc. Jpn. 2015, 135, 1281–1289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koivicco, R.; Loponen, J.; Pihlaja, K.; Jormalainen, V. High-perfomance liquid chromatographic analysis of florotannins from the brown alga Fucus vesiculosus. Phytochem. Anal. 2007, 18, 326–332. [Google Scholar] [CrossRef]
- Bogolitsyn, K.G.; Druzhinina, A.S.; Ovchinnikov, D.V.; Kaplitin, P.A.; Shulgina, E.V.; Parshina, A.E. Polyphenols of brown algae. Chem. Veget. Raw Mater. 2018, 3, 5–21. (In Russian) [Google Scholar] [CrossRef]
- Ryu, Y.B.; Jeong, H.J.; Yoon, S.Y.; Park, J.Y.; Kim, Y.M.; Park, S.J.; Rho, M.C.; Kim, S.J.; Lee, W.S. Influenza virus neuraminidase inhibitory activity of phlorotannins from the edible brown alga Ecklonia cava. J. Agric. Food Chem. 2011, 59, 6467–6473. [Google Scholar] [CrossRef]
- Tsybalova, L.M.; Kiselev, O.I. Universal flu vaccines. Developments, use prospects. Voprosy Virol. 2012, 1, 9–14. [Google Scholar]
- Zhang, W.; Oda, T.; Yu, Q.; Jin, J.O. Fucoidan from Macrocystis pyrifera has powerful immune-modulatory effects compared to three other fucoidans. Mar. Drugs 2015, 13, 1084–1104. [Google Scholar] [CrossRef] [PubMed]
- Semakova, A.P.; Mikshis, N.I. Adjuvant technologies in the creation of modern vaccines. Probl. Espec. Danger. Infect. 2016, 2, 28–35. (In Russian) [Google Scholar]
- Petrovsky, N.; Cooper, P.D. Carbohydrate-based immune adjuvants. Expert Rev. Vaccines 2011, 10, 523–537. [Google Scholar] [CrossRef] [PubMed]
- Makarenkova, I.D.; Logunov, D.Y.; Tukhvatulin, A.I.; Semenov, I.B.; Zvyagintseva, T.N.; Gorbach, V.I.; Ermakova, S.P.; Besednova, N.N. Sulfated polysaccharides of brown seaweeds are ligands of Toll-like receptors. Biochem. (Moscow). Suppl. Ser. B Biomed. Chem. 2012, 6, 75–80. [Google Scholar] [CrossRef]
- Lin, C.C.; Pan, I.H.; Li, Y.R.; Pan, Y.G.; Lin, M.K.; Lu, Y.H.; Wu, H.C.; Chu, C.L. The adjuvant effects of high-molecule-weight polysaccharides purified from Antrodia cinnamomea on dendritic cell function and DNA vaccines. PLoS ONE 2015, 10, e0116191. [Google Scholar] [CrossRef] [PubMed]
- Kuznetsova, T.A.; Ivanushko, L.A.; Persiyanova, E.V.; Shutikova, A.L.; Ermakova, S.P.; Khotimchenko, M.Y.; Besednova, N.N. Evaluation of adjuvant effects of fucoidan from brown seaweed Fucus evanescens and its structural analogues for the strengthening vaccines effectiveness. Biomeditsinskaya Khimiya 2017, 63, 553–558. (In Russian) [Google Scholar] [CrossRef] [PubMed]
- Negishi, H.; Mori, M.; Mori, H.; Yamori, Y. Supplementation of elderly Japanese men and women with fucoidan from seaweed increases immune responses to seasonal influenza vaccination. J. Nutr. 2013, 143, 1794–1798. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Direct Acting Antiviral Drugs | Mechanism of Action of Antiviral Drugs | Side Effects |
|---|---|---|
| Oseltamivir (Tamiflu) | Competitive and selective inhibitor of neuraminidase of influenza viruses A and B | The gastrointestinal tract can be involved (nausea and vomiting; diarrhea, abdominal bloating, and fecal incontinence); central nervous system (dizziness, migraine, sleep disturbance, weakness); respiratory tract system (bronchitis, cough, infections of the upper respiratory tract); generalized pain |
| Zanamivir (Relenza) | Selective inhibitor of neuraminidase of influenza viruses A, inhibitor of replication and release of new virus particles | Allergies, breathing problems, dermatological disorders |
| Umifenovir (Arbidol) | A specific inhibitor of the fusion of a viral lipid membrane with cell membranes. Interacts with NA, prevents its conformation, which is necessary for the fusion of NA with endosome membranes. Interferon inductor. Stimulator of humoral and cellular immunity | Pruritus, rash, angioedema, urticaria, anaphylaxis |
| Riamylovir (Triazavirin) | Inhibitor of viral RNA synthesis and replication of genomic fragments | Allergies; the gastrointestinal tract can be involved (nausea and vomiting; diarrhea, abdominal bloating, and fecal incontinence) |
| Rimantadine, amantadine | M2 channel blockers. An inhibitor of the early stage of virus reproduction from the moment it enters the cell until the beginning of the transcription process. RNA inhibitor | The gastrointestinal tract can be involved (diarrhea); central nervous system (dizziness, migraine, sleep disturbance, weakness); respiratory tract system (cough); generalized pain; allergies |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Besednova, N.; Zaporozhets, T.; Kuznetsova, T.; Makarenkova, I.; Fedyanina, L.; Kryzhanovsky, S.; Malyarenko, O.; Ermakova, S. Metabolites of Seaweeds as Potential Agents for the Prevention and Therapy of Influenza Infection. Mar. Drugs 2019, 17, 373. https://doi.org/10.3390/md17060373
Besednova N, Zaporozhets T, Kuznetsova T, Makarenkova I, Fedyanina L, Kryzhanovsky S, Malyarenko O, Ermakova S. Metabolites of Seaweeds as Potential Agents for the Prevention and Therapy of Influenza Infection. Marine Drugs. 2019; 17(6):373. https://doi.org/10.3390/md17060373
Chicago/Turabian StyleBesednova, Natalia, Tatiana Zaporozhets, Tatiana Kuznetsova, Ilona Makarenkova, Lydmila Fedyanina, Sergey Kryzhanovsky, Olesya Malyarenko, and Svetlana Ermakova. 2019. "Metabolites of Seaweeds as Potential Agents for the Prevention and Therapy of Influenza Infection" Marine Drugs 17, no. 6: 373. https://doi.org/10.3390/md17060373
APA StyleBesednova, N., Zaporozhets, T., Kuznetsova, T., Makarenkova, I., Fedyanina, L., Kryzhanovsky, S., Malyarenko, O., & Ermakova, S. (2019). Metabolites of Seaweeds as Potential Agents for the Prevention and Therapy of Influenza Infection. Marine Drugs, 17(6), 373. https://doi.org/10.3390/md17060373