First Insight on the Mucus of the Annelid Myxicola infundibulum (Polychaeta, Sabellidae) as a Potential Prospect for Drug Discovery





Abstract
:1. Introduction
2. Results
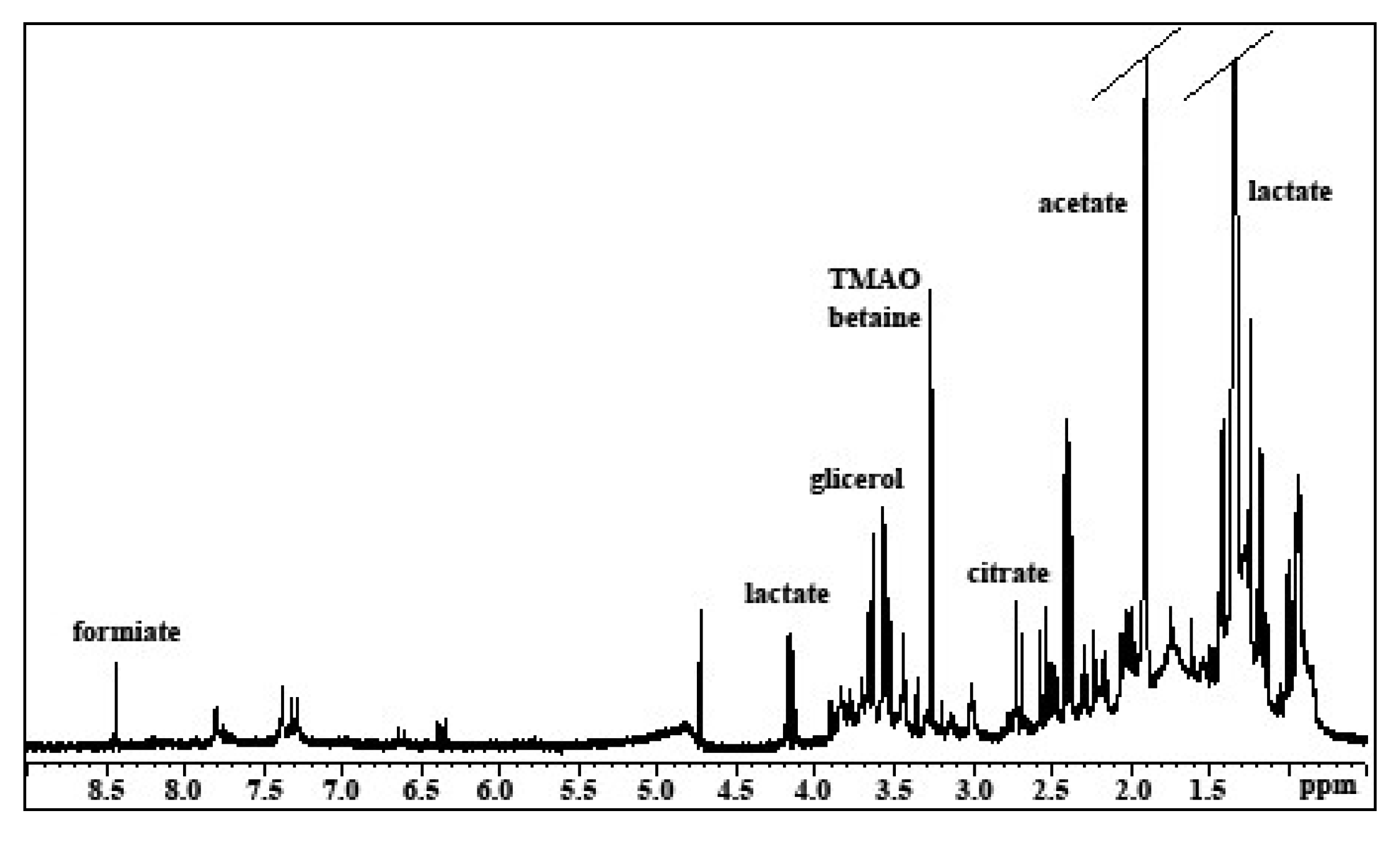
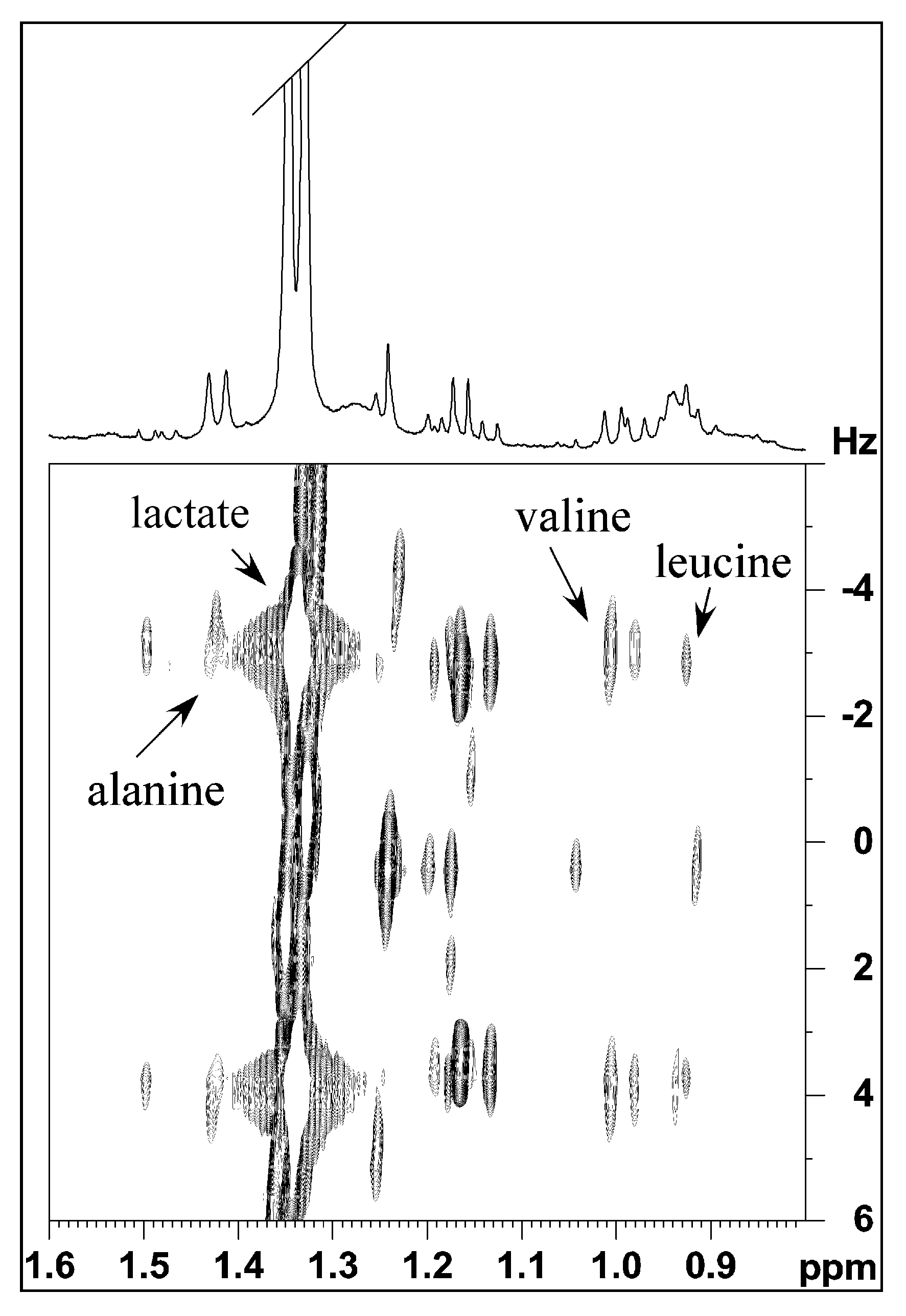
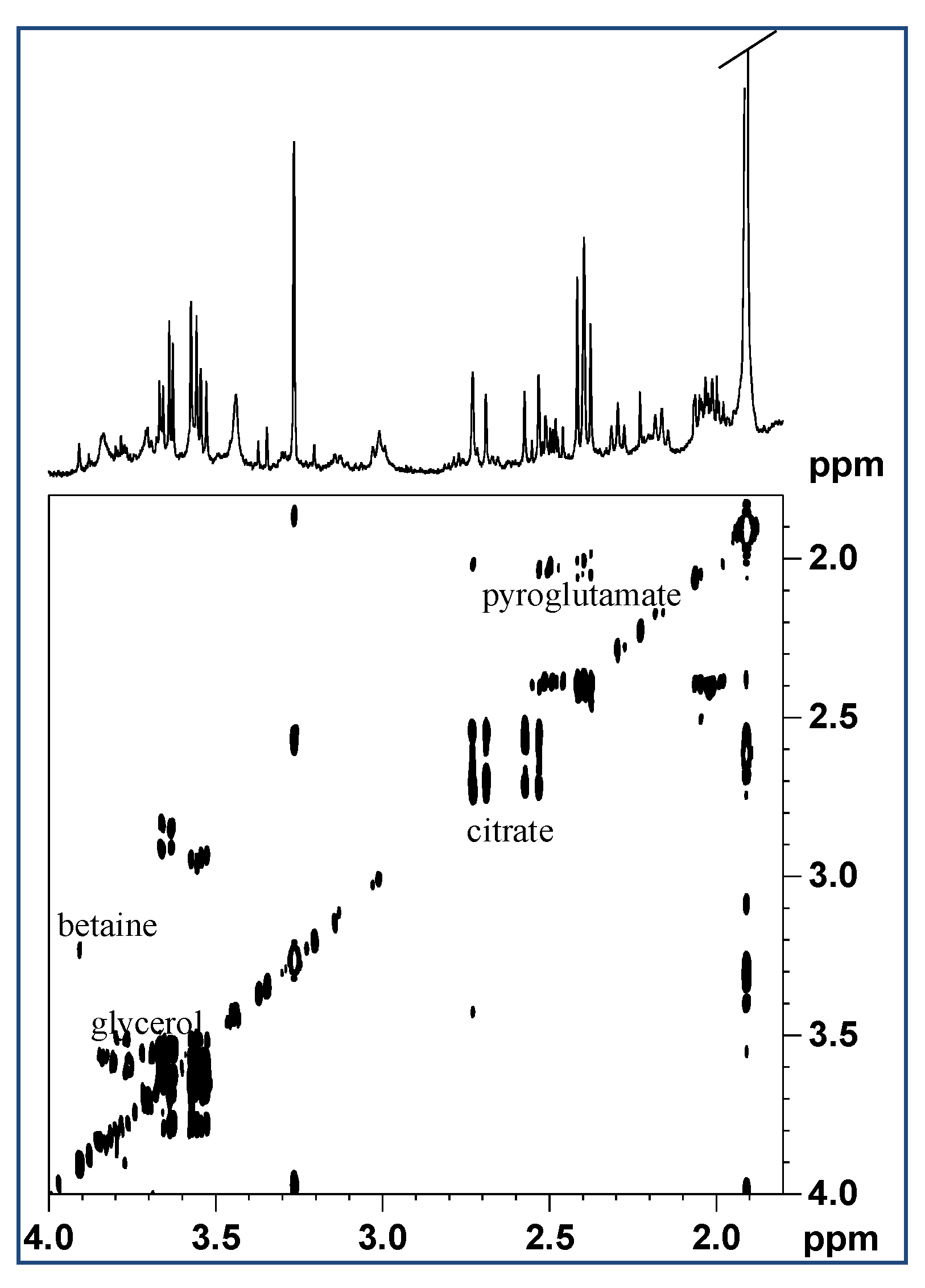
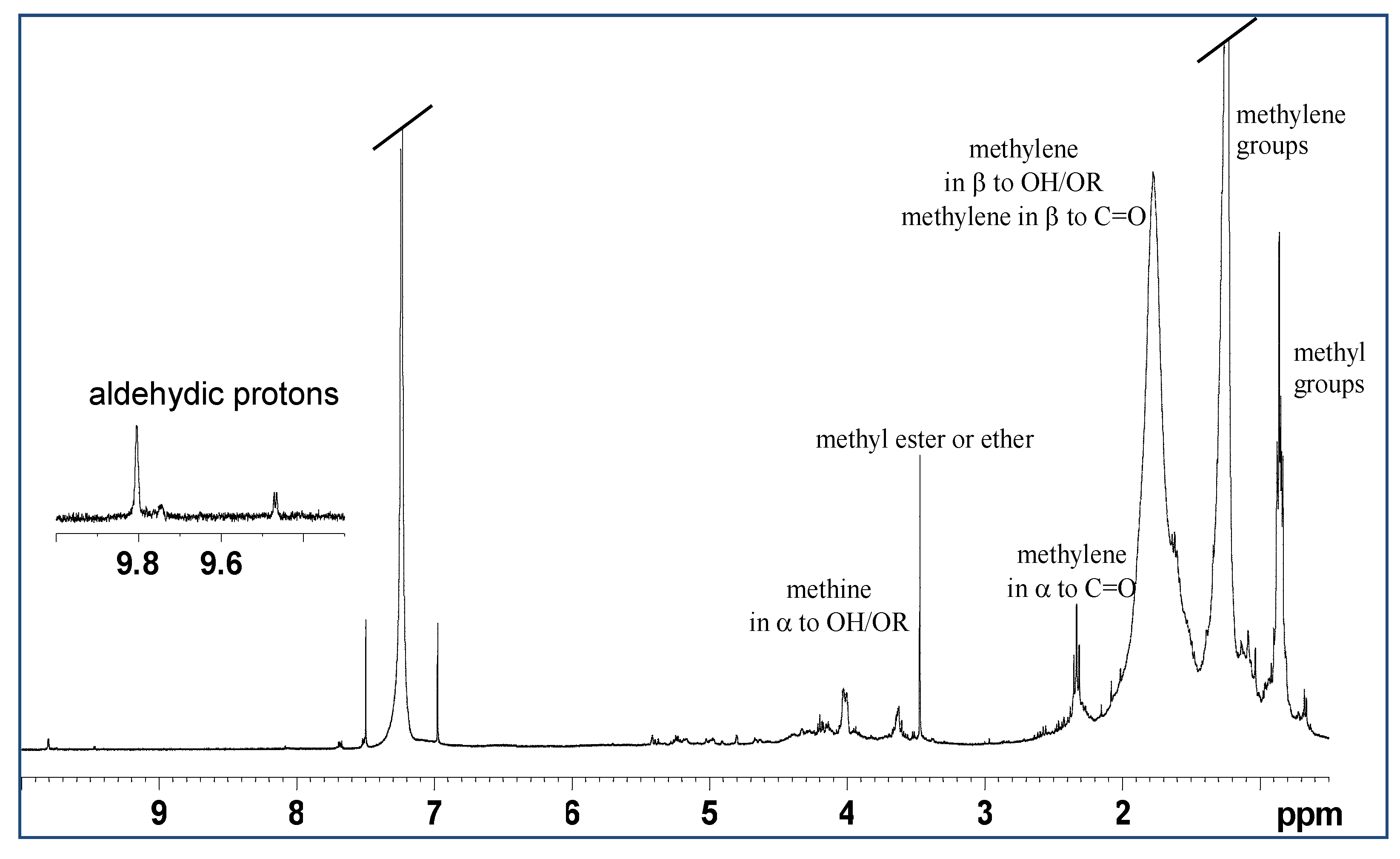
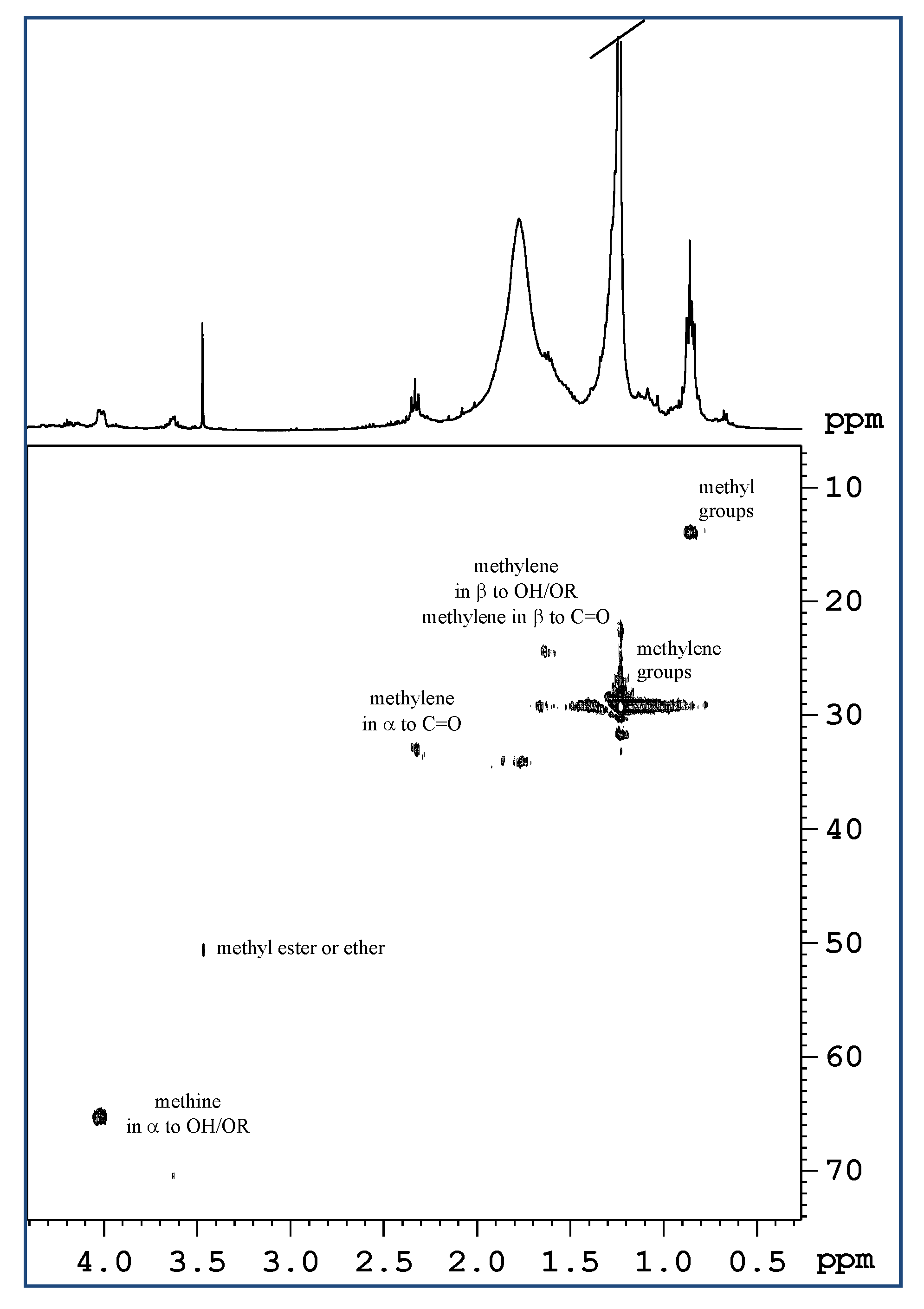
2.1. Analysis of Mucus Composition by NMR Spectroscopy
2.1.1. Aqueous Extract
2.1.2. Lipidic Extract
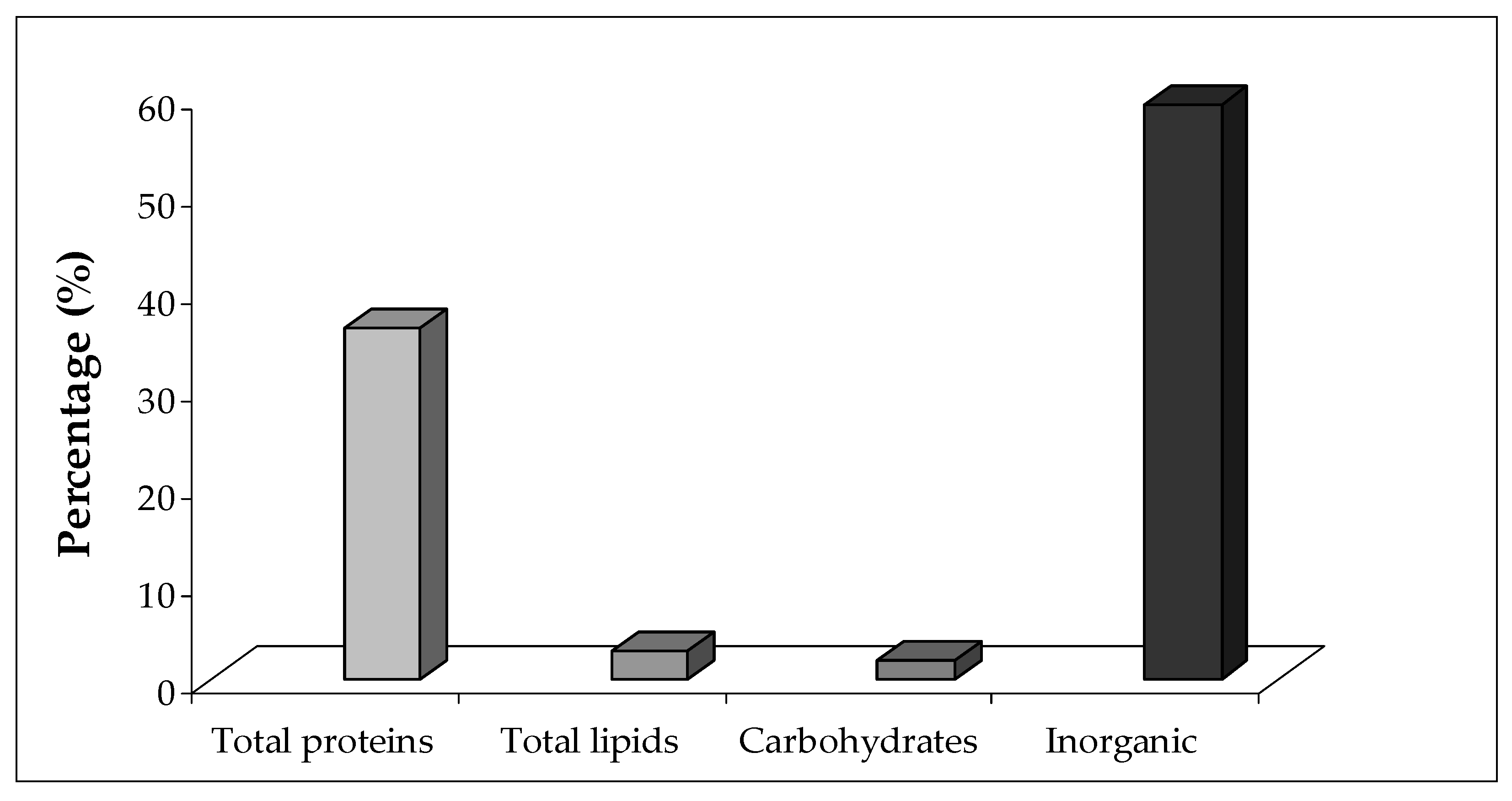
2.2. Water and Inorganic Content
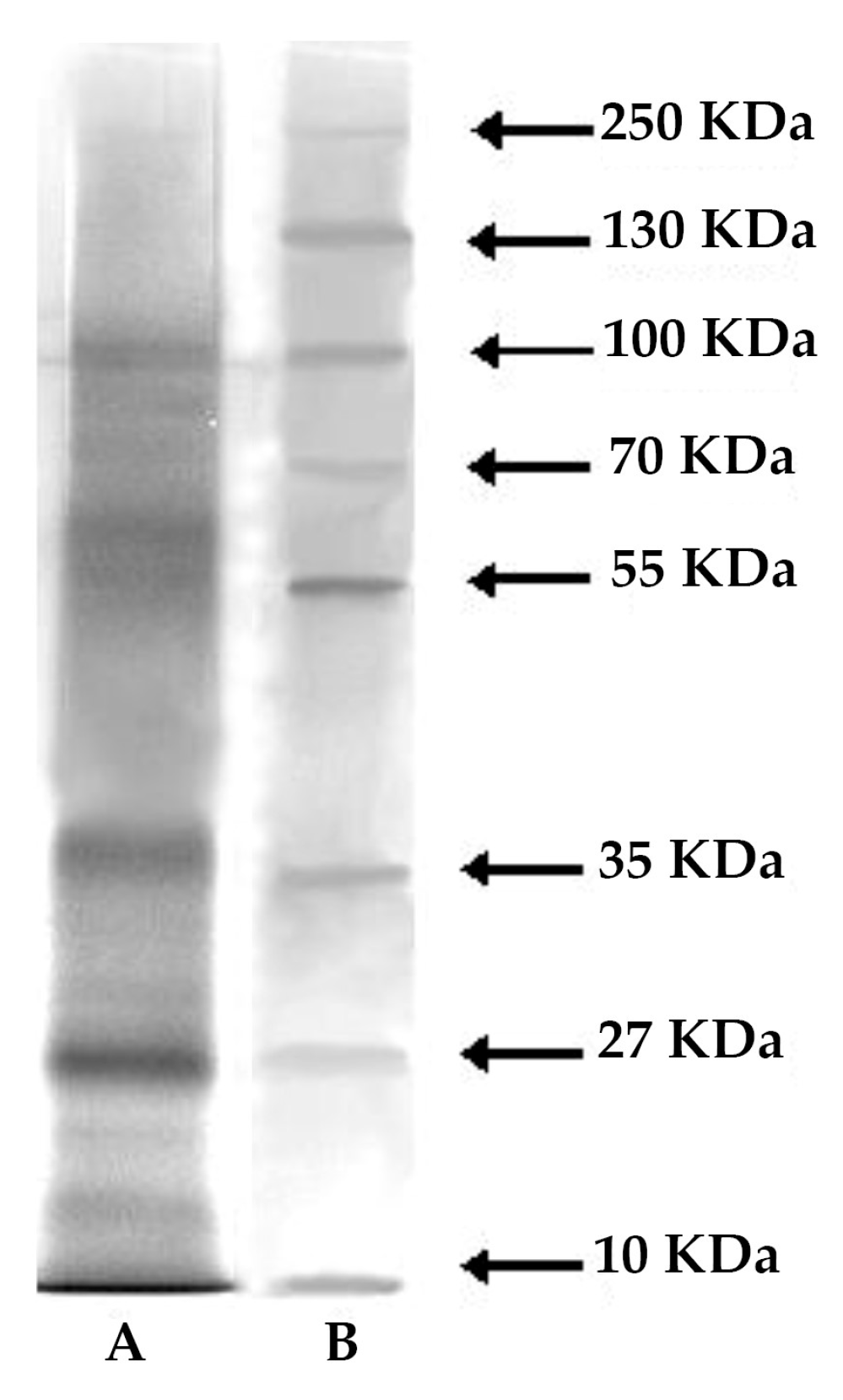
2.3. Protein, Lipid, and Carbohydrate Content
2.4. Antibacterial Activity
2.5. Antioxidant Activity
3. Discussion
4. Materials and Methods
4.1. Animals and Sample Preparation
4.2. NMR Spectroscopy
4.3. Water, Lipid, Protein, and Carbohydrate Content
4.4. Electrophoresis
4.5. Antibacterial Activity
4.6. Antioxidant Activity
4.6.1. Oxygen Radical Absorbance Capacity Assay (ORAC)
4.6.2. Trolox Equivalent Antioxidant Capacity Assay (TEAC)
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bansil, R.; Turner, B.S. Mucin structure, aggregation, physiological functions and biomedical applications. J. Colloid Interface Sci. 2006, 11, 164–170. [Google Scholar] [CrossRef]
- Martin, R.; Walther, P. Protective mechanisms against the action of nematocysts in the epidermis of Cratena peregrina and Flabellina affinis (Gastropoda, Nudibranchia). Zoomorphology 2003, 122, 25–35. [Google Scholar] [CrossRef]
- Derby, C.D. Escape by inking and secreting: Marine molluscs avoid predators through a rich array of chemicals and mechanisms. Biol. Bull. 2007, 213, 274–289. [Google Scholar] [CrossRef] [PubMed]
- Calow, P. Why some metazoan mucus secretions are more susceptible to microbial attack than others. Am. Nat. 1979, 114, 149–152. [Google Scholar] [CrossRef]
- Azam, F. Microbial control of oceanic carbon flux: The plot thickens. Science 1998, 280, 694–696. [Google Scholar] [CrossRef]
- Conley, K.R.; Lombard, F.; Sutherland, K.R. Mammoth grazers on the ocean’s minuteness: A review of selective feeding using mucous meshes. Proc. Biol. Soc. B 2018, 285, 20180056. [Google Scholar] [CrossRef] [PubMed]
- Bonar, D.B. Feeding and tube construction in Chone mollis Bush (Polychaeta, Sabellidae). J. Exp. Mar. Biol. Ecol. 1972, 9, 1–18. [Google Scholar] [CrossRef]
- Giangrande, A.; Licciano, M.; Pagliara, P.; Gambi, M.C. Gametogenesis and larval development in Sabella spallanzanii (Polychaeta: Sabellidae) from the Mediterranean Sea. Mar. Biol. 2000, 136, 847–861. [Google Scholar] [CrossRef]
- Stabili, L.; Schirosi, R.; Licciano, M.; Giangrande, A. The mucus of Sabella spallanzanii (Annelida, Polychaeta): Its involvement in chemical defence and fertilization success. J. Exp. Mar. Biol. Ecol. 2009, 374, 144–149. [Google Scholar] [CrossRef]
- Mastrodonato, M.; Lepore, E.; Gherardi, M.; Zizza, S.; Sciscioli, M.; Ferri, D. Histochemical and ultrastructural analysis of the epidermal gland cells of Branchioma luctuosum (Polichaeta, Sabellidae). Invertebr. Biol. 2005, 124, 303–309. [Google Scholar] [CrossRef]
- Nozais, C.; Duchene, J.C.; Bhaud, M. Control of position in the water column by the larvae of Poecilochaetus serpens, (Polychaeta): The importance of mucus secretion. J. Exp. Mar. Biol. Ecol. 1997, 210, 91–106. [Google Scholar] [CrossRef]
- Dales, R.P. The coelomic and peritoneal cell systems of some sabellid polychaetes. Q. J. Microsc. Sci. 1961, 3, 327–346. [Google Scholar]
- Canicattì, C.; Ville, P.; Pagliara, P.; Roch, P. Hemolysins from the mucus of Spirographis spallanzanii (Polychaeta: Sabellidae). Mar. Biol. 1992, 114, 453–458. [Google Scholar] [CrossRef]
- Iwanaga, S.; Lee, B.L. Recent advances in the innate immunity of invertebrate animals. J. Biochem. Mol. Biol. 2005, 38, 128–150. [Google Scholar] [CrossRef] [PubMed]
- M’Beri, M. Dégranulation des Coelomocytes au Cours des Réactions Immunitaires d’un Invertébré Marin: Nereis diversicolor (Annélides polychète): Caractérisation des Coelomocytes d’après Leurs Récepteurs Membranaires, Identification et Rôle Biologique des Produits Extrudés. Ph.D. Thesis, Universitè de Lille I, Lille, France, 1988. [Google Scholar]
- Perin, J.P.; Jolles, P. The lysozyme from Nephthys hombergi (Annelida). Biochim. Biophys. Acta 1972, 263, 683–689. [Google Scholar] [CrossRef]
- Zapata Vivens, E.; Nusetti, O.A.; Marcano, L.; Esclapes, M.M.; Arredondo, L. Immunological responses and wound healing in the polychaete Eurythoe complanata (Annelida: Amphinomidae) exposed to copper. Cienc. Mar. 2005, 31, 1–10. [Google Scholar] [CrossRef]
- Moraes, T.B.; Ferreira, J.L.; da Rosa, C.E.; Sandrini, J.Z.; Votto, A.P.; Trindade, G.S.; Geracitano, L.A.; Abreu, P.C.; Monserrat, J.M. Antioxidant properties of the mucus secreted by Laeonereis acuta (Polychaeta, Nereididae): A defense against environmental pro-oxidants? Comp. Biochem. Physiol. C 2006, 142, 292–300. [Google Scholar] [CrossRef]
- Papaspyrou, S.; Gregersen, T.; Kristensen, E.; Christensen, B.; Cox, R.P. Microbial reaction rates and bacterial communities in sediment surrounding burrows of two nereidid polychaetes (Nereis diversicolor and N. virens). Mar. Biol. 2006, 148, 541–550. [Google Scholar] [CrossRef]
- Mouneyrac, C.; Mastain, O.; Amiard, J.C.; Amiard-Triquet, C.; Beaunier, P.; Jeantet, A.Y.; Smith, B.D.; Rainbow, P.S. Trace-metal detoxication and tolerance of the estuarine worm Hediste diversicolor chronically exposed in their environment. Mar. Biol. 2003, 143, 731–744. [Google Scholar] [CrossRef]
- Gelain, D.P.; Behr, G.A.; de Oliveira, R.B.; Trujillo, M. Antioxidant therapies for neurodegenerative diseases: mechanisms, current trends, and perspectives. Oxid. Med. Cell Longev. 2012, 2012, 895153. [Google Scholar] [CrossRef]
- Pachiayappan, A.; Muthuvel, A.; Sadhasivam, G. In vitro antioxidant activity of different gastropods, bivalves and echinoderm by solvent extraction method. Int. J. Pharm. Sci. Res. 2014, 5, 2539–2545. [Google Scholar] [CrossRef]
- Shahidi, F. Maximizing the Value of Marine by-Products; Woodhead Publishing Ltd.: Cambridge, UK, 2006; pp. 1–560. ISBN 9781845690137. [Google Scholar]
- Stabili, L.; Giangrande, A.; Pizzolante, G.; Caruso, G.; Alifano, P. Characterization of vibrios diversity in the mucus of the polychaete Myxicola infundibulum (Annelida, Polichaeta). Microb. Ecol. 2014, 67, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.M.; Quick, T.J.; Peter, S.T.R.L. Differences in the composition of adhesive and non-adhesive mucus from the limpet Lottia limatula. Biol. Bull. 1999, 196, 34–44. [Google Scholar] [CrossRef] [PubMed]
- Smith, A.M.; Morin, M.C. Biochemical differences between trail mucus and adhesive mucus from marsh periwinkles. Biol. Bull. 2002, 203, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Bansil, R.; Turner, B.S. The biology of mucus: composition, synthesis and organization. Adv. Drug Deliv. Rev. 2018, 124, 3–15. [Google Scholar] [CrossRef] [PubMed]
- Santos, R.; da Costa, G.; Franco, C.; Gomes-Alves, P.; Flammang, P.; Coelho, A.V. First insights into the biochemistry of tube foot adhesive from the sea urchin Paracentrotus lividus (Echinoidea, Echinodermata). Mar. Biotechnol. 2009, 11, 686–698. [Google Scholar] [CrossRef]
- Stabili, L.; Schirosi, R.; Di Benedetto, A.; Merendino, A.; Villanova, R.; Giangrande, A. First insights into the biochemistry of Sabella spallanzanii (Annelida: Polychaeta) mucus: A potentially unexplored resource for applicative purposes. J. Mar. Biol. Assoc. UK 2011, 91, 199–208. [Google Scholar] [CrossRef]
- Flammang, P.; Santos, R.; Haesaerts, D. Echinoderm adhesive secretions: From experimental characterization to evidence-based complementary and alternative medicine biotechnological applications. In Progress in Molecular and Subcellular Biology, Marine Molecular Biotechnology, Echinodermata; Matranga, V., Ed.; Springer: Berlin, Germany, 2005; pp. 201–220. ISBN 978-3-540-24402-8. [Google Scholar]
- Smith, A.M. The biochemistry and mechanics of gastropod adhesive gels. In Biological Adhesives; Smith, A.M., Callow, J.A., Eds.; Springer: Berlin, Germany, 2006; pp. 167–182. ISBN 978-3-540-31048-8. [Google Scholar]
- Kamino, K. Barnacle underwater attachment. In Biological Adhesives; Smith, A.M., Callow, J.A., Eds.; Springer: Berlin, Germany, 2006; pp. 145–166. ISBN 3-540-31048-7. [Google Scholar]
- Jolles, P.; Jolles, J. What’s new in lysozyme research?—Always a model system, today as yesterday. Mol. Cell Biochem. 1984, 6, 165–189. [Google Scholar] [CrossRef]
- Dhainaut, A.; Scaps, P. Immune defense and biological responses induced by toxics in Annelida. Can. J. Zool. 2001, 79, 233–253. [Google Scholar] [CrossRef]
- Paul, V.J.; Puglisi, M.P. Chemical mediation of interactions among marine organisms. Nat. Prod. Rep. 2004, 21, 189–209. [Google Scholar] [CrossRef]
- Dobretsov, S.; Dahms, H.U.; Tsoi, M.Y.; Qjan, P.Y. Chemical control of epibiosis by Hong Kong sponges: The effect of sponge extracts on micro- and macrofouling communities. Mar. Ecol. Prog. Ser. 2005, 297, 119–129. [Google Scholar] [CrossRef]
- Kelly, S.R.; Garo, E.; Jensen, P.R.; Fenical, W.; Pawlik, J.R. Effects of Caribbean sponge secondary metabolites on bacterial surface colonization. Aquat. Microb. Ecol. 2005, 40, 191–203. [Google Scholar] [CrossRef]
- Zachary, D.; Hoffmann, D. Lysozyme is stored in the granules of certain haemocyte types in Locusta. J. Insect. Physiol. 1984, 30, 405–411. [Google Scholar] [CrossRef]
- Marcano, L.; Nusetti, O.; Rodríguez-Grau, J.; Briceño, J.; Vilas, J. Coelomic fluid lysozyme activity induction in the polychaete Eurythoe complanata as a biomarker of heavy metal toxicity. Bull. Environ. Contamin. Toxicol. 1997, 59, 22–28. [Google Scholar] [CrossRef]
- Parisien, A.; Allain, B.; Zhang, J.; Mandeville, R.; Lan, C.Q. Novel alternatives to antibiotics: Bacteriophages, bacterial cell wall hydrolases, and antimicrobial peptides. J. Appl. Microbiol. 2008, 104, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Boucher, H.W.; Talbot, G.H.; Benjamin, D.K.; Bradley, J.; Guidos, R.J.; Jones, R.N.; Murray, B.E.; Bonomo, R.A.; Gilbert, D. 10 × 20 progress—Development of new drugs active against gram negative bacilli: An update from the infectious diseases society of America. Clin. Infect. Dis. 2013, 56, 1685–1694. [Google Scholar] [CrossRef]
- Ibrahim, H.R.; Aoki, T.; Pellegrini, A. Strategies for new antimicrobial proteins and peptides: Lysozyme and aprotinin as model molecules. Curr. Pharm. Des. 2002, 8, 671–693. [Google Scholar] [CrossRef]
- Niyonsaba, F.; Ogawa, H. Protective roles of the skin against infection: Implication of naturally occurring human antimicrobial agents β-defensins, cathelicidin LL-37 and lysozyme. J. Dermatol. Sci. 2005, 40, 157–168. [Google Scholar] [CrossRef]
- McNeil, D.S.; Nuccio, M.L.; Hanson, A.D. Betaines and related osmoprotectants. Targets for metabolic engineering of stress resistance. Plant. Physiol. 1999, 120, 945–949. [Google Scholar] [CrossRef]
- Oren, A. A hundred years of Dunaliella research: 1905–2005. Saline Syst. 2005, 1, 2. [Google Scholar] [CrossRef]
- Muscatine, L. Glycerol excretion by symbiotic algae from corals and Tridacna and its control by the host. Science 1967, 156, 516–519. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Ou, B.; Prior, R.L. The chemistry behind antioxidant capacity assay. J. Agric. Food Chem. 2005, 53, 1841–1856. [Google Scholar] [CrossRef] [PubMed]
- Zulueta, A.; Esteve, M.J.; Frigola, A. ORAC and TEAC assays comparison to measure the antioxidant capacity of food products. Food Chem. 2009, 114, 310–316. [Google Scholar] [CrossRef]
- Sarmadi, B.H.; Ismail, A. Antioxidative peptides from food proteins: A review. Peptides 2010, 31, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, P.M.; Hégaret, H.; Lambert, C.; Wikfors, G.H.; Le Goïc, N.; Shumway, S.E.; Soudant, P. Immunological responses of the manila clam (Ruditapes philippinarum) with varying parasite (Perkinsus olseni) burden, during a long-term exposure to the harmful alga, Karenia selliformis, and possible interactions. Toxicon 2008, 51, 563–573. [Google Scholar] [CrossRef] [PubMed]
- Livingstone, D.R. Oxidative stress in aquatic Organisms in relation to pollution and aquaculture. Revue Méd. Vét. 2003, 154, 427–430. [Google Scholar]
- Alemán, A.; Giménez, B.; Montero, P.; Gómez-Guillén, M.C. Antioxidant activity of several marine skin gelatins. LWT-Food Sci. Technol. 2011, 44, 407–413. [Google Scholar] [CrossRef]
- Elias, R.J.; Kellerby, S.S.; Decker, E.A. Antioxidant activity of proteins and peptides. Crit. Rev. Food Sci. Nutr. 2008, 48, 430–441. [Google Scholar] [CrossRef]
- Abele-Oeschger, D.; Oeschger, R.; Theede, H. Biochemical adaptations of Nereis diversicolor (Polychaeta) to temporarily increased hydrogen peroxide levels in intertidal sandflats. Mar. Ecol. Prog. Ser. 1994, 106, 101–110. [Google Scholar] [CrossRef]
- Abele-Oeschger, D.; Oeschger, R. Enzymatic antioxidant protection in spawn, larvae and adult worms of Phyllodoce mucosa (Polychaeta). Ophelia 1995, 43, 101–110. [Google Scholar] [CrossRef]
- Geracitano, L.A.; Monserrat, J.M.; Bianchini, A. Physiological and antioxidant enzyme responses to acute and chronic exposure of Laeonereis acuta (Polychaeta, Nereididae) to copper. J. Exp. Mar. Biol. Ecol. 2002, 227, 145–156. [Google Scholar] [CrossRef]
- Hermes-Lima, M. Oxygen in biology and biochemistry: Role of free radicals. In Functional Metabolism: Regulation and Adaptation; Storey, K.B., Ed.; John Wiley & Sons: Hoboken, NJ, USA, 2004; pp. 319–368. ISBN 978-0-471-41090-4. [Google Scholar]
- Da Rosa, C.E.; Souza, M.S.; Yunes, J.S.; Proença, L.A.O.; Nery, L.E.M.; Monserrat, J.M. Cyanobacterial blooms in estuarine ecosystems: Characteristics and effects on Laeonereis acuta (Polychaeta, Nereididae). Mar. Pollut. Bull. 2005, 50, 956–964. [Google Scholar] [CrossRef] [PubMed]
- Da Rosa, C.E.; Iurman, M.G.; Abreu, P.C.; Geracitano, L.A.; Monserrat, J.M. Antioxidant mechanisms of the nereidid Laeonereis acuta (Annelida:Polychaeta) to cope with environmental hydrogen peroxide. Physiol. Biochem. Zool. 2005, 78, 641–649. [Google Scholar] [CrossRef] [PubMed]
- Chai, T.T.; Law, Y.C.; Wong, F.C.; Kim, S.K. Enzyme-assisted discovery of antioxidant peptides from edible marine invertebrates: A review. Mar. Drugs 2017, 15, 42. [Google Scholar] [CrossRef] [PubMed]
- Stabili, L.; Schirosi, R.; Licciano, M.; Giangrande, A. Role of Myxicola infundibulum (Polychaeta, Annelida) mucus: From bacterial control to nutritional home site. J. Exp. Mar. Biol. Ecol. 2014, 461, 344–349. [Google Scholar] [CrossRef]
- Standal, I.B.; Axelson, A.E.; Aursand, M. Differentiation of fish oils according to species by 13C-NMR regiospecific analyses of triacyglycerols. J. Am. Oil Chem. Soc. 2009, 86, 401–407. [Google Scholar] [CrossRef]
- Stabili, L.; Acquaviva, M.I.; Biandolino, F.; Cavallo, R.A.; De Pascali, S.A.; Fanizzi, F.P.; Narracci, M.; Cecere, E.; Petrocelli, A. Biotechnological potential of the seaweed Cladophora rupestris (Chlorophyta, Cladophorales) lipidic extract. New Biotechnol. 2014, 31, 436–444. [Google Scholar] [CrossRef] [PubMed]
- Del Coco, L.; Papadia, P.; De Pascali, S.A.; Bressani, G.; Storelli, C.; Zonno, V.; Fanizzi, F.P. Comparison among different gilthead sea bream (Sparus aurata) farming systems: Activity of intestinal and hepatic enzymes and 13C-NMR analysis of lipids. Nutrients 2009, 1, 291–301. [Google Scholar] [CrossRef]
- Kamal, C.; Sethuraman, M.G. Caulerpin—A bis-indole alkaloid as a green inhibitor for the corrosion of mild steel in 1 M HCl solution from the marine alga Caulerpa racemosa. Ind. Eng. Chem. Res. 2012, 51, 10399–10407. [Google Scholar] [CrossRef]
- Folch, J.; Less, M.; Stone Stanley, G.H. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–508. [Google Scholar]
- Zöllner, N.; Kirsch, K. Determination of the total lipid concentration in serum. Z. Gesamte Exp. Med. 1962, 135, 545–549. [Google Scholar] [CrossRef]
- Bradford, M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of proteins dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Kennedy, J.F.; Pagliuca, G. Chapter 2. Oligosaccharides. In Carbohydrate Analysis A Practical Approach, 2nd ed.; Chapling, M.F., Kennedy, J.F., Eds.; Oxford University Press: New York, NY, USA, 1994; pp. 43–72. ISBN 9780199634491. [Google Scholar]
- Laemmli, V.H. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nat. Lond. 1970, 227, 680–682. [Google Scholar] [CrossRef] [PubMed]
- Hames, B.D. One-dimensional polyacrylamide gel electrophoresis. In Gel Electrophoresis of Proteins: A Practical Approach; Hames, B.D., Rickwood, D., Eds.; Oxford University Press: New York, NY, USA, 1990; pp. 1–147. ISBN 0199636419. [Google Scholar]
- Stabili, L.; Acquaviva, M.I.; Cavallo, R.A.; Gerardi, C.; Narracci, M.; Pagliara, P. Screening of three echinoderm species as new opportunity for drug discovery: Their bioactivities and antimicrobial properties. J. Evid. Based Complement. Altern. Med. 2018, 2018, 7891748. [Google Scholar] [CrossRef] [PubMed]
- Dávalos, A.; Gómez-Cordovés, C.; Bartolomé, B. Commercial dietary antioxidant supplements assayed for their antioxidant activity by different methodologies. J. Agric. Food 2003, 51, 2512–2519. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Element | Content % |
|---|---|
| C | 2.34 ± 0.02 |
| H | 1.79 ± 0.11 |
| N | 0.66 ± 0.02 |
| Fe | absent |
| Ca | 1.08 ± 0.02 |
| Mg | 2.82 ± 0.02 |
| Zn | 0.04 ± 0.005 |
| Cu | absent |
| K | 1.35 ±0.02 |
| Na | 14.87 ± 0.10 |
| Cl | 34.13 ± 0.15 |
| P | absent |
| Sn | absent |
| Se | absent |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stabili, L.; Licciano, M.; Giangrande, A.; Gerardi, C.; De Pascali, S.A.; Fanizzi, F.P. First Insight on the Mucus of the Annelid Myxicola infundibulum (Polychaeta, Sabellidae) as a Potential Prospect for Drug Discovery. Mar. Drugs 2019, 17, 396. https://doi.org/10.3390/md17070396
Stabili L, Licciano M, Giangrande A, Gerardi C, De Pascali SA, Fanizzi FP. First Insight on the Mucus of the Annelid Myxicola infundibulum (Polychaeta, Sabellidae) as a Potential Prospect for Drug Discovery. Marine Drugs. 2019; 17(7):396. https://doi.org/10.3390/md17070396
Chicago/Turabian StyleStabili, Loredana, Margherita Licciano, Adriana Giangrande, Carmela Gerardi, Sandra Angelica De Pascali, and Francesco Paolo Fanizzi. 2019. "First Insight on the Mucus of the Annelid Myxicola infundibulum (Polychaeta, Sabellidae) as a Potential Prospect for Drug Discovery" Marine Drugs 17, no. 7: 396. https://doi.org/10.3390/md17070396
APA StyleStabili, L., Licciano, M., Giangrande, A., Gerardi, C., De Pascali, S. A., & Fanizzi, F. P. (2019). First Insight on the Mucus of the Annelid Myxicola infundibulum (Polychaeta, Sabellidae) as a Potential Prospect for Drug Discovery. Marine Drugs, 17(7), 396. https://doi.org/10.3390/md17070396