Discovery of Two New Sorbicillinoids by Overexpression of the Global Regulator LaeA in a Marine-Derived Fungus Penicillium dipodomyis YJ-11

 , , , and
, , , and 
Abstract
:1. Introduction
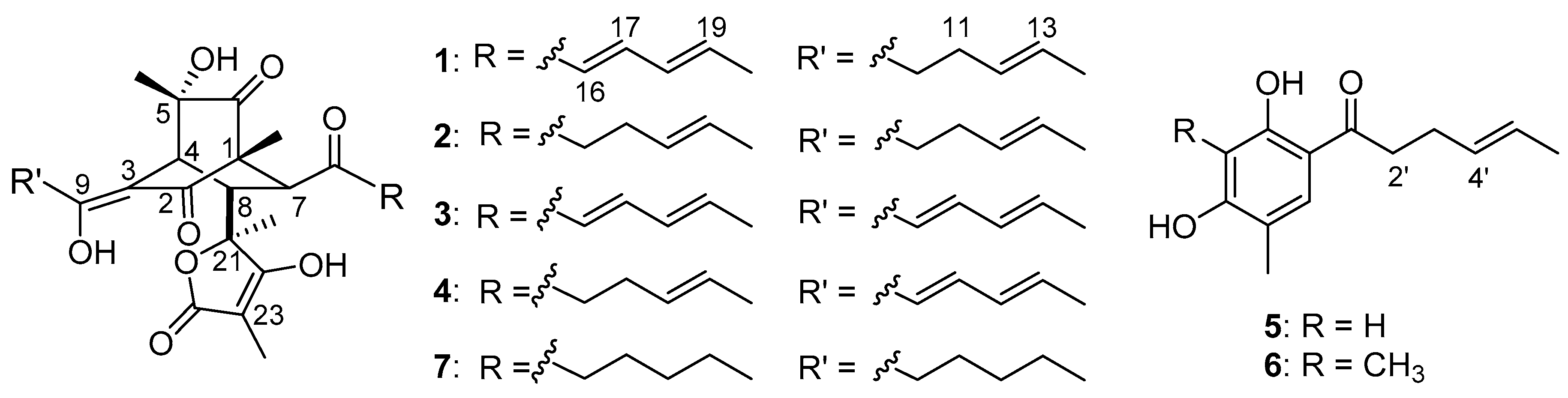
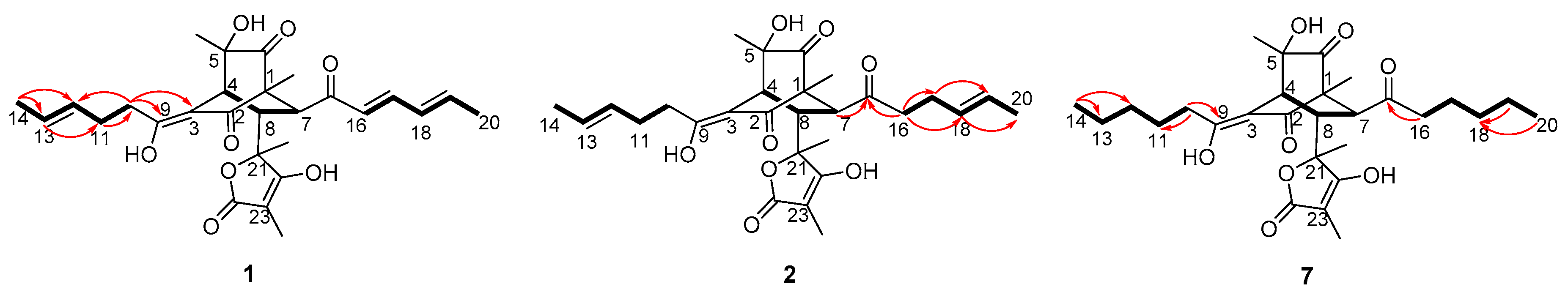
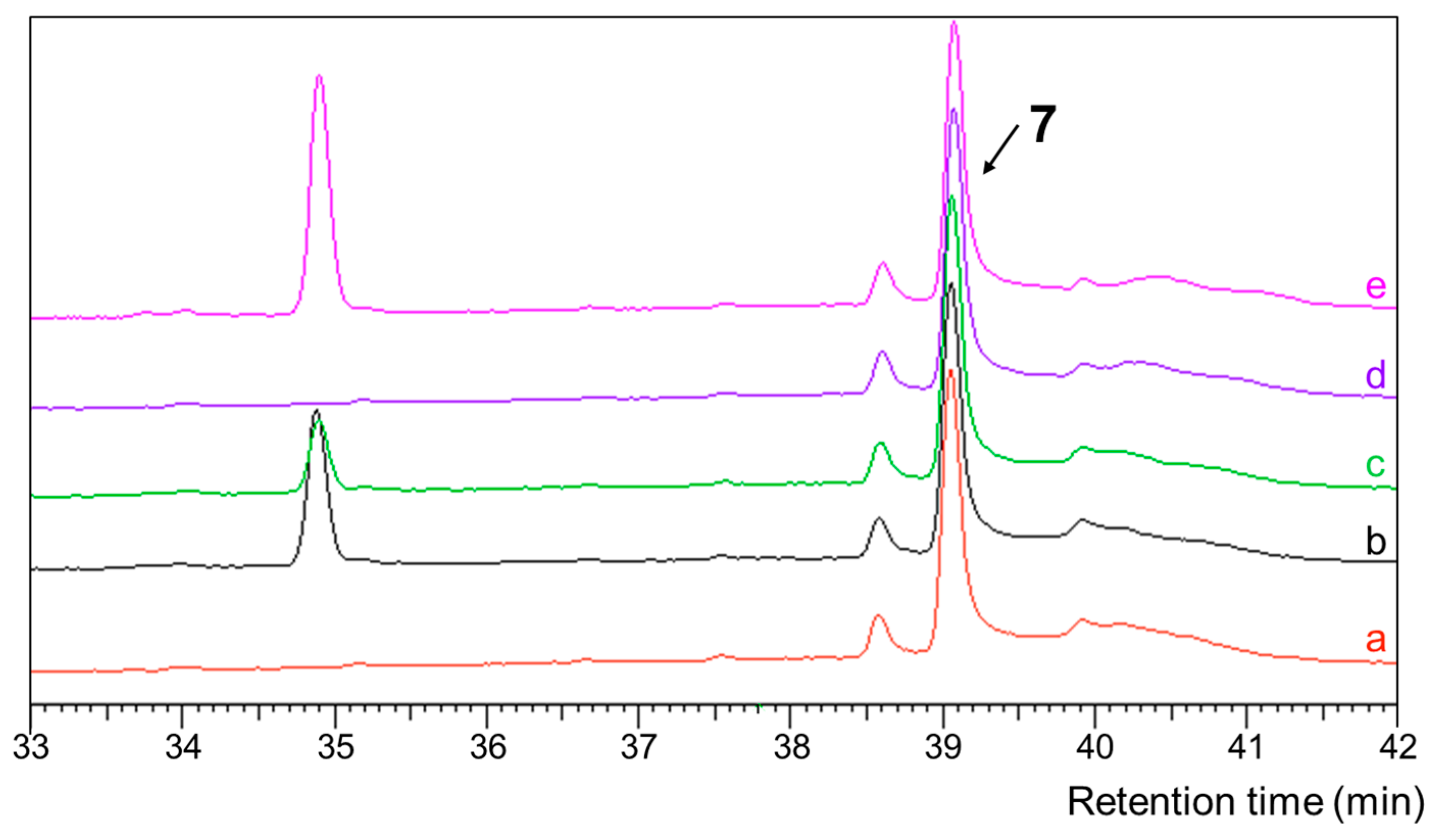
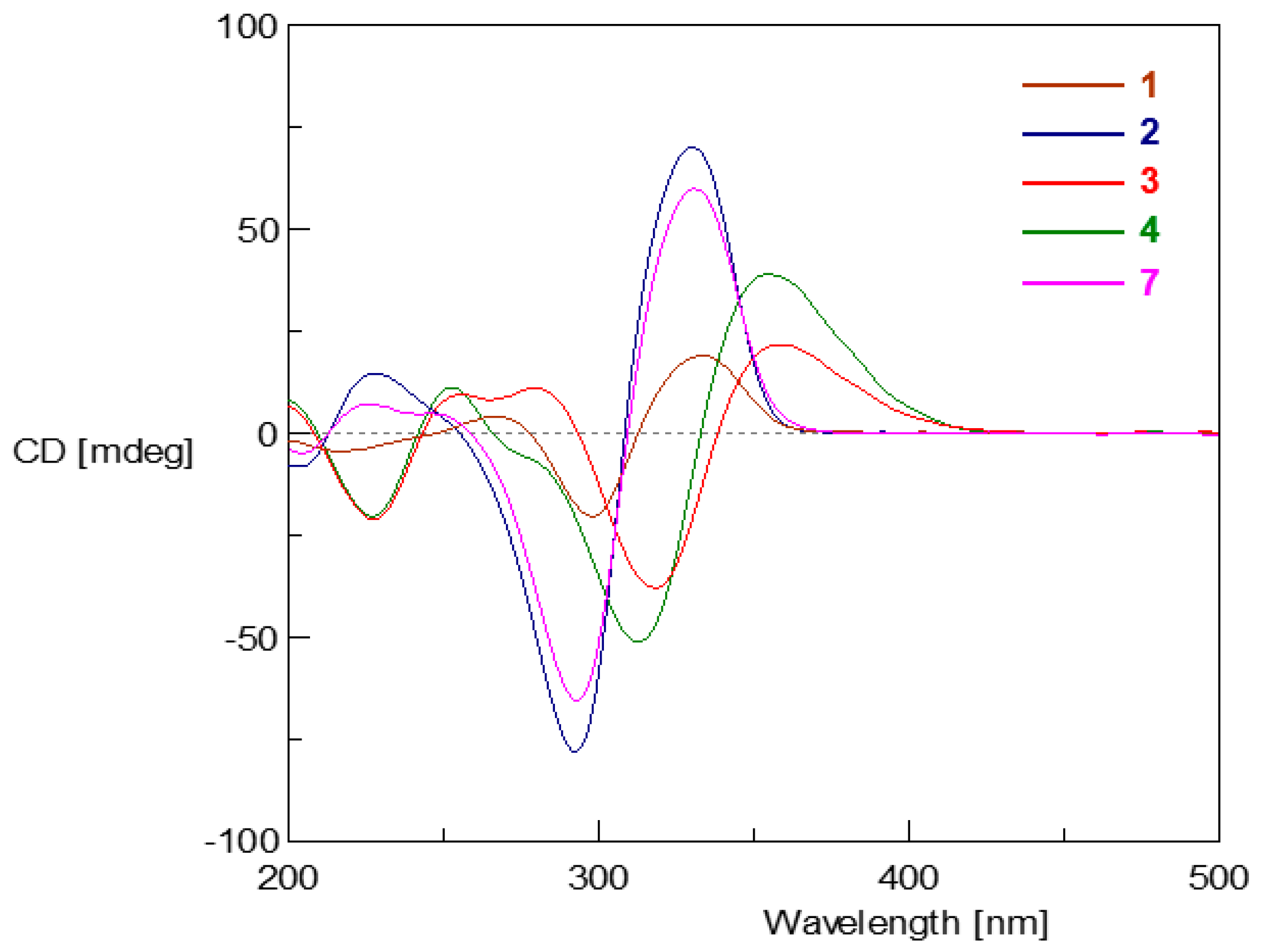
2. Results and Discussion
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Materials and Culture Conditions
3.3. Sequence Analysis of the PdLaeA Gene
3.4. Construction of the PdLaeA Expression Vector
3.5. Fungal Protoplast Formation and Transformation
3.6. Transformants Screening
3.7. Fermentation and Extraction
3.8. Compound Isolation
3.9. Hydrogenation of Compounds 1–4
3.10. Assay of Cytotoxicity, Antimicrobial and Antioxidation Activity
3.11. Assay of Siderophore Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brakhage, A.A.; Schuemann, J.; Bergmann, S.; Scherlach, K.; Schroeckh, V.; Hertweck, C. Activation of fungal silent gene clusters: A new avenue to drug discovery. Prog. Drug Res. 2008, 66, 3–12. [Google Scholar] [CrossRef]
- Netzker, T.; Fischer, J.; Weber, J.; Mattern, D.J.; König, C.C.; Valiante, V.; Schroeckh, V.; Brakhage, A.A. Microbial communication leading to the activation of silent fungal secondary metabolite gene clusters. Front. Microbiol. 2015, 6, 299. [Google Scholar] [CrossRef] [PubMed]
- Rutledge, P.J.; Challis, G.L. Discovery of microbial natural products by activation of silent biosynthetic gene clusters. Nat. Rev. Microbiol. 2015, 13, 509–523. [Google Scholar] [CrossRef] [PubMed]
- Bok, J.W.; Keller, N.P. LaeA, a Regulator of Secondary Metabolism in Aspergillus spp. Eukaryot. Cell 2004, 3, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.S.; Lee, J.H.; Lee, I. Strain improvement by overexpression of the laeA gene in Monascus pilosus for the production of monascus-fermented rice. J. Microbiol. Biotechnol. 2013, 23, 959–965. [Google Scholar] [CrossRef] [PubMed]
- Hong, E.J.; Kim, N.K.; Lee, D.; Kim, W.G.; Lee, I. Overexpression of the laeA gene leads to increased production of cyclopiazonic acid in Aspergillus fumisynnematus. Fungal Biol. 2015, 119, 973–983. [Google Scholar] [CrossRef] [PubMed]
- Linde, T.; Zoglowek, M.; Lubeck, M.; Frisvad, J.C.; Lubeck, P.S. The global regulator LaeA controls production of citric acid and endoglucanases in Aspergillus carbonarius. J. Ind. Microbiol. Biot. 2016, 43, 1139–1147. [Google Scholar] [CrossRef] [PubMed]
- Chettri, P.; Bradshaw, R.E. LaeA negatively regulates dothistromin production in the pine needle pathogen Dothistroma septosporum. Fungal Genet. Biol. 2016, 97, 24–32. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.; Wang, M.; Li, L.; Si, J.; Song, B.; Zhou, C.; Yu, M.; Wang, X.; Zhang, Y.; Ding, G.; et al. Overexpression of the Global Regulator LaeA in Chaetomium globosum Leads to the Biosynthesis of Chaetoglobosin, Z.J. Nat. Prod. 2016, 79, 2487–2494. [Google Scholar] [CrossRef]
- Kosalkova, K.; Garcia-Estrada, C.; Ullan, R.V.; Godio, R.P.; Feltrer, R.; Teijeira, F.; Mauriz, E.; Martin, J.F. The global regulator LaeA controls penicillin biosynthesis, pigmentation and sporulation, but not roquefortine C synthesis in Penicillium chrysogenum. Biochimie 2009, 91, 214–225. [Google Scholar] [CrossRef]
- Liu, Q.; Cai, L.; Shao, Y.; Zhou, Y.; Li, M.; Wang, X.; Chen, F. Inactivation of the global regulator LaeA in Monascus ruber results in a species-dependent response in sporulation and secondary metabolism. Fungal Biol. 2016, 120, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Estiarte, N.; Lawrence, C.B.; Sanchis, V.; Ramos, A.J.; Crespo-Sempere, A. LaeA and VeA are involved in growth morphology, asexual development, and mycotoxin production in Alternaria alternata. Int. J. Food Microbiol. 2016, 238, 153–164. [Google Scholar] [CrossRef] [PubMed]
- Keller, N.P.; Turner, G.; Bennett, J.W. Fungal secondary metabolism—From biochemistry to genomics. Nat. Rev. Microbiol. 2005, 3, 937–947. [Google Scholar] [CrossRef] [PubMed]
- Bok, J.W.; Noordermeer, D.; Kale, S.P.; Keller, N.P. Secondary metabolic gene cluster silencing in Aspergillus nidulans. Mol. Microbiol. 2006, 61, 1636–1645. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.C.; Sun, C.; Xu, J.; Zhang, X.; Luo, H.M.; Ji, A.J.; Hu, Y.L.; Song, J.Y.; Chen, S.L. Progress in the study of Velvet and LaeA proteins and their relation to the development and bioactive compounds in medicinal fungi. Yao Xue Xue Bao 2014, 49, 1520–1527. [Google Scholar] [PubMed]
- Abe, N.; Murata, T.; Hirota, A. Novel oxidized sorbicillin dimers with 1,1-diphenyl-2-picrylhydrazyl-radical scavenging activity from a fungus. Biosci. Biotechnol. Biochem. 1998, 62, 2120–2126. [Google Scholar] [CrossRef] [PubMed]
- Andrade, R.; Ayer, W.A.; Trifonov, L.S. The metabolites of Trichoderma Longibrachiatum. III. Two new tetronic acids: 5-hydroxyvertinolide and bislongiquinolide. Aust. J. Chem. 1997, 50, 255–257. [Google Scholar] [CrossRef]
- Shirota, O.; Pathak, V.; Hossain, C.F.; Sekita, S.; Takatori, K.; Satake, M. Structural elucidation of trichotetronines: Polyketides possessing a bicyclo[2.2.2]octane skeleton with tetronic acid moiety isolated from Trichoderma sp. J. Chem. Soc. Perkin Trans. 1997, 29, 2961–2964. [Google Scholar] [CrossRef]
- Evidente, A.; Andolfi, A.; Cimmino, A.; Ganassi, S.; Altomare, C.; Favilla, M.; Cristofaro, A.D.; Vitagliano, S.; Sabatini, M.A. Bisorbicillinoids Produced by the Fungus Trichoderma Citrinoviride Affect Feeding Preference of the Aphid Schizaphis Graminum. J. Chem. Ecol. 2009, 35, 533–541. [Google Scholar] [CrossRef]
- Du, L.; Zhu, T.; Li, L.; Cai, S.; Zhao, B.; Gu, Q. Cytotoxic sorbicillinoids and bisorbicillinoids from a marine-derived fungus Trichoderma sp. Chem. Pharm. Bull. 2009, 57, 220–223. [Google Scholar] [CrossRef]
- Trifonov, L.S.; Bieri, J.H.; Prewo, R.; Dreiding, A.S.; Hoesch, L.; Rast, D.M. Isolation and structure elucidation of three metabolites from Verticillium intertextum: Sorbicillin, dihydrosorbicillin and bisvertinoquinol. Tetrahedron 1983, 39, 4243–4256. [Google Scholar] [CrossRef]
- Maskey, R.P.; Grün-Wollny, I.; Laatsch, H. Sorbicillin analogues and related dimeric compounds from Penicillium Notatum. J. Nat. Prod. 2005, 68, 865–870. [Google Scholar] [CrossRef]
- Balde, E.S.; Andolfi, A.; Bruyère, C.; Cimmino, A.; Lamoral-Theys, D.; Vurro, M.; Damme, M.V.; Altomare, C.; Mathieu, V.; Kiss, R.; et al. Investigations of fungal secondary metabolites with potential anticancer activity. J. Nat. Prod. 2010, 73, 969. [Google Scholar] [CrossRef]
- Harned, A.M.; Volp, K.A. The sorbicillinoid family of natural products: Isolation, biosynthesis, and synthetic studies. Nat. Prod. Rep. 2011, 28, 1790. [Google Scholar] [CrossRef]
- Tang, M.C.; Cui, X.; He, X.; Ding, Z.; Zhu, T.; Tang, Y.; Li, D. Late-Stage Terpene Cyclization by an Integral Membrane Cyclase in the Biosynthesis of Isoprenoid Epoxycyclohexenone Natural Products. Org. Lett. 2017, 19, 5376–5379. [Google Scholar] [CrossRef]
- Yu, G.; Wu, G.; Sun, Z.; Zhang, X.; Che, Q.; Gu, Q.; Zhu, T.; Li, D.; Zhang, G. Cytotoxic Tetrahydroxanthone Dimers from the Mangrove-Associated Fungus Aspergillus versicolor HDN1009. Mar. Drugs 2018, 16, 335. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. Molecular Evolutionary Genetics Analysis Version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Mitchell, A.L.; Attwood, T.K.; Babbitt, P.C.; Blum, M.; Bork, P.; Bridge, A.; Brown, S.D.; Chang, H.Y.; El-Gebali, S.; Fraser, M.I.; et al. InterPro in 2019: Improving coverage, classification and access to protein sequence annotations. Nucleic Acids Res. 2019, 47, 351–360. [Google Scholar] [CrossRef]
- Ohashi, M.; Liu, F.; Hai, Y.; Chen, M.; Tang, M.C.; Yang, Z.; Sato, M.; Watanabe, K.; Houk, K.N.; Tang, Y. SAM-dependent enzyme-catalysed pericyclic reactions in natural product biosynthesis. Nature 2017, 549, 502–506. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; He, X.; Che, Q.; Zhang, G.; Zhu, T.; Gu, Q.; Li, D. Sorbicillasins A-B and Scirpyrone K from a Deep-Sea-Derived Fungus, Phialocephala sp. FL30r. Mar. Drugs 2018, 16, 245. [Google Scholar] [CrossRef]
- Schwyn, B.; Neilands, J.B. Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 1987, 160, 47–60. [Google Scholar] [CrossRef]
- Jang, J.H.; Kanoh, K.; Adachi, K.; Matsuda, S.; Shizuri, Y. Tenacibactins a-d, hydroxamate siderophores from a marine-derived bacterium, Tenacibaculum sp. a4k-17. J. Nat. Prod. 2007, 70, 563–566. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | 1 | 2 | 3 [16] | 7 |
|---|---|---|---|---|
| 1 | -- | -- | -- | -- |
| 2 | -- | -- | -- | -- |
| 3 | -- | -- | -- | -- |
| 4 | 3.36 s | 3.42 s | 3.36 br. s | 3.39 s |
| 5 | -- | -- | -- | -- |
| 6 | -- | -- | -- | -- |
| 7 | 2.95 d (7.0) | 2.65 d (6.6) | 3.43 d (4.8) | 2.75 d (7.3) |
| 8 | 3.20 br. d (7.0) | 3.15 br. d (6.6) | 3.21 dd (1.2, 4.8) | 3.08 br. d (7.3) |
| 9 | -- | -- | -- | -- |
| 10 | 2.51 t (7.4) | 2.45-2.60a | 6.12 d (15.0) | 2.40–2.50 a |
| 11 | 2.42 m | 2.39 m | 7.33 dd (15.0, 10.6) | 1.70 m |
| 12 | 5.44 dt (15.3, 6.7) | 5.44–5.55 a | 6.28 dd (14.4, 10.6) | 1.30–1.40 a |
| 13 | 5.52 dq (15.3, 6.0) | 5.44–5.55 a | 6.24 dq (14.4, 6.8) | 1.30–1.40 a |
| 14 | 1.64 d (6.0) | 1.59–1.67 a | 1.88 d (6.8) | 0.87 t (7.3) |
| 15 | -- | -- | -- | -- |
| 16 | 6.02 d (15.0) | 2.45–2.60 a | 6.13 d (15.0) | 2.40–2.50 a |
| 17 | 7.16 dd (15.0, 10.6) | 2.11, 2.19 m | 7.22 dd (15.0, 10.4) | 1.48 m |
| 18 | 6.24 dd (15.0, 10.6) | 5.34 m | 6.21 dd (15.2, 10.4) | 1.20–1.30 a |
| 19 | 6.42 m | 5.44–5.55 a | 6.38 dq (15.2, 7.2) | 1.20–1.30 a |
| 20 | 1.93 d (6.7) | 1.59–1.67 a | 1.90 d (7.2) | 0.89 t (6.5) |
| 21 | -- | -- | -- | -- |
| 22 | -- | -- | -- | -- |
| 23 | -- | -- | -- | -- |
| 24 | -- | -- | -- | -- |
| 1-CH3 | 1.05 s | 1.13 s | 1.12 s | 1.11 s |
| 5-CH3 | 1.34 s | 1.31 s | 1.29 s | 1.30 s |
| 21-CH3 | 1.57 s | 1.55 s | 1.51 s | 1.50 s |
| 23-CH3 | 1.61 s | 1.71 s | 1.58 s | 1.67 s |
| 9-OH | 14.45 br. s | 14.45 br. s | 13.99 br. s | 14.46 br. s |
| No. | 1 | 2 | 3 [16] | 7 |
|---|---|---|---|---|
| 1 | 61.7 | 61.4 | 62.6 | 61.4 |
| 2 | 193.0 | 192.9 | 194.9 | 192.9 |
| 3 | 108.4 | 108.4 | 108.5 | 106.3 |
| 4 | 42.8 | 42.7 | 42.4 | 42.9 |
| 5 | 75.3 | 75.2 | 75.0 | 75.3 |
| 6 | 209.9 | 209.1 | 208.3 | 209.2 b |
| 7 | 50.7 | 54.0 | 51.3 | 54.3 |
| 8 | 42.0 | 42.6 | 43.6 | 42.7 |
| 9 | 181.3 | 181.2 | 169.8 | 182.4 |
| 10 | 32.5 | 32.4 | 117.6 | 32.4 |
| 11 | 28.4 | 28.4 | 143.9 | 25.2 |
| 12 | 129.1 | 129.0 | 130.3 | 22.5 |
| 13 | 126.9 | 126.9 | 140.9 | 31.9 |
| 14 | 17.8 | 17.8 | 18.9 | 14.0 |
| 15 | 202.1 | 213.2 | 202.7 | 214.3 |
| 16 | 127.2 | 45.9 | 127.0 | 46.0 |
| 17 | 148.3 | 25.9 | 148.0 | 22.5 |
| 18 | 130.1 | 128.5 | 131.0 | 31.0 |
| 19 | 146.0 | 126.8 | 145.5 | 29.8 |
| 20 | 19.2 | 17.8 | 19.1 | 14.0 |
| 21 | 83.1 | 84.0 | 83.2 | 83.1 |
| 22 | 175.8 | 177.3 | 176.5 | 176.0 |
| 23 | 98.8 | 97.8 | 98.2 | 98.2 |
| 24 | 174.4 | 175.4 | 174.9 | 173.6 |
| 1-CH3 | 10.5 | 10.6 | 11.0 | 10.7 |
| 5-CH3 | 24.1 | 24.0 | 23.5 | 24.2 |
| 21-CH3 | 21.8 | 21.3 | 23.1 | 21.5 |
| 23-CH3 | 6.1 | 5.9 | 6.3 | 6.1 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, J.; Han, H.; Zhang, X.; Ma, C.; Sun, C.; Che, Q.; Gu, Q.; Zhu, T.; Zhang, G.; Li, D. Discovery of Two New Sorbicillinoids by Overexpression of the Global Regulator LaeA in a Marine-Derived Fungus Penicillium dipodomyis YJ-11. Mar. Drugs 2019, 17, 446. https://doi.org/10.3390/md17080446
Yu J, Han H, Zhang X, Ma C, Sun C, Che Q, Gu Q, Zhu T, Zhang G, Li D. Discovery of Two New Sorbicillinoids by Overexpression of the Global Regulator LaeA in a Marine-Derived Fungus Penicillium dipodomyis YJ-11. Marine Drugs. 2019; 17(8):446. https://doi.org/10.3390/md17080446
Chicago/Turabian StyleYu, Jing, Huan Han, Xianyan Zhang, Chuanteng Ma, Chunxiao Sun, Qian Che, Qianqun Gu, Tianjiao Zhu, Guojian Zhang, and Dehai Li. 2019. "Discovery of Two New Sorbicillinoids by Overexpression of the Global Regulator LaeA in a Marine-Derived Fungus Penicillium dipodomyis YJ-11" Marine Drugs 17, no. 8: 446. https://doi.org/10.3390/md17080446
APA StyleYu, J., Han, H., Zhang, X., Ma, C., Sun, C., Che, Q., Gu, Q., Zhu, T., Zhang, G., & Li, D. (2019). Discovery of Two New Sorbicillinoids by Overexpression of the Global Regulator LaeA in a Marine-Derived Fungus Penicillium dipodomyis YJ-11. Marine Drugs, 17(8), 446. https://doi.org/10.3390/md17080446