Effects of Temperature, Growth Media, and Photoperiod on Growth and Toxin Production of Azadinium spinosum


Abstract
:
1. Introduction
2. Results and Discussion
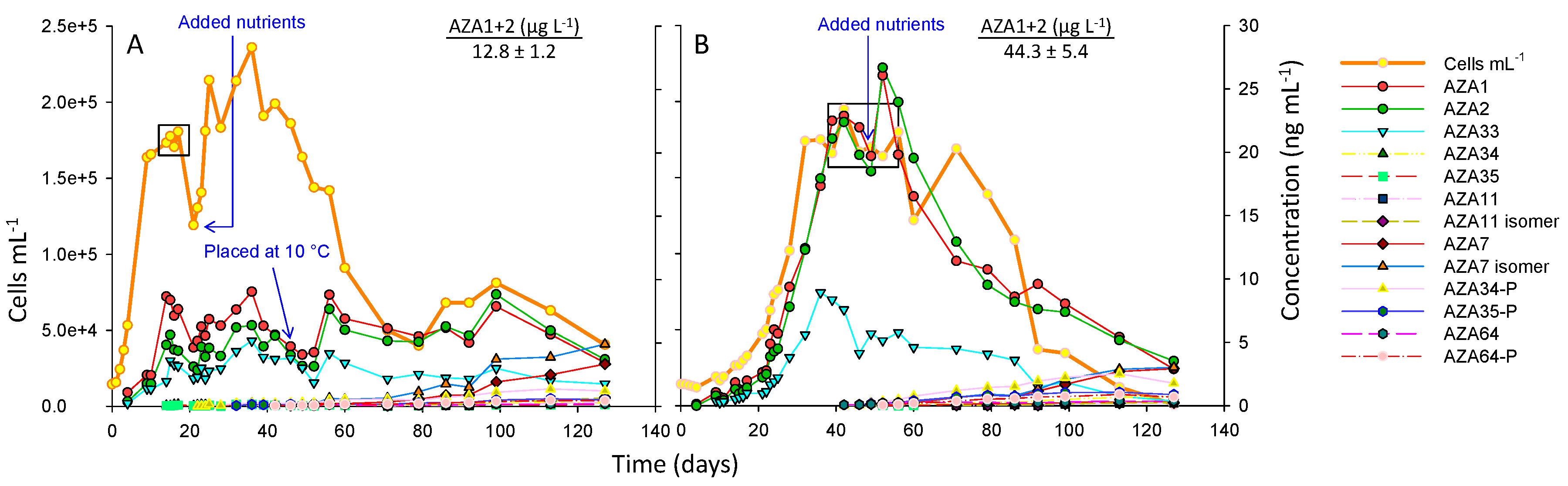
2.1. Impact of Temperature
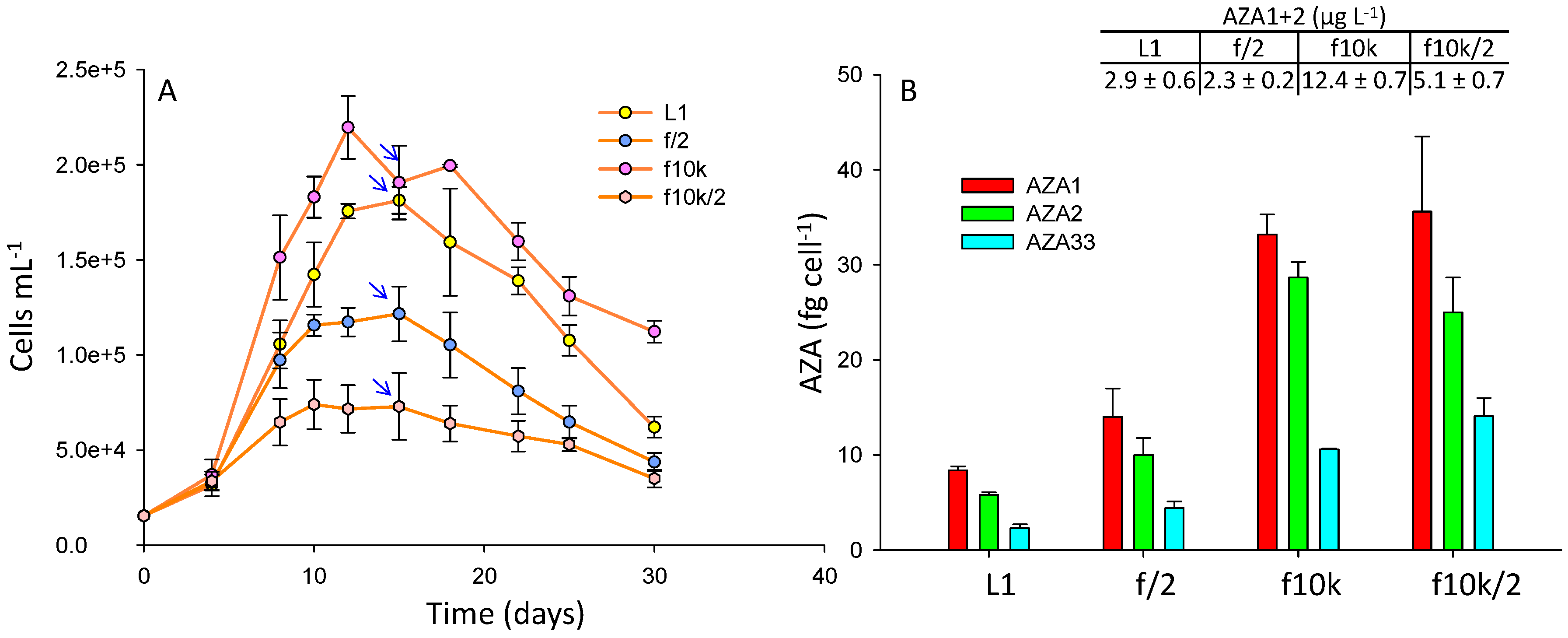
2.2. Impact of Growth Media
2.3. Impact of Photoperiod
2.4. Toxin Profiles
3. Materials and Methods
3.1. Culture Conditions
3.1.1. Culture Sampling
3.1.2. Impact of Temperature
3.1.3. 5 L Culture Flask Harvesting
3.1.4. SPATTs
3.1.5. Periodate Cleavage
3.1.6. Impact of Growth Media
3.1.7. Impact of Photoperiod
3.2. Mass Spectrometry
3.3. Reagents
3.4. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Satake, M.; Ofuji, K.; Naoki, H.; James, K.J.; Furey, A.; McMahon, T.; Silke, J.; Yasumoto, T. Azaspiracid, a new marine toxin having unique spiro ring assemblies, isolated from Irish mussels, Mytilus edulis. J. Am. Chem. Soc. 1998, 120, 9967–9968. [Google Scholar] [CrossRef]
- Rehmann, N.; Hess, P.; Quilliam, M.A. Discovery of new analogs of the marine biotoxin azaspiracid in blue mussels (Mytilus edulis) by ultra-performance liquid chromatography/tandem mass spectrometry. Rapid Commun. Mass Spectrom. 2008, 22, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Krock, B.; Tillmann, U.; Witt, M.; Gu, H. Azaspiracid variability of Azadinium poporum (Dinophyceae) from the China sea. Harmful Algae 2014, 36, 22–28. [Google Scholar] [CrossRef]
- Rossi, R.; Dell’Aversano, C.; Krock, B.; Ciminiello, P.; Percopo, I.; Tillmann, U.; Soprano, V.; Zingone, A. Mediterranean Azadinium dexteroporum (Dinophyceae) produces six novel azaspiracids and azaspiracid-35: A structural study by a multi-platform mass spectrometry approach. Anal. Bioanal. Chem 2017, 409, 1121–1134. [Google Scholar] [CrossRef] [PubMed]
- Kilcoyne, J.; McCarron, P.; Twiner, M.J.; Rise, F.; Hess, P.; Wilkins, A.L.; Miles, C.O. Identification of 21,22-dehydroazaspiracids in mussels (Mytilus edulis) and in vitro toxicity of azaspiracid-26. J. Nat. Prod. 2018, 81, 885–893. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Tillmann, U.; Adams, N.G.; Krock, B.; Stutts, W.L.; Deeds, J.R.; Han, M.S.; Trainer, V.L. Identification of Azadinium species and a new azaspiracid from Azadinium poporum in Puget sound, Washington State, USA. Harmful Algae 2017, 68, 152–167. [Google Scholar] [CrossRef] [PubMed]
- Krock, B.; Tillmann, U.; Tebben, J.; Trefault, N.; Gu, H. Two novel azaspiracids from Azadinium poporum, and a comprehensive compilation of azaspiracids produced by Amphidomataceae, (Dinophyceae). Harmful Algae 2019, 82, 1–8. [Google Scholar] [CrossRef]
- Anon. Commission Regulation (EU) No 15/2011 of 10th January 2011 amending regulation (EC) no 2074/2005 as regards recognised testing methods for detecting marine biotoxins in live bivalve molluscs. Off. J. Eur. Union 2011, L6, 3–6. [Google Scholar]
- HABs. Database. Available online: https://www.marine.ie/Home/site-area/data-services/interactive-maps/weekly-hab-bulletin (accessed on 18 June 2019).
- Kilcoyne, J.; Nulty, C.; Jauffrais, T.; McCarron, P.; Hervé, F.; Wilkins, A.L.; Foley, B.; Rise, F.; Crain, S.; Twiner, M.J.; et al. Isolation, structure elucidation, relative LC-MS response, and in vitro toxicity of azaspiracids from the dinoflagellate Azadinium spinosum. J. Nat. Prod. 2014, 77, 2465–2474. [Google Scholar] [CrossRef]
- Krock, B.; Tillmann, U.; Potvin, É.; Jeong, H.J.; Drebing, W.; Kilcoyne, J.; Bickmeyer, U.; Göthel, Q.; Köck, M. Structural elucidation and toxicity of AZA36 and AZA37, two new azaspiracids isolated from Azadinium poporum. Mar. Drugs 2015, 13, 6687–6702. [Google Scholar] [CrossRef]
- Jauffrais, T.; Herrenknecht, C.; Séchet, V.; Sibat, M.; Tillmann, U.; Krock, B.; Kilcoyne, J.; Miles, C.O.; McCarron, P.; Amzil, Z.; et al. Quantitative analysis of azaspiracids in Azadinium spinosum cultures. Anal. Bioanal. Chem 2012, 403, 833–846. [Google Scholar] [CrossRef] [PubMed]
- Kilcoyne, J.; McCarron, P.; Twiner, M.J.; Nulty, C.; Wilkins, A.L.; Rise, F.; Quilliam, M.A.; Miles, C.O. Epimers of azaspiracids: isolation, structural elucidation, relative LC-MS response, and in vitro toxicity of 37-epi-azaspiracid-1. Chem. Res. Toxicol. 2014, 27, 587–600. [Google Scholar] [CrossRef] [PubMed]
- Tillmann, U.; Edvardsen, B.; Krock, B.; Smith, K.F.; Paterson, R.F.; Voß, D. Diversity, distribution, and azaspiracids of Amphidomataceae (Dinophyceae) along the Norwegian coast. Harmful Algae 2018, 80, 15–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Twiner, M.J.; Hess, P.; Bottein Dechraoui, M.Y.; McMahon, T.; Samons, M.S.; Satake, M.; Yasumoto, T.; Ramsdell, J.S.; Doucette, G.J. Cytotoxic and cytoskeletal effects of azaspiracid-1 on mammalian cell lines. Toxicon 2005, 45, 891–900. [Google Scholar] [CrossRef] [PubMed]
- Twiner, M.J.; Rehmann, N.; Hess, P.; Doucette, G.J. Azaspiracid shellfish poisoning: a review on the chemistry, ecology, and toxicology with an emphasis on human health impacts. Mar. Drugs 2008, 6, 39–72. [Google Scholar] [CrossRef] [PubMed]
- Vale, C.; Wandscheer, C.; Nicolaou, K.C.; Frederick, M.O.; Alfonso, C.; Vieytes, M.R. Cytotoxic effect of azaspiracid-2 and azaspiracid-2-methyl ester in cultured neurons: involvement of the c-Jun N-terminal kinase. J. Neurosci. Res. 2008, 86, 2952–2962. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Lepage, K.T.; Frederick, M.O.; Nicolaou, K.C.; Murray, T.F. Involvement of caspase activation in azaspiracid-induced neurotoxicity in neocortical neurons. Toxicol. Sci. 2010, 114, 323–334. [Google Scholar] [CrossRef] [PubMed]
- Roman, Y.; Alfonso, A.; Vieytes, M.R.; Ofuji, K.; Satake, M.; Yasumoto, T.; Botana, L.M. Effects of azaspiracids 2 and 3 on intracellular cAMP, [Ca2+], and pH. Chem. Res. Toxicol. 2004, 17, 1338–1349. [Google Scholar] [CrossRef] [PubMed]
- Twiner, M.J.; Doucette, G.J.; Rasky, A.; Huang, P.X.; Roth, B.L.; Sanguinetti, M.C. The marine algal toxin azaspiracid is an open state blocker of hERG potassium channels. Chem. Res. Toxicol. 2012, 25, 1975–1984. [Google Scholar] [CrossRef]
- Twiner, M.J.; Hanagriff, J.C.; Butler, S.; Madhkoor, A.K.; Doucette, G.J. Induction of apoptosis pathways in several cell lines following exposure to the marine algal toxin azaspiracid-1. Chem. Res. Toxicol. 2012, 25, 1493–1501. [Google Scholar] [CrossRef]
- Assunção, J.; Guedes, A.C.; Malcata, F.X. Biotechnological and pharmacological applications of biotoxins and other bioactive molecules from dinoflagellates. Mar. Drugs 2017, 15, 393. [Google Scholar] [CrossRef] [PubMed]
- Ofuji, K.; Satake, M.; McMahon, T.; Silke, J.; James, K.J.; Naoki, H.; Oshima, Y.; Yasumoto, T. Two analogs of azaspiracid isolated from mussels, Mytilus edulis, involved in human intoxications in Ireland. Nat. Toxins 1999, 7, 99–102. [Google Scholar] [CrossRef]
- Kilcoyne, J.; Keogh, A.; Clancy, G.; LeBlanc, P.; Burton, I.; Quilliam, M.A.; Hess, P.; Miles, C.O. Improved isolation procedure for azaspiracids from shellfish, structural elucidation of azaspiracid-6, and stability studies. J. Agric. Food Chem. 2012, 60, 2447–2455. [Google Scholar] [CrossRef] [PubMed]
- Ueoka, R.; Ito, A.; Izumikawa, M.; Maeda, S.; Takagi, M.; Shin-ya, K.; Yoshida, M.; van Soest, R.W.M.; Matsunaga, S. Isolation of azaspiracid-2 from a marine sponge Echinoclathria sp. as a potent cytotoxin. Toxicon 2009, 53, 680–684. [Google Scholar] [CrossRef] [PubMed]
- Perez, R.; Rehmann, N.; Crain, S.; LeBlanc, P.; Craft, C.; MacKinnon, S.; Reeves, K.; Burton, I.; Walter, J.A.; Hess, P.; et al. The preparation of certified calibration solutions for azaspiracid-1, -2, and -3, potent marine biotoxins found in shellfish. Anal. Bioanal. Chem. 2010, 398, 2243–2252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelin, M.; Kilcoyne, J.; Nulty, C.; Crain, S.; Hess, P.; Tubaro, A.; Sosa, S. Toxic equivalency factors (TEFs) after acute oral exposure of azaspiracid -1, -2 and -3 in mice. Toxicol. Lett. 2018, 282, 136–146. [Google Scholar] [CrossRef] [PubMed]
- Nicolaou, K.C.; Li, Y.W.; Uesaka, N.; Koftis, T.V.; Vyskocil, S.; Ling, T.T.; Govindasamy, M.; Qian, W.; Bernal, F.; Chen, D.Y.K. Total synthesis of the proposed azaspiracid-1 structure, part 1: construction of the enantiomerically pure C1-C20, C21-C27, and C28-C40 fragments. Angew. Chem. Int. Edit. 2003, 42, 3643–3648. [Google Scholar] [CrossRef]
- Nicolaou, K.C.; Frederick, M.O.; Petrovic, G.; Cole, K.P.; Loizidou, E.Z. Total synthesis and confirmation of the revised structures of azaspiracid-2 and azaspiracid-3. Angew. Chem. Int. Edit. 2006, 45, 2609–2615. [Google Scholar] [CrossRef]
- Kenton, N.T.; Adu-Ampratwum, D.; Okumu, A.A.; Zhang, Z.; Chen, Y.; Nguyen, S.; Xu, J.; Ding, Y.; McCarron, P.; Kilcoyne, J.; et al. Total synthesis of (6R,10R,13R,14R,16R,17R,19S,20R,21R,24S,25S,28S,30S,32R,33R,34R,36S,37S,39R)-azaspiracid-3 reveals non-identity with the natural product. Angew. Chem. Int. Ed. 2018, 57, 805–809. [Google Scholar] [CrossRef]
- McCarron, P.; Kilcoyne, J.; Miles, C.O.; Hess, P. Formation of azaspiracids-3, -4, -6, and -9 via decarboxylation of carboxyazaspiracid metabolites from shellfish. J. Agric. Food Chem. 2009, 57, 160–169. [Google Scholar] [CrossRef]
- Krock, B.; Tillmann, U.; John, U.; Cembella, A.D. Characterization of azaspiracids in plankton size-fractions and isolation of an azaspiracid-producing dinoflagellate from the North sea. Harmful Algae 2009, 8, 254–263. [Google Scholar] [CrossRef]
- Salas, R.; Tillmann, U.; John, U.; Kilcoyne, J.; Burson, A.; Cantwell, C.; Hess, P.; Jauffrais, T.; Silke, J. The role of Azadinium spinosum (Dinophyceae) in the production of azaspiracid shellfish poisoning in mussels. Harmful Algae 2011, 10, 774–783. [Google Scholar] [CrossRef]
- Jauffrais, T.; Séchet, V.; Herrenknecht, C.; Truquet, P.; Savar, V.; Tillmann, U.; Hess, P. Effect of environmental and nutritional factors on growth and azaspiracid production of the dinoflagellate Azadinium spinosum. Harmful Algae 2013, 27, 138–148. [Google Scholar] [CrossRef]
- Tillmann, U.; Elbrächter, M.; Krock, B.; John, U.; Cembella, A.D. Azadinium spinosum gen. et sp. nov. (Dinophyceae) identified as a primary producer of azaspiracid toxins. Eur. J. Phycol. 2009, 44, 63–79. [Google Scholar] [CrossRef]
- Li, A.; Jiang, B.; Chen, H.; Gu, H. Growth and toxin production of Azadinium poporum strains in batch cultures under different nutrient conditions. Ecotoxicol. Environ. Saf. 2016, 127, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Jauffrais, T.; Kilcoyne, J.; Séchet, V.; Herrenknecht, C.; Truquet, P.; Hervé, F.; Bérard, J.B.; Nulty, C.; Taylor, S.; Tillmann, U.; et al. Production and isolation of azaspiracid-1 and -2 from Azadinium spinosum culture in pilot scale photobioreactors. Mar. Drugs 2012, 10, 1360–1382. [Google Scholar] [CrossRef] [PubMed]
- Kilcoyne, J.; Fux, E. Strategies for the elimination of matrix effects in the liquid chromatography tandem mass spectrometry analysis of the lipophilic toxins okadaic acid and azaspiracid-1 in molluscan shellfish. J. Chromatogr. A 2010, 1217, 7123–7130. [Google Scholar] [CrossRef] [PubMed]
- Mitrovic, S.M.; James, K.J.; Furey, A.; McKenzie, L. Effects of selenium, iron and cobalt addition to growth of the toxic marine dinoflagellate, Protoceratium reticulatum, in culture. J. Expt. Mar. Biol. Ecol. 2004, 313, 337–351. [Google Scholar] [CrossRef]
- Keller, M.D.; Selvin, R.C.; Claus, W.; Guillard, R.R.L. Media for the culture of oceanic ultraphytoplankton. J. Phycol. 1987, 23, 633–638. [Google Scholar] [CrossRef]
- Tillmann, U.; Trefault, N.; Krock, B.; Parada-Pozo, G.; de la Iglesia, R.; Vásquez, M. Identification of Azadinium poporum (Dinophyceae) in the southeast Pacific: morphology, molecular phylogeny, and azaspiracid profile characterization. J. Plankton Res. 2017, 39, 350–367. [Google Scholar]
- Tillmann, U.; Borel, C.M.; Barrera, F.; Lara, R.; Krock, B.; Almandoz, G.O.; Witt, M.; Trefault, N. Azadinium poporum from the Argentine continental shelf, southwestern Atlantic, produces azaspiracid-2 and azaspiracid-2 phosphate. Harmful Algae 2016, 51, 40–55. [Google Scholar] [CrossRef] [PubMed]
- Luo, Z.; Krock, B.; Giannakourou, A.; Venetsanopoulou, A.; Pagou, K.; Tillmann, U.; Hu, G. Sympatric occurrence of two Azadinium poporum ribotypes in the eastern Mediterranean sea. Harmful Algae 2018, 78, 75–85. [Google Scholar] [CrossRef] [PubMed]
- Fux, E.; Marcaillou, C.; Mondeguer, F.; Biré, R.; Hess, P. Field and mesocosm trials on passive sampling for the study of adsorption and desorption behaviour of lipophilic toxins with a focus on OA and DTX1. Harmful Algae 2008, 7, 574–583. [Google Scholar] [CrossRef] [Green Version]
- Fux, E.; Biré, R.; Hess, P. Comparative accumulation and composition of lipophilic marine biotoxins in passive samplers and in mussels (M. edulis) on the west coast of Ireland. Harmful Algae 2009, 8, 523–537. [Google Scholar] [CrossRef]
- Guillard, R.R.L.; Hargraves, P.E. Stichochrysis immobilis is a diatom, not a chrysophyte. Phycol. 1993, 32, 234–236. [Google Scholar] [CrossRef]
- Guillard, R.R.L.; Ryther, J.H. Studies on marine planktonic diatoms I. Cyclotella nana Hustedt and Detonula confervacea (Cleve) Gran. Can. J. Microbiol. 1962, 8, 229–239. [Google Scholar] [CrossRef] [PubMed]
- MacKenzie, L.; Beuzenberg, V.; Holland, P.; McNabb, P.; Selwood, A. Solid Phase Adsorption Toxin Tracking (SPATT): A new monitoring tool that simulates the biotoxin contamination of filter feeding bivalves. Toxicon 2004, 44, 901–918. [Google Scholar] [CrossRef]
- Holmes, M.J.; Lewis, R.J.; Poli, M.A.; Gillespie, N.C. Strain dependent production of ciguatoxin precursors (gambiertoxins) by Gambierdiscus toxicus (Dinophyceae) in culture. Toxicon 1991, 29, 761–775. [Google Scholar] [CrossRef]
- Kilcoyne, J.; Twiner, M.J.; McCarron, P.; Crain, S.; Giddings, S.D.; Wilkins, A.L.; Hess, P.; Miles, C.O. Structure elucidation, relative LC-MS and in vitro toxicity of azaspiracids 7–10 isolated from mussels (Mytilus edulis). J. Agric. Food Chem. 2015, 63, 5083–5091. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 18 °C | 10 °C | |||||
|---|---|---|---|---|---|---|
| AZA1 | AZA2 | AZA33 | AZA1 | AZA2 | AZA33 | |
| % filtrate * | 19.5 | 65.9 | 70.5 | 22.7 | 59.4 | 34.3 |
| % adsorbed onto culture flask walls | 6.2 | 24.7 | 29.5 | 14.7 | 35.7 | 65.7 |
| % C3 hydroxylated # | 74.3 | 9.5 | 62.7 | 4.9 | ||
| AZA | Molecular Formula ([M + H]+) | Measured m/z [M + H]+ | Δ (ppm) |
|---|---|---|---|
| AZA34-P | C45H71NO12PO3 | 896.4547 | −0.99 |
| AZA35-P | C46H73NO12PO3 | 910.4738 | 2.21 |
| AZA64 | C44H68NO12 | 802.4742 | 0.74 |
| AZA64-P | C44H69NO12PO3 | 882.4394 | −0.61 |
| AZA | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Temperature | 1 | 2 | 33 | 34 | 35 | 11 | 11 isomer | 7 | 7 isomer | 34-P | 35-P | 64 | 64-P |
| 10 °C | 5.8 | 6.4 | 0.6 | 1.4 | 0.6 | 0.2 | 0.3 | 4.8 | 4.9 | 3.6 | 1.2 | 0.6 | 1.0 |
| 18 °C | 3.3 | 3.7 | 0.7 | 0.4 | 0.3 | 0.2 | 0.4 | 4.1 | 5.4 | 2.1 | 0.7 | 0.3 | 0.4 |
| AZA | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Depth (m) | 1 | 2 | 33 | 34 | 35 | 11 | 11 isomer | 7 | 7 isomer | 34-P | 35-P | 64 | 64-P | 1-P | 2-P |
| 0 | 1031.2 | 423.5 | 47.7 | 10.1 | 1.1 | 4.4 | 1.1 | 48.5 | 10.9 | 5.9 | 1.1 | 132.5 | 32.2 | 67.5 | 24.3 |
| 5 | 999.1 | 383.2 | 46.7 | 6.8 | n.d. | 2.8 | 0.6 | 1.1 | 10.1 | 3.1 | 0.5 | 86.6 | 34.9 | 58.5 | 38.5 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kilcoyne, J.; McCoy, A.; Burrell, S.; Krock, B.; Tillmann, U. Effects of Temperature, Growth Media, and Photoperiod on Growth and Toxin Production of Azadinium spinosum. Mar. Drugs 2019, 17, 489. https://doi.org/10.3390/md17090489
Kilcoyne J, McCoy A, Burrell S, Krock B, Tillmann U. Effects of Temperature, Growth Media, and Photoperiod on Growth and Toxin Production of Azadinium spinosum. Marine Drugs. 2019; 17(9):489. https://doi.org/10.3390/md17090489
Chicago/Turabian StyleKilcoyne, Jane, Amy McCoy, Stephen Burrell, Bernd Krock, and Urban Tillmann. 2019. "Effects of Temperature, Growth Media, and Photoperiod on Growth and Toxin Production of Azadinium spinosum" Marine Drugs 17, no. 9: 489. https://doi.org/10.3390/md17090489
APA StyleKilcoyne, J., McCoy, A., Burrell, S., Krock, B., & Tillmann, U. (2019). Effects of Temperature, Growth Media, and Photoperiod on Growth and Toxin Production of Azadinium spinosum. Marine Drugs, 17(9), 489. https://doi.org/10.3390/md17090489