Worms’ Antimicrobial Peptides

 ,
,  , , , ,
, , , ,  , ,
, ,  and
and 
Abstract
:1. Introduction
2. AMPs Diversity in Annelids and Nematodes
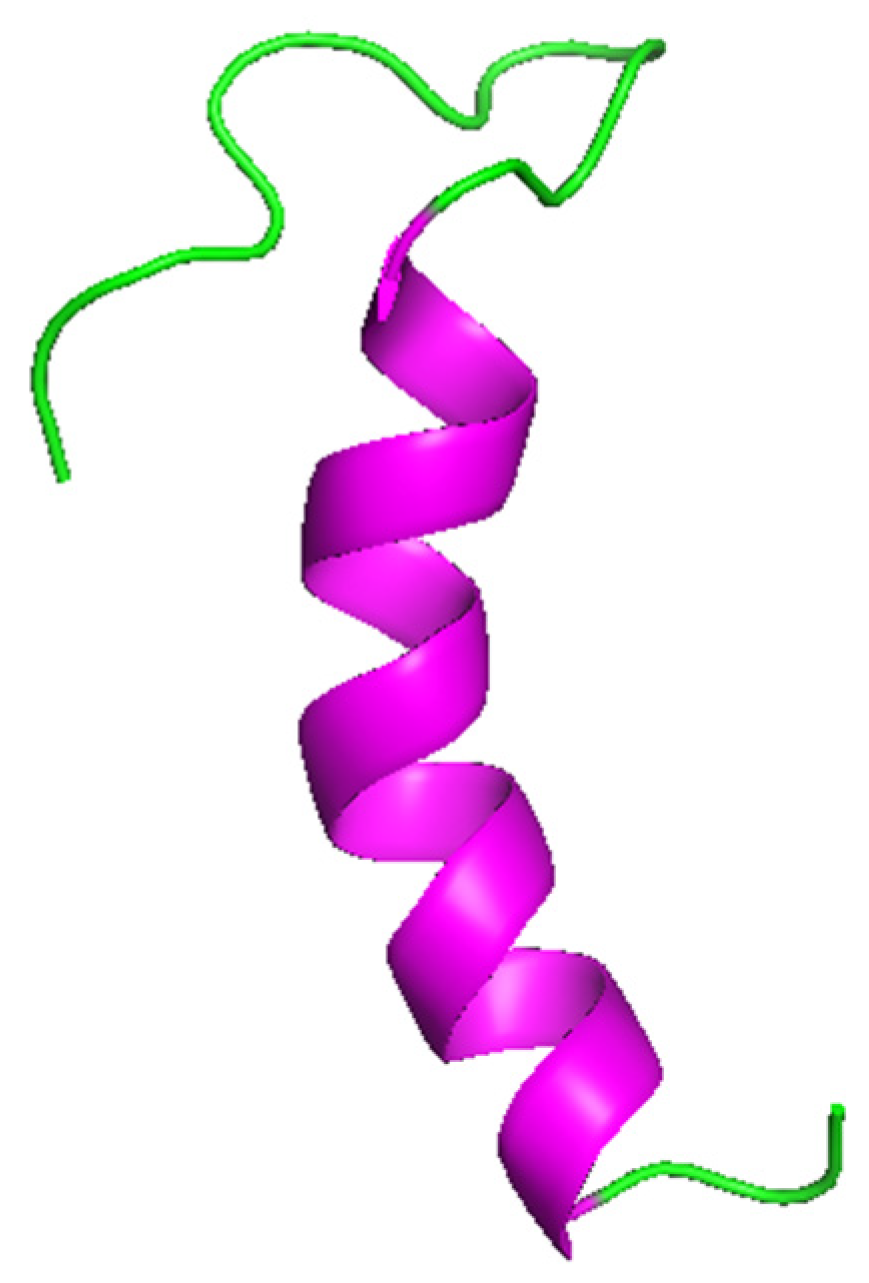
2.1. α-helix Peptides
2.1.1. α-helix Peptides in Nematodes
Cecropin and Caenopore Families
2.1.2. α-helix Peptides in Annellids
Hedistin
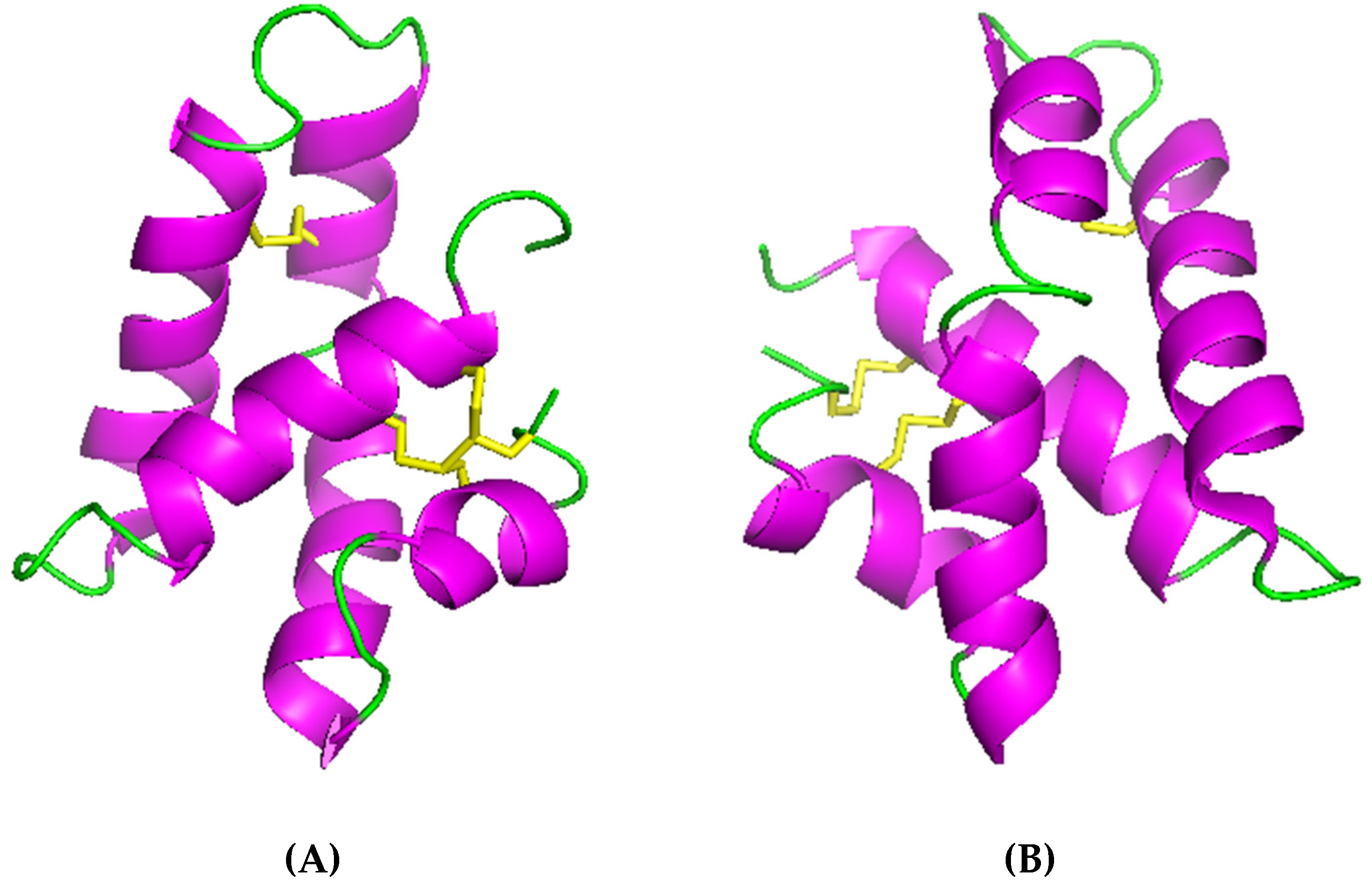
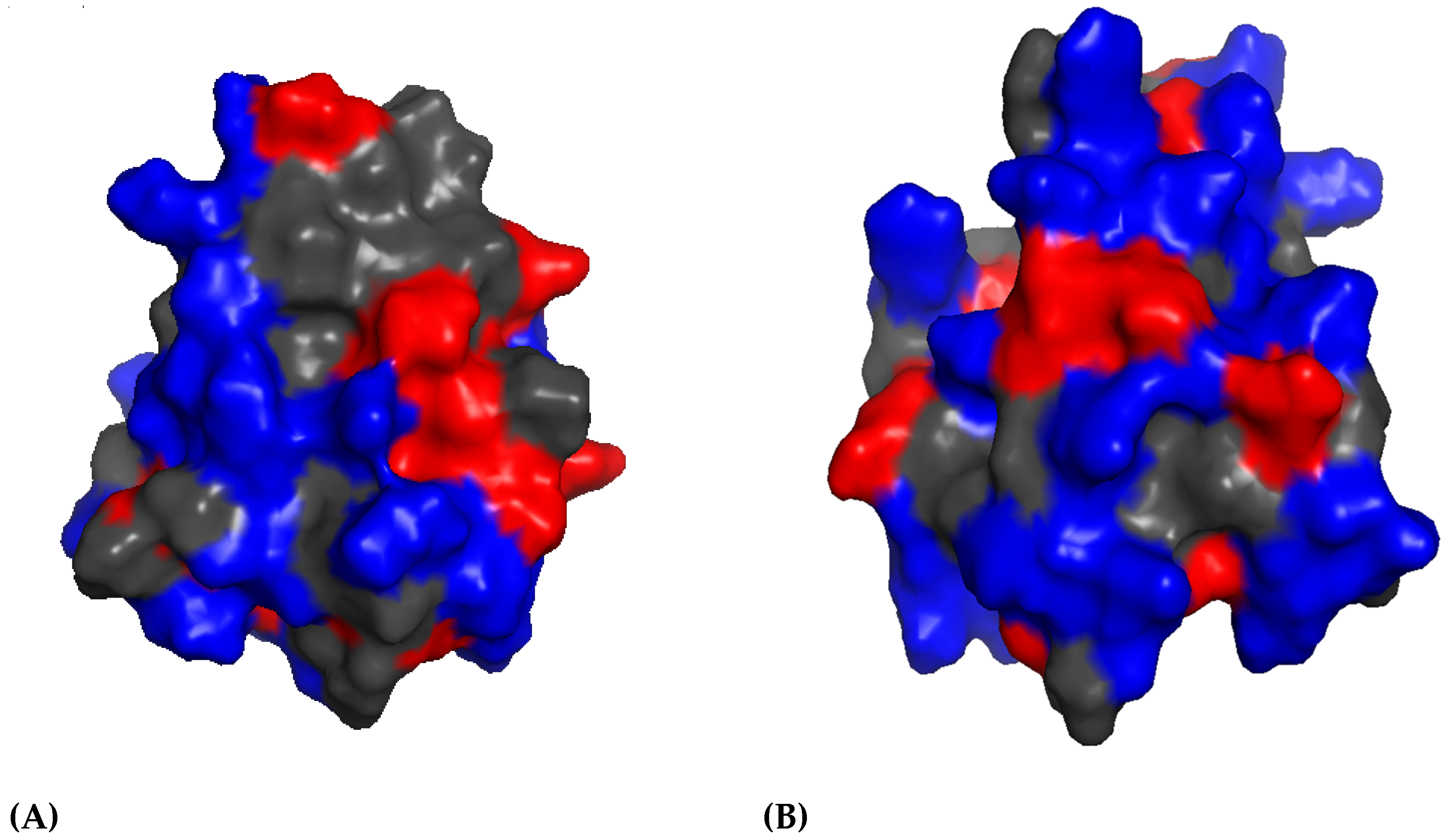
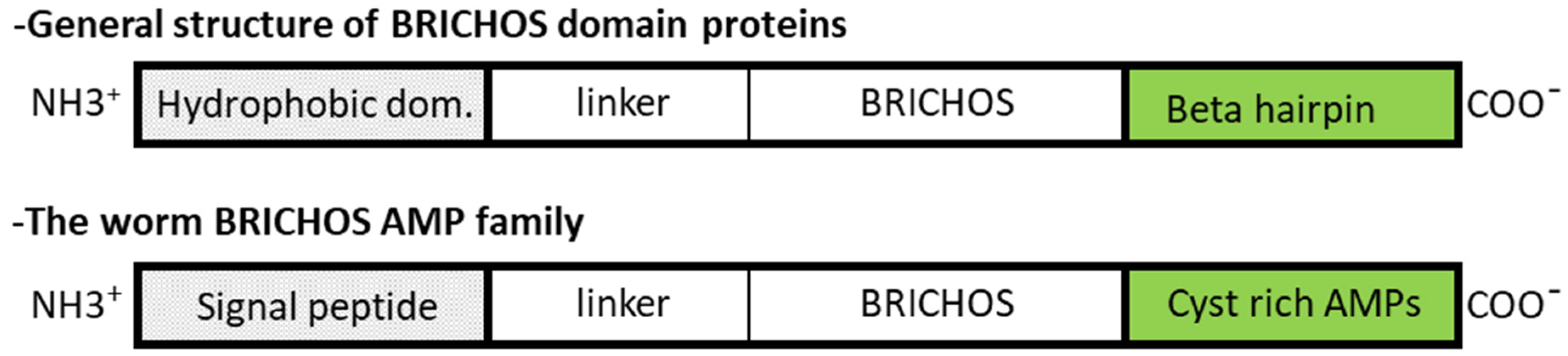
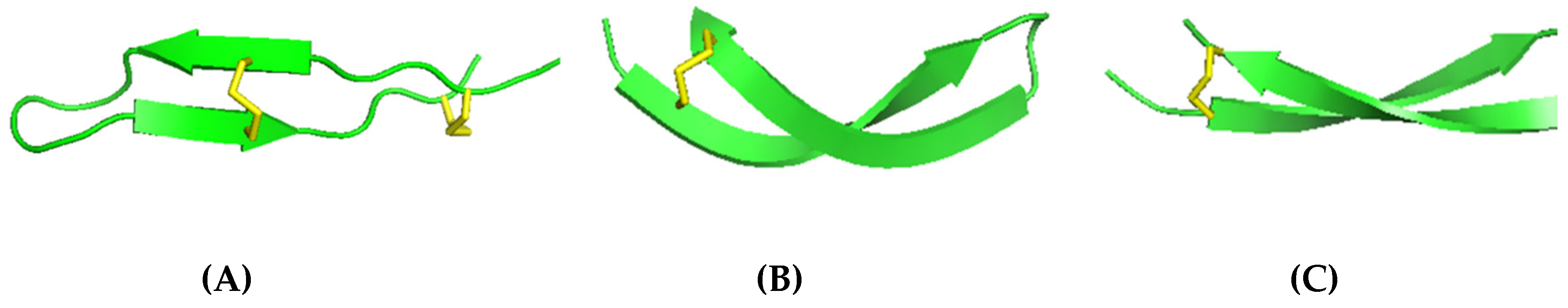
2.2. β-sheet Peptides in Annelids
BRICHOS-AMPs Family
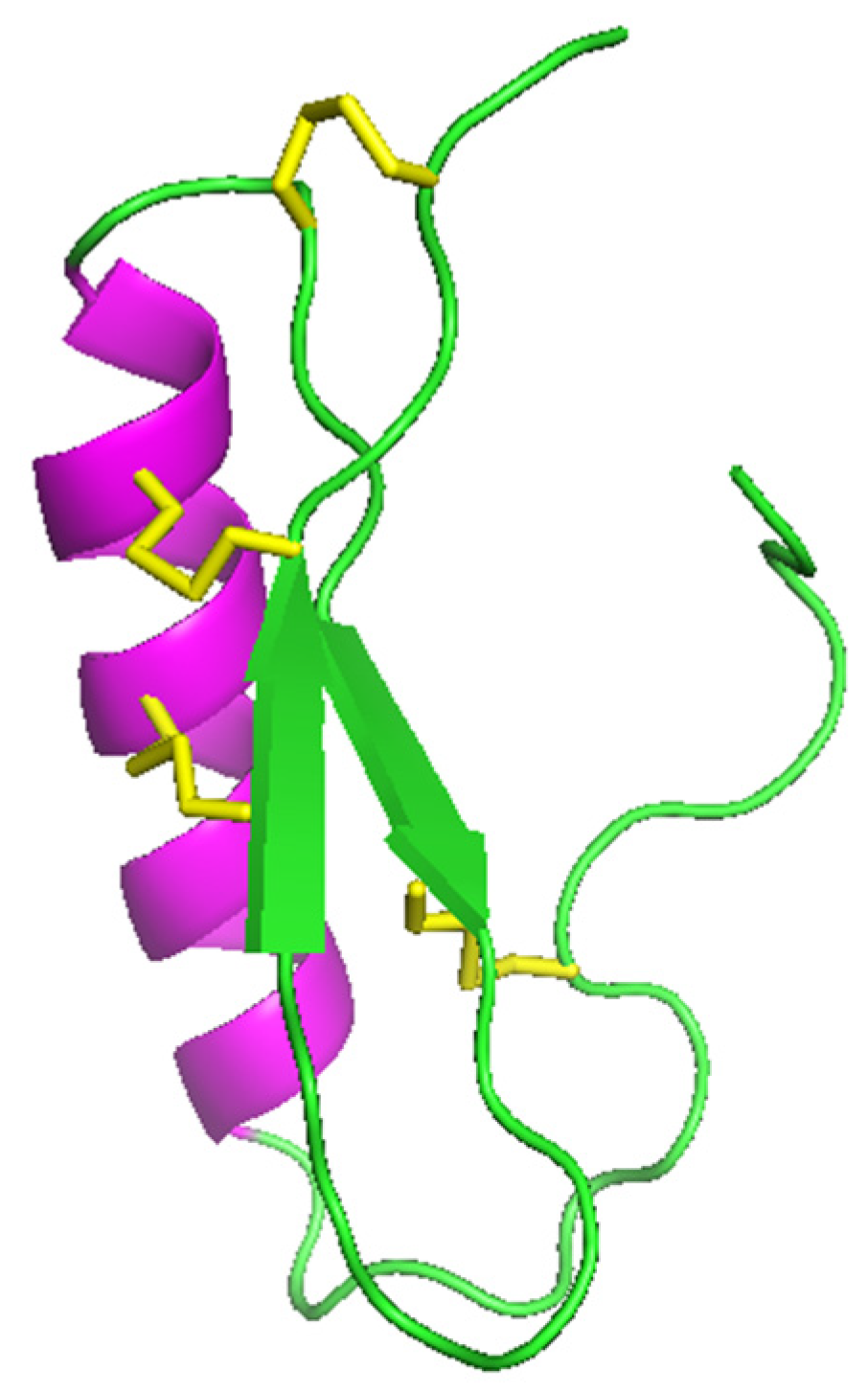
2.3. Mixed α-helix/β-sheet Peptides
2.3.1. Mixed α-helix/β-sheet Peptides in Nematodes
The ABF Family
2.3.2. Mixed α-helix/β-sheet Peptides in Annelids
Macin Family
2.4. Peptides Enriched with One or Two Specific Amino Acids
2.4.1. Peptides Enriched with One or Two Specific Amino Acids in Nematodes
Neuropeptide-Like Peptides and Caenacins
2.4.2. Peptides enriched with one or two specific amino acids in annelids
Lumbricin Family
2.5. Peptides Derived from Larger Molecules in Annelids
2.5.1. Perinerin
2.5.2. Ms-Hemerycin
3. Conclusions and Perspectives
Funding
Acknowledgments
Conflicts of Interest
References
- O’Connell, K.M.G.; Hodgkinson, J.T.; Sore, H.F.; Welch, M.; Salmond, G.P.C.; Spring, D.R. Combating Multidrug-Resistant Bacteria: Current Strategies for the Discovery of Novel Antibacterials. Angew. Chem. Int. Ed. 2013, 52, 10706–10733. [Google Scholar] [CrossRef] [PubMed]
- Pasupuleti, M.; Schmidtchen, A.; Malmsten, M. Antimicrobial peptides: Key components of the innate immune system. Crit. Rev. Biotechnol. 2012, 32, 143–171. [Google Scholar] [CrossRef] [PubMed]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef] [PubMed]
- Bulet, P.; Stöcklin, R.; Menin, L. Anti-microbial peptides: From invertebrates to vertebrates. Immunol. Rev. 2004, 198, 169–184. [Google Scholar] [CrossRef] [PubMed]
- Jenssen, H.; Hamill, P.; Hancock, R.E.W. Peptide Antimicrobial Agents. Clin. Microbiol. Rev. 2006, 19, 491–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, S.; Sönnichsen, F.D.; Hung, C.W.; Tholey, A.; Boidin-Wichlacz, C.; Haeusgen, W.; Gelhaus, C.; Desel, C.; Podschun, R.; Waetzig, V.; et al. Macin Family of Antimicrobial Proteins Combines Antimicrobial and Nerve Repair Activities. J. Biol. Chem. 2012, 287, 14246–14258. [Google Scholar] [CrossRef] [Green Version]
- Steckbeck, J.D.; Deslouches, B.; Montelaro, R.C. Antimicrobial peptides: New drugs for bad bugs? Expert Opin. Biol. Ther. 2014, 14, 11–14. [Google Scholar] [CrossRef]
- Boman, H.G. Peptide Antibiotics and their Role in Innate Immunity. Annu. Rev. Immunol. 1995, 13, 61–92. [Google Scholar] [CrossRef]
- Brogden, K.A. Antimicrobial peptides: Pore formers or metabolic inhibitors in bacteria? Nat. Rev. Genet. 2005, 3, 238–250. [Google Scholar] [CrossRef]
- Da Costa, J.P.; Cova, M.; Ferreira, R.; Vitorino, R. Antimicrobial peptides: An alternative for innovative medicines? Appl. Microbiol. Biotechnol. 2015, 99, 2023–2040. [Google Scholar] [CrossRef]
- Marr, A.; Gooderham, W.; Hancock, R. Antibacterial peptides for therapeutic use: Obstacles and realistic outlook. Curr. Opin. Pharmacol. 2006, 6, 468–472. [Google Scholar] [CrossRef] [PubMed]
- Sherlina Daphny, C.; Arputha Bibiana, M.; Vengatesan, R.; Selvamani, P.; Latha, S. Antimicrobial Peptides-A milestone for developing antibiotics against drug resistant infectious pathogens. J. Pharm. Sci. Res. 2015, 7, 226–230. [Google Scholar]
- Macke, E.; Tasiemski, A.; Callens, M.; Decaestecker, E.; Massol, F. Life history and eco-evolutionary dynamics in light of the gut microbiota. Oikos 2017, 126, 508–531. [Google Scholar] [CrossRef] [Green Version]
- Ovchinnikova, T.V.; Aleshina, G.M.; Balandin, S.V.; Krasnosdembskaya, A.D.; Markelov, M.L.; Frolova, E.I.; Leonova, Y.F.; Tagaev, A.A.; Krasnodembsky, E.G.; Kokryakov, V.N. Purification and primary structure of two isoforms of arenicin, a novel antimicrobial peptide from marine polychaetaArenicola marina. FEBS Lett. 2004, 577, 209–214. [Google Scholar] [CrossRef] [PubMed]
- Tasiemski, A.; Massol, F.; Cuvillier-Hot, V.; Boidin-Wichlacz, C.; Roger, E.; Rodet, F.; Fournier, I.; Thomas, F.; Salzet, M. Reciprocal immune benefit based on complementary production of antibiotics by the leech Hirudo verbana and its gut symbiont Aeromonas veronii. Sci. Rep. 2015, 5, 17498. [Google Scholar] [CrossRef] [PubMed]
- Amato, K.R. Incorporating the gut microbiota into models of human and non-human primate ecology and evolution. Am. J. Phys. Anthr. 2016, 159, 196–215. [Google Scholar] [CrossRef]
- Sousa, R.; Dias, S.; Antunes, C. Subtidal macrobenthic structure in the lower lima estuary, NW of Iberian Peninsula. Ann. Zool. Fennici 2007, 44, 303–313. [Google Scholar] [CrossRef]
- Zhao, C.; Liaw, L.; Lee, I.H.; I Lehrer, R. cDNA cloning of three cecropin-like antimicrobial peptides (Styelins) from the tunicate, Styela clava. FEBS Lett. 1997, 412, 144–148. [Google Scholar] [CrossRef]
- Andersson, M.; Boman, A.; Boman, H.G. Ascaris nematodes from pig and human make three anti-bacterial peptides: Isolation of cecropin P1 and two ASABF peptides. Cell. Mol. Life Sci. 2003, 60, 599–606. [Google Scholar] [CrossRef]
- Meyer, V.; Jung, S. Antifungal Peptides of the AFP Family Revisited: Are These Cannibal Toxins? Microorganisms 2018, 6, 50. [Google Scholar] [CrossRef]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [PubMed]
- Yeaman, M.R.; Yount, N.Y. Unifying themes in host defence effector polypeptides. Nat. Rev. Genet. 2007, 5, 727–740. [Google Scholar] [CrossRef] [PubMed]
- Peschel, A.; Sahl, H.G. The co-evolution of host cationic antimicrobial peptides and microbial resistance. Nat. Rev. Genet. 2006, 4, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Malanovic, N.; Lohner, K. Antimicrobial Peptides Targeting Gram-Positive Bacteria. Pharmaceuticals 2016, 9, 59. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.L. Antimicrobial peptides stage a comeback. Nat. Biotechnol. 2013, 31, 379–382. [Google Scholar] [CrossRef] [PubMed]
- Salzet, M.; Tasiemski, A.; Cooper, E. Innate immunity in lophotrochozoans: The annelids. Curr. Pharm. Des. 2006, 12, 3043–3050. [Google Scholar] [CrossRef] [PubMed]
- Bogaerts, A.; Beets, I.; Schoofs, L.; Verleyen, P. Antimicrobial peptides in Caenorhabditis elegans. Invertebr. Surviv. J. 2010, 7, 45–52. [Google Scholar]
- Berlov, M.N.; Maltseva, A.L. Immunity of the lugworm Arenicola marina: Cells and molecules. Invertebr. Surviv. J. 2016, 13, 247–256. [Google Scholar]
- Tarr, D.E.K. Distribution and characteristics of ABFs, cecropins, nemapores, and lysozymes in nematodes. Dev. Comp. Immunol. 2012, 36, 502–520. [Google Scholar] [CrossRef]
- Cyrino, M.; Coutinho, L.; Teixeira, V.L.; Simone, C.; Santos, G. A Review of “Polychaeta” Chemicals and their Possible Ecological Role. J. Chem. Ecol. 2017, 44, 1–23. [Google Scholar]
- Lee, J.Y.; Boman, A.; Sun, C.X.; Andersson, M.; Jörnvall, H.; Mutt, V.; Boman, H.G. Antibacterial peptides from pig intestine: Isolation of a mammalian cecropin. Proc. Natl. Acad. Sci. USA 1989, 86, 9159–9162. [Google Scholar] [CrossRef] [PubMed]
- Hultmark, D.; Steiner, H.; Rasmuson, T.; Boman, H.G. Insect immunity. Purification and properties of three inducible bactericidal proteins from hemolymph of immunized pupae of Hyalophora cecropia. JBIC J. Biol. Inorg. Chem. 1980, 106, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Lockey, T.D.; Ourth, D.D. Formation of Pores in Escherichia coli Cell Membranes by a Cecropin Isolated from Hemolymph of Heliothis virescens Larvae. JBIC J. Biol. Inorg. Chem. 1996, 236, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Pütsep, K.; Normark, S.; Boman, H.G. The origin of cecropins; implications from synthetic peptides derived from ribosomal protein L1. FEBS Lett. 1999, 451, 249–252. [Google Scholar] [CrossRef] [Green Version]
- Kato, Y.; Komatsu, S. ASABF, a Novel Cysteine-rich Antibacterial Peptide Isolated from the Nematode Ascaris suum: Purification, primary structure, and molecular cloning of cDNA. J. Biol. Chem. 1996, 271, 30493–30498. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Kato, Y. Common structural properties specifically found in the CSαβ-type antimicrobial peptides in nematodes and mollusks: Evidence for the same evolutionary origin? Dev. Comp. Immunol. 2003, 27, 499–503. [Google Scholar] [CrossRef]
- Froy, O. Convergent evolution of invertebrate defensins and nematode antibacterial factors. Trends Microbiol. 2005, 13, 314–319. [Google Scholar] [CrossRef] [PubMed]
- Bányai, L.; Patthy, L. Amoebapore homologs of Caenorhabditis elegans. Biochim. Biophys. Acta (BBA) Protein Struct. Mol. Enzym. 1998, 1429, 259–264. [Google Scholar] [CrossRef]
- Leippe, M. Antimicrobial and cytolytic polypeptides of amoeboid protozoa - effector molecules of primitive phagocytes. Dev. Comp. Immunol. 1999, 23, 267–279. [Google Scholar] [CrossRef]
- Couillault, C.; Pujol, N.; Reboul, J.; Sabatier, L.; Guichou, J.F.; Kohara, Y.; Ewbank, J.J. TLR-independent control of innate immunity in Caenorhabditis elegans by the TIR domain adaptor protein TIR-1, an ortholog of human SARM. Nat. Immunol. 2004, 5, 488–494. [Google Scholar] [CrossRef]
- Sperstad, S.V.; Haug, T.; Blencke, H.M.; Styrvold, O.B.; Li, C.; Stensvåg, K. Antimicrobial peptides from marine invertebrates: Challenges and perspectives in marine antimicrobial peptide discovery. Biotechnol. Adv. 2011, 29, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.H.; Park, C.B.; Yoon, Y.G.; Kim, S.C. Lumbricin I, a novel proline-rich antimicrobial peptide from the earthworm: Purification, cDNA cloning and molecular characterization. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 1998, 1408, 67–76. [Google Scholar] [CrossRef]
- Schikorski, D.; Cuvillier-Hot, V.; Leippe, M.; Boidin-Wichlacz, C.; Slomianny, C.; Macagno, E.; Salzet, M.; Tasiemski, A. Microbial challenge promotes the regenerative process of the injured central nervous system of the medicinal leech by inducing the synthesis of antimicrobial peptides in neurons and microglia. J. Immunol. 2008, 181, 1083–1095. [Google Scholar] [CrossRef] [PubMed]
- Tasiemski, A.; Vandenbulcke, F.; Mitta, G.; Lemoine, J.; Lefebvre, C.; Sautiere, P.-E.; Salzet, M. Molecular Characterization of Two Novel Antibacterial Peptides Inducible upon Bacterial Challenge in an Annelid, the Leech Theromyzon tessulatum. J. Biol. Chem. 2004, 279, 30973–30982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanchez-Pulido, L.; Devos, D.; Valencia, A. BRICHOS: A conserved domain in proteins associated with dementia, respiratory distress and cancer. Trends Biochem. Sci. 2002, 27, 329–332. [Google Scholar] [CrossRef]
- Knight, S.D.; Presto, J.; Linse, S.; Johansson, J. The brichos Domain, Amyloid Fibril Formation, and Their Relationship. Biochemistry 2013, 52, 7523–7531. [Google Scholar] [CrossRef] [PubMed]
- Papot, C.; Massol, F.; Jollivet, D.; Tasiemski, A. Antagonistic evolution of an antibiotic and its molecular chaperone: How to maintain a vital ectosymbiosis in a highly fluctuating habitat. Sci. Rep. 2017, 7, 389. [Google Scholar]
- Tasiemski, A.; Jung, S.; Boidin-Wichlacz, C.; Jollivet, D.; Cuvillier-Hot, V.; Pradillon, F.; Vetriani, C.; Hecht, O.; Sönnichsen, F.D.; Gelhaus, C.; et al. Characterization and Function of the First Antibiotic Isolated from a Vent Organism: The Extremophile Metazoan Alvinella pompejana. PLoS ONE 2014, 9, e95737. [Google Scholar] [CrossRef] [PubMed]
- Pan, W.; Liu, X.; Ge, F.; Han, J.; Zheng, T. Perinerin, a novel antimicrobial peptide purified from the clamworm Perinereis aibuhitensis grube and its partial characterization. J. Biochem. 2004, 135, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Tasiemski, A.; Schikorski, D.; Le Marrec-Croq, F.; Camp, C.P.V.; Boidin-Wichlacz, C.; Sautière, P.E. Hedistin: A novel antimicrobial peptide containing bromotryptophan constitutively expressed in the NK cells-like of the marine annelid, Nereis diversicolor. Dev. Comp. Immunol. 2007, 31, 749–762. [Google Scholar] [CrossRef]
- Seo, J.K.; Nam, B.H.; Go, H.J.; Jeong, M.; Lee, K.Y.; Cho, S.M.; Lee, I.A.; Park, N.G. Hemerythrin-related antimicrobial peptide, msHemerycin, purified from the body of the Lugworm, Marphysa sanguinea. Fish Shellfish. Immunol. 2016, 57, 49–59. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Harroun, T.A.; Weiss, T.M.; Ding, L.; Huang, H.W. Barrel-stave model or toroidal model? A case study on melittin pores. Biophys. J. 2001, 81, 1475–1485. [Google Scholar] [CrossRef]
- Ladokhin, A.S.; White, S.H. ‘Detergent-like’ permeabilization of anionic lipid vesicles by melittin. Biochim. Biophys. Acta (BBA) Biomembr. 2001, 1514, 253–260. [Google Scholar] [CrossRef]
- Epand, R.F.; Maloy, L.; Ramamoorthy, A.; Epand, R.M. Amphipathic Helical Cationic Antimicrobial Peptides Promote Rapid Formation of Crystalline States in the Presence of Phosphatidylglycerol: Lipid Clustering in Anionic Membranes. Biophys. J. 2010, 98, 2564–2573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nicolas, P. Multifunctional host defense peptides: Intracellular-targeting antimicrobial peptides. FEBS J. 2009, 276, 6483–6496. [Google Scholar] [CrossRef] [PubMed]
- Giangaspero, A.; Sandri, L.; Tossi, A.; Database, A.S. Amphipathic a helical antimicrobial peptides activity. Sabouraudia 2001, 5600, 5589–5600. [Google Scholar]
- Powers, J.P.S.; Hancock, R.E. The relationship between peptide structure and antibacterial activity. Peptides 2003, 24, 1681–1691. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Haney, E.F.; Vogel, H.J. The expanding scope of antimicrobial peptide structures and their modes of action. Trends Biotechnol. 2011, 29, 464–472. [Google Scholar] [CrossRef]
- Reddy, K.; Yedery, R.; Aranha, C. Antimicrobial peptides: Premises and promises. Int. J. Antimicrob. Agents 2004, 24, 536–547. [Google Scholar] [CrossRef]
- Kylsten, P.; Samakovlis, C.; Hultmark, D. The cecropin locus in Drosophila; a compact gene cluster involved in the response to infection. EMBO J. 1990, 9, 217–224. [Google Scholar] [CrossRef]
- Liang, Y.; Wang, J.X.; Zhao, X.F.; Du, X.J.; Xue, J.F. Molecular cloning and characterization of cecropin from the housefly (Musca domestica), and its expression in Escherichia coli. Dev. Comp. Immunol. 2006, 30, 249–257. [Google Scholar] [CrossRef] [PubMed]
- Pillai, A.; Ueno, S.; Zhang, H.; Lee, J.M.; Kato, Y. Cecropin P1 and novel nematode cecropins: A bacteria-inducible antimicrobial peptide family in the nematode Ascaris suum. Biochem. J. 2005, 390, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Baek, M.H.; Kamiya, M.; Kushibiki, T.; Nakazumi, T.; Tomisawa, S.; Abe, C.; Kumaki, Y.; Kikukawa, T.; Demura, M.; Kawano, K.; et al. Lipopolysaccharide-bound structure of the antimicrobial peptide cecropin P1 determined by nuclear magnetic resonance spectroscopy. J. Pept. Sci. 2016, 22, 214–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tassanakajon, A.; Somboonwiwat, K.; Amparyup, P. Sequence diversity and evolution of antimicrobial peptides in invertebrates. Dev. Comp. Immunol. 2014, 48, 324–341. [Google Scholar] [CrossRef] [PubMed]
- Satchell, D.P.; Sheynis, T.; Shirafuji, Y.; Kolusheva, S.; Ouellette, A.J.; Jelinek, R. Interactions of Mouse Paneth Cell α-Defensins and α-Defensin. J. Biol. Chem. 2003, 278, 13838–13846. [Google Scholar] [CrossRef] [PubMed]
- Andrä, J.; Berninghausen, O.; Leippe, M. Cecropins, antibacterial peptides from insects and mammals, are potently fungicidal against Candida albicans. Med Microbiol. Immunol. 2001, 189, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Hee Lee, I.N.; Cho, Y.; Lehrer, R.I. Effects of pH and salinity on the antimicrobial properties of clavanins. Infect. Immun. 1997, 65, 2898–2903. [Google Scholar] [Green Version]
- Chiou, P.P.; Chen, M.J.; Lin, C.; Khoo, J.; Larson, J.; Holt, R.; Leong, J.; Thorgarrd, G.; Chen, T.T. Production of Homozygous Transgenic Rainbow Trout with Enhanced Disease Resistance. Mar. Biotechnol. 2014, 299–308. [Google Scholar] [CrossRef]
- Han, Y.; Chen, T.T. Fish and Shellfish Immunology A pathway-focused RT-qPCR array study on immune relevant genes in rainbow trout (Oncorhynchus mykiss) harboring cecropin P1 transgene. Fish Shellfish Immunol. 2019, 89, 1–11. [Google Scholar] [CrossRef]
- Song, K.D.; Lee, W.K. Antibacterial Activity of Recombinant Pig Intestinal Parasite Cecropin P4 Peptide Secreted from Pichia pastoris. Asian Australas. J. Anim. Sci. 2014, 27, 278–283. [Google Scholar] [CrossRef] [Green Version]
- Bruhn, H. A short guided tour through functional and structural features of saposin-like proteins. Biochem. J. 2005, 389, 249–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roeder, T.; Stanisak, M.; Gelhaus, C.; Bruchhaus, I.; Grotzinger, J.; Leippe, M. Caenopores are antimicrobial peptides in the nematode Caenorhabditis elegans instrumental in nutrition and immunity. Dev. Comp. Immunol. 2010, 34, 203–209. [Google Scholar] [CrossRef] [PubMed]
- Dierking, K.; Yang, W.; Schulenburg, H. Antimicrobial effectors in the nematode Caenorhabditis elegans: An outgroup to the Arthropoda. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150299. [Google Scholar] [CrossRef] [PubMed]
- Ewbank, J.J.; Zugasti, O.C. elegans: Model host and tool for antimicrobial drug discovery. Dis. Model. Mech. 2011, 4, 300–304. [Google Scholar] [CrossRef] [PubMed]
- Hoeckendorf, A.; Stanisak, M.; Leippe, M. The saposin-like protein SPP-12 is an antimicrobial polypeptide in the pharyngeal neurons ofCaenorhabditis elegansand participates in defence against a natural bacterial pathogen. Biochem. J. 2012, 445, 205–212. [Google Scholar] [CrossRef]
- Wong, D.; Bazopoulou, D.; Pujol, N.; Tavernarakis, N.; Ewbank, J.J. Genome-wide investigation reveals pathogen-specific and shared signatures in the response of Caenorhabditis elegans to infection. Genome Biol. 2007, 8, R194. [Google Scholar] [CrossRef] [PubMed]
- Mysliwy, J.; Dingley, A.J.; Stanisak, M.; Jung, S.; Lorenzen, I.; Roeder, T.; Leippe, M.; Grötzinger, J. Caenopore-5: The three-dimensional structure of an antimicrobial protein from Caenorhabditis elegans. Dev. Comp. Immunol. 2010, 34, 323–330. [Google Scholar] [CrossRef]
- Shinnar, A.E.; Butler, K.L.; Park, H.J. Cathelicidin family of antimicrobial peptides: Proteolytic processing and protease resistance. Bioorganic Chem. 2003, 31, 425–436. [Google Scholar] [CrossRef]
- Tasiemski, A. Antimicrobial peptides in annelids. Lab. Neuroimmunol. Annelides. 2008, 75–82. [Google Scholar]
- Xu, G.; Wu, M.; Wang, L.; Zhang, X.; Cao, S.; Liu, M.; Cui, Y. Conformational and dynamics simulation study of antimicrobial peptide hedistin—Heterogeneity of its helix–turn–helix motif. Biochim. Biophys. Acta (BBA) Biomembr. 2009, 1788, 2497–2508. [Google Scholar] [CrossRef]
- Hoegenhaug, K.H.; Mygind, P.H.; Kruse, T.; Segura, D.R.; Sandvang, D.H.; Neve, S. Antimicrobial Peptide Variants and Polynucleotides Encoding Same. US Patent US8835604B2, 16 September 2014. [Google Scholar]
- Willander, H.; Hermansson, E.; Johansson, J.; Presto, J. BRICHOS domain associated with lung fibrosis, dementia and cancer—A chaperone that prevents amyloid fibril formation? FEBS J. 2011, 278, 3893–3904. [Google Scholar] [CrossRef] [PubMed]
- Sommer, A.; Pörtner, H. Metabolic cold adaptation in the lugworm Arenicola marina: Comparison of a North Sea and a White Sea population. Mar. Ecol. Prog. Ser. 2002, 240, 171–182. [Google Scholar] [CrossRef]
- Spodsberg, N. Polypeptides Having Antimicrobial Activity and Polynucleotides Encoding Same. US Patent 7745576B2, 29 June 2010. [Google Scholar]
- Desbruyères, D.; Chevaldonné, P.; Alayse, A.M.; Jollivet, D.; Lallier, F.; Jouin-Toulmond, C.; Zal, F.; Sarradin, P.M.; Cosson, R.; Caprais, J.C.; et al. Biology and ecology of the “Pompeii worm” (Alvinella pompejana Desbruyères and Laubier), a normal dweller of an extreme deep-sea environment: A synthesis of current knowledge and recent developments. Deep. Sea Res. Part II Top. Stud. Oceanogr. 1998, 45, 383–422. [Google Scholar] [CrossRef]
- Blake, J.A.; Grassle, J.P.; Eckelbarger, K.J. Capitella teleta, a new species designation for the opportunistic and experimental Capitella sp. I, with a review of the literature for confirmed records. Zoosymposia 2009, 53, 25–53. [Google Scholar]
- Cho, J.; Lee, D.G. The characteristic region of arenicin-1 involved with a bacterial membrane targeting mechanism. Biochem. Biophys. Res. Commun. 2011, 405, 422–427. [Google Scholar] [CrossRef] [PubMed]
- Ovchinnikova, T.V.; Shenkarev, Z.O.; Nadezhdin, K.D.; Balandin, S.V.; Zhmak, M.N.; Kudelina, I.A.; Finkina, E.I.; Kokryakov, V.N.; Arseniev, A.S. Recombinant expression, synthesis, purification, and solution structure of arenicin. Biochem. Biophys. Res. Commun. 2007, 360, 156–162. [Google Scholar] [CrossRef]
- Andrä, J.; Jakovkin, I.; Grötzinger, J.; Hecht, O.; Krasnosdembskaya, A.D.; Goldmann, T.; Gutsmann, T.; Leippe, M. Structure and mode of action of the antimicrobial peptide arenicin. Biochem. J. 2008, 410, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Park, K.H.; Lee, J.; Kim, J.; Shin, S.Y.; Park, Y.; Hahm, K.; Kim, Y. Cell Selectivity of Arenicin-1 and Its Derivative with Two Disulfide Bonds. Bull. Korean Chem. Soc. 2008, 29, 1190–1194. [Google Scholar] [Green Version]
- Fahrner, R.L.; Dieckmann, T.; Harwig, S.S.; Lehrer, R.I.; Eisenberg, D.; Feigon, J. Solution structure of protegrin-1, a broad-spectrum antimicrobial peptide from porcine leukocytes. Chem. Biol. 1996, 3, 543–550. [Google Scholar] [CrossRef] [Green Version]
- Laederach, A.; Andreotti, A.H.; Fulton, D.B. Solution and Micelle-Bound Structures of Tachyplesin I and Its Active Aromatic. Society 2002, 12359–12368. [Google Scholar] [CrossRef]
- Kuzmin, D.V.; Emelianova, A.A.; Kalashnikova, M.B.; Panteleev, P.V.; Balandin, S.V.; Serebrovskaya, E.O.; Belogurova-Ovchinnikova, O.Y.; Ovchinnikova, T.V. Comparative in vitro study on cytotoxicity of recombinant β-hairpin peptides. Chem. Biol. Drug Des. 2018, 91, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Edwards, I.A.; Elliott, A.G.; Kavanagh, A.M.; Zuegg, J.; Blaskovich, M.A.T.; Cooper, M.A. Contribution of Amphipathicity and Hydrophobicity to the Antimicrobial Activity and Cytotoxicity of β-Hairpin Peptides. ACS Infect. Dis. 2016, 2, 442–450. [Google Scholar] [CrossRef] [PubMed]
- Panteleev, P.V; Bolosov, I.A.; Ovchinnikova, T.V. Bioengineering and functional characterization of arenicin shortened analogs with enhanced antibacterial activity and cell selectivity. J. Pept. Sci. 2016, 22, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Shai, Y. Mode of action of membrane active antimicrobial peptides. Biopolymer 2002, 66, 236–248. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.U.; Kang, D.I.; Zhu, W.L.; Shin, S.Y.; Hahm, K.S.; Kim, Y. Solution structures and biological functions of the antimicrobial peptide, arenicin-1, and its linear derivative. Biopolymer 2007, 88, 208–216. [Google Scholar] [CrossRef]
- Panteleev, P.V.; Myshkin, M.Y.; Shenkarev, Z.O.; Ovchinnikova, T. V Biochemical and Biophysical Dimerization of the antimicrobial peptide arenicin plays a key role in the cytotoxicity but not in the antibacterial activity. Biochem. Biophys. Res. Commun. 2017, 482, 1320–1326. [Google Scholar] [CrossRef]
- Choi, H.; Lee, D.G. Synergistic effect of antimicrobial peptide arenicin-1 in combination with antibiotics against pathogenic bacteria. Res. Microbiol. 2012, 163, 479–486. [Google Scholar] [CrossRef]
- Park, C.; Lee, D.G. Fungicidal effect of antimicrobial peptide arenicin-1. Biochim. Biophys. Acta (BBA) Biomembr. 2009, 1788, 1790–1796. [Google Scholar] [CrossRef] [Green Version]
- Shenkarev, Z.O.; Balandin, S.V.; Trunov, K.I.; Paramonov, A.S.; Sukhanov, S.V.; Barsukov, L.I.; Arseniev, A.S.; Ovchinnikova, T.V. Molecular Mechanism of Action of β-Hairpin Antimicrobial Peptide Arenicin: Oligomeric Structure in Dodecylphosphocholine Micelles and Pore Formation in Planar Lipid Bilayers. Biochemistry 2011, 50, 6255–6265. [Google Scholar] [CrossRef]
- Sychev, S.V.; Sukhanov, S.V.; Panteleev, P.V.; O Shenkarev, Z.; Ovchinnikova, T.V. Marine antimicrobial peptide arenicin adopts a monomeric twisted β-hairpin structure and forms low conductivity pores in zwitterionic lipid bilayers. Pept. Sci. 2018, 110, e23093. [Google Scholar] [CrossRef]
- Yeaman, M.R.; Yount, N.Y.; Hauger, R.L.; Grigoriadis, D.E.; Dallman, M.F.; Plotsky, P.M.; Vale, W.W.; Dautzenberg, F.M. Mechanisms of Antimicrobial Peptide Action and Resistance. Pharmacol. Rev. 2003, 55, 27–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ovchinnikova, T.V.; Shenkarev, Z.O.; Balandin, S.V.; Nadezhdin, K.D.; Paramonov, A.S.; Kokryakov, V.N.; Arseniev, A.S. Molecular insight into mechanism of antimicrobial action of the β-hairpin peptide arenicin: Specific oligomerization in detergent micelles. Biopolymer 2008, 89, 455–464. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.; Lee, D.G. The antimicrobial peptide arenicin-1 promotes generation of reactive oxygen species and induction of apoptosis. Biochim. Biophys. Acta (BBA) Gen. Subj. 2011, 1810, 1246–1251. [Google Scholar] [CrossRef] [PubMed]
- Andrä, J.; Hammer, M.U.; Grötzinger, J.; Jakovkin, I.; Lindner, B.; Vollmer, E.; Fedders, H.; Leippe, M.; Gutsmann, T. Significance of the cyclic structure and of arginine residues for the antibacterial activity of arenicin-1 and its interaction with phospholipid and lipopolysaccharide model membranes. Biol. Chem. 2009, 390, 337–349. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Wang, X.; Teng, D.; Mao, R.; Hao, Y.; Feng, X. Deleting the first disulphide bond in an arenicin derivative enhances its expression in Pichia pastoris. Lett. Appl. Microbiol. 2017, 65, 241–248. [Google Scholar] [CrossRef] [PubMed]
- Yang, N.; Liu, X.; Teng, D.; Li, Z.; Wang, X.; Mao, R.; Wang, X.; Hao, Y.; Wang, J. Antibacterial and detoxifying activity of NZ17074 analogues with multi-layers of selective antimicrobial actions against Escherichia coli and Salmonella enteritidis. Sci. Rep. 2017, 7, 3392. [Google Scholar] [CrossRef] [PubMed]
- Maltseva, A.L.; Kotenko, O.N.; Kokryakov, V.N.; Starunov, V.V.; Krasnodembskaya, A.D. Expression pattern of arenicins—The antimicrobial peptides of polychaete Arenicola marina. Front. Physiol. 2014, 5, 1–11. [Google Scholar] [CrossRef]
- Panteleev, P.V.; Tsarev, A.V.; Bolosov, I.A.; Paramonov, A.S.; Marggraf, M.B.; Sychev, S.V.; Shenkarev, Z.O.; Ovchinnikova, T.V. Novel Antimicrobial Peptides from the Arctic Polychaeta Nicomache minor Provide New Molecular Insight into Biological Role of the BRICHOS Domain. Mar. Drugs 2018, 16, 401. [Google Scholar] [CrossRef]
- Shcherbakova, T.D.; Tzetlin, A.B.; Mardashova, M.V.; Sokolova, O.S. Fine structure of the tubes of Maldanidae (Annelida). J. Mar. Biol. Assoc. UK. 2017, 97, 1177–1187. [Google Scholar] [CrossRef] [Green Version]
- Kato, Y.; Aizawa, T.; Hoshino, H.; Kawano, K.; Nitta, K.; Zhang, H. abf-1 and abf-2, ASABF-type antimicrobial peptide genes in Caenorhabditis elegans. Biochem. J. 2002, 361, 221. [Google Scholar] [CrossRef]
- Kato, Y. Humoral Defense of the Nematode Ascaris suum: Antibacterial, Bacteriolytic and Agglutinating Activities in the Body Fluid. Zoöl. Sci. 1995, 12, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Tarr, D.E.K. Nematode antimicrobial peptides. Invertebr. Surviv. J. 2012, 122–133. [Google Scholar]
- Minaba, M.; Ueno, S.; Pillai, A.; Kato, Y. Evolution of ASABF (Ascaris suum antibacterial factor)-type antimicrobial peptides in nematodes: Putative rearrangement of disulfide bonding patterns. Dev. Comp. Immunol. 2009, 33, 1147–1150. [Google Scholar] [CrossRef] [PubMed]
- Pillai, A.; Ueno, S.; Zhang, H.; Kato, Y. Induction of ASABF (Ascaris suum antibacterial factor)-type antimicrobial peptides by bacterial injection: Novel members of ASABF in the nematode Ascaris suum. Biochem. J. 2003, 371, 663–668. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Yoshida, S.; Aizawa, T.; Murakami, R.; Suzuki, M.; Koganezawa, N.; Matsuura, A.; Miyazawa, M.; Kawano, K.; Nitta, K.; et al. In Vitro Antimicrobial Properties of Recombinant ASABF, an Antimicrobial Peptide Isolated from the Nematode Ascaris suum. Antimicrob. Agents Chemother. 2000, 44, 2701–2705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, S.; Dingley, A.J.; Augustin, R.; Anton-Erxleben, F.; Stanisak, M.; Gelhaus, C.; Gutsmann, T.; Hammer, M.U.; Podschun, R.; Bonvin, A.M.J.J.; et al. Hydramacin-1, structure and antibacterial activity of a protein from the basal metazoan hydra. J. Biol. Chem. 2009, 284, 1896–1905. [Google Scholar] [CrossRef] [PubMed]
- Gerdol, M.; De Moro, G.; Manfrin, C.; Venier, P.; Pallavicini, A. Big defensins and mytimacins, new AMP families of the Mediterranean mussel Mytilus galloprovincialis. Dev. Comp. Immunol. 2012, 36, 390–399. [Google Scholar] [CrossRef]
- Wilkialis, J.; Davies, R.W. The population ecology of the leech (Hirudinoidea: Glossiphoniidae) Theromyzon tessulatum. Can. J. Zoöl. 1980, 58, 906–912. [Google Scholar] [CrossRef]
- Duval, C. Hirudo medicinalis: De sa physiologie à l’hirudothérapie Thèse pour le diplôme d’état de Docteur en pharmacie. Pharm. Sci. 2013, dumas-00831872. [Google Scholar]
- Hung, C.W.; Jung, S.; Grötzinger, J.; Gelhaus, C.; Leippe, M.; Tholey, A. Determination of disulfide linkages in antimicrobial peptides of the macin family by combination of top-down and bottom-up proteomics. J. Proteom. 2014, 103, 216–226. [Google Scholar] [CrossRef]
- Tasiemski, A.; Salzet, M. Leech immunity: From brain to peripheral responses. Adv. Exp. Med. Biol. 2010, 708, 80–104. [Google Scholar] [PubMed]
- Li, C.; Nelson, L.S.; Kim, K.; Nathoo, A.; Hart, A.C. Neuropeptide Gene Families in the Nematode Caenorhabditis elegansa. Ann. NY Acad. Sci. 1999, 897, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Nathoo, A.N.; Moeller, R.A.; Westlund, B.A.; Hart, A.C. Identification of neuropeptide-like protein gene families in Caenorhabditis elegans and other species. Proc. Natl. Acad. Sci. USA 2001, 98, 14000–14005. [Google Scholar] [CrossRef] [PubMed]
- McVeigh, P.; Alexander-Bowman, S.; Veal, E.; Mousley, A.; Marks, N.J.; Maule, A.G. Neuropeptide-like protein diversity in phylum Nematoda. Int. J. Parasitol. 2008, 38, 1493–1503. [Google Scholar] [CrossRef] [PubMed]
- Lim, M.P.; Firdaus-Raih, M.; Nathan, S. Nematode Peptides with Host-Directed Anti-inflammatory Activity Rescue Caenorhabditis elegans from a Burkholderia pseudomallei Infection. Front. Microbiol. 2016, 7, 807. [Google Scholar] [CrossRef]
- Wang, X.; Wang, X.; Zhang, Y.; Qu, X.; Yang, S. An antimicrobial peptide of the earthworm Pheretima tschiliensis: cDNA cloning, expression and immunolocalization. Biotechnol. Lett. 2003, 25, 1317–1323. [Google Scholar] [CrossRef]
- Bodó, K.; Boros, Á.; Rumpler, É.; Molnár, L.; Böröcz, K.; Németh, P.; Engelmann, P. Identification of novel lumbricin homologues in Eisenia andrei earthworms. Dev. Comp. Immunol. 2019, 90, 41–46. [Google Scholar] [CrossRef]
- Li, W.; Li, S.; Zhong, J.; Zhu, Z.; Liu, J.; Wang, W. A novel antimicrobial peptide from skin secretions of the earthworm, Pheretima guillelmi (Michaelsen). Peptides 2011, 32, 1146–1150. [Google Scholar] [CrossRef]
- Bilej, M.; Procházková, P.; Šilerová, M.; Josková, R. Earthworm Immunity. In Advances in Experimental Medicine and Biology; Springer Nature: Boston, MA, USA, 2010; Volume 708, pp. 66–79. [Google Scholar]
- Gupta, S.; Yadav, S. Immuno-defense Strategy in Earthworms: A Review Article. Int. J. Curr. Microbiol. Appl. Sci. 2016, 5, 1022–1035. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Li, M.; Xi, T. Cloning and Expression of a Clamworm Antimicrobial Peptide Perinerin in Pichia pastoris. Curr. Microbiol. 2009, 58, 384–388. [Google Scholar] [CrossRef]
- Sun, F.; Zhou, Q.; Wang, M.; An, J. Joint stress of copper and petroleum hydrocarbons on the polychaete Perinereis aibuhitensis at biochemical levels. Ecotoxicol. Environ. Saf. 2009, 72, 1887–1892. [Google Scholar] [CrossRef] [PubMed]
- Smith, V.J.; Desbois, A.P.; Dyrynda, E.A. Conventional and Unconventional Antimicrobials from Fish, Marine Invertebrates and Micro-algae. Mar. Drugs 2010, 8, 1213–1262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tortorella, E.; Tedesco, P.; Esposito, F.P.; January, G.G.; Fani, R.; Jaspars, M.; De Pascale, D. Antibiotics from Deep-Sea Microorganisms: Current Discoveries and Perspectives. Mar. Drugs 2018, 16, 355. [Google Scholar] [CrossRef] [PubMed]
- Bulgheresi, S. All the microbiology nematodes can teach us. FEMS Microbiol. Ecol. 2016, 92, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Douglas, A.E. Conflict, cheats and the persistence of symbioses. New Phytol. 2008, 177, 849–858. [Google Scholar] [CrossRef] [PubMed]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Worm Phylum | Dates | AMP Families | References |
|---|---|---|---|
| Nematodes | 1989 | Cecropins | [31] |
| 1996 | ABFs | [35] | |
| 1998 | Coenopores | [38] | |
| 2002 | Caenacins | [39] | |
| 2004 | Nlps | [40] | |
| Annelids | 1998 | Lumbricins | [42] |
| 2004 | Macins | [44] | |
| 2004 and 2013 | BRICHOS-AMPs | [14,46] | |
| 2004 | Perinerin | [49] | |
| 2006 | Hedistin | [50] | |
| 2016 | Ms-Hemerycin | [51] |
| Structure (Group) | AMPs | Worm Phylum | Worm Habitat |
|---|---|---|---|
| Linear α-helix (i) | Cecropins | Nematode | Terrestrial |
| Caenopores | Nematode | Terrestrial | |
| Hedistin | Annelid | Marine | |
| β-sheet (ii) | BRICHOS-AMPs | Annelid | Marine |
| Mixed α-helix/β-sheet (iii) | ABFs | Nematode | Terrestrial |
| Macins | Annelid | Freshwater | |
| Enriched with specific amino acids (iv) | Neuropeptide-like | Nematode | Terrestrial |
| Caenacins | Nematode | Terrestrial | |
| Lumbricins | Annelid | Marine and Freshwater | |
| Derived from larger molecules (v) | Perinerin | Annelid | Marine |
| Ms-Hemerycin | Annelid | Marine |
| Microorganisms | CECROPINS | CAENOPORES | HEDISTIN | BRICHOS FAMILY | ABFS | MACINS | LUMBRICINS | PERINERIN | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| P1 | P2 | P3 | P4 | SSP1 | SSP5 | SSP12 | arenicin1 | arenicin2 | alvinellacin | nicomicin1 | AS-α | CE-2 | Hm-N | Tt-T | Hm-T | PG | 1 | ||||
| G R A M N E G A T I V E | Escherichia coli | 0.3–0.5 | 30 | 9 | 20 | 0.1 | 0.8–1.6 | 4 | 4 | 0.012–0.024 | 2–16 | 50 | 25 | 25 | 20 | 12 | 12.5–25 | ||||
| Pseudomonas aeruginosa | 0.4–0.5 | 20 | 20 | 20 | 2 | 32 | 3.1–9.2 | ||||||||||||||
| Pseudomonas sp. | 0.001–0.003 | ||||||||||||||||||||
| Salmonella enterica | 0.6 | ||||||||||||||||||||
| Salmonella typhimurium | 0.4–0.5 | 20 | 8 | 8 | |||||||||||||||||
| Proteus mirabilis | 0.6 | ||||||||||||||||||||
| Proteus vulgaris | 10 | ||||||||||||||||||||
| Klebsiella pneumoniae | 0.5 | 2–4 | 70 | 0.9 | |||||||||||||||||
| Vibrio alginolyticus | 0.4 | ||||||||||||||||||||
| Vibrio diabolicus | 0.048–0.096 | ||||||||||||||||||||
| Vibrio MPV19 | 0.012–0.024 | ||||||||||||||||||||
| Listonella anguillarum | 3.1 | ||||||||||||||||||||
| Bdellovibrio bacteriovorus | 0.5 | 0.06 | |||||||||||||||||||
| Agrobacterium tumefaciens | 5 | 10 | 0.05 | ||||||||||||||||||
| Serratia sp. | 2.5 | 16 | |||||||||||||||||||
| G R A M P O S I T I V E | Micrococcus luteus | 8 | 30 | 8 | 8 | 0.4–0.8 | 2.6 | 0.125 | 0.8 | 0.165–0.33 | 25–50 | ||||||||||
| Micrococcus nishinomiyaensis | 0.4–0.8 | 1.95–3.8 | |||||||||||||||||||
| Staphylococcus aureus | 22.2 | 8 | 3 | 3 | 3–6 | 2–8 | 0.048–0.096 | 2 | 0.6 | 6.25 | 100 | 5 | 16 | ||||||||
| Staphylococcus epidermidis | 4–8 | ||||||||||||||||||||
| Streptococcus mutans | 30 | ||||||||||||||||||||
| Bacillus megaterium | 0.1 | 0.05 | 0.275 | 2.6 | 0.012–-0.024 | 0.20 | 0.39 | 2.5-5 | |||||||||||||
| Bacillus subtilis | 2 | 20 | 10 | 20 | 0.31 | 0.062 | 1.2 | 12 | |||||||||||||
| Bacillus thuringiensis | 10 | ||||||||||||||||||||
| Kocuria varians | 0.5 | 0.008 | |||||||||||||||||||
| Enterococcus faecium | 3.4–4 | 12.5 | |||||||||||||||||||
| Enterococcus faecalis | 9.4 | ||||||||||||||||||||
| Planococcus citreus | 0.03 | ||||||||||||||||||||
| Listeria monocytogenes | 4.1 | 0.6 | 0.6–0.8 | ||||||||||||||||||
| Y E A S T S | Candida albicans | 200 | 200 | 200 | 200 | 4.5–9 | 4.5–9 | 10 | 16 | ||||||||||||
| Candida krusei | 10 | 0.3 | |||||||||||||||||||
| Candida parapsilosis | 4.5 | ||||||||||||||||||||
| Trichosporon beigelii | 4.5 | ||||||||||||||||||||
| Trichophyton rubrum | 9 | ||||||||||||||||||||
| Malassezia furfur | 9 | ||||||||||||||||||||
| Fusarium solani | 50 | ||||||||||||||||||||
| Saccharomyces cerevisiae | 300 | 300 | 300 | 300 | 12 | ||||||||||||||||
| Pichia anomala | 30 | 0.08 | |||||||||||||||||||
| Paecilomyces heliothis | 12.5–25 | ||||||||||||||||||||
| Kluyveromyces thermotolerans | 3 | 0.3 | |||||||||||||||||||
| REFERENCES | [62,67] | [62] | [62] | [62] | [72] | [72] | [75] | [50] | [14,87,98,99,100] | [88] | [48] | [110] | [35,117] | [113] | [6] | [6,44] | [43] | [130] | [42] | [49] | |
| AMP Name | Amino Acid Sequence | Hydrophobicity | Net Charge At pH 7 |
|---|---|---|---|
| Arenicin-1 | RWCVYAYVRVRGVLVRYRRCW | 42% | +6 |
| Arenicin-2 | RWCVYAYVRIRGVLVRYRRCW | 42% | +6 |
| Arenicin-3 | GFCWYVCVYRNGVRVCYRRCN | 28% | +4 |
| Alvinellacin | RGCYTRCWKVGRNGRVCMRVCT | 22% | +6 |
| Nicomicin-1 | GFWSSVWDGAKNVGTAIIKNAKVCVYAVCVSHK | 45% | +3 |
| Nicomicin-2 | GFWSSVWDGAKNVGTAIIRNAKVCVYAVCVSHK | 45% | +3 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bruno, R.; Maresca, M.; Canaan, S.; Cavalier, J.-F.; Mabrouk, K.; Boidin-Wichlacz, C.; Olleik, H.; Zeppilli, D.; Brodin, P.; Massol, F.; et al. Worms’ Antimicrobial Peptides. Mar. Drugs 2019, 17, 512. https://doi.org/10.3390/md17090512
Bruno R, Maresca M, Canaan S, Cavalier J-F, Mabrouk K, Boidin-Wichlacz C, Olleik H, Zeppilli D, Brodin P, Massol F, et al. Worms’ Antimicrobial Peptides. Marine Drugs. 2019; 17(9):512. https://doi.org/10.3390/md17090512
Chicago/Turabian StyleBruno, Renato, Marc Maresca, Stéphane Canaan, Jean-François Cavalier, Kamel Mabrouk, Céline Boidin-Wichlacz, Hamza Olleik, Daniela Zeppilli, Priscille Brodin, François Massol, and et al. 2019. "Worms’ Antimicrobial Peptides" Marine Drugs 17, no. 9: 512. https://doi.org/10.3390/md17090512
APA StyleBruno, R., Maresca, M., Canaan, S., Cavalier, J. -F., Mabrouk, K., Boidin-Wichlacz, C., Olleik, H., Zeppilli, D., Brodin, P., Massol, F., Jollivet, D., Jung, S., & Tasiemski, A. (2019). Worms’ Antimicrobial Peptides. Marine Drugs, 17(9), 512. https://doi.org/10.3390/md17090512