A New Look for the Red Macroalga Palmaria palmata: A Seafood with Polar Lipids Rich in EPA and with Antioxidant Properties


 ,
, 
 and
and
Abstract
:1. Introduction
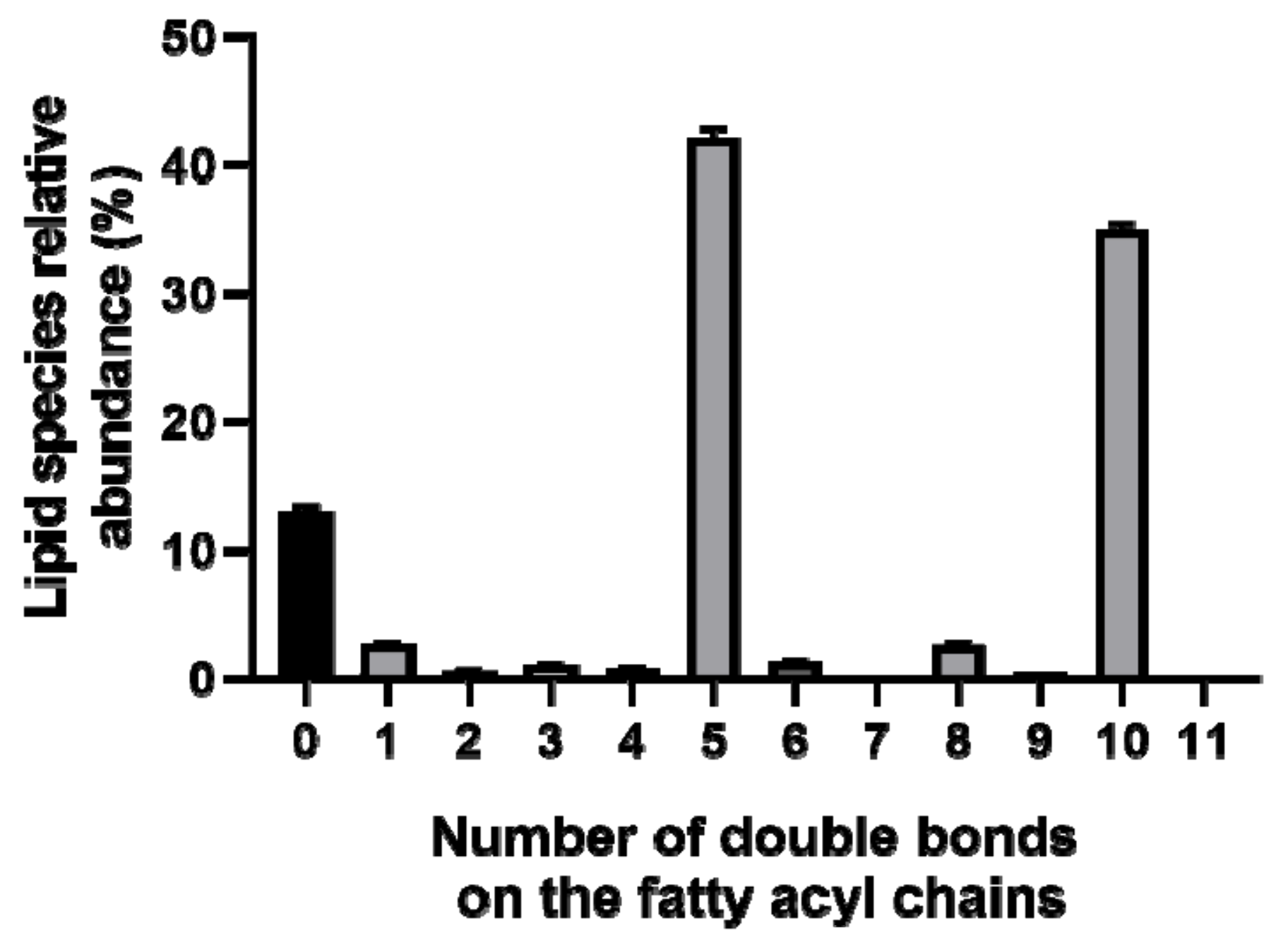
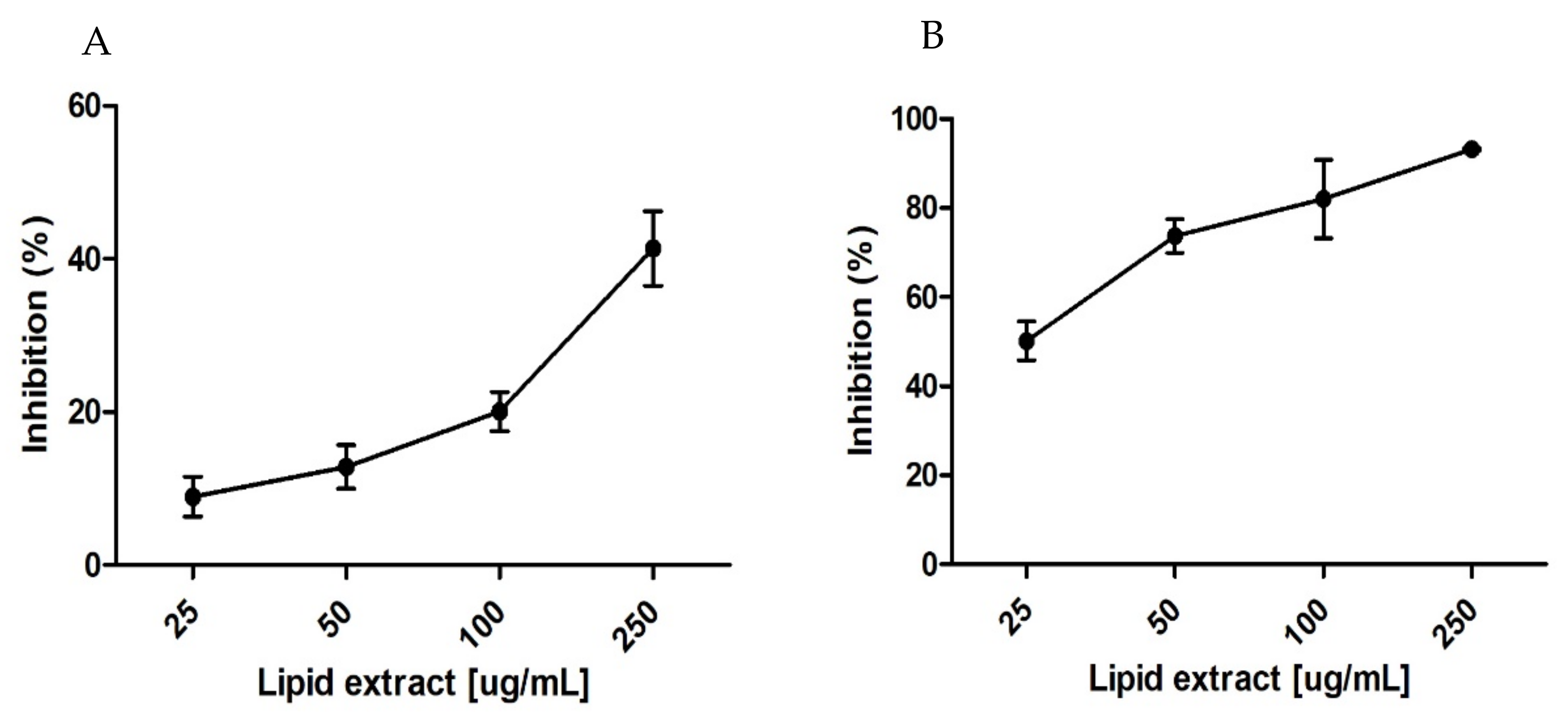
2. Results
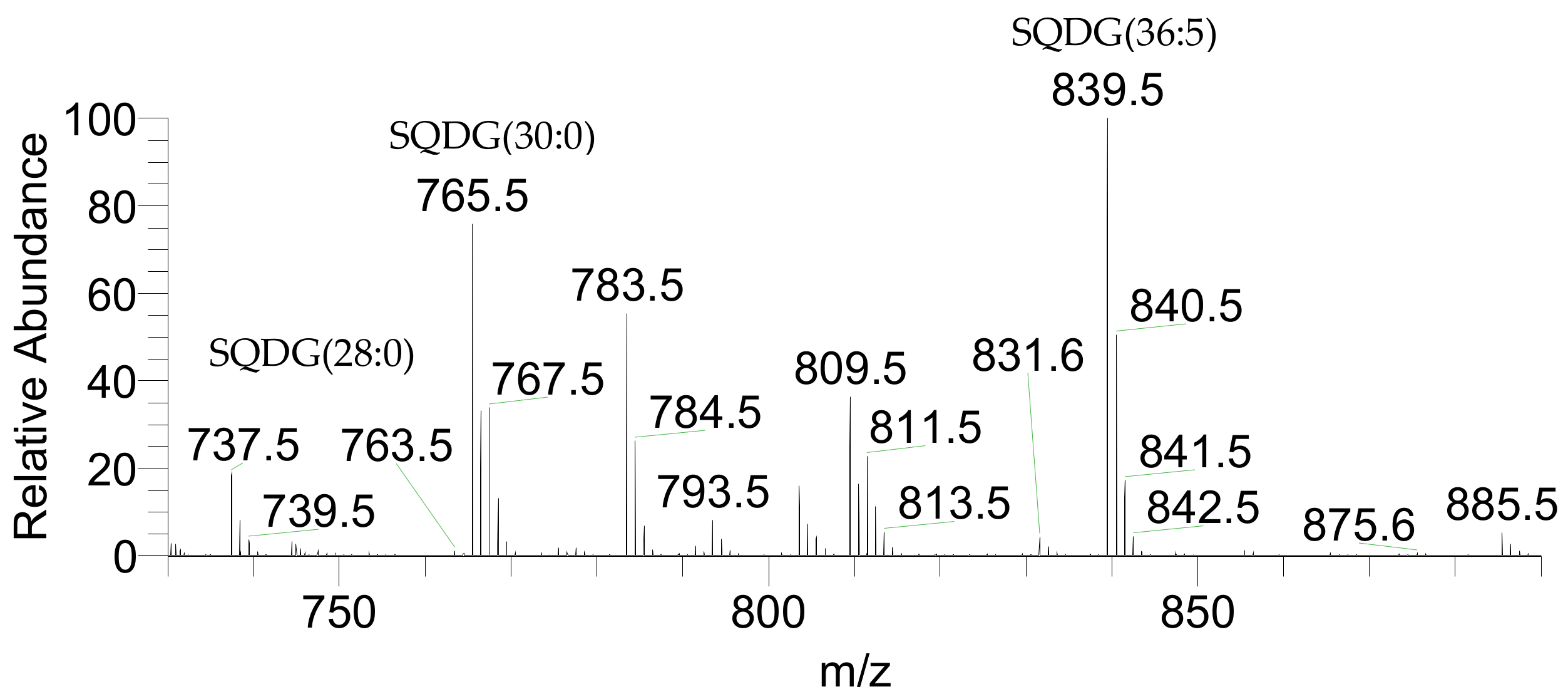
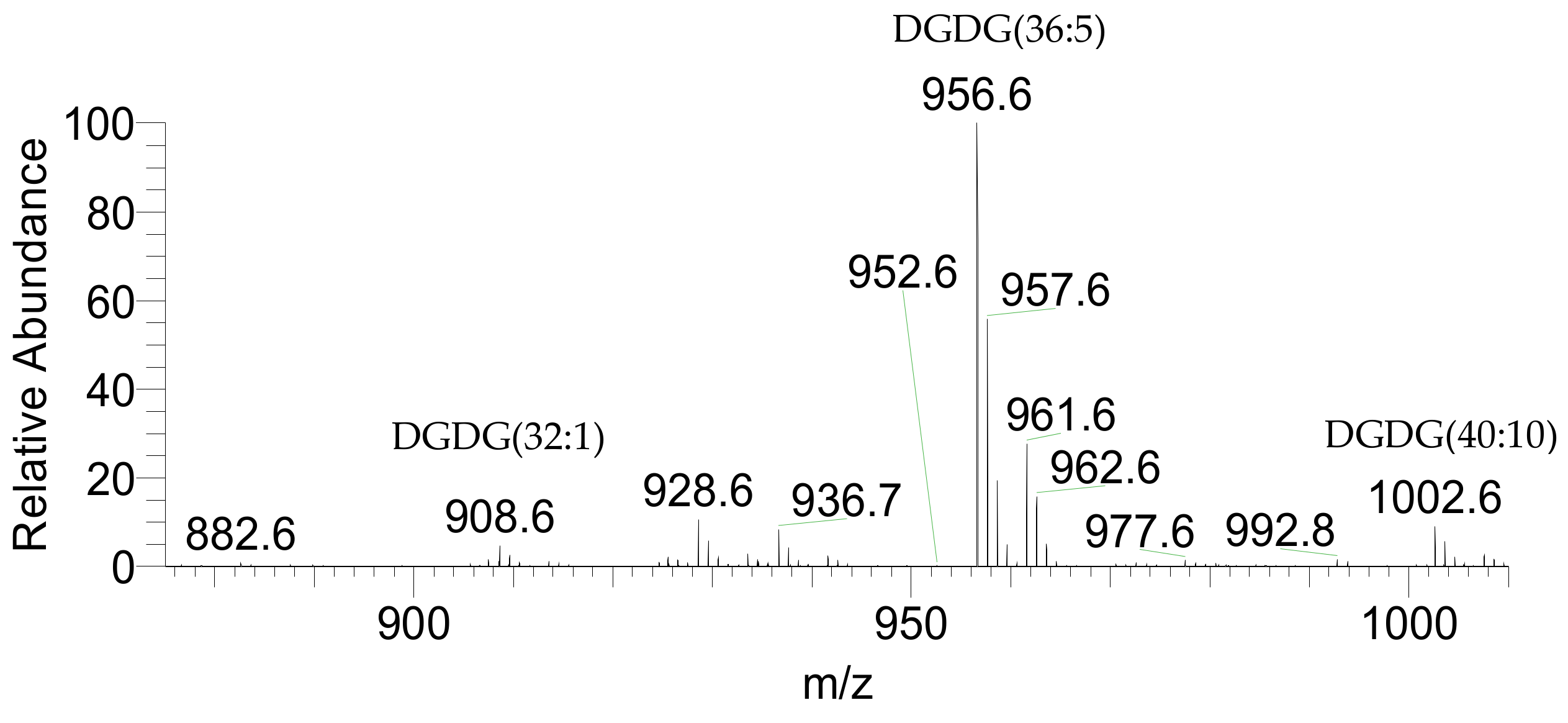
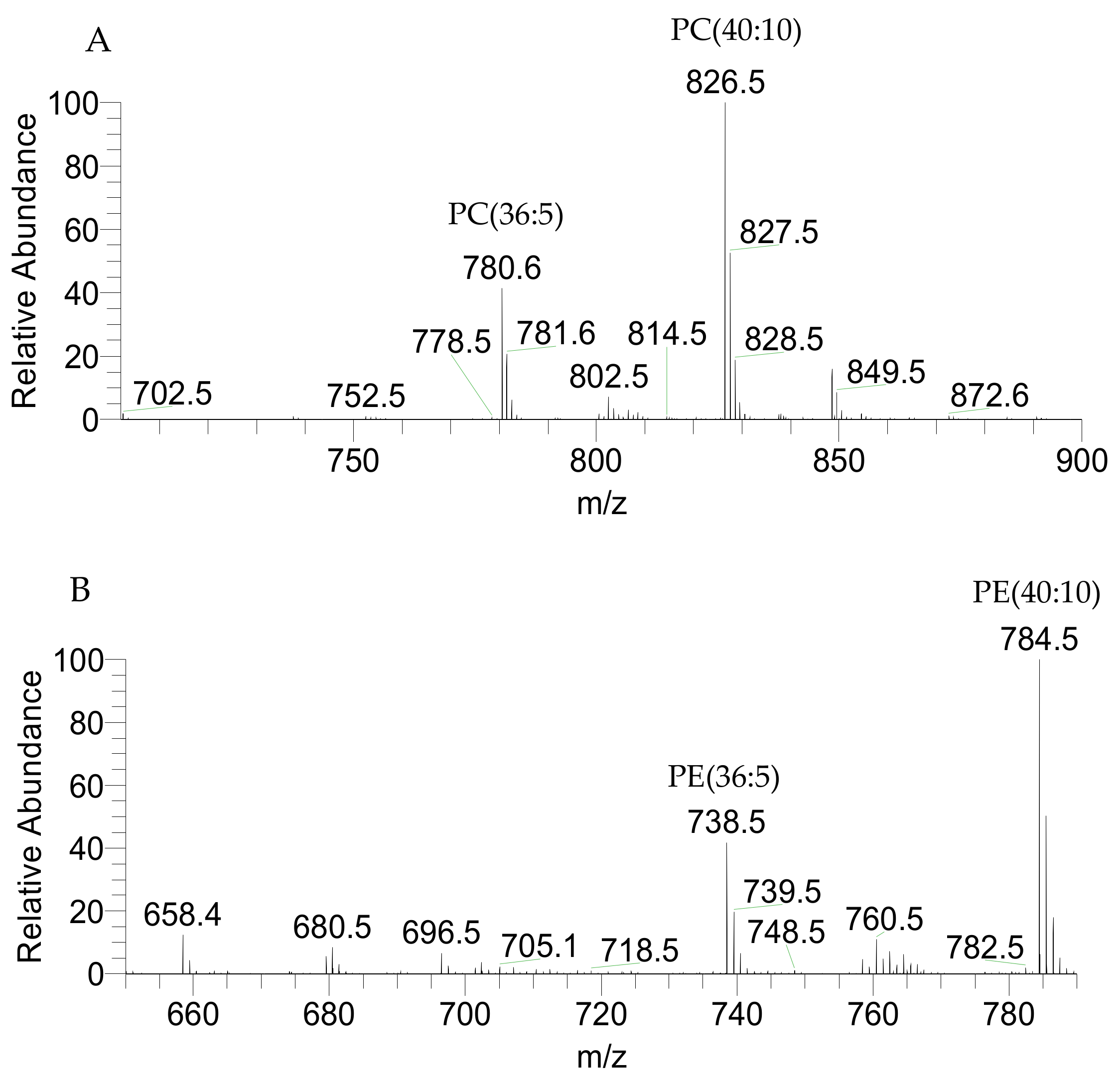
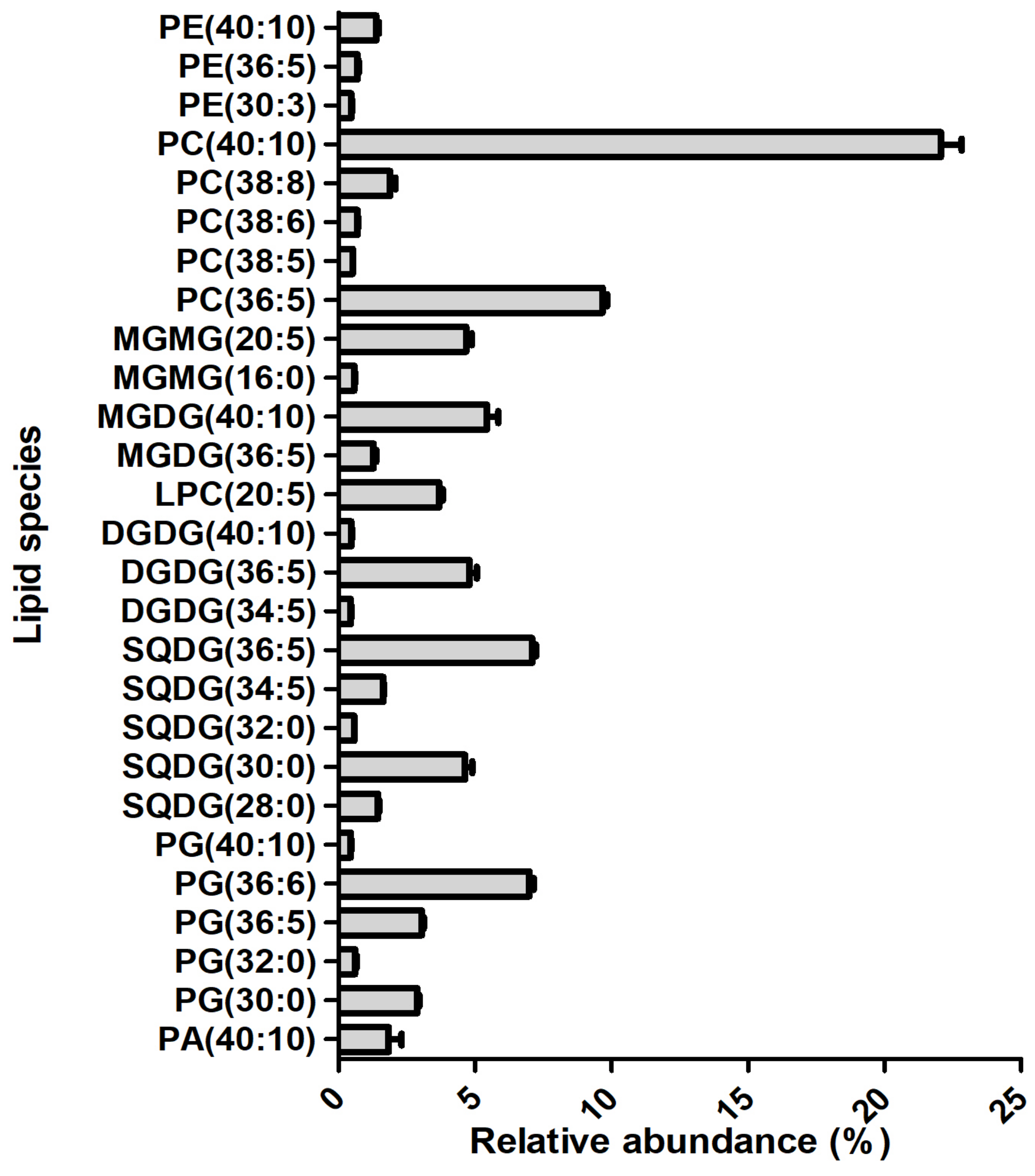
2.1. Identification of the of Palmaria Palmata Lipidome
2.2. Evaluation of Antioxidant Activity
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Seaweed Biomass
4.3. Lipid Extraction
4.4. Fatty Acid Analysis by Gas Chromatography Mass Spectrometry
4.5. Polar Lipid Analysis by Hydrophilic Interaction Liquid Chromatography Mass Spectrometry (HILIC-ESI-MS)
4.6. Data Analysis
4.7. 2-Diphenyl-1-Picrylhydrazyl Radical Assay—DPPH Radical Scavenging Activity
4.8. 2,20-Azino-bis-3-Ethylbenzothiazoline-6-Sulfonic Acid Radical Cation Assay—ABTS Radical Scavenging Activity
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- McHugh, D.J. A Guide to the Seaweed Industry; FAO Fisheries Technical Paper; FAO: Rome, Italy, 2003; p. 105. ISBN 9-25-104958-0. [Google Scholar]
- Bixler, H.J.; Porse, H. A decade of change in the seaweed hydrocolloids industry. J. Appl. Phycol. 2011, 23, 321–335. [Google Scholar] [CrossRef]
- Sutherland, J.E.; Lindstrom, S.C.; Nelson, W.A.; Brodie, J.; Lynch, M.D.J.; Hwang, M.S.; Choi, H.; Miyata, M.; Kikuchi, N.; Oliveira, M.C.; et al. A new look at an ancient order: Generic revision of the Bangiales (Rhodophyta). J. Phycol. 2011, 47, 1131–1151. [Google Scholar] [CrossRef] [PubMed]
- Cultured Aquatic Species Information Programme. Porphyra spp. Cultured Aquatic Species Information Programme; Chen, J., Xu, P., Eds.; FAO Fisheries and Aquaculture Department: Rome, Italy, 2005. [Google Scholar]
- Grote, B. Recent developments in aquaculture of Palmaria palmata (Linnaeus) (Weber & Mohr 1805): Cultivation and uses. Rev. Aquac. 2019, 11, 25–41. [Google Scholar]
- Morgan, K.C.; Wright, J.L.C.; Simpson, F.J. Review of chemical constituents of the red alga Palmaria palmata (dulse). Econ. Bot. 1980, 34, 28–50. [Google Scholar] [CrossRef]
- Mishra, V.K.; Temelli, F.; Ooraikul, B.; Shacklock, P.F.; Craigie, J.S. Lipids of the Red Alga, Palmaria palmata. Bot. Mar. 1993, 36, 169–174. [Google Scholar] [CrossRef]
- Van Ginneken, V.J.; Helsper, J.P.; de Visser, W.; van Keulen, H.; Brandenburg, W.A. Polyunsaturated fatty acids in various macroalgal species from north Atlantic and tropical seas. Lipids Health Dis. 2011, 10, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Borow, K.M.; Nelson, J.R.; Mason, R.P. Biologic plausibility, cellular effects, and molecular mechanisms of eicosapentaenoic acid (EPA) in atherosclerosis. Atherosclerosis 2015, 242, 357–366. [Google Scholar] [CrossRef] [Green Version]
- Gdula-Argasińska, J.; Czepiel, J.; Woźniakiewicz, A.; Wojtoń, K.; Grzywacz, A.; Woźniakiewicz, M.; Jurczyszyn, A.; Perucki, W.; Librowski, T. N-3 Fatty acids as resolvents of inflammation in the A549 cells. Pharmacol. Rep. 2015, 67, 610–615. [Google Scholar] [CrossRef]
- Meyer, B.J. Are we consuming enough long chain omega-3 polyunsaturated fatty acids for optimal health? Prostaglandins Leukot. Essent. Fat. Acids 2011, 85, 275–280. [Google Scholar] [CrossRef] [Green Version]
- Eitsuka, T.; Nakagawa, K.; Igarashi, M.; Miyazawa, T. Telomerase inhibition by sulfoquinovosyldiacylglycerol from edible purple laver (Porphyra yezoensis). Cancer Lett. 2004, 212, 15–20. [Google Scholar] [CrossRef]
- Ohta, K.; Mizushina, Y.; Hirata, N.; Takemura, M.; Sugawara, F.; Matsukage, A.; Yoshida, S.; Sakaguchi, K. Sulfoquinovosyldiacylglycerol, KM043, a new potent inhibitor of eukaryotic DNA polymerases and HIV-reverse transcriptase type 1 from a marine red alga, Gigartina tenella. Chem. Pharm. Bull. (Tokyo) 1998, 46, 684–686. [Google Scholar] [CrossRef] [PubMed]
- Banskota, A.H.; Stefanova, R.; Sperker, S.; Lall, S.P.; Craigie, J.S.; Hafting, J.T.; Critchley, A.T. Polar lipids from the marine macroalga Palmaria palmata inhibit lipopolysaccharide-induced nitric oxide production in RAW264.7 macrophage cells. Phytochemistry 2014, 101, 101–108. [Google Scholar] [CrossRef] [PubMed]
- Banskota, A.H.; Stefanova, R.; Sperker, S.; Melanson, R.; Osborne, J.A.; O’Leary, S.J.B. Five new galactolipids from the freshwater microalga Porphyridium aerugineum and their nitric oxide inhibitory activity. J. Appl. Phycol. 2013, 25, 951–960. [Google Scholar] [CrossRef]
- El Baz, F.K.; El Baroty, G.S.; Abd El Baky, H.H.; Abd El Salam, O.I.; Ibrahim, E.A. Structural characterization and Biological Activity of Sulfolipids from selected Marine Algae. Grasas y Aceites 2013, 64, 561–571. [Google Scholar]
- Parveez, A.A.; Rasheed, U.M.; Noorani, K.P.M.; Reehana, N.; Santhoshkumar, S.; Imran, Y.M.M.; Alharbi, S.N.; Arunachalam, C.; Alharbi, A.S.; Akbarsha, M.A.; et al. In vitro antibacterial activity of MGDG-palmitoyl from Oscillatoria acuminata NTAPC05 against extended-spectrum β-lactamase producers. J. Antibiot. (Tokyo) 2017, 70, 754–762. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Li, Y.L.; Shen, W.Z.; Rui, W.; Ma, X.J.; Cen, Y.Z. Antiviral activity of a sulfoquinovosyldiacylglycerol (SQDG) compound isolated from the green alga Caulerpa racemosa. Bot. Mar. 2007, 50, 185–190. [Google Scholar] [CrossRef]
- Terme, N.; Boulho, R.; Kucma, J.-P.; Bourgougnon, N.; Bedoux, G. Radical scavenging activity of lipids from seaweeds isolated by solid-liquid extraction and supercritical fluids. OCL 2018, 25, D505. [Google Scholar] [CrossRef]
- Chakraborty, K.; Maneesh, A.; Makkar, F. Antioxidant Activity of Brown Seaweeds. J. Aquat. Food Prod. Technol. 2017, 26, 406–419. [Google Scholar] [CrossRef]
- Da Costa, E.; Melo, T.; Moreira, A.S.P.; Alves, E.; Domingues, P.; Calado, R.; Abreu, M.H.; Domingues, M.R. Decoding bioactive polar lipid profile of the macroalgae Codium tomentosum from a sustainable IMTA system using a lipidomic approach. Algal Res. 2015, 12, 388–397. [Google Scholar] [CrossRef]
- Da Costa, E.; Azevedo, V.; Melo, T.; Rego, A.M.; Evtuguin, D.V.; Domingues, P.; Calado, R.; Pereira, R.; Abreu, M.H.; Domingues, M.R. High-Resolution Lipidomics of the Early Life Stages of the Red Seaweed Porphyra dioica. Molecules 2018, 23, 187. [Google Scholar] [CrossRef]
- Da Costa, E.; Melo, T.; Moreira, A.S.P.; Bernardo, C.; Helguero, L.; Ferreira, I.; Cruz, M.T.; Rego, A.M.; Domingues, P.; Calado, R.; et al. Valorization of Lipids from Gracilaria sp. through Lipidomics and Decoding of Antiproliferative and Anti-Inflammatory Activity. Mar. Drugs 2017, 15, 62. [Google Scholar] [CrossRef] [PubMed]
- Melo, T.; Alves, E.; Azevedo, V.; Martins, A.S.; Neves, B.; Domingues, P.; Calado, R.; Abreu, H.; Domingues, M.R. Lipidomics as a new approach for the bioprospecting of marine macroalgae - unraveling the polar lipid and fatty acid composition of Chondrus crispus. Algal Res. 2015, 8, 181–191. [Google Scholar] [CrossRef]
- Lopes, D.; Moreira, A.S.P.; Rey, F.; da Costa, E.; Melo, T.; Maciel, E.; Rego, A.; Abreu, M.H.; Domingues, P.; Calado, R.; et al. Lipidomic signature of the green macroalgae Ulva rigida farmed in a sustainable integrated multi-trophic aquaculture. J. Appl. Phycol. 2019, 31, 1369–1381. [Google Scholar] [CrossRef]
- Byju, K.; Vasundhara, G.; Anuradha, V.; Nair, S.M.; Kumar, N.C. Presence of Phytol, a Precursor of Vitamin E in Chaetomorpha Antinnina. Mapana-J. Sci. 2013, 12, 57–65. [Google Scholar] [CrossRef]
- Guschina, I.A.; Harwood, J.L. Lipids and lipid metabolism in eukaryotic algae. Prog. Lipid Res. 2006, 45, 160–186. [Google Scholar] [CrossRef] [PubMed]
- Li-Beisson, Y.; Thelen, J.J.; Fedosejevs, E.; Harwood, J.L. The lipid biochemistry of eukaryotic algae. Prog. Lipid Res. 2019, 74, 31–68. [Google Scholar] [CrossRef] [PubMed]
- Petroutsos, D.; Amiar, S.; Abida, H.; Dolch, L.J.; Bastien, O.; Rébeillé, F.; Jouhet, J.; Falconet, D.; Block, M.A.; McFadden, G.I.; et al. Evolution of galactoglycerolipid biosynthetic pathways—From cyanobacteria to primary plastids and from primary to secondary plastids. Prog. Lipid Res. 2014, 54, 68–85. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.-L.; Wang, B.-G. Antioxidant capacity and lipophilic content of seaweeds collected from the Qingdao coastline. J. Agric. Food Chem. 2004, 52, 4993–4997. [Google Scholar] [CrossRef]
- Mozaffarian, D.; Ascherio, A.; Hu, F.B.; Stampfer, M.J.; Willett, W.C.; Siscovick, D.S.; Rimm, E.B. Interplay between different polyunsaturated fatty acids and risk of coronary heart disease in men. Circulation 2005, 111, 157–164. [Google Scholar] [CrossRef]
- Marik, P.E.; Varon, J. Omega-3 dietary supplements and the risk of cardiovascular events: A systematic review. Clin. Cardiol. 2009, 32, 365–372. [Google Scholar] [CrossRef]
- SanGiovanni, J.P.; Parra-Cabrera, S.; Colditz, G.A.; Berkey, C.S.; Dwyer, J.T. Meta-analysis of Dietary Essential Fatty Acids and Long-Chain Polyunsaturated Fatty Acids as They Relate to Visual Resolution Acuity in Healthy Preterm Infants. Pediatrics 2004, 105, 1292–1298. [Google Scholar] [CrossRef] [PubMed]
- Rennie, K.L.; Hughes, J.; Lang, R.; Jebb, S.A. Nutritional management of rheumatoid arthritis: A review of the evidence. J. Hum. Nutr. Diet. 2003, 16, 97–109. [Google Scholar] [CrossRef] [PubMed]
- Barros, R.; Moreira, A.; Fonseca, J.; Delgado, L.; Graça Castel-Branco, M.; Haahtela, T.; Lopes, C.; Moreira, P. Dietary intake of α-linolenic acid and low ratio of n -6: N -3 PUFA are associated with decreased exhaled NO and improved asthma control. Br. J. Nutr. 2011, 106, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Chopin, T.; Cooper, J.A.; Reid, G.; Cross, S.; Moore, C. Open-water integrated multi-trophic aquaculture: Environmental biomitigation and economic diversification of fed aquaculture by extractive aquaculture. Rev. Aquac. 2012, 4, 209–220. [Google Scholar] [CrossRef]
- Paiva, L.; Lima, E.; Neto, A.I.; Baptista, J. Seasonal Variability of the Biochemical Composition and Antioxidant Properties of Fucus spiralis at Two Azorean Islands. Mar. Drugs 2018, 16, 248. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Machado, D.I.; López-Cervantes, J.; López-Hernández, J.; Paseiro-Losada, P. Fatty acids, total lipid, protein and ash contents of processed edible seaweeds. Food Chem. 2004, 85, 439–444. [Google Scholar] [CrossRef]
- Lagarde, M.; Bernoud, N.; Brossard, N.; Lemaitre-Delaunay, D.; Thiès, F.; Croset, M.; Lecerf, J. Lysophosphatidylcholine as a preferred carrier form of docosahexaenoic acid to the brain. J. Mol. Neurosci. 2001, 16, 201–204. [Google Scholar] [CrossRef]
- Picq, M.; Chen, P.; Perez, M.; Michaud, M.; Véricel, E.; Guichardant, M.; Lagarde, M. DHA Metabolism: Targeting the Brain and Lipoxygenation. Mol. Neurobiol. 2010, 42, 48–51. [Google Scholar] [CrossRef] [Green Version]
- Krimbou, L.; Hajj Hassan, H.; Blain, S.; Rashid, S.; Denis, M.; Marcil, M.; Genest, J. Biogenesis and speciation of nascent apoA-I-containing particles in various cell lines. J. Lipid Res. 2005, 46, 1668–1677. [Google Scholar] [CrossRef] [Green Version]
- Magun, A.M.; Mish, B.; Glickman, R.M. Intracellular apoA-I and apoB distribution in rat intestine is altered by lipid feeding. J. Lipid Res. 1988, 29, 1107–1116. [Google Scholar]
- Wang, H.; Du, J.; Lu, S.; Yao, Y.; Hunter, F.; Black, D.D. Regulation of intestinal apolipoprotein A-I synthesis by dietary phosphatidylcholine in newborn swine. Lipids 2001, 36, 683–687. [Google Scholar] [CrossRef] [PubMed]
- Hung, M.-C.; Shibasaki, K.; Yoshida, R.; Sato, M.; Imaizumi, K. Learning behaviour and cerebral protein kinase C, antioxidant status, lipid composition in senescence-accelerated mouse: Influence of a phosphatidylcholine–vitamin B12 diet. Br. J. Nutr. 2001, 86, 163–171. [Google Scholar] [CrossRef] [PubMed]
- Schneider, H.; Braun, A.; Füllekrug, J.; Stremmel, W.; Ehehalt, R. Lipid Based Therapy for Ulcerative Colitis—Modulation of Intestinal Mucus Membrane Phospholipids as a Tool to Influence Inflammation. Int. J. Mol. Sci. 2010, 11, 4149–4164. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.-Y.; Moriyama, T.; Uezu, E.; Uezu, K.; Hirata, R.; Yohena, N.; Masuda, Y.; Kokubu, Y.; Yamamoto, S. Administration of Phosphatidylcholine Increases Brain Acetylcholine Concentration and Improves Memory in Mice with Dementia. J. Nutr. 1995, 125, 1484–1489. [Google Scholar] [PubMed]
- Buang, Y.; Wang, Y.-M.; Cha, J.-Y.; Nagao, K.; Yanagita, T. Dietary phosphatidylcholine alleviates fatty liver induced by orotic acid. Nutrition 2005, 21, 867–873. [Google Scholar] [CrossRef] [PubMed]
- Deiana, S.; Platt, B.; Riedel, G. The cholinergic system and spatial learning. Behav. Brain Res. 2011, 221, 389–411. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Gammon, M.D.; Zeisel, S.H.; Bradshaw, P.T.; Wetmur, J.G.; Teitelbaum, S.L.; Neugut, A.I.; Santella, R.M.; Chen, J. High intakes of choline and betaine reduce breast cancer mortality in a population-based study. FASEB J. 2009, 23, 4022–4028. [Google Scholar] [CrossRef]
- Xu, X.; Gammon, M.D.; Zeisel, S.H.; Lee, Y.L.; Wetmur, J.G.; Teitelbaum, S.L.; Bradshaw, P.T.; Neugut, A.I.; Santella, R.M.; Chen, J. Choline metabolism and risk of breast cancer in a population-based study. FASEB J. 2008, 22, 2045–2052. [Google Scholar] [CrossRef]
- Lee, J.E.; Giovannucci, E.; Fuchs, C.S.; Willett, W.C.; Zeisel, S.H.; Cho, E. Choline and betaine intake and the risk of colorectal cancer in men. Cancer Epidemiol. Biomark. Prev. 2010, 19, 884–887. [Google Scholar] [CrossRef]
- Burri, L.; Hoem, N.; Banni, S.; Berge, K. Marine Omega-3 phospholipids: Metabolism and biological activities. Int. J. Mol. Sci. 2012, 13, 15401–15419. [Google Scholar] [CrossRef]
- Alavi, M.; Karimi, N.; Safaei, M. Application of Various Types of Liposomes in Drug Delivery Systems. Adv. Pharm. Bull. 2017, 7, 3–9. [Google Scholar] [CrossRef] [PubMed]
- Silva AMS, A.A. Chemical Study and Biological Activity Evaluation of Two Azorean Macroalgae: Ulva rigida and Gelidium microdon. Oceanogr. Open Access 2013, 1, 1–7. [Google Scholar] [CrossRef]
- Lee, J.-C.; Hou, M.-F.; Huang, H.-W.; Chang, F.-R.; Yeh, C.-C.; Tang, J.-Y.; Chang, H.-W. Marine algal natural products with anti-oxidative, anti-inflammatory, and anti-cancer properties. Cancer Cell Int. 2013, 13, 55. [Google Scholar] [CrossRef] [PubMed]
- Mendes, M.; Pereira, R.; Sousa Pinto, I.; Carvalho, A.P.; Gomes, A.M. Antimicrobial activity and lipid profile of seaweed extracts from the North Portuguese Coast. Int. Food Res. J. 2013, 20, 3337–3345. [Google Scholar]
- Arunkumar, K.; Selvapalam, N.; Rengasamy, R. The antibacterial compound sulphoglycerolipid 1-0 palmitoyl-3-0(6-sulpho-α-quinovopyranosyl)-glycerol from Sargassum wightii Greville (Phaeophyceae). Bot. Mar. 2005, 48, 441–445. [Google Scholar] [CrossRef]
- Ohta, K.; Mizushima, Y.; Hirata, N.; Takemura, M.; Sugawara, F.; Matsukage, A.; Yoshida, S.; Sakaguchi, K. Action of a New Mammalian DNA Polymerase Inhibitor, Sulfoquinovosyldiacylglycerol. Biol. Pharm. Bull. 1999, 22, 111–116. [Google Scholar] [CrossRef]
- Winget, R.R. Anti-Inflammatory Compositions Containing Monogalactosyl Dieicosapentaenoyl Glycerol and Methods Relating Thereto. U.S. Patent No. 5,767,095, 16 June 1998. [Google Scholar]
- Khansari, N.; Shakiba, Y.; Mahmoudi, M. Chronic inflammation and oxidative stress as a major cause of age-related diseases and cancer. Recent Pat. Inflamm. Allergy Drug Discov. 2009, 3, 73–80. [Google Scholar] [CrossRef]
- Liguori, I.; Russo, G.; Curcio, F.; Bulli, G.; Aran, L.; Della-Morte, D.; Gargiulo, G.; Testa, G.; Cacciatore, F.; Bonaduce, D.; et al. Oxidative stress, aging, and diseases. Clin. Interv. Aging 2018, 13, 757–772. [Google Scholar] [CrossRef]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free radicals, antioxidants and functional foods: Impact on human health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef]
- Arulselvan, P.; Fard, M.T.; Tan, W.S.; Gothai, S.; Fakurazi, S.; Norhaizan, M.E.; Kumar, S.S. Role of Antioxidants and Natural Products in Inflammation. Oxid. Med. Cell. Longev. 2016, 2016, 5276130. [Google Scholar] [CrossRef]
- Xu, D.-P.; Li, Y.; Meng, X.; Zhou, T.; Zhou, Y.; Zheng, J.; Zhang, J.-J.; Li, H.-B. Natural Antioxidants in Foods and Medicinal Plants: Extraction, Assessment and Resources. Int. J. Mol. Sci. 2017, 18, 96. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, C.; Sørensen, A.-D.M.; Holdt, S.L.; Akoh, C.C.; Hermund, D.B. Source, Extraction, Characterization, and Applications of Novel Antioxidants from Seaweed. Annu. Rev. Food Sci. Technol. 2019, 10, 541–568. [Google Scholar] [CrossRef] [PubMed]
- Rey, F.; Lopes, D.; Maciel, E.; Monteiro, J.; Skjermo, J.; Funderud, J.; Raposo, D.; Domingues, P.; Calado, R.; Domingues, M.R. Polar lipid profile of Saccharina latissima, a functional food from the sea. Algal Res. 2019, 39, 101473. [Google Scholar] [CrossRef]
- Da Costa, E.; Domingues, P.; Melo, T.; Coelho, E.; Pereira, R.; Calado, R.; Abreu, H.M.; Domingues, R.M. Lipidomic Signatures Reveal Seasonal Shifts on the Relative Abundance of High-Valued Lipids from the Brown Algae Fucus vesiculosus. Mar. Drugs 2019, 17, 335. [Google Scholar] [CrossRef] [PubMed]
- Rico, D.; Diana, A.B.M.; Milton-Laskibar, I.; Fernández-Quintela, A.; Silván, J.M.; Rai, D.K.; Choudhary, A.; Peñas, E.; de Luis, D.A.; Martínez-Villaluenga, C. Characterization and in vitro evaluation of seaweed species as potential functional ingredients to ameliorate metabolic syndrome. J. Funct. Foods 2018, 46, 185–194. [Google Scholar] [CrossRef] [Green Version]
- Leri, A.C.; Dunigan, M.R.; Wenrich, R.L.; Ravel, B. Particulate organohalogens in edible brown seaweeds. Food Chem. 2019, 272, 126–132. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.; Alves, C.; Freitas, R.; Martins, A.; Pinteus, S.; Ribeiro, J.; Gaspar, H.; Alfonso, A.; Pedrosa, R. Antioxidant and neuroprotective potential of the brown seaweed Bifurcaria bifurcata in an in vitro Parkinson’s disease model. Mar. Drugs 2019, 17, 85. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Zhang, J.; Fan, J.; Clark, J.; Shen, P.; Li, Y.; Zhang, C. Microwave assisted extraction of phenolic compounds from four economic brown macroalgae species and evaluation of their antioxidant activities and inhibitory effects on α-amylase, α-glucosidase, pancreatic lipase and tyrosinase. Food Res. Int. 2018, 113, 288–297. [Google Scholar] [CrossRef]
- Yuan, Y.V.; Carrington, M.F.; Walsh, N.A. Extracts from dulse (Palmaria palmata) are effective antioxidants and inhibitors of cell proliferation in vitro. Food Chem. Toxicol. 2005, 43, 1073–1081. [Google Scholar] [CrossRef]
- Pang, J.R.; Goh, V.M.J.; Tan, C.Y.; Phang, S.M.; Wong, K.H.; Yow, Y.Y. Neuritogenic and in vitro antioxidant activities of Malaysian Gracilaria manilaensis Yamamoto & Trono. J. Appl. Phycol. 2018, 30, 3253–3260. [Google Scholar]
- Lim, S.; Choi, A.H.; Kwon, M.; Joung, E.J.; Shin, T.; Lee, S.G.; Kim, N.G.; Kim, H.R. Evaluation of antioxidant activities of various solvent extract from Sargassum serratifolium and its major antioxidant components. Food Chem. 2019, 278, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Melo, T.; Marques, S.S.; Ferreira, I.; Cruz, M.T.; Domingues, P.; Segundo, M.A.; Domingues, M.R.M. New Insights into the Anti-Inflammatory and Antioxidant Properties of Nitrated Phospholipids. Lipids 2018, 53, 117–131. [Google Scholar] [CrossRef] [PubMed]
- Magalhães, L.M.; Segundo, M.A.; Reis, S.; Lima, J.L.F.C. Automatic method for determination of total antioxidant capacity using 2,2-diphenyl-1-picrylhydrazyl assay. Anal. Chim. Acta 2006, 558, 310–318. [Google Scholar] [CrossRef]
- Magalhães, L.M.; Barreiros, L.; Maia, M.A.; Reis, S.; Segundo, M.A. Rapid assessment of endpoint antioxidant capacity of red wines through microchemical methods using a kinetic matching approach. Talanta 2012, 97, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Ozgen, M.; Reese, R.N.; Tulio, A.Z.; Scheerens, J.C.; Miller, A.R. Modified 2,2-azino-bis-3-ethylbenzothiazoline-6-sulfonic acid (ABTS) method to measure antioxidant capacity of selected small fruits and comparison to ferric reducing antioxidant power (FRAP) and 2,2′-diphenyl-1- picrylhydrazyl (DPPH) methods. J. Agric. Food Chem. 2006, 54, 1151–1157. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid | Relative Abundance (%) ± SD |
|---|---|
| 14:0 | 5.32 ± 0.44 |
| 16:0 | 24.32 ± 1.11 |
| 16:1(n-7) | 2.03 ± 0.43 |
| 18:0 | 12.45 ± 6.74 |
| 18:1 | 2.82 ± 0.54 |
| 18:2 | 0.45 ± 0.19 |
| 20:4(n-6) | 0.92 ± 0.17 |
| 20:5(n-3) | 51.68 ± 6.47 |
| Theoretical m/z | Observed m/z | Error (ppm) | Lipid Species (C:N) | Fatty Acyl Chains | Formula |
|---|---|---|---|---|---|
| 527.2526 | 527.2541 | 2.84 | SQMG(14:0) | 14:0 | C23H43O11S |
| 555.2839 | 555.2855 | 2.88 | SQMG(16:0) | 16:0 | C25H47O11S |
| 737.4510 | 737.4529 | 2.58 | SQDG(28:0) | 14:0/14:0 | C37H69O12S |
| 763.4666 | 763.4685 | 2.49 | SQDG(30:1) | 14:0/16:1 | C39H71O12S |
| 765.4823 | 765.4797 | −3.40 | SQDG(30:0) | 14:0/16:0 | C39H73O12S |
| 785.4510 | 785.4531 | 2.67 | SQDG(32:4) | 16:3/16:1 | C41H69O12S |
| 787.4666 | 787.4630 | −4.57 | SQDG(32:3) | 16:3/16:0 | C41H71O12S |
| 789.4823 | 789.4861 | 4.81 | SQDG(32:2) | 18:2/14:0 | C41H73O12S |
| 791.4979 | 791.5001 | 2.78 | SQDG(32:1) | 18:1/14:0 | C41H75O12S |
| 793.5136 | 793.5156 | 2.52 | SQDG(32:0) | 16:0/16:0 | C41H77O12S |
| 811.4666 | 811.4685 | 2.34 | SQDG(34:5) | 20:5/14:0 | C43H71O12S |
| 819.5292 | 819.5308 | 1.95 | SQDG(34:1) | 20:1/14:0 | C43H79O12S |
| 821.5449 | 821.5461 | 1.46 | SQDG(34:0) | * | C43H81O12S |
| 837.4823 | 837.4834 | 1.31 | SQDG(36:6) | 20:5/16:1 | C45H73O12S |
| 839.4979 | 839.4998 | 2.26 | SQDG(36:5) | 20:5/16:0 | C45H75O12S |
| 847.5605 | 847.5618 | 1.53 | SQDG(36:1) | 20:1/16:0 | C45H83O12S |
| 859.4666 | 859.4682 | 1.86 | SQDG(38:9) | 22:5/16:4 and 20:5/18:4 | C47H71O12S |
| 875.5918 | 875.5939 | 2.40 | SQDG(38:1) | 24:1/14:0 | C47H87O12S |
| 885.4823 | 885.4843 | 2.26 | SQDG(40:10) | 20:5/20:5 | C49H73O12S |
| Theoretical m/z | Observed m/z | Error (ppm) | Lipid Species (C:N) | Fatty Acyl Chains | Formula |
|---|---|---|---|---|---|
| 482.3329 | 482.3326 | −0.62 | MGMG(14:0) | 14:0 | C23H48NO9 |
| 502.3016 | 502.3014 | −0.40 | MGMG(16:4) | 16:4 | C25H44NO9 |
| 504.3173 | 504.3184 | 2.18 | MGMG(16:3) | 16:3 | C25H46NO9 |
| 506.3329 | 506.3331 | 0.39 | MGMG(16:2) | 16:2 | C25H48NO9 |
| 510.3642 | 510.3640 | −0.39 | MGMG(16:0) | 16:0 | C25H52NO9 |
| 530.3329 | 530.3330 | 0.19 | MGMG(18:4) | 18:4 | C27H48NO9 |
| 536.3799 | 536.3799 | 0.00 | MGMG(18:1) | 18:1 | C27H54NO9 |
| 556.3486 | 556.3485 | −0.18 | MGMG(20:5) | 20:5 | C29H50NO9 |
| 766.5469 | 766.5494 | 3.26 | MGDG(34:5) | 20:5/14:0 | C43H76NO10 |
| 774.6090 | 774.6104 | 1.81 | MGDG(34:1) | 18:1/16:0 | C43H84NO10 |
| 794.5782 | 794.5778 | −0.50 | MGDG(36:5) | 20:5/16:0 | C45H80NO10 |
| 822.6095 | 822.6101 | 0.73 | MGDG(38:5) | * | C47H84NO10 |
| 840.5626 | 840.5627 | 0.12 | MGDG(40:10) | 20:5/20:5 | C49H78NO10 |
| 904.6878 | 904.6877 | −0.11 | MGDG(44:6) | * | C53H94NO10 |
| 672.4170 | 672.4172 | 0.30 | DGMG(16:0) | 16:0 | C31H62NO14 |
| 698.4327 | 698.4326 | −0.11 | DGMG(18:1) | 18:1 | C33H64NO14 |
| 718.4014 | 718.4015 | 0.14 | DGMG(20:5) | 20:5 | C35H60NO14 |
| 882.6154 | 882.6173 | 2.15 | DGDG(30:0) | * | C45H88O15N |
| 906.6154 | 906.6166 | 1.32 | DGDG(32:2) | * | C47H88O15N |
| 908.6310 | 908.6309 | −0.11 | DGDG(32:1) | 18:1/14:0 | C47H90O15N |
| 928.5997 | 928.5997 | 0.00 | DGDG(34:5) | 20:5/14:0 | C49H86O15N |
| 932.6310 | 932.6319 | 0.97 | DGDG(34:3) | * | C49H90O15N |
| 934.6467 | 934.645 | −1.82 | DGDG(34:2) | * | C49H92O15N |
| 936.6623 | 936.6623 | 0.00 | DGDG(34:1) | 18:1/16:0 | C49H94O15N |
| 956.6310 | 956.6317 | 0.73 | DGDG(36:5) | 20:5/16:0 | C51H90O15N |
| 984.6623 | 984.6617 | −0.61 | DGDG(38:5) | * | C53H94O15N |
| 1002.6154 | 1002.6160 | 0.60 | DGDG(40:10) | 20:5/20:5 | C55H88O15N |
| Theoretical m/z | Observed m/z | Error (ppm) | Lipid Species (C:N) | Fatty Acyl Chains | Formula |
|---|---|---|---|---|---|
| 468.3090 | 468.3092 | 0.43 | LPC(14:0) | 14:0 | C22H47NO7P |
| 494.3247 | 494.3242 | −1.01 | LPC(16:1) | 16:1 | C24H49NO7P |
| 496.3403 | 496.3401 | −0.40 | LPC(16:0) | 16:0 | C24H51NO7P |
| 516.3090 | 516.3090 | 0.00 | LPC(18:4) | 18:4 | C26H47NO7P |
| 518.3247 | 518.3236 | −2.12 | LPC(18:3) | 18:3 | C26H49NO7P |
| 520.3403 | 520.3395 | −1.54 | LPC(18:2) | 18:2 | C26H51NO7P |
| 522.3560 | 522.3564 | 0.77 | LPC(18:1) | 18:1 | C26H53NO7P |
| 542.3247 | 542.3246 | −0.18 | LPC(20:5) | 20:5 | C28H49NO7P |
| 568.3403 | 568.3405 | 0.35 | LPC(22:6) | 22:6 | C30H51NO7P |
| 570.3560 | 570.3559 | −0.18 | LPC(22:5) | 22:5 | C30H53NO7P |
| 706.5387 | 706.5387 | 0.00 | PC(30:0) | 14:0/16:0 | C38H77NO8P |
| 724.4917 | 724.4916 | −0.14 | PC(32:5) | * | C40H71NO8P |
| 726.5074 | 726.5070 | −0.55 | PC(32:4) | 14:0/18:4 | C40H73NO8P |
| 728.5230 | 728.5233 | 0.41 | PC(32:3) | 14:0/18:3 | C40H75NO8P |
| 730.5387 | 730.5387 | 0.00 | PC(32:2) | 14:0/18:2 and 16:0/16:2 | C40H77NO8P |
| 732.5543 | 732.5541 | −0.27 | PC(32:1) | 16:0/16:1 and 14:0/18:1 | C40H79NO8P |
| 734.5700 | 734.5696 | −0.54 | PC(32:0) | * | C40H81NO8P |
| 750.5074 | 750.5059 | −2.00 | PC(34:6) | * | C42H73NO8P |
| 752.5230 | 752.523 | 0.00 | PC(34:5) | 14:0/20:5 | C42H75NO8P |
| 754.5387 | 754.538 | −0.93 | PC(34:4) | 16:0/18:4 | C42H77NO8P |
| 756.5543 | 756.5541 | −0.26 | PC(34:3) | 16:0/18:3 | C42H79NO8P |
| 758.5700 | 758.5699 | −0.13 | PC(34:2) | 16:0/18:2 | C42H81NO8P |
| 760.5856 | 760.5856 | 0.00 | PC(34:1) | 16:0/18:1 | C42H83NO8P |
| 774.5073 | 774.5058 | −1.94 | PC(36:8) | 16:3/20:5 | C44H73NO8P |
| 776.5230 | 776.5215 | −1.93 | PC(36:7) | 16:2/20:5 | C44H75NO8P |
| 778.5387 | 778.5386 | −0.13 | PC(36:6) | 16:1/20:5 | C44H77NO8P |
| 780.5543 | 780.5544 | 0.13 | PC(36:5) | 16:0/20:5 | C44H79NO8P |
| 784.5856 | 784.5823 | −4.21 | PC(36:3) | * | C44H83NO8P |
| 786.6013 | 786.6002 | −1.40 | PC(36:2) | * | C44H85NO8P |
| 788.6169 | 788.6153 | −2.03 | PC(36:1) | 14:0/22:1 | C44H87NO8P |
| 800.5230 | 800.5225 | −0.62 | PC(38:9) | 18:4/20:5 | C46H75NO8P |
| 802.5387 | 802.5362 | −3.12 | PC(38:8) | 18:3/20:5 | C46H77NO8P |
| 806.5700 | 806.5699 | −0.12 | PC(38:6) | 18:1/20:5 | C46H81NO8P |
| 808.5856 | 808.5845 | −1.36 | PC(38:5) | 18:0/20:5 | C46H83NO8P |
| 810.6013 | 810.597 | −5.30 | PC(38:4) | * | C46H85NO8P |
| 816.6482 | 816.6486 | 0.49 | PC(38:1) | * | C46H91NO8P |
| 826.5387 | 826.5391 | 0.48 | PC(40:10) | 20:5/20:5 | C48H77NO8P |
| 834.6013 | 834.5997 | −1.92 | PC(40:6) | 20:1/20:5 | C48H85NO8P |
| 836.6169 | 836.6153 | −1.91 | PC(40:5) | * | C48H87NO8P |
| 854.5700 | 854.5696 | −0.47 | PC(42:10) | 22:5/20:5 | C50H81NO8P |
| 862.6326 | 862.6334 | 0.93 | PC(42:6) | 22:1/20:5 | C50H89NO8P |
| 864.6482 | 864.6459 | −2.66 | PC(42:5) | 22:0/20:5 | C50H91NO8P |
| 882.6013 | 882.6026 | 1.47 | PC(44:10) | * | C52H85NO8P |
| 890.6639 | 890.6639 | 0.00 | PC(44:6) | 24:1/20:5 | C52H93NO8P |
| 480.3090 | 480.3092 | 0.42 | LPE(18:1) | 18:1 | C23H47NO7P |
| 500.2777 | 500.2776 | −0.20 | LPE(20:5) | 20:5 | C25H43NO7P |
| 658.4448 | 658.4426 | −3.34 | PE(30:3) | * | C35H65NO8P |
| 688.4917 | 688.4936 | 2.76 | PE(32:2) | 16:1/16:1 | C37H71NO8P |
| 690.5074 | 690.5088 | 2.03 | PE(32:1) | 16:0/16:1 and 14:0/18:1 | C37H73NO8P |
| 710.4761 | 710.4753 | −1.13 | PE(34:5) | 14:0/20:5 | C39H69NO8P |
| 712.4917 | 712.4887 | −4.21 | PE(34:4) | 16:0/18:4 | C39H71NO8P |
| 714.5074 | 714.5084 | 1.40 | PE(34:3) | 16:0/18:3 and 16:1/18:2 | C39H73NO8P |
| 716.5230 | 716.5233 | 0.42 | PE(34:2) | 16:1/18:1 and 16:0/18:2 | C39H75NO8P |
| 718.5387 | 718.5385 | −0.28 | PE(34:1) | 16:0/18:1 | C39H77NO8P |
| 736.4917 | 736.4912 | −0.68 | PE(36:6) | * | C41H71NO8P |
| 738.5074 | 738.5073 | −0.14 | PE(36:5) | 16:0/20:5 | C41H73NO8P |
| 742.5387 | 742.5381 | −0.81 | PE(36:3) | * | C41H77NO8P |
| 760.4917 | 760.489 | −3.55 | PE(38:8) | * | C43H71NO8P |
| 764.5230 | 764.5239 | 1.18 | PE(38:6) | 18:1/20:5 | C43H75NO8P |
| 766.5387 | 766.5396 | 1.17 | PE(38:5) | 16:0/22:5 and 18:0/20:5 | C43H77NO8P |
| 784.4917 | 784.492 | 0.38 | PE(40:10) | 20:5/20:5 | C45H71NO8P |
| Theoretical m/z | Observed m/z | Error (ppm) | Lipid Species (C:N) | Fatty Acyl Chains | Formula |
|---|---|---|---|---|---|
| 455.2410 | 455.2425 | 3.29 | LPG(14:0) | 14:0 | C20H40O9P |
| 481.2566 | 481.258 | 2.91 | LPG(16:1) | 16:1 | C22H42O9P |
| 483.2723 | 483.2741 | 3.72 | LPG(16:0) | 16:0 | C22H44O9P |
| 529.2566 | 529.2583 | 3.21 | LPG(20:5) | 20:5 | C26H42O9P |
| 691.4550 | 691.4565 | 2.17 | PG(30:1) | 14:0/16:1 | C36H68O10P |
| 693.4707 | 693.4724 | 2.45 | PG(30:0) | 14:0/16:0 | C36H70O10P |
| 719.4863 | 719.4879 | 2.22 | PG(32:1) | 14:0/18:1 and 16:0/16:1 | C38H72O10P |
| 721.502 | 721.5039 | 2.63 | PG(32:0) | 16:0/16:0 and 14:0/18:0 | C38H74O10P |
| 739.455 | 739.4583 | 4.46 | PG(34:5) | 14:0/20:5 | C40H68O10P |
| 745.502 | 745.5033 | 1.74 | PG(34:2) | 16:0/18:2 and 16:1/18:1 | C40H74O10P |
| 747.5176 | 747.5189 | 1.74 | PG(34:1) | 16:0/18:1 and 14:0/20:1 | C40H76O10P |
| 749.5333 | 749.5345 | 1.60 | PG(34:0) | 16:0/18:0 and 14:0/20:0 | C40H78O10P |
| 765.4707 | 765.4735 | 3.66 | PG(36:6) | 16:1/20:5 | C40H70O10P |
| 767.4863 | 767.4871 | 1.04 | PG(36:5) | * | C42H72O10P |
| 773.5333 | 773.5348 | 1.94 | PG(36:2) | 18:1/18:1 | C42H78O10P |
| 775.5489 | 775.5508 | 2.45 | PG(36:1) | 14:0/22:1 | C42H80O10P |
| 777.5646 | 777.5662 | 2.06 | PG(36:0) | * | C42H82O10P |
| 783.4812 | 783.4810 | −0.29 | PG(36:5-OH) | 16:0-OH/20:5 | C42H72O11P |
| 801.5646 | 801.5668 | 2.74 | PG(38:2) | 16:0/24:2 | C44H82O10P |
| 805.5959 | 805.5945 | −1.74 | PG(38:0) | * | C44H86O10P |
| 813.4707 | 813.4736 | 3.60 | PG(40:10) | 20:5/20:5 | C46H70O10P |
| 829.5959 | 829.599 | 3.74 | PG(40:2) | 16:1/24:1 and 14:0/26:2 | C46H86O10P |
| 831.6115 | 831.6138 | 2.77 | PG(40:1) | 14:0/26:1 | C46H88O10P |
| 859.6428 | 859.6452 | 2.79 | PG(42:1) | 16:0/26:1 | C48H92O10P |
| 855.5024 | 855.5039 | 1.75 | PI(36:5) | 16:0/20:5 | C45H76O13P |
| 881.5180 | 881.5204 | 2.72 | PI(38:6) | 18:1/20:5 | C47H78O13P |
| 901.4867 | 901.4888 | 2.33 | PI(40:10) | 20:5/20:5 | C49H74O13P |
| 965.6119 | 965.6138 | 1.97 | PI(44:6) | * | C53H90O13P |
| 739.4339 | 739.4346 | 0.95 | PA(40:10) | 20:5/20:5 | C43H64O8P |
| 743.4652 | 743.4669 | 2.29 | PA(40:8) | 20:4/20:4 | C43H68O8P |
| Theoretical m/z | Observed m/z | Error (ppm) | Lipid Species (C:N) | Fatty Acyl Chains | Formula |
|---|---|---|---|---|---|
| 892.5915 | 892.5933 | 2.02 | PI-Cer(d38:2) | * | C46H87NO13P |
| 918.6072 | 918.6096 | 2.61 | PI-Cer(d40:3) | * | C48H89NO13P |
| 920.6228 | 920.6217 | −1.19 | PI-Cer(d40:2) | * | C48H91NO13P |
| 922.6385 | 922.6353 | −3.47 | PI-Cer(d40:1) | * | C48H93NO13P |
| 946.6385 | 946.6403 | 1.90 | PI-Cer(d42:3) | * | C50H93NO13P |
| 948.6541 | 948.6535 | −0.63 | PI-Cer(d42:2) | * | C50H95NO13P |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lopes, D.; Melo, T.; Meneses, J.; Abreu, M.H.; Pereira, R.; Domingues, P.; Lillebø, A.I.; Calado, R.; Domingues, M.R. A New Look for the Red Macroalga Palmaria palmata: A Seafood with Polar Lipids Rich in EPA and with Antioxidant Properties. Mar. Drugs 2019, 17, 533. https://doi.org/10.3390/md17090533
Lopes D, Melo T, Meneses J, Abreu MH, Pereira R, Domingues P, Lillebø AI, Calado R, Domingues MR. A New Look for the Red Macroalga Palmaria palmata: A Seafood with Polar Lipids Rich in EPA and with Antioxidant Properties. Marine Drugs. 2019; 17(9):533. https://doi.org/10.3390/md17090533
Chicago/Turabian StyleLopes, Diana, Tânia Melo, Joana Meneses, Maria H. Abreu, Rui Pereira, Pedro Domingues, Ana I. Lillebø, Ricardo Calado, and M. Rosário Domingues. 2019. "A New Look for the Red Macroalga Palmaria palmata: A Seafood with Polar Lipids Rich in EPA and with Antioxidant Properties" Marine Drugs 17, no. 9: 533. https://doi.org/10.3390/md17090533
APA StyleLopes, D., Melo, T., Meneses, J., Abreu, M. H., Pereira, R., Domingues, P., Lillebø, A. I., Calado, R., & Domingues, M. R. (2019). A New Look for the Red Macroalga Palmaria palmata: A Seafood with Polar Lipids Rich in EPA and with Antioxidant Properties. Marine Drugs, 17(9), 533. https://doi.org/10.3390/md17090533