Exploiting Mannuronan C-5 Epimerases in Commercial Alginate Production

 ,
,
Abstract
:1. Introduction
2. Results and Discussion
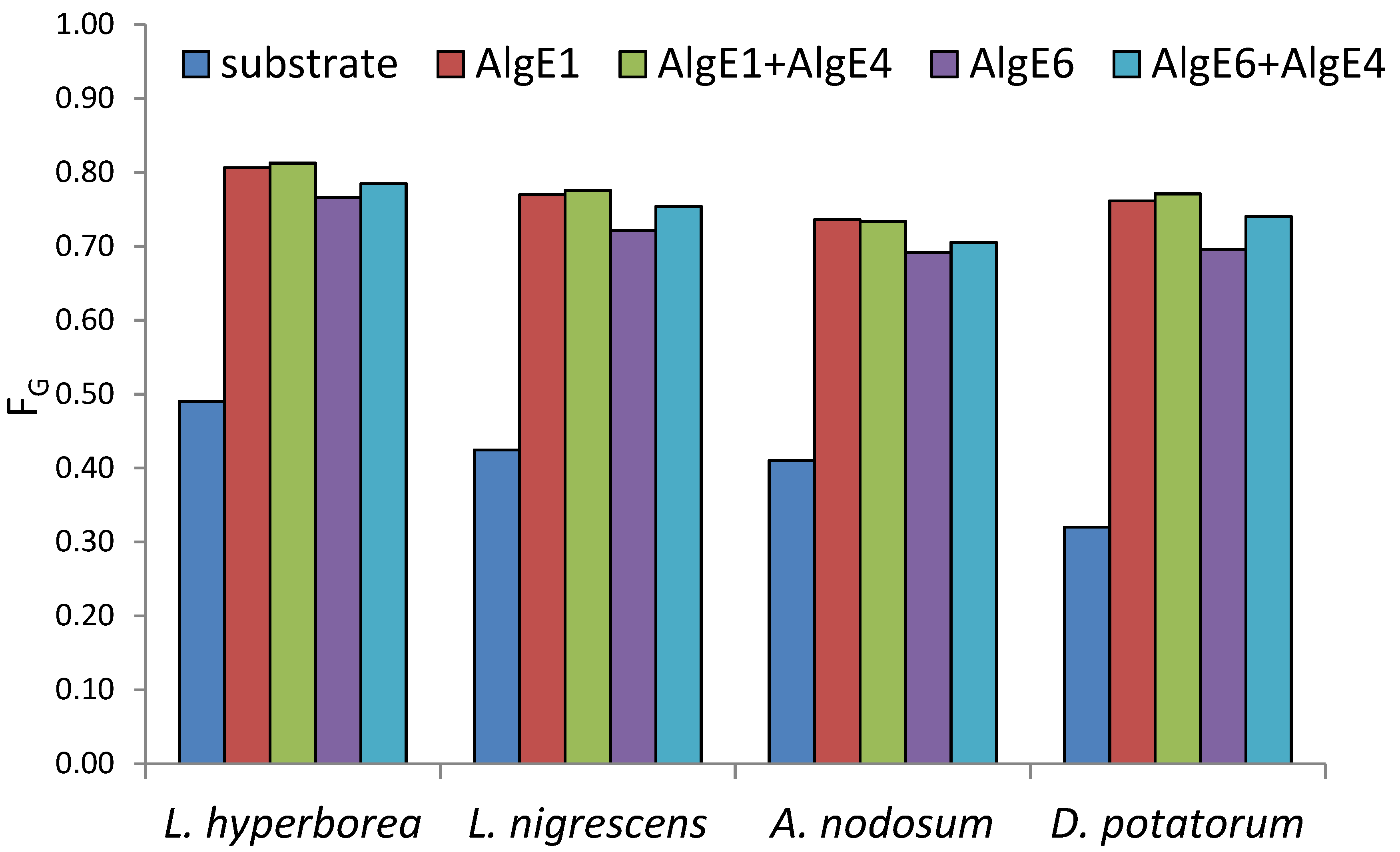
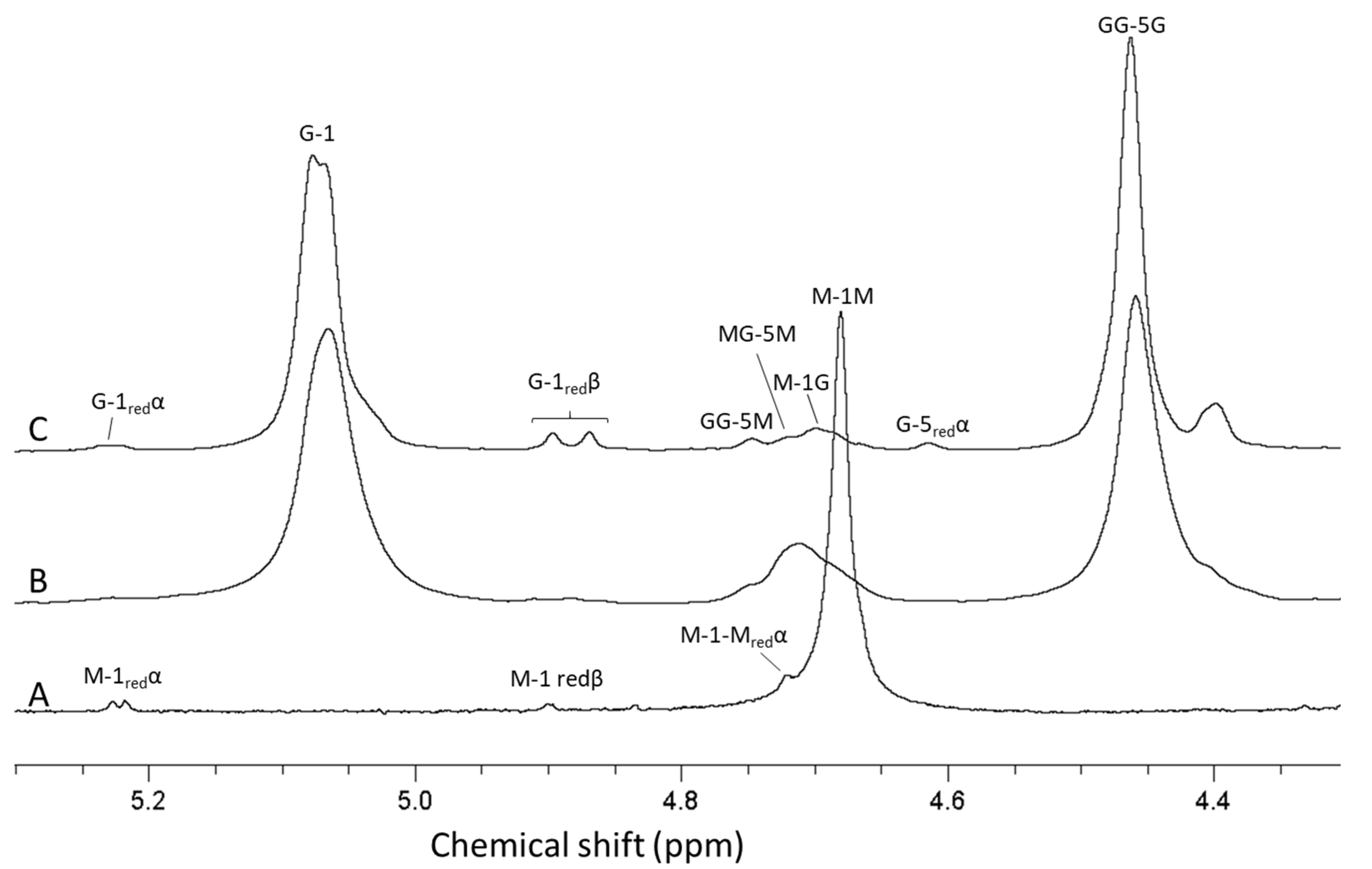
2.1. Seaweed Alginates Rich in Mannuronic Acid Can Be Upgraded and Used for Production of Guluronate Oligomers
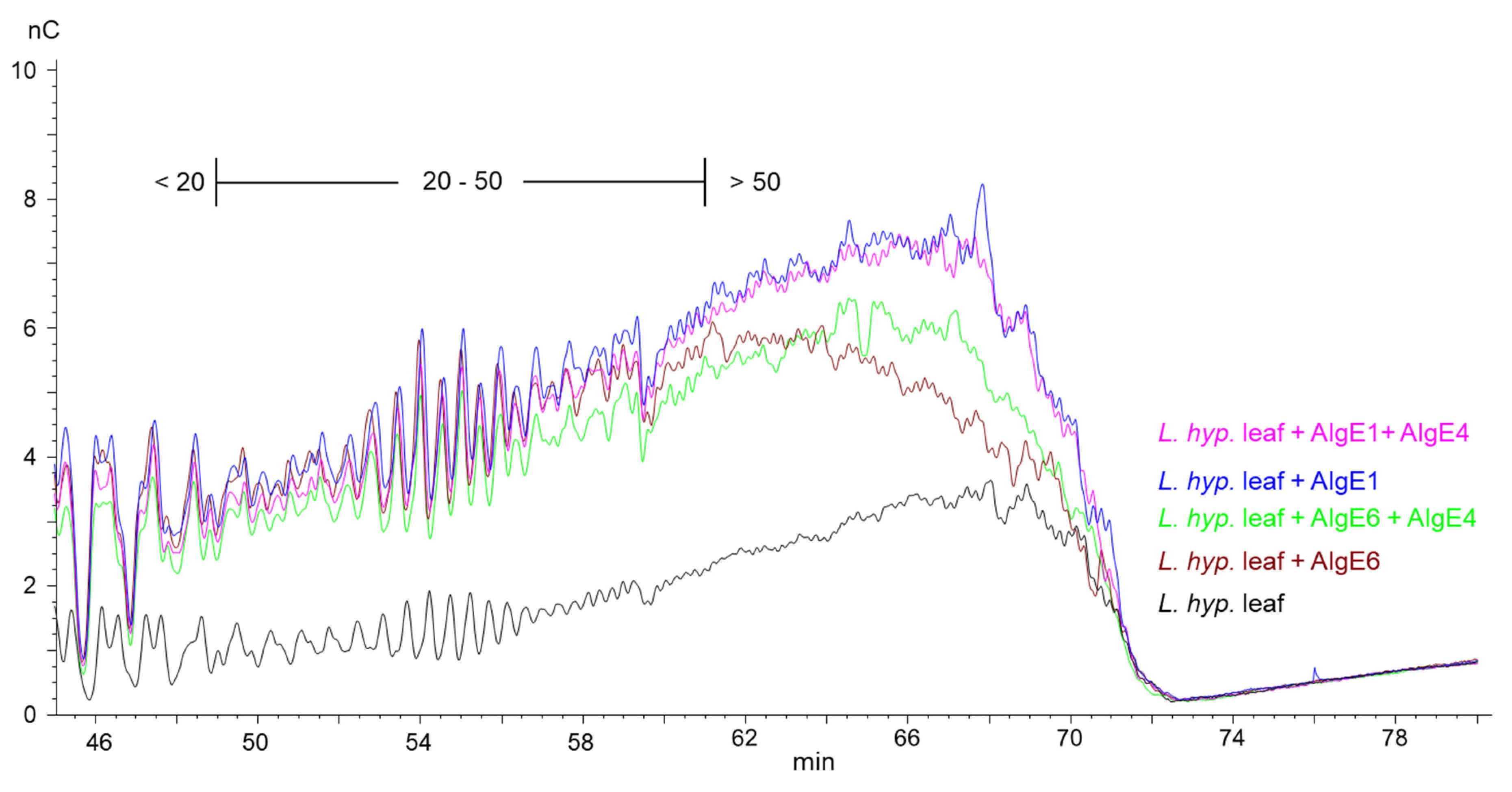
2.2. Guluronate Oligomers Prepared from Bacterially Produced Mannuronan
2.3. Guluronate Oligomers Prepared by In Vitro Epimerization Potentiate the Effect of Antibiotics Similar to OligoG CF-5/20
3. Materials and Methods
3.1. Bacterial Production and Isolation of Mannuronan
3.2. Seaweed Alginates
3.3. Epimerization of Alginate by Mannuronan C-5 Epimerases
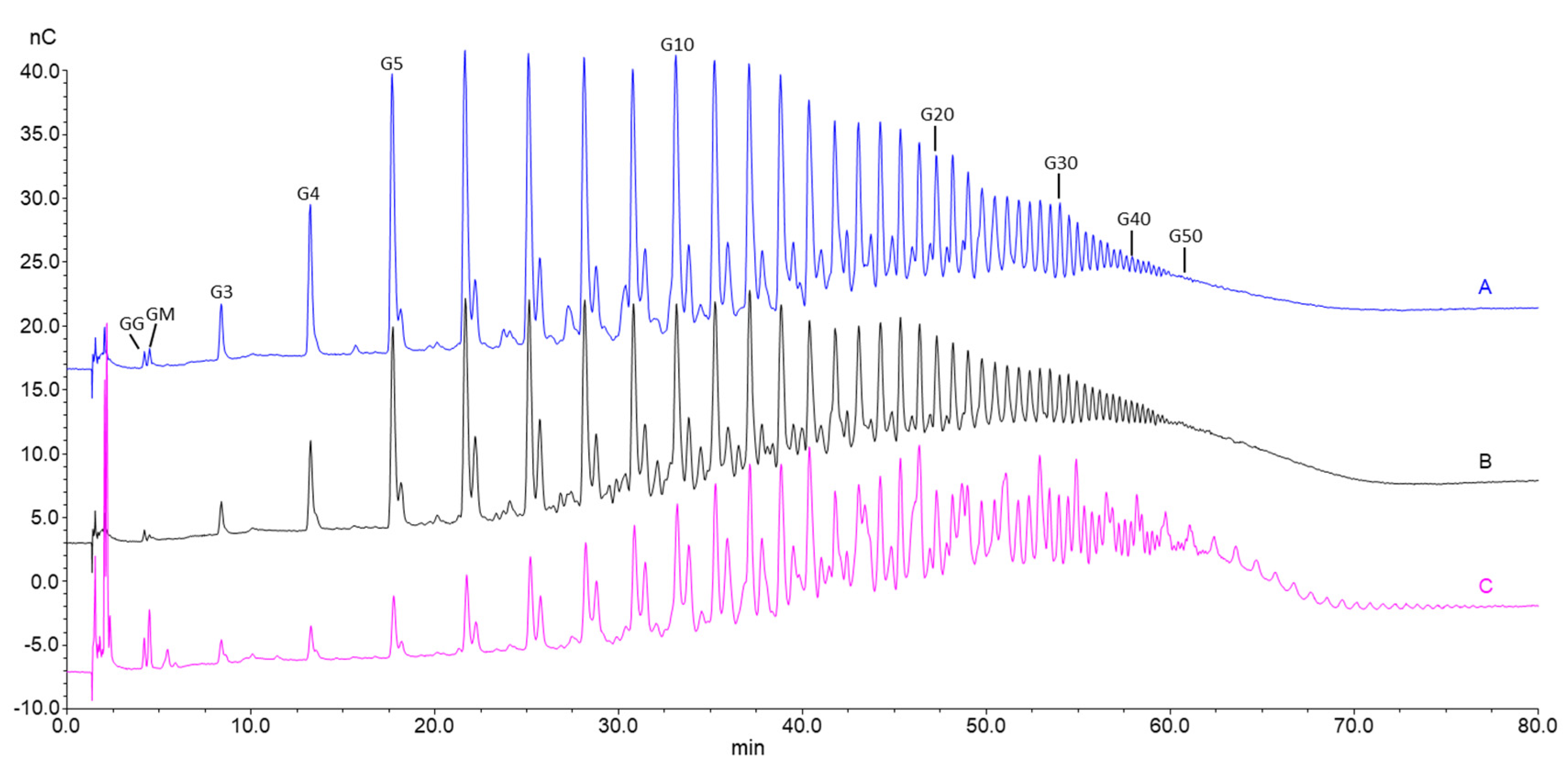
3.4. Production of Oligomers by Acid Hydrolysis of High Molecular Weight Alginate
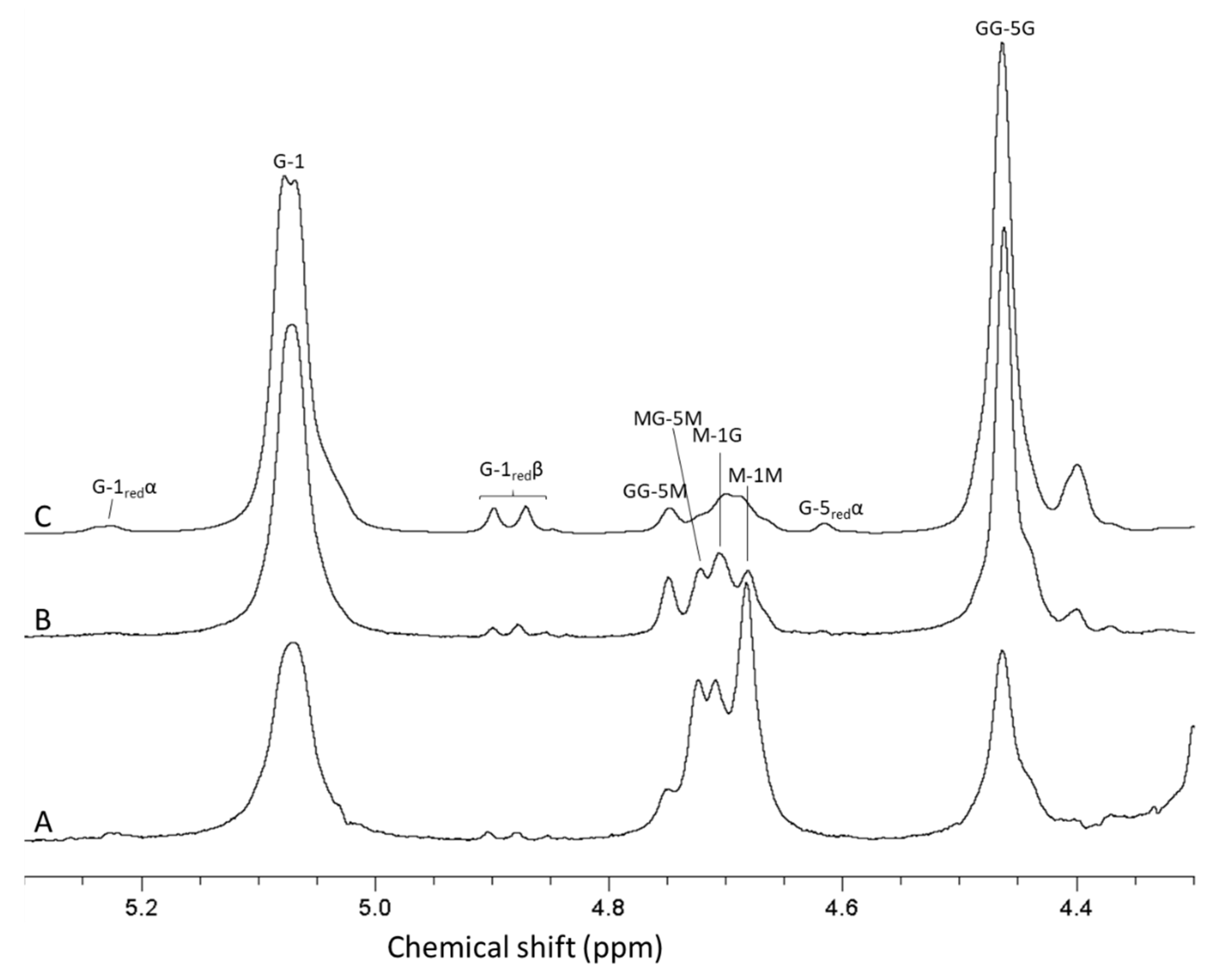
3.5. Characterization and Purification of Alginate Oligomers
3.6. Minimum Inhibitory Concentration Assays
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Drummond, D.W.; Hirst, E.L.; Percival, E. The constitution of alginic acid. J. Chem. Soc. 1962, 1208–1216. [Google Scholar] [CrossRef]
- Fischer, F.G.; Dorfel, H. Polyuronic acid in brown algae. Hoppe Seylers Z Physiol. Chem. 1955, 302, 186–203. [Google Scholar] [CrossRef] [PubMed]
- Skjåk-Bræk, G. Alginates-biosyntheses and some structure-function-relationships relevant to biomedical and biotechnological applications. Biochem. Soc. Trans. 1992, 20, 27–33. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haug, A.; Larsen, B.; Smidsrød, O. Uronic acid sequence in alginate from different sources. Carbohydrate Res. 1974, 32, 217–225. [Google Scholar] [CrossRef]
- Smidsrød, O.; Draget, K.I. Chemistry and physical properties of alginates. Carbohydr. Eur. 1996, 14, 6–13. [Google Scholar]
- Smidsrød, O.; Haug, A. Dependence upon uronic acid composition of some ion-exchange properties of alginates. Acta Chemica Scandinavia 1968, 22, 1989–1997. [Google Scholar] [CrossRef]
- Skjåk-Bræk, G.; Donati, I.; Paoletti, S. Alginate hydrogels: Properties Applications; Pan Stanford Publishing Pte. Ltd.: Singapore, 2015. [Google Scholar]
- Guo, X.; Wang, Y.; Qin, Y.M.; Shen, P.L.; Peng, Q. Structures, properties and application of alginic acid: A review. Int. J. Biol. Macromol. 2020, 162, 618–628. [Google Scholar] [CrossRef]
- Onsøyen, E. Commercial applications of alginates. Carbohydr. Eur. 1996, 14, 26–31. [Google Scholar]
- Page, W.J.; Sadoff, H.L. Relationship between calcium and uronic acids in the encystment of Azotobacter vinelandii. J. Bacteriol. 1975, 122, 1. [Google Scholar] [CrossRef] [Green Version]
- Aarstad, O.A.; Stanisci, A.; Sætrom, G.I.; Tøndervik, A.; Sletta, H.; Aachmann, F.L.; Skjåk-Bræk, G. Biosynthesis and Function of Long Guluronic Acid-Blocks in Alginate Produced by Azotobacter vinelandii. Biomacromolecules 2019, 20, 1613–1622. [Google Scholar] [CrossRef] [Green Version]
- Holtan, S.; Bruheim, P.; Skjåk-Bræk, G. Mode of action and subsite studies of the guluronan block-forming mannuronan C-5 epimerases AlgE1 and AlgE6. Biochem. J. 2006, 395, 319–329. [Google Scholar] [CrossRef] [PubMed]
- Ertesvåg, H.; Valla, S.; Skjåk-Bræk, G. Enzymatic Alginate Modification; Springer: Berlin/Heidelberg, Germany, 2009; Volume 13. [Google Scholar]
- Fischl, R.; Bertelsen, K.; Gaillard, F.; Coelho, S.; Michel, G.; Klinger, M.; Boyen, C.; Czjzek, M.; Herve, C. The cell-wall active mannuronan C5-epimerases in the model brown alga Ectocarpus: From gene context to recombinant protein. Glycobiology 2016, 26, 973–983. [Google Scholar] [CrossRef] [Green Version]
- Inoue, A.; Satoh, A.; Morishita, M.; Tokunaga, Y.; Miyakawa, T.; Tanokura, M.; Ojima, T. Functional heterologous expression and characterization of mannuronan C5-epimerase from the brown alga Saccharina japonica. Algal. Res. 2016, 16, 282–291. [Google Scholar] [CrossRef] [Green Version]
- Mørch, Y.A.; Donati, I.; Strand, B.L.; Skjåk-Bræk, G. Molecular engineering as an approach to design new functional properties of alginate. Biomacromolecules 2007, 8, 2809–2814. [Google Scholar] [CrossRef] [PubMed]
- Sandvig, I.; Karstensen, K.; Rokstad, A.M.; Aachmann, F.L.; Formo, K.; Sandvig, A.; Skjåk-Bræk, G.; Strand, B.L. RGD-peptide modified alginate by a chemoenzymatic strategy for tissue engineering applications. J. Biomed. Mater. Res. Part A 2015, 103, 896–906. [Google Scholar] [CrossRef] [PubMed]
- Arlov, O.; Aachmann, F.L.; Sundan, A.; Espevik, T.; Skjåk-Bræk, G. Heparin-like properties of sulfated alginates with defined sequences and sulfation degrees. Biomacromolecules 2014, 15, 2744–2750. [Google Scholar] [CrossRef] [PubMed]
- Nordgard, C.T.; Draget, K.I. Oligosaccharides as modulators of reheology in complex mucous systems. Biomacromolecules 2011, 12, 3084–3090. [Google Scholar] [CrossRef]
- Sletmoen, M.; Maurstad, G.; Nordgard, C.T.; Draget, K.I.; Stokke, B.T. Oligoguluronate induced competitive displacement of mucin-alginate interactions: Relevance for mucolytic function. Soft Matter. 2012, 8, 8413–8421. [Google Scholar] [CrossRef]
- Vitko, M.; Valerio, D.M.; Rye, P.D.; Onsoyen, E.; Myrset, A.H.; Dessen, A.; Drumm, M.L.; Hodges, C.A. A novel guluronate oligomer improves intestinal transit and survival in cystic fibrosis mice. J. Cyst. Fibros 2016, 15, 745–751. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, M.F.; Powell, L.C.; Menzies, G.E.; Lewis, P.D.; Hawkins, K.; Wright, C.; Doull, I.; Walsh, T.R.; Onsøyen, E.; Dessen, A.; et al. A new class of safe oligosaccharide polymer therapy to modify the mucus barrier of chronic respiratory disease. Mol. Pharm. 2016, 13, 863–872. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, M.F.; Oakley, J.L.; Brilliant, C.D.; Rye, P.D.; Forton, J.; Doull, I.J.M.; Ketchell, I.; Hill, K.E.; Thomas, D.W.; Lewis, P.D. Mucin structural interactions with an alginate oligomer mucolytic in cystic fibrosis sputum. Vib. Spectrosc. 2019, 103, 7. [Google Scholar] [CrossRef]
- Ermund, A.; Recktenwald, C.V.; Skjåk-Bræk, G.; Meiss, L.N.; Onsøyen, E.; Rye, P.D.; Dessen, A.; Myrset, A.H.; Hansson, G.C. OligoG CF-5/20 normalizes cystic fibrosis mucus by chelating calcium. Clin. Exp. Pharmacol. Physiol. 2017, 44, 639–647. [Google Scholar] [CrossRef] [PubMed]
- Jack, A.A.; Khan, S.; Powell, L.C.; Pritchard, M.F.; Beck, K.; Sadh, H.; Sutton, L.; Cavaliere, A.; Florance, H.; Rye, P.D.; et al. Alginate oligosaccharide-induced modification of the lasI-lasR and rhlI-rhlR quorum-sensing systems in pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2018, 62, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, L.C.; Pritchard, M.F.; Ferguson, E.L.; Powell, K.A.; Patel, S.U.; Rye, P.D.; Sakellakou, S.M.; Buurma, N.J.; Brilliant, C.D.; Copping, J.M.; et al. Targeted disruption of the extracellular polymeric network of Pseudomonas aeruginosa biofilms by alginate oligosaccharides. N.P.J. Biofilms Microbiomes 2018, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.Z.; Song, Z.J.; Ciofu, O.; Onsoyen, E.; Rye, P.D.; Hoiby, N. OligoG CF-5/20 disruption of mucoid pseudomonas aeruginosa biofilm in a murine lung infection model. Antimicrob. Agents Chemother. 2016, 60, 2620–2626. [Google Scholar]
- Khan, S.; Tøndervik, A.; Sletta, H.; Klinkenberg, G.; Emanuel, C.; Onsøyen, E.; Myrvold, R.; Howe, R.A.; Walsh, T.R.; Hill, K.E.; et al. Overcoming drug resistance with alginate oligosaccharides able to potentiate the action of selected antibiotics. Antimicrob. Agents Chemother. 2012, 56, 5134–5141. [Google Scholar] [CrossRef] [Green Version]
- Tøndervik, A.; Sletta, H.; Klinkenberg, G.; Emanuel, C.; Powell, L.C.; Pritchard, M.F.; Khan, S.; Craine, K.M.; Onsøyen, E.; Rye, P.D.; et al. Alginate oligosaccharides inhibit fungal cell growth and potentiate the activity of antifungals against Candida and aspergillus spp. PLoS ONE 2014, 9, 10. [Google Scholar] [CrossRef] [Green Version]
- Pritchard, M.F.; Powell, L.C.; Jack, A.A.; Powell, K.; Beck, K.; Florance, H.; Forton, J.; Rye, P.D.; Dessen, A.; Hill, K.E.; et al. A Low-molecular-weight alginate oligosaccharide disrupts pseudomonal microcolony formation and enhances antibiotic effectiveness. Antimicrob. Agents Chemother. 2017, 61, 14. [Google Scholar] [CrossRef] [Green Version]
- Holtan, S.; Zhang, Q.J.; Strand, W.I.; Skjåk-Bræk, G. Characterization of the hydrolysis mechanism of polyalternating alginate in weak acid and assignment of the resulting MG-oligosaccharides by NMR spectroscopy and ESI-mass spectrometry. Biomacromolecules 2006, 7, 2108–2121. [Google Scholar] [CrossRef]
- Gimmestad, M.; Sletta, H.; Ertesvåg, H.; Bakkevig, K.; Jain, S.; Suh, S.; Skjåk-Bræk, G.; Ellingsen, T.E.; Ohman, D.E.; Valla, S. The Pseudomonas fluorescens AlgG protein, but not its mannuronan C-5-epimerase activity, is needed for alginate polymer formation. J. Bacteriol. 2003, 185, 3515–3523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ertesvåg, H.; Skjåk-Bræk, G. Modification of Alginate Using Mannuronan C-5 Epimerases; Humana Press Inc.: Totowa, NJ, USA, 1999; Volume 10. [Google Scholar]
- Sletta, H.; Nedal, A.; Aune, T.E.V.; Hellebust, H.; Hakvåg, S.; Aune, R.; Ellingsen, T.E.; Valla, S.; Brautaset, T. Broad-host-range plasmid pJB658 can be used for industrial-level production of a secreted host-toxic single-chain antibody fragment in Escherichia coli. Appl. Environ. Microbiol. 2004, 70, 7033–7039. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanisci, A.; Aarstad, O.A.; Tøndervik, A.; Sletta, H.; Dypås, L.B.; Skjåk-Bræk, G.; Aachmann, F.L. Overall size of mannuronan C5-Epimerases influences their ability to epimerize modified alginates and alginate gels. Carbohydr. Polym. 2018, 180, 256–263. [Google Scholar] [CrossRef] [PubMed]
- Østgaard, K. Enzymatic microassay for the determination and characterization of alginates. Carbohydr. Polym. 1992, 19, 51–59. [Google Scholar]
- Ballance, S.; Holtan, S.; Aarstad, O.A.; Sikorski, P.; Skjåk-Bræk, G.; Christensen, B.E. Application of high-performance anion-exchange chromatography with pulsed amperometric detection and statistical analysis to study oligosaccharide distributions -a complementary method to investigate the structure and some properties of alginates. J. Chromatogr. A 2005, 1093, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Grasdalen, H. High field, 1HNMR spectroscopy of alginate: Sequential structure and linkage conformations. Carbohydr. Res. 1983, 118, 255–260. [Google Scholar] [CrossRef]
- Aarstad, O.A.; Tøndervik, A.; Sletta, H.; Skjåk-Bræk, G. Alginate Sequencing: An Analysis of Block Distribution in Alginates Using Specific Alginate Degrading Enzymes. Biomacromolecules 2012, 13, 106–116. [Google Scholar] [CrossRef]
- Stenvik, J.; Sletta, H.; Grimstad, O.; Pukstad, B.; Ryan, L.; Aune, R.; Strand, W.; Tøndervik, A.; Torp, S.H.; Skjåk-Bræk, G.; et al. Alginates induce differentiation and expression of CXCR7 and CXCL12/SDF-1 in human keratinocytesu the role of calcium. J. Biomed. Mater. Res. Part. A 2012, 100, 2803–2812. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Guluronic Acid Content (%) | Molecular Weight (kDa) | DPn | |
|---|---|---|---|
| Mannuronan | 0 | 100–150 | n.d. |
| alginate from L. hyperborea leaf | 49 | 250 | n.d. |
| OligoG CF-5/20 | 90–95 | 5 | 15–17 |
| oligomers prepared from epimerized L. hyperborea leaf alginate | 90 | 6.5 | 20 |
| oligomers prepared from epimerized mannuronan | 93 | 5.2 | 17 |
| Pseudomonas aeruginosa | Acinetobacter baumannii | ||||||
|---|---|---|---|---|---|---|---|
| Antibiotic | Oligomer Conc. (%) | OligoG CF-5/20 | Oligomers Prepared from Mannuronan | Oligomers Prepared from L. hyperborea Leaf Alginate | OligoG CF-5/20 | Oligomers Prepared from Mannuronan | Oligomers Prepared from L. hyperborea Leaf Alginate |
| Aztreonam | 0 | 4 | 4 | 4 | 256 | 256 | 256 |
| 3 | 2 | 2 | 2 | 128 | 128 | 128 | |
| Ciprofloxacin | 0 | 0.125 | 0.125 | 0.125 | 2 | 4 | 4 |
| 3 | 0.063 | 0.031 | 0.063 | <0,5 | <0.5 | <0.5 | |
| Ceftazidime | 0 | 2 | 2 | 2 | >512 | >512 | >512 |
| 3 | 1 | 1 | 1 | 256 | 512 | 256 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tøndervik, A.; Aarstad, O.A.; Aune, R.; Maleki, S.; Rye, P.D.; Dessen, A.; Skjåk-Bræk, G.; Sletta, H. Exploiting Mannuronan C-5 Epimerases in Commercial Alginate Production. Mar. Drugs 2020, 18, 565. https://doi.org/10.3390/md18110565
Tøndervik A, Aarstad OA, Aune R, Maleki S, Rye PD, Dessen A, Skjåk-Bræk G, Sletta H. Exploiting Mannuronan C-5 Epimerases in Commercial Alginate Production. Marine Drugs. 2020; 18(11):565. https://doi.org/10.3390/md18110565
Chicago/Turabian StyleTøndervik, Anne, Olav A. Aarstad, Randi Aune, Susan Maleki, Philip D. Rye, Arne Dessen, Gudmund Skjåk-Bræk, and Håvard Sletta. 2020. "Exploiting Mannuronan C-5 Epimerases in Commercial Alginate Production" Marine Drugs 18, no. 11: 565. https://doi.org/10.3390/md18110565
APA StyleTøndervik, A., Aarstad, O. A., Aune, R., Maleki, S., Rye, P. D., Dessen, A., Skjåk-Bræk, G., & Sletta, H. (2020). Exploiting Mannuronan C-5 Epimerases in Commercial Alginate Production. Marine Drugs, 18(11), 565. https://doi.org/10.3390/md18110565