Properties of Carotenoids in Fish Fitness: A Review



Abstract
:1. Introduction
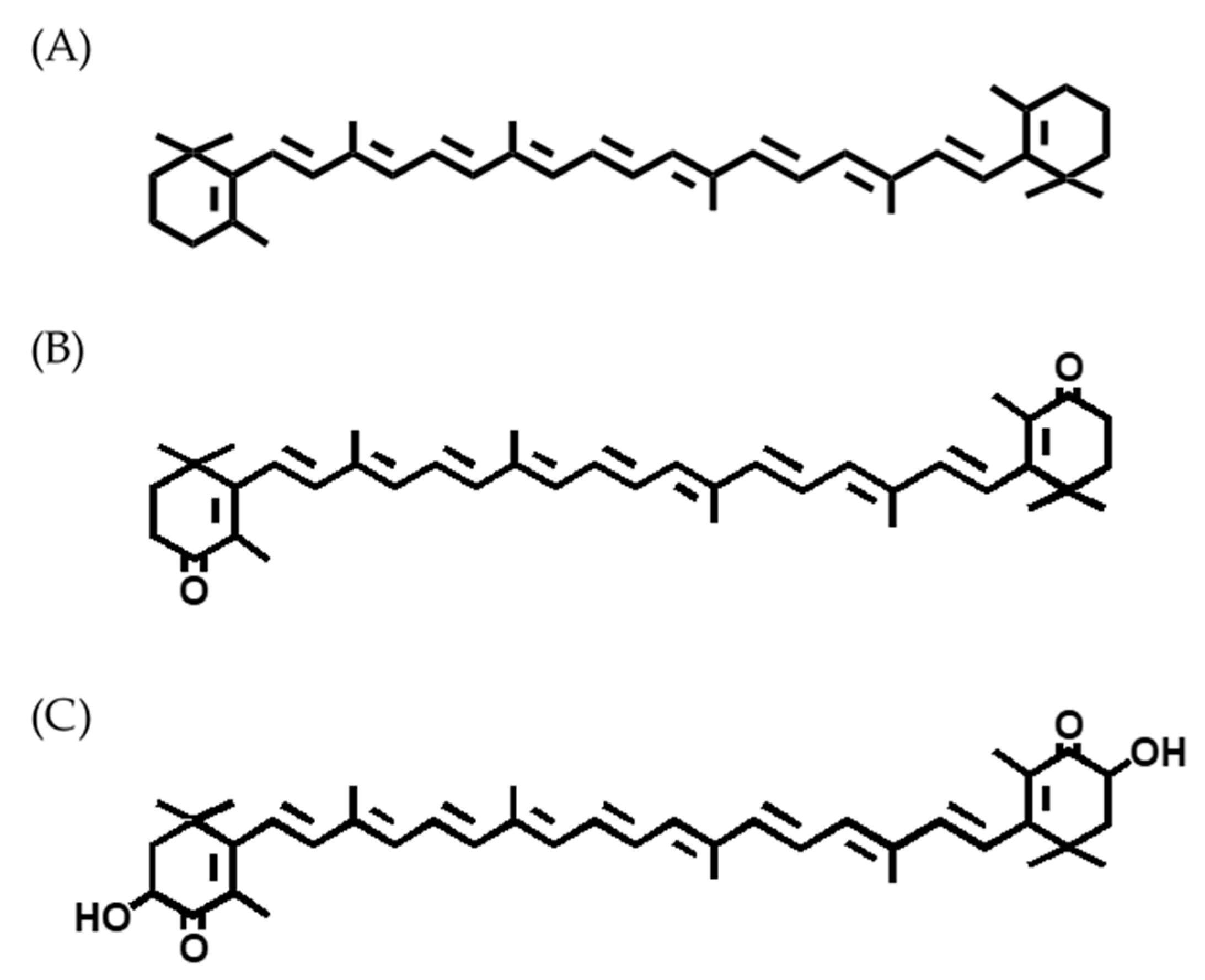
2. Sources of Carotenoids
3. Activities of Carotenoids in Farmed Fish
3.1. Effects of Important Carotenoids Such as ASX on the Fish Immune System
3.2. Effects of Carotenoids, Particularly ASX, on Cellular Damage and Oxidative Stress in Fish
3.3. Molecular Mechanisms of Carotenoid’s Actions in the Body
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Meyers, S.P. Developments in world aquaculture, feed formulations, and role of carotenoids. Pure Appl. Chem. 1994, 66, 1069–1076. [Google Scholar] [CrossRef] [Green Version]
- Fletcher, T.C. Dietary effects on stress and health. In Fish Stress and Health in Aquaculture; Iwama, G.K., Pickering, A.D., Sumpter, J.P., Schreck, C.B., Eds.; Cambridge University Press: Cambridge, UK, 1997; pp. 223–246. [Google Scholar]
- Gatesoupe, F.J. The use of probiotics in aquaculture. Aquaculture 1999, 180, 147–164. [Google Scholar] [CrossRef]
- Olafsen, J.A. Interactions between fish larvae and bacteria in marine aquaculture. Aquaculture 2001, 200, 223–247. [Google Scholar] [CrossRef]
- Irianto, A.; Austin, B. Probiotics in aquaculture. J. Fish Dis. 2002, 25, 633–642. [Google Scholar] [CrossRef]
- Vine, N.G.; Leukes, W.D.; Kaiser, H. Probiotics in marine larviculture. FEMS Microbiol. Rev. 2006, 30, 404–427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakano, T. Microorganisms. In Micronutrients and Health of Cultured Fish; Nakagawa, H., Sato, M., Eds.; Koseisha Koseikaku: Tokyo, Japan, 2003; pp. 95–106. [Google Scholar]
- Pohlenz, C.; Gatlin III, D.M. Interrelationships between fish nutrition and health. Aquaculture 2014, 431, 111–117. [Google Scholar] [CrossRef]
- Burr, G.; Gatlin III, D.M.; Ricke, S. Microbial ecology of the gastrointestinal tract of fish and the potential application of prebiotics and probiotics in finfish aquaculture. J. World Aquacult. Soc. 2005, 36, 425–436. [Google Scholar] [CrossRef]
- Maita, M. Fish health assessment. In Dietary Supplements for the Health and Quality of Cultured Fish; Nakagawa, H., Sato, M., Gatlin, D.M., III, Eds.; CAB International: Oxford, UK, 2007; pp. 86–108. [Google Scholar]
- Nakano, T. Microorganisms. In Dietary Supplements for the Health and Quality of Cultured Fish; Nakagawa, H., Sato, M., Gatlin, D.M., III, Eds.; CAB International: Oxfordshire, UK, 2007; pp. 86–108. [Google Scholar]
- Takeuchi, T. Amino acids, peptides. In Dietary Supplements for the Health and Quality of Cultured Fish; Nakagawa, H., Sato, M., Gatlin, D.M., III, Eds.; CAB International: Oxford, UK, 2007; pp. 47–63. [Google Scholar]
- Koshio, S. Vitamins. In Dietary Supplements for the Health and Quality of Cultured Fish; Nakagawa, H., Sato, M., Gatlin, D.M., III, Eds.; CAB International: Oxford, UK, 2007; pp. 35–46. [Google Scholar]
- Ishikawa, M. Lipids. In Dietary Supplements for the Health and Quality of Cultured Fish; Nakagawa, H., Sato, M., Gatlin, D.M., III, Eds.; CAB International: Oxford, UK, 2007; pp. 64–73. [Google Scholar]
- Gatlin III, D.M.; Barrows, F.T.; Brown, P.; Dabrowski, K.; Gaylord, T.G.; Hardy, R.W.; Herman, E.; Hu, G.; Krogdahl, A.; Nelson, R.; et al. Expanding the utilization of sustainable plant products in aquafeeds: A review. Aquacult. Res. 2007, 38, 551–579. [Google Scholar] [CrossRef]
- Nakano, T.; Osatomi, K.; Miura, N.; Aikawa-Fukuda, Y.; Kanai, K.; Yoshida, A.; Shirakawa, H.; Yamauchi, A.; Yamaguchi, T.; Ochiai, Y. Effect of bacterial infection on the expression of stress proteins and antioxidative enzymes in Japanese flounder. In Evolution of Marine Coastal Ecosystems under the Pressure of Global Changes; Ceccaldi, H.J., Henocque, Y., Komatsu, T., Prouzet, P., Sautour, B., Yoshida, J., Eds.; Springer Nature: Cham, Switzerland, 2020; pp. 111–127. [Google Scholar]
- Yonar, M.E. The effect of lycopene on oxytetracycline-induced oxidative stress and immunosuppression in rainbow trout (Oncorhynchus mykiss, W.). Fish Shellfish Immunol. 2012, 32, 994–1001. [Google Scholar] [CrossRef]
- Elia, A.C.; Ciccotelli, V.; Pacini, N.; Dorr, A.J.M.; Gili, M.; Natali, M.; Gasco, L.; Prearo, M.; Abete, M.C. Transferability of oxytetracycline (OTC) from feed to carp muscle and evaluation of the antibiotic effects on antioxidant systems in liver and kidney. Fish Physiol. Biochem. 2014, 40, 1055–1068. [Google Scholar] [CrossRef]
- Burridge, L.; Weis, J.S.; Cabello, F.; Pizarro, J.; Bostick, K. Chemical use in aquaculture: A review of current practices and possible environmental effects. Aquaculture 2010, 306, 7–23. [Google Scholar] [CrossRef]
- Nakano, T.; Hayashi, S.; Nagamine, N. Effect of excessive doses of oxytetracycline on stress-related biomarker expression in coho salmon. Environ. Sci. Pollut. Res. 2018, 25, 7121–7128. [Google Scholar] [CrossRef] [PubMed]
- Takaichi, S. Carotenoids. In Carotenoids-Biological Functions and Diversity; Takaichi, S., Ed.; Shokabo: Tokyo, Japan, 2006; pp. 1–20. [Google Scholar]
- Khachik, F. Distribution and metabolism of dietary carotenoids in humans as a criterion for development of nutritional supplements. Pure Appl. Chem. 2006, 78, 1551–1557. [Google Scholar] [CrossRef]
- Maoka, T. Diversity and biological functions of natural carotenoids. FFI J. 2015, 220, 118–124. [Google Scholar]
- Maoka, T. Recent progress in structural studies of carotenoids in animals and plants. Arch. Biochem. Biophys. 2009, 483, 191–195. [Google Scholar] [CrossRef]
- Maoka, T. Experimental techniques for structural elucidation and analysis of natural carotenoids. Oreoscience 2012, 12, 485–501. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Concepcion, M.; Avalos, J.; Bonet, M.L.; Boronat, A.; Gomez-Gomez, L.; Hornero-Mendez, D.; Limon, M.C.; Melendez-Martinez, A.J.; Olmedilla-Alonso, B.; Palou, A.; et al. A global perspective on carotenoids: Metabolism, biotechnology, and benefits for nutrition and health. Prog. Lipid Res. 2018, 70, 62–93. [Google Scholar] [CrossRef] [Green Version]
- Lim, K.C.; Yusoff, F.M.; Shariff, M.; Kamarudin, M.S. Astaxanthin as feed supplement in aquatic animals. Rev. Aquac. 2018, 10, 738–773. [Google Scholar] [CrossRef]
- Britton, G. Carotenoid research: History and new perspectives for chemistry in biological systems. Biochim. Biophys. Acta Mol. Cell. Biol. Lipids 2020, 1865, 158699. [Google Scholar] [CrossRef]
- Milani, A.; Basirnejad, M.; Shahbazi, S.; Bolhassani, A. Carotenoids: Biochemistry, pharmacology and treatment. Br. J. Pharmacol. 2017, 174, 1290–1324. [Google Scholar] [CrossRef] [Green Version]
- Nakano, T.; Wazawa, M.; Yamaguchi, T.; Sato, M.; Iwama, G.K. Positive biological actions of astaxanthin in rainbow trout. Mar. Biotechnol. 2004, 6, S100–S105. [Google Scholar]
- Maoka, T. Carotenoids as natural functional pigments. J. Nat. Med. 2020, 74, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, H.D.; Youn, Y.K.; Shin, W.G. Positive effects of astaxanthin of lipid profiles and oxidative stress in overweight subjects. Plant Foods Hum. Nutr. 2011, 66, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Bonet, M.L.; Canas, J.A.; Ribot, J.; Palou, A. Carotenoids and their conversion products in the control of adipocyte function, adiposity and obesity. Arch. Biochem. Biophys. 2015, 572, 112–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aslani, B.A.; Ghobadi, S. Studies on oxidants and antioxidants with a brief glance at their relevance to the immune system. Life Sci. 2016, 146, 163–173. [Google Scholar] [CrossRef]
- Aoi, W.; Naito, Y.; Takanami, Y.; Ishii, T.; Kawai, Y.; Akagiri, S.; Kato, Y.; Osawa, T.; Yoshikawa, T. Astaxanthin improves muscle lipid metabolism in exercise via inhibitory effect of oxidative CPT I modification. Biochem. Biophys. Res. Commun. 2008, 366, 892–897. [Google Scholar] [CrossRef]
- Amaro, H.M.; Barros, R.; Guedes, A.C.; Sousa-Pinto, I.; Malcata, F.X. Microalgal compounds modulate carcinogenesis in the gastrointestinal tract. Trends Biotechnol. 2013, 31, 92–98. [Google Scholar] [CrossRef]
- Krinsky, N.I.; Johnson, E.J. Carotenoid actions and their relation to health and disease. Mol. Aspects Med. 2005, 26, 459–516. [Google Scholar] [CrossRef]
- Kishimoto, Y.; Yoshida, H.; Kondo, K. Potential anti-atherosclerotic properties of astaxanthin. Mar. Drugs 2016, 14, 35. [Google Scholar] [CrossRef]
- World Health Organization. IARC, Carotenoids. In IARC Handbooks of Cancer Prevention; International Agency for Research on Cancer, IARC/WHO: Lyon, France, 1998; pp. 15–45. [Google Scholar]
- Hussein, G.; Nakamura, M.; Zhao, Q.; Iguchi, H.; Goto, H.; Sankawa, U.; Watanabe, H. Anti-hypertensive and neuroprotecitve effects of astaxanthin in experimental animals. Biol. Pharm. Bull. 2005, 28, 47–52. [Google Scholar] [CrossRef] [Green Version]
- Hughes, D.A. Dietary carotenoids and human immune function. Nutrition 2001, 17, 823–827. [Google Scholar] [CrossRef]
- Guerin, M.; Huntley, M.E.; Olaizola, M. Haematococcus astaxanthin: Applications for human health and nutrition. Trends Biotechnol. 2003, 21, 210–216. [Google Scholar] [CrossRef]
- Gammone, M.A.; Riccioni, G.; D’Orazio, N. Marine carotenoids against oxidative stress: Effects on human health. Mar. Drugs 2015, 13, 6226–6246. [Google Scholar] [CrossRef] [PubMed]
- Nakano, T.; Tosa, M.; Takeuchi, M. Improvement of biochemical features in fish health by red yeast and synthetic astaxanthin. J. Agric. Food Chem. 1995, 43, 1570–1573. [Google Scholar] [CrossRef]
- Nakano, T.; Miura, Y.; Wazawa, M.; Sato, M.; Takeuchi, M. Red yeast Phaffia rhodozyma reduces susceptibility of liver homogenate to lipid peroxidation in rainbow trout. Fish. Sci. 1999, 65, 961–962. [Google Scholar] [CrossRef] [Green Version]
- Nakano, T.; Kanmuri, T.; Sato, M.; Takeuchi, M. Effect of astaxanthin rich red yeast (Phaffia rhodozyma) on oxidative stress in rainbow trout. Biochim. Biophys. Acta 1999, 1426, 119–125. [Google Scholar] [CrossRef]
- Miyashita, K.; Hosokawa, M. Focus on a brown seaweed carotenoid, fucoxanthin. FFI J. 2015, 220, 101–109. [Google Scholar]
- Rao, A.V.; Rao, L.G. Carotenoids and human health. Pharmacol. Res. 2007, 55, 207–216. [Google Scholar] [CrossRef]
- Pashkow, F.J.; Watumull, D.G.; Campbell, C.L. Astaxanthin: A novel potential treatment for oxidative stress and inflammation in cardiovascular disease. Am. J. Cardiol. 2008, 101, S58–S68. [Google Scholar] [CrossRef]
- Nishino, H.; Murakoshi, M.; Tokuda, H.; Satomi, Y. Cancer prevention by carotenoids. Arch. Biochem. Biophys. 2009, 483, 165–168. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, H. Multiple mechanisms of anti-cancer effects exerted by astaxanthin. Mar. Drugs 2015, 13, 4310–4330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, H.; Yanai, H.; Ito, K.; Koikeda, T.; Tsukahara, H.; Tada, N. Administration of natural astaxanthin increase serum HDL-cholesterol and adiponectin in subjects with mild hyperlipidemia. Atherosclerosis 2010, 209, 520–523. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Niu, H.; Shao, A.; Wu, C.; Dixon, B.J.; Zhang, J.; Yang, S.; Wang, Y. Astaxanthin as a potential neuroprotective agent for neurological diseases. Mar. Drugs 2015, 13, 5750–5766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tomita, Y. Functions and biological activities in animal. In Carotenoids-Biological Functions and Diversity; Takaichi, S., Ed.; Shokabo: Tokyo, Japan, 2006; pp. 67–107. [Google Scholar]
- Rowles, J.L., 3rd; Erdman, J.W., Jr. Carotenoids and their role in cancer prevention. Biochim. Biophys. Acta Mol. Cell. Biol. Lipids 2020, 1865, 158613. [Google Scholar] [CrossRef]
- Eggersdorfer, M.; Wyss, A. Carotenoids in human nutrition and health. Arch. Biochem. Biophys. 2018, 652, 18–26. [Google Scholar] [CrossRef]
- Bae, M.; Kim, M.B.; Park, Y.K.; Lee, J.Y. Health benefits of fucoxanthin in the prevention of chronic diseases. Biochim. Biophys. Acta Mol. Cell. Biol. Lipids 2020, 1865, 158618. [Google Scholar] [CrossRef]
- Le Goff, M.; Le Ferrec, E.; Mayer, C.; Mimouni, V.; Lagadic-Gossmann, D.; Schoefs, B.; Ulmann, L. Microalgal carotenoids and phytosterols regulate biochemical mechanisms involved in human health and disease prevention. Biochimie 2019, 167, 106–118. [Google Scholar] [CrossRef]
- Zerres, S.; Stahl, W. Carotenoids in human skin. Biochim. Biophys. Acta 2020, 1865, 158588. [Google Scholar] [CrossRef]
- Zheng, J.; Manabe, Y.; Sugawara, T. Siphonaxanthin, a carotenoid from green algae Codium cylindricum, protects Ob/Ob mice fed on a high-fat diet against lipotoxicity by ameliorating somatic stresses and restoring anti-oxidative capacity. Nutr. Res. 2020, 77, 29–42. [Google Scholar] [CrossRef]
- Komatsu, T.; Sasaki, S.; Manabe, Y.; Hirata, T.; Sugawara, T. Preventive effect of dietary astaxanthin on UVA-induced skin photoaging in hairless mice. PLoS ONE 2017, 12, e0171178. [Google Scholar] [CrossRef] [Green Version]
- Mularczyk, M.; Michalak, I.; Marycz, K. Astaxanthin and other nutrients from Haematococcus pluvialis-multifunctional applications. Mar. Drugs 2020, 18, 459. [Google Scholar] [CrossRef] [PubMed]
- Nakao, T.; Sakata, S. Carotenoids powder as a food ingredient. FFI J. 2019, 224, 428–431. [Google Scholar]
- Sathasivam, R.; Ki, J.S. A review of the biological activities of microalgal carotenoids and their potential use in healthcare and cosmetic industries. Mar. Drugs 2018, 16, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- FAO. The State of World Fishery and Aquaculture 2020 (SOFIA); Food and Agriculture Organization of the United Nations: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Olson, J.A. Vitamin A and carotenoids as antioxidants in a physiological context. J. Nutr. Sci. Vitaminol. 1993, 39, S57–S65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mannazzu, I.; Landolfo, S.; Lopes de Silva, T.; Buzzini, P. Red yeasts and carotenoid production: Outlining a future for non-conventional yeasts of biotechnological interest. World J. Microbiol. Biotechnol. 2015, 31, 1665–1673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arathi, B.P.; Sowmya, R.R.; Vijay, K.; Baskaran, V.; Lakshminarayana, R. Metabolomics of carotenids: The challenges and prospects—A review. Trends Food Sci. Technol. 2015, 45, 105–117. [Google Scholar] [CrossRef] [Green Version]
- Storebakken, T.; No, H.K. Pigmentation of rainbow trout. Aquaculture 1992, 100, 209–229. [Google Scholar] [CrossRef]
- Meyers, S.P. The biological/nutritional role of astaxanthin in salmonids and other aquatic species. In Proceedings of the 1st International Symposium of Natural Colorants for Foods, Nutraceuticals, Beverages and Confectionery, Amherst, MA, USA, 7–10 November 1993; pp. 7–10. [Google Scholar]
- Hertrampf, J.W.; Piedad-Pascual, F. Feed carotenoids. In Handbook on Ingredients for Aquaculture Feeds; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 142–156. [Google Scholar]
- Baker, R.; Gunther, C. The role of carotenoids in consumer choice and the likely benefits from their inclusion into products for human consumption. Trends Food Sci. Technol. 2004, 15, 484–488. [Google Scholar] [CrossRef]
- Faehnrich, B.; Lukas, B.; Humer, E.; Zebeli, Q. Phytogenic pigments in animal nutrition: Potentials and risks. J. Sci. Food Agric. 2016, 96, 1420–1430. [Google Scholar] [CrossRef]
- Sahin, K.; Yazlak, H.; Orhan, C.; Tuzcu, M.; Akdemir, F.; Sahin, N. The effect of lycopene on antioxidant status in rainbow trout (Oncorhynchus mykiss) reared under high stocking density. Aquaculture 2014, 418, 132–138. [Google Scholar] [CrossRef]
- Lorenz, R.T.; Cysewski, T. Commercial potential for Haematococcus microalgae as a natural source of astaxanthin. Trends Biotechnol. 2000, 18, 160–166. [Google Scholar] [CrossRef]
- Lee, C.R.; Pham, M.A.; Lee, S.M. Effect of dietary paprika and lipid levels on growth and skin pigmentation of pale chub (Zacco platypus). Asian Aust. J. Anim. Sci. 2010, 23, 724–732. [Google Scholar] [CrossRef]
- Hertrampf, J.W.; Piedad-Pascual, F. Feed yeast. In Handbook on Ingredients for Aquaculture Feeds; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2000; pp. 157–175. [Google Scholar]
- Guedes, A.C.; Amaro, H.M.; Malcata, F.X. Microalgae as sources of carotenoids. Mar. Drugs 2011, 9, 625–644. [Google Scholar] [CrossRef] [PubMed]
- Buyukcapar, H.M.; Yanar, M.; Yanar, Y. Pigmentation of rainbow trout (Oncorhynchus mykiss) with carotenoids from Marigold flower (Tagetes erecta) and red paper (Capsicum annum). Turk. J. Vet. Anim. Sci. 2007, 31, 7–12. [Google Scholar]
- Ambati, R.R.; Phang, S.M.; Ravi, S.; Aswathanarayana, R.G. Astaxanthin: Sources, extraction, stability, biological activities and its commercial applications—A review. Mar. Drugs 2014, 12, 128–152. [Google Scholar] [CrossRef]
- Rodriguez-Amaya, D.B. Natural food colorants: Current knowledge and research trends. FFI J. 2020, 225, 111–130. [Google Scholar]
- Pereira da Costa, D.; Campos Miranda-Filho, K. The use of carotenoid pigments as food additives for aquatic organisms and their functional roles. Rev. Aquaculture 2019, 12, 1567–1578. [Google Scholar] [CrossRef]
- Widjaja-Adhi, M.A.K.; Golczak, M. The molecular aspects of absorption and metabolism of carotenoids and retinoids in vertebrates. Biochim. Biophys. Acta Mol. Cell. Biol. Lipids 2020, 1865, 158571. [Google Scholar] [CrossRef]
- Takaichi, S. Separation and analytical methods. In Carotenoids-Biological Functions and Diversity; Takaichi, S., Ed.; Shokabo: Tokyo, Japan, 2006; pp. 157–199. [Google Scholar]
- Nakano, T.; Shirakawa, H.; Yeo, G.; Devlin, R.H.; Soga, T. Metabolome profiling of growth hormone transgenic coho salmon by capillary electrophoresis time-of-flight mass spectrometry. In Oceanography Challenges to Future Earth; Komatsu, T., Ceccaldi, H.J., Yoshida, J., Prouzet, P., Henocque, Y., Eds.; Springer Nature: Cham, Switzerland, 2019; pp. 223–234. [Google Scholar]
- Hirayama, A.; Kami, K.; Sugimoto, M.; Sugawara, M.; Toki, N.; Onozuka, H.; Kinoshita, T.; Saito, N.; Ochiai, A.; Tomita, M.; et al. Quantitative metabolome profiling of colon and stomach cancer microenvironment by capillary electrophoresis time-of-flight mass spectrometry. Cancer Res. 2009, 69, 4918–4925. [Google Scholar] [CrossRef] [Green Version]
- Miki, W. Carotenoids. In Micronutrients and Health of Cultured Fish; Nakagawa, H., Sato, M., Eds.; Koseisha Koseikaku: Tokyo, Japan, 2003; pp. 42–53. [Google Scholar]
- Karnaukhov, V.N. Carotenoids: Recent progress, problems and prospects. Comp. Biochem. Physiol. 1990, 95B, 1–20. [Google Scholar] [CrossRef]
- Ahmadi, M.R.; Bazyar, A.A.; Safi, S.; Ytrestoyl, T.; Bjerkeng, B. Effects of dietary astaxanthin supplementation on reproductive characteristics of rainbow trout (Oncorhynchus mykiss). J. Appl. Ichthyol. 2006, 22, 388–394. [Google Scholar] [CrossRef]
- Maita, M.; Kiron, V. Nutrition and health in fish. In Nutrition and Feeding in Fish and Crustaceans; Watanabe, T., Ed.; Koseisha Koseikaku: Tokyo, Japan, 2009; pp. 251–269. [Google Scholar]
- Rossi, L.T.; Sharpen, A.R.; Zimmermann, J.A.; Olivero, C.R.; Zbrun, M.V.; Frizzo, L.S.; Signorini, M.L.; Bacchetta, C.; Cian, R.E.; Cazenave, J.; et al. Intestinal microbiota modulation in juvenile Pacú (Piaractus mesopotamicus) by supplementation with Pyropia columbina and β-carotene. Aquacult. Int. 2020, 28, 1001–1016. [Google Scholar] [CrossRef]
- Von Lintig, J.; Moon, J.; Lee, J.; Ramkumar, S. Carotenoid metabolism at the intestinal barrier. Biochim. Biophys. Acta Mol. Cell. Biol. Lipids 2020, 1865. [Google Scholar] [CrossRef] [PubMed]
- Oroian, M.; Escriche, I. Antioxidants: Characterization, natural sources, extraction and analysis. Food Res. Int. 2015, 74, 10–36. [Google Scholar] [CrossRef] [PubMed]
- Pisoschi, A.M.; Pop, A. The role of antioxidants in the chemistry of oxidative stress: A review. Eur. J. Med. Chem. 2015, 97, 55–74. [Google Scholar] [CrossRef] [PubMed]
- Secombes, C.J. The nonspecific immune system: Cellular defenses. In The Fish Immune System; Iwama, G.K., Nakanishi, T., Eds.; Academic Press: San Diego, CA, USA, 1996; pp. 63–103. [Google Scholar]
- Virág, L.; Jaén, R.I.; Regdon, Z.; Boscá, L.; Prieto, P. Self-defense of macrophages against oxidative injury: Fighting for their own survival. Redox Biol. 2019, 26, 101261. [Google Scholar] [CrossRef]
- Taniguchi, N.; Endo, T. Cross talk of SOD, NO, and glutathione metabolism. In Biochemistry in Oxidative Stress and Redox; Taniguchi, N., Yodoi, J., Eds.; Kyoritsu Shuppan: Tokyo, Japan, 2000; pp. 1–11. [Google Scholar]
- Sies, H.; Berndt, C.; Jones, D.P. Oxidative Stress. Annu. Rev. Biochem. 2017, 86, 715–748. [Google Scholar] [CrossRef]
- Forlenza, M.; Scharsack, J.P.; Kachamakova, N.M.; Taverne-Thiele, A.J.; Rombout, J.H.W.M.; Wiegertjes, G.F. Differential contribution of neutrophilic granulocytes and macrophages to nitrosative stress in a host-parasite animal model. Moll. Immunol. 2008, 45, 3178–3189. [Google Scholar] [CrossRef] [PubMed]
- Dixon, B.; Barreda, D.R.; Sunyer, J.O. Perspective on the development and validation of Ab reagents to fish immune proteins for the correct assessment of immune function. Front. Immunol. 2018, 9, 2957. [Google Scholar] [CrossRef]
- Zou, J.; Secombes, C.J. The function of fish cytokines. Biology 2016, 5, 23. [Google Scholar] [CrossRef]
- Abd El-Gawad, E.A.; Wang, H.-P.; Yao, H. Diet supplemented with synthetic carotenoids: Effects on growth performance and biochemical and immunological parameters of yellow perch (Perca flavescens). Front. Physiol. 2019, 10, 1056. [Google Scholar] [CrossRef] [PubMed]
- Li, M.Y.; Sun, L.; Niu, X.T.; Chen, X.M.; Tian, J.X.; Kong, Y.D.; Wang, G.Q. Astaxanthin protects lipopolysaccharide-induced inflammatory response in Channa argus through inhibiting NF-kappa B and MAPKs signaling pathways. Fish Shellfish Immunol. 2019, 86, 280–286. [Google Scholar] [CrossRef] [PubMed]
- Petit, J.; David, L.; Dirks, R.; Wiegertjes, G.F. Genomic and transcriptomic approaches to study immunology in cyprinids: What is next? Dev. Comp. Immunol. 2017, 75, 48–62. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Qu, Y.K.; Wang, A.M.; Yu, Y.B.; Yang, W.P.; Lv, F.; Nie, Q. Effects of carotenoids on the growth performance, biochemical parameters, immune responses and disease resistance of yellow catfish (Pelteobagrus fulvidraco) under high-temperature stress. Aquaculture 2019, 503, 293–303. [Google Scholar] [CrossRef]
- Lim, K.C.; Yusoff, F.M.; Shariff, M.; Kamarudin, M.S.; Nagao, N. Dietary supplementation of astaxanthin enhances hemato-biochemistry and innate immunity of Asian seabass Lates calcarifer (Bloch, 1790). Aquaculture 2019, 512, 734339. [Google Scholar] [CrossRef]
- Cheng, C.H.; Guo, Z.X.; Ye, C.X.; Wang, A.L. Effect of dietary astaxanthin on the growth performance, non-specific immunity, and antioxidant capacity of pufferfish (Takifugu obscurus) under high temperature stress. Fish Physiol. Biochem. 2018, 44, 209–218. [Google Scholar] [CrossRef]
- Anbazahan, S.M.; Mari, L.S.; Yogeshwari, G.; Jagruthi, C.; Thirumurugan, R.; Arockiaraj, J.; Velanganni, A.A.; Krishnamoorthy, P.; Balasundaram, C.; Harikrishnan, R. Immune response and disease resistance of carotenoids supplementation diet in Cyprinus carpio against Aeromonas hydrophila. Fish Shellfish Immunol. 2014, 40, 9–13. [Google Scholar] [CrossRef]
- Abdollahi, Y.; Ahmadifard, N.; Agh, N.; Rahmanifarah, K.; Hejazi, M.A. β-Carotene-enriched Artemia as a natural carotenoid improved skin pigmentation and enhanced the mucus immune responses of platyfish Xiphophorus maculatus. Aquacult. Int. 2019, 27, 1847–1858. [Google Scholar] [CrossRef]
- Alishahi, M.; Karamifar, M.; Mesbah, M. Effects of astaxanthin and Dunaliella salina on skin carotenoids, growth performance and immune response of Astronotus ocellatus. Aquacult. Int. 2015, 23, 1239–1248. [Google Scholar] [CrossRef]
- Villena, J.; Alvarez, S.; Kitazawa, H. Introduction. In Probiotics: Immunobiotics and immunogenics; Kitazawa, H., Villena, J., Alvarez, S., Eds.; CRC Press: Boca Raton, FL, USA, 2014; pp. 1–11. [Google Scholar]
- Katsuyama, M. Carotenoids as vitamin A precursors. In Carotenoids in Marine Organisms; Miki, W., Ed.; Koseisha Koseikaku: Tokyo, Japan, 1993; pp. 23–34. [Google Scholar]
- Iwama, G.K.; Afonso, L.O.B.; Vijayan, M.M. Stress in fishes. In The Physiology of Fishes, 3rd ed.; Evans, D.H., Claiborne, J.B., Eds.; CRC Press: Boca Raton, FL, USA, 2006; pp. 319–342. [Google Scholar]
- Nakano, T.; Afonso, L.O.; Beckman, B.R.; Iwama, G.K.; Devlin, R.H. Acute physiological stress down-regulates mRNA expressions of growth-related genes in coho salmon. PLoS ONE 2013, 8, e71421. [Google Scholar] [CrossRef]
- Basu, N.; Nakano, T.; Grau, E.G.; Iwama, G.K. The effects of cortisol on heat shock protein 70 levels in two fish species. Gen. Comp. Endocrinol. 2001, 124, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Barton, B.A.; Iwama, G.K. Physiological change in fish from stress in aquaculture with emphasis on the response and effects of corticosteroids. Annu. Rev. Fish Dis. 1991, 1, 3–26. [Google Scholar] [CrossRef]
- Wu, H.; Aoki, A.; Arimoto, T.; Nakano, T.; Ohnuki, H.; Murata, M.; Ren, H.; Endo, H. Fish stress become visible: A new attempt to use biosensor for real-time monitoring fish stress. Biosens. Bioelectron. 2015, 67, 503–510. [Google Scholar] [CrossRef] [PubMed]
- Nakano, T.; Kameda, M.; Shoji, Y.; Hayashi, S.; Yamaguchi, T.; Sato, M. Effect of severe environmental thermal stress on redox state in salmon. Redox Biol. 2014, 2, 772–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, H.; Fujii, Y.; Nakano, T.; Arimoto, T.; Murata, M.; Matsumoto, H.; Yoshiura, Y.; Ohnuki, H.; Endo, H. Development of a novel enhanced biosensor system for real-time monitoring of fish stress using a self-assembled monolayer. Biosensors 2019, 19, 1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakano, T.; Wu, H.; Arimoto, T.; Endo, H.; Yamaguchi, T.; Ochiai, Y. Some artificially applied stressors have a positive effect on fish fitness. In Proceedings of the JSFS 85th Anniversary-Commemorative International Symposium “Fisheries Science for Future Generations”, Tokyo, Japan, 22–24 September 2017; Available online: http://www.jsfs.jp/office/annual_meeting/meeting-program/85th/proceeding/pdfs/06010.pdf (accessed on 15 August 2020).
- Livingstone, D.R. Contaminant-stimulated reactive oxygen species production and oxidative damage in aquatic organisms. Mar. Pollut. Bull. 2001, 42, 656–666. [Google Scholar] [CrossRef]
- Valavanidis, A.; Vlahogianni, T.; Dassenakis, M.; Scoullos, M. Molecular biomarkers of oxidative stress in aquatic organisms in relation to toxic environmental pollutants. Ecotoxicol. Environ. Saf. 2006, 64, 178–189. [Google Scholar] [CrossRef]
- Martinez-Alvarez, R.M.; Morales, A.E.; Sanz, A. Antioxidant defenses in fish: Biotic and abiotic factors. Rev. Fish Biol. Fish. 2005, 15, 75–88. [Google Scholar] [CrossRef]
- Lushchak, V.I. Environmentally induced oxidative stress in aquatic animals. Aquat. Toxicol. 2011, 101, 13–30. [Google Scholar] [CrossRef]
- Lesser, M.P. Oxidative stress in marine environments: Biochemistry and physiological ecology. Annu. Rev. Physiol. 2006, 68, 253–278. [Google Scholar] [CrossRef] [Green Version]
- Nakano, T.; Sato, M.; Takeuchi, M. Superoxide dismutase activity in the skin of fish. J. Fish Biol. 1993, 43, 492–496. [Google Scholar] [CrossRef]
- Nakano, T.; Sato, M.; Takeuchi, M. Partial purification and properties of glutathione peroxidase from carp hepatopancreas. Comp. Biochem. Physiol. 1992, 102B, 31–35. [Google Scholar] [CrossRef]
- Nakano, T.; Sato, M.; Takeuchi, M. Glutathione peroxidase of fish. J. Food Sci. 1992, 57, 1116–1119. [Google Scholar] [CrossRef]
- Arrigo, A.P. Gene expression and the thiol redox state. Free Radic. Biol. Med. 1999, 27, 936–944. [Google Scholar] [CrossRef]
- Sies, H. Glutathione and its role in cellular functions. Free Radic. Biol. Med. 1999, 27, 916–921. [Google Scholar] [CrossRef]
- Iwama, G.K.; Thomas, P.T.; Forsyth, R.B.; Vijayan, M.M. Heat shock protein expression in fish. Rev. Fish Biol. Fish. 1998, 8, 35–56. [Google Scholar] [CrossRef]
- Hochachka, P.W.; Somero, G.N. Temperature. In Biochemical Adaptation; Hochachka, P.W., Somero, G.N., Eds.; Oxford University Press: Oxford, UK, 2002; pp. 290–449. [Google Scholar]
- Basu, N.; Todgham, A.E.; Ackerman, P.A.; Bibeau, M.R.; Nakano, K.; Shulte, P.M.; Iwama, G.K. Heat shock protein genes and their functional significance in fish. Gene 2002, 295, 173–183. [Google Scholar] [CrossRef]
- Roberts, R.J.; Agius, C.; Saliba, C.; Bossier, P.; Sung, Y.Y. Heat shock proteins (chaperones) in fish and shellfish and their potential role in relation to fish health: A review. J. Fish Dis. 2010, 33, 789–801. [Google Scholar] [CrossRef]
- Feder, M.E.; Hofmann, G.E. Heat-shock proteins, molecular chaperones, and the stress response. Annu. Rev. Physiol. 1999, 61, 243–282. [Google Scholar] [CrossRef] [Green Version]
- Deane, E.E.; Woo, N.Y.S. Advances and perspectives on the regulation and expression of piscine heat shock proteins. Rev. Fish Biol. Fish. 2011, 21, 153–185. [Google Scholar] [CrossRef]
- Regoli, F.; Giuliani, M.E. Oxidative pathways of chemical toxicity and oxidative stress biomarkers in marine organisms. Mar. Environ. Res. 2014, 93, 106–117. [Google Scholar] [CrossRef] [PubMed]
- Srikanth, K.; Pereira, E.; Duarte, A.C.; Ahmad, I. Glutathione and its dependent enzymes’ modulatory responses to toxic metals and metalloids in fish-a review. Environ. Sci. Pollut. Res. 2013, 20, 2133–2149. [Google Scholar] [CrossRef] [PubMed]
- Ho, E.; Galougahi, K.K.; Liu, C.C.; Bhindi, R.; Figtree, G.A. Biological markers of oxidative stress: Applications to cardiovascular research and practice. Redox Biol. 2013, 1, 483–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ameur, W.B.; De Lapuente, J.; Megdiche, Y.E.; Barhoumi, B.; Trabelsi, S.; Camps, L.; Serret, J.; Ramos-Lopez, D.; Gonzalez-Linares, J.; Driss, M.R.; et al. Oxidative stress, genotoxicity and histopathology biomarker responses in mullet (Mugil cephalus) and sea bass (Dicentrarchus labrax) liver from Bizerte Lagoon (Tunisia). Mar. Pollut. Bull. 2012, 64, 241–251. [Google Scholar] [CrossRef] [PubMed]
- Heise, K.; Puntarulo, S.; Portner, H.O.; Abele, D. Production of reactive oxygen species by isolated mitochondria of the Antarctic bivalve Laternula elliptica (King and Broderip) under heat stress. Comp. Biochem. Physiol. 2003, 134C, 79–90. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Abdel-Tawwab, M.; Abdel-Latif, H.M.R. Lycopene reduces the impacts of aquatic environmental pollutants and physical stressors in fish. Rev. Aquac. 2020, 12, 2511–2526. [Google Scholar] [CrossRef]
- Pan, C.H.; Chien, Y.H.; Wang, Y.J. Antioxidant defence to ammonia stress of characins (Hyphessobrycon eques Steindachner) fed diets supplemented with carotenoids. Aquac. Nutr. 2011, 17, 258–266. [Google Scholar] [CrossRef]
- Hata, Y. Lipid hydroperoxide and disease. In Lipid Peroxides and Nutrition; Igarashi, O., Kaneda, T., Fukuba, H., Mino, M., Eds.; Koseikan: Tokyo, Japan, 1986; pp. 169–214. [Google Scholar]
- Ando, S. Carotenoid transport in fishes. In Carotenoids in Marine Organisms; Miki, W., Ed.; Koseisha Koseikaku: Tokyo, Japan, 1993; pp. 49–58. [Google Scholar]
- Tan, K.; Zhang, H.; Lim, L.-S.; Ma, H.; Li, S.; Zheng, H. Roles of carotenoids in invertebrate immunology. Front. Immunol. 2020, 10. [Google Scholar] [CrossRef]
- Al-Amin, M.M.; Akhter, S.; Hasan, A.T.; Alam, T.; Nageeb Hasan, S.M.; Saifullah, A.R.; Shohel, M. The antioxidant effect of astaxanthin is higher in young mice than aged: A region specific study on brain. Metab. Brain Dis. 2015, 30, 1237–1246. [Google Scholar] [CrossRef]
- Müller, L.; Catalano, A.; Simone, R.; Cittadini, A.; Fröhlich, K.; Böhm, V.; Palozza, P. Antioxidant capacity of tomato seed oil in solution and its redox properties in cultured macrophages. J. Agric. Food Chem. 2013, 61, 346–354. [Google Scholar] [CrossRef]
- Fleischmann, C.; Bar-Ilan, N.; Horowitz, M.; Bruchim, Y.; Deuster, P.; Heled, Y. Astaxanthin supplementation impacts the cellular HSP expression profile during passive heating. Cell Stress Chaperones 2020, 25, 549–558. [Google Scholar] [CrossRef] [PubMed]
- Miki, W. Biological functions and activities of animal carotenoids. Pure Appl. Chem. 1991, 63, 141–146. [Google Scholar] [CrossRef]
- Skibsted, L.H. Carotenoids in antioxidant networks, colorants or radical scavengers. J. Agric. Food Chem. 2012, 60, 2409–2417. [Google Scholar] [CrossRef] [PubMed]
- Bast, A.; Haenen, G.R.; van den Berg, R.; van den Berg, H. Antioxidant effects of carotenoids. Int. J. Vitam. Nutr. Res. 1998, 68, 399–403. [Google Scholar] [PubMed]
- Kaulmann, A.; Bohn, T. Carotenoids, inflammation, and oxidative stress-implications of cellular signaling pathways and relation to chronic disease prevention. Nutr. Res. 2014, 34, 907–929. [Google Scholar] [CrossRef] [PubMed]
- Niu, T.; Xuan, R.; Jiang, L.; Wu, W.; Zhen, Z.; Song, Y.; Hong, L.; Zheng, K.; Zhang, J.; Xu, Q.; et al. Astaxanthin induces the Nrf2/HO-1 antioxidant pathway in human umbilical vein endothelial cells by generating trace amounts of ROS. J. Agric. Food Chem. 2018, 66, 1551–1559. [Google Scholar] [CrossRef]
- Young, A.J.; Lowe, G.M. Antioxidant and prooxidant properties of carotenoids. Arch. Biochem. Biophys. 2001, 385, 20–27. [Google Scholar] [CrossRef]
- Ribeiro, D.; Freitas, M.; Silva, A.M.S.; Carvalho, F.; Fernandes, E. Antioxidant and pro-oxidant activities of carotenoids and their oxidation products. Food Chem. Toxicol. 2018, 120, 681–699. [Google Scholar] [CrossRef]
- Jomova, K.; Valko, M. Health protective effects of carotenoids and their interactions with other biological antioxidants. Eur. J. Med. Chem. 2013, 70, 102–110. [Google Scholar] [CrossRef]
- Droge, W. Free radicals in the physiological control of cell function. Physiol. Rev. 2002, 82, 47–95. [Google Scholar] [CrossRef]
- Allen, R.G.; Tresini, M. Oxidative stress and gene regulation. Free Radic. Biol. Med. 2000, 28, 463–499. [Google Scholar] [CrossRef]
- Burri, B.J. Carotenoids and gene expression. Nutrition 2000, 16, 577–578. [Google Scholar] [CrossRef]
- Suzuki, Y.J.; Forman, H.J.; Sevanian, A. Oxidants as stimulators of signal transduction. Free Radic. Biol. Med. 1997, 22, 269–285. [Google Scholar] [CrossRef]
- Schwarz, K.B. Oxidative stress during viral infection: A review. Free Radic. Biol. Med. 1996, 21, 641–649. [Google Scholar] [CrossRef]
- Osatomi, K.; Kanai, K.; Hara, K.; Ishihara, T. Changes in Cu, Zn-SOD activity in Japanese flounder Paralichtys olivaceus with bacterial infection. Nippon Suisan Gakkaishi 2002, 68, 207–213. [Google Scholar] [CrossRef] [Green Version]
- Maehara, K.; Hasegawa, T.; Xiao, H.; Takeuchi, A.; Abe, R.; Isobe, K. Cooperative interaction of NF-kB and C/EBP binding sites is necessary for manganese superoxide dismutase gene transcription mediated by lipopolysaccharide and interferon-γ. FEBS Lett. 1999, 449, 115–119. [Google Scholar] [CrossRef] [Green Version]
- Brady, T.C.; Chang, L.Y.; Day, B.J.; Crapo, J.D. Extracellular superoxide dismutase is upregulated with inducible nitric oxide synthase after NF-κB activation. Am. J. Physiol. Lung Cell Mol. Physiol. 1997, 273, L1002–L1006. [Google Scholar] [CrossRef]
- Qadri, I.; Iwahashi, M.; Capasso, J.M.; Hopken, M.W.; Flores, S.; Schaack, J.; Simon, F.R. Induced oxidative stress and activated expression of manganese superoxide dismutase during hepatitis C virus replication: Role of JNK, p38 MAPK and AP-1. Biochem. J. 2004, 378, 919–928. [Google Scholar] [CrossRef]
- Wang, J.Y.; Lee, Y.J.; Chou, M.C.; Chang, R.; Chiu, C.H.; Liang, Y.J.; Wu, L.S. Astaxanthin protects steroidogenesis from hydrogen peroxide-induced oxidative stress in mouse Leydig cells. Mar. Drugs 2015, 13, 1375–1388. [Google Scholar] [CrossRef] [Green Version]
- Stentiford, G.D.; Bateman, I.J.; Hinchliffe, S.J.; Bass, D.; Hartnell, R.; Santos, E.M.; Devlin, M.J.; Feist, S.W.; Taylor, N.G.H.; Verner-Jeffreys, D.W.; et al. Sustainable aquaculture through the one health lens. Nature Food 2020, 1, 468–474. [Google Scholar] [CrossRef]
- Naylor, R.L.; Hardy, R.W.; Bureau, D.P.; Chiu, A.; Elliott, M.; Farrell, A.P.; Forster, I.; Gatlin, D.M.; Goldburg, R.J.; Hua, K.; et al. Feeding aquaculture in an era of finite resources. Proc. Natl. Acad. Sci. USA 2009, 106, 15103–15110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naylor, R.L.; Goldburg, R.J.; Primavera, J.H.; Kautsky, N.; Beveridge, M.C.M.; Clay, J.; Folke, C.; Lubchenco, J.; Mooney, H.; Troell, M. Effect of aquaculture on world fish supplies. Nature 2000, 405, 1017–1024. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costello, C.; Cao, L.; Gelcich, S.; Cisneros-Mata, M.A.; Free, C.M.; Froehlich, H.E.; Golden, C.D.; Ishimura, G.; Maier, J.; Macadam-Somer, I.; et al. The future of food from the sea. Nature 2020. [Google Scholar] [CrossRef] [PubMed]
- FAO. FAO-Yearbook of Fishery and Aquaculture tatistics 2017; Food and Agriculture Organization of the United Nations: Rome, Italy, 2019; pp. 81–84. Available online: http://www.fao.org/3/ca5495t/CA5495T.pdf (accessed on 15 August 2020).
- DBJ. Monthly Topics on Global Fish Farming; Development Bank of Japan: Tokyo, Japan, 2014; pp. 7–32. [Google Scholar]
- Nakano, T.; Shoji, Y.; Shirakawa, H.; Suda, Y.; Yamaguchi, T.; Sato, M.; Devlin, R.H. Daily expression patterns of growth-related genes in growth hormone transgenic coho salmon, Oncorhynchus kisutch. La Mer 2011, 49, 111–117. [Google Scholar]
- Nakano, T. Rising to the challenge of reconstructing the coastal fisheries environment following the massive tsunami in Japan: The national 10-year “Tohoku Ecosystem-Associated Marine Sciences (TEAMS)”. In Marine Productivity: Perturbations and Resilience of Socio-Ecosystems; Ceccaldi, H.J., Henocque, Y., Koike, Y., Komatsu, T., Stora, G., Tusseau-Vuillemin, M.H., Eds.; Springer International Publishin: Cham, Switzerland, 2015; pp. 63–68. [Google Scholar]
- Ushio, H.; Kitazawa, D.; Mizuno, H. An innovation of salmonid aquaculture for reconstruction following the Great East Japan Earthquake. In eReconstruction and Renovation Efforts Following the Great East Japan Earthquake by Development of New Technologies; Takuchi, T., Sato, M., Watabe, S., Eds.; Koseisha Koseikaku: Tokyo, Japan, 2017; pp. 43–52. [Google Scholar]
- Edge, R.; Truscott, T.G. Prooxidant and antioxidant reaction mechanisms of carotene and radical interactions with vitamins E and C. Nutrition 1997, 13, 992–994. [Google Scholar] [CrossRef]
- Burton, G.W.; Ingold, K.U. β-Carotene: An unusual type of lipid antioxidant. Science 1984, 224, 569–573. [Google Scholar] [CrossRef]
- Nakano, T.; Shiba, Y.; Yamaguchi, T.; Sato, M.; Ohshima, H. Natural marine products inhibit formation of 8-chloroguanosine mediated by hypochlorous acid, myeloperoxidase and activated neutrophils. Free Radic. Biol. Med. 2012, 53, S95. [Google Scholar] [CrossRef]
- Suzuki, T.; Nakano, T.; Masuda, M.; Ohshima, H. Epigallocatechin gallate markedly enhances formation of 8-oxo-7,8-dihydro-2’-deoxyguanosine with hypochlorous acid. Free Radic. Biol. Med. 2004, 36, 1087–1093. [Google Scholar] [CrossRef]
- Ohshima, H.; Yoshie, Y.; Auriol, S.; Gilibert, I. Antioxidant and pro-oxidant actions of flavonoids: Effects on DNA damage induced by nitric oxide, peroxynitrite and nitroxyl anion. Free Radic. Biol. Med. 1998, 25, 1057–1065. [Google Scholar] [CrossRef]
- Masuda, M.; Suzuki, T.; Friesen, M.D.; Ravanat, J.L.; Cadet, J.; Pignatelli, B.; Nishino, H.; Ohshima, H. Chlorination of guanosine and other nucleosides by hypochlorous acid and myeloperoxidase of activated human neutrophils: Catalysis by nicotine and trimethylamine. J. Biol. Chem. 2001, 276, 40486–40496. [Google Scholar] [CrossRef] [Green Version]
- Nakano, T.; Masuda, M.; Suzuki, T.; Ohshima, H. Inhibition by polyphenolic phytochemicals and sulfurous compounds of the formation of 8-chloroguanosine mediated by hypochlorous acid, human myeloperoxidase, and activated human neutrophils. Biosci. Biotechnol. Biochem. 2012, 76, 2208–2213. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Source |
|---|
| Synthetic astaxanthin (CAROPHYLL Pink, DSM Nutritional Products Ltd.) |
| Synthetic canthaxanthin (CAROPHYLL Red, DSM Nutritional Products Ltd.) |
| Red yeast Xanthophyllomyces dendrnrhous (formerly designated Phaffia rhodozyma) |
| Green microalgae Haematococcus pluvialis |
| Bacteria Paracoccus carotinifaciens |
| Krill Euphausia superba |
| Alga Spirulina spp. |
| Crustacean exoskeleton and meal |
| Red pepper paprika Capsicum annuum L |
| Marigold |
| Biological Function |
|---|
| Antioxidant activity |
| Anti-stress |
| Anti-inflammatory |
| Egg quality |
| Growth performance |
| Immune system |
| Lipid metabolism |
| Liver function |
| Muscle pigmentation |
| Photoprotection |
| Provitamin A activity |
| Reproduction |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nakano, T.; Wiegertjes, G. Properties of Carotenoids in Fish Fitness: A Review. Mar. Drugs 2020, 18, 568. https://doi.org/10.3390/md18110568
Nakano T, Wiegertjes G. Properties of Carotenoids in Fish Fitness: A Review. Marine Drugs. 2020; 18(11):568. https://doi.org/10.3390/md18110568
Chicago/Turabian StyleNakano, Toshiki, and Geert Wiegertjes. 2020. "Properties of Carotenoids in Fish Fitness: A Review" Marine Drugs 18, no. 11: 568. https://doi.org/10.3390/md18110568
APA StyleNakano, T., & Wiegertjes, G. (2020). Properties of Carotenoids in Fish Fitness: A Review. Marine Drugs, 18(11), 568. https://doi.org/10.3390/md18110568