Protection of Alcohol Dehydrogenase against Freeze–Thaw Stress by Ice-Binding Proteins Is Proportional to Their Ice Recrystallization Inhibition Property



{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
2.1. IBPs Show Resistance to the Repeated Freeze–Thaw Cycle
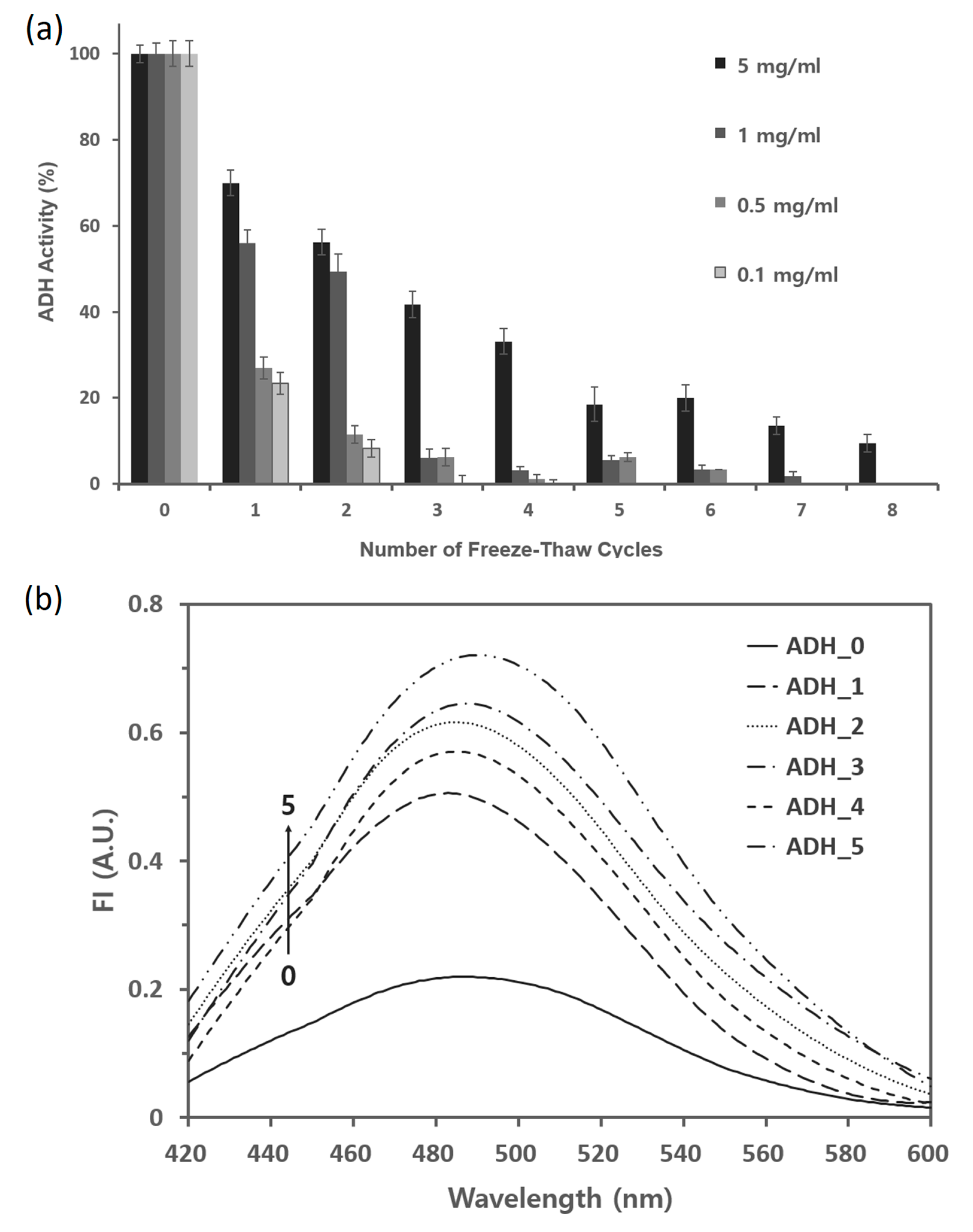
2.2. ADH Is Susceptible to the Repeated Freeze–Thaw Cycles
2.3. Activity of ADH Is Rescued Proportional to the IRI Property of IBPs
3. Materials and Methods
3.1. Chemicals and Proteins
3.2. Ice Recrystallization Inhibition Assay
3.3. Freeze–Thaw of ADH and Its Activity Assay
3.4. ANS Fluorescence Measurement
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Davies, P.L. Ice-binding proteins: A remarkable diversity of structures for stopping and starting ice growth. Trends Biochem. Sci. 2014, 39, 548–555. [Google Scholar] [CrossRef]
- Kim, H.J.; Lee, J.H.; Hur, Y.B.; Lee, C.W.; Park, S.-H.; Koo, B.-W. Marine antifreeze proteins: Structure, function, and application to cryopreservation as a potential cryoprotectant. Mar. Drugs 2017, 15, 27. [Google Scholar] [CrossRef] [Green Version]
- Davies, P.L.; Sykes, B.D. Antifreeze proteins. Curr. Opin. Struct. Biol. 1997, 7, 828–834. [Google Scholar] [CrossRef]
- Raymond, J.A.; Janech, M.G. Cryoprotective property of diatom ice-active substance. Cryobiology 2003, 46, 203–204. [Google Scholar] [CrossRef]
- Griffith, M.; Lumb, C.; Wiseman, S.B.; Wisniewski, M.; Johnson, R.W.; Marangoni, A.G. Antifreeze proteins modify the freezing process in planta. Plant. Physiol 2005, 138, 330–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janech, M.; Krell, A.; Mock, T.; Kang, J.-S.; Raymond, J. Ice-binding proteins from sea ice diatoms (Bacillariophyceae). J. Phycol. 2006, 42, 410–416. [Google Scholar] [CrossRef] [Green Version]
- Raymond, J.A.; Knight, C.A. Ice binding, recrystallization inhibition, and cryoprotective properties of ice-active substances associated with Antarctic sea ice diatoms. Cryobiology 2003, 46, 174–181. [Google Scholar] [CrossRef]
- Knight, C.A.; Wen, D.; Laursen, R.A. Nonequilibrium antifreeze peptides and the recrystallization of ice. Cryobiology 1995, 32, 23–34. [Google Scholar] [CrossRef]
- Raymond, J.A.; Fritsen, C.H. Semipurification and ice recrystallization inhibition activity of ice-active substances associated with Antarctic photosynthetic organisms. Cryobiology 2001, 43, 63–70. [Google Scholar] [CrossRef]
- Rodriguez, C.; Sajjadi, S.; Abrol, R.; Wen, X. A beetle antifreeze protein protects lactate dehydrogenase under freeze-thawing. Int. J. Biol. Macromol. 2019, 136, 1153–1160. [Google Scholar] [CrossRef]
- Knight, C.A.; Duman, J.G. Inhibition of recrystallization of ice by insect thermal hysteresis proteins: A possible cryoprotective role. Cryobiology 1986, 23, 256–262. [Google Scholar] [CrossRef]
- Halwani, D.O.; Brockbank, K.G.; Duman, J.G.; Campbell, L.H. Recombinant Dendroides canadensis antifreeze proteins as potential ingredients in cryopreservation solutions. Cryobiology 2014, 68, 411–418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitchell, D.E.; Fayter, A.E.R.; Deller, R.C.; Hasan, M.; Gutierrez-Marcos, J.; Gibson, M.I. Ice-recrystallization inhibiting polymers protect proteins against freeze-stress and enable glycerol-free cryostorage. Mater. Horiz. 2018, 6, 364–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leavy, O. Therapeutic antibodies: Past, present and future. Nat. Rev. Immunol. 2010, 10, 297. [Google Scholar] [CrossRef] [PubMed]
- Scott, A.M.; Wolchok, J.D.; Old, L.J. Antibody therapy of cancer. Nat. Rev. Cancer 2012, 12, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Gil, D.; Schrum, A.G. Strategies to stabilize compact folding and minimize aggregation of antibody-based fragments. Adv. Biosci. Biotechnol. 2013, 4, 73–84. [Google Scholar] [CrossRef] [Green Version]
- Kotsovilis, S.; Andreakos, E. Therapeutic human monoclonal antibodies in inflammatory diseases. Methods Mol. Biol. 2014, 1060, 37–59. [Google Scholar] [CrossRef]
- Lee, J.H.; Park, A.K.; Do, H.; Park, K.S.; Moh, S.H.; Chi, Y.M.; Kim, H.J. Structural basis for the antifreeze activity of an ice-binding protein from an Arctic yeast. J. Biol. Chem. 2012, 287, 11460–11468. [Google Scholar] [CrossRef] [Green Version]
- Do, H.; Kim, S.J.; Kim, H.J.; Lee, J.H. Structure-based characterization and antifreeze properties of a hyperactive ice-binding protein from the Antarctic bacterium Flavobacterium frigoris PS1. Acta Crystallogr. D Biol. Crystallogr. 2014, 70, 1061–1073. [Google Scholar] [CrossRef]
- Lee, J.; Kim, S.K.; Youm, H.W.; Kim, H.J.; Lee, J.R.; Suh, C.S.; Kim, S.H. Effects of three different types of antifreeze proteins on mouse ovarian tissue cryopreservation and transplantation. PLoS ONE 2015, 10, e0126252. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.H.; Lee, H.J.; Kim, H.J.; Lee, J.H.; Ko, Y.; Kim, S.M.; Lee, J.R.; Suh, C.S.; Kim, S.H. Effects of antifreeze proteins on the vitrification of mouse oocytes: Comparison of three different antifreeze proteins. Hum. Reprod. 2015, 30, 2110–2119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.R.; Youm, H.W.; Lee, H.J.; Jee, B.C.; Suh, C.S.; Kim, S.H. Effect of antifreeze protein on mouse ovarian tissue cryopreservation and transplantation. Yonsei Med. J. 2015, 56, 778–784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carpenter, J.F.; Hansen, T.N. Antifreeze protein modulates cell survival during cryopreservation: Mediation through influence on ice crystal growth. Proc. Natl. Acad. Sci. USA 1992, 89, 8953–8957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amir, G.; Rubinsky, B.; Kassif, Y.; Horowitz, L.; Smolinsky, A.K.; Lavee, J. Preservation of myocyte structure and mitochondrial integrity in subzero cryopreservation of mammalian hearts for transplantation using antifreeze proteins—An electron microscopy study. Eur. J. Cardiothorac. Surg. 2003, 24, 292–297. [Google Scholar] [CrossRef] [Green Version]
- Klinman, J.P.; Welsh, K. The zinc content of yeast alcohol dehydrogenase. Biochem. Biophys. Res. Commun. 1976, 70, 878–884. [Google Scholar] [CrossRef]
- Zheng, Y.G.; Yin, H.H.; Yu, D.F.; Chen, X.; Tang, X.L.; Zhang, X.J.; Xue, Y.P.; Wang, Y.J.; Liu, Z.Q. Recent advances in biotechnological applications of alcohol dehydrogenases. Appl. Microbiol. Biotechnol. 2017, 101, 987–1001. [Google Scholar] [CrossRef]
- Suzuki, T.; Imamura, K.; Yamamoto, K.; Satoh, T.; Okazaki, M. Thermal stabilization of freeze-dried enzymes by sugars. J. Chem. Eng. Jpn. 1997, 30, 609–613. [Google Scholar] [CrossRef] [Green Version]
- Bolivar, J.M.; Wilson, L.; Ferrarotti, S.A.; Guisán, J.M.; Fernández-Lafuente, R.; Mateo, C. Improvement of the stability of alcohol dehydrogenase by covalent immobilization on glyoxyl-agarose. J. Biotechnol. 2006, 125, 85–94. [Google Scholar] [CrossRef]
- Miroliaei, M.; Ranjbar, B.; Naderi-Manesh, H.; Nemat-Gorgani, M. Thermal denaturation of yeast alcohol dehydrogenase and protection of secondary and tertiary structural changes by sugars: CD and fluorescence studies. Enzyme Microb. Technol. 2007, 40, 896–901. [Google Scholar] [CrossRef]
- Cao, E.; Chen, Y.; Cui, Z.; Foster, P.R. Effect of freezing and thawing rates on denaturation of proteins in aqueous solutions. Biotechnol. Bioeng. 2003, 82, 684–690. [Google Scholar] [CrossRef]
- Knight, C.A.; Hallett, J.; DeVries, A.L. Solute effects on ice recrystallization: An assessment technique. Cryobiology 1988, 25, 55–60. [Google Scholar] [CrossRef]
- Gabellieri, E.; Strambini, G.B. Perturbation of protein tertiary structure in frozen solutions revealed by 1-anilino-8-naphthalene sulfonate fluorescence. Biophys. J. 2003, 85, 3214–3220. [Google Scholar] [CrossRef] [Green Version]
- Semisotnov, G.V.; Rodionova, N.A.; Razgulyaev, O.I.; Uversky, V.N.; Gripas’, A.F.; Gilmanshin, R.I. Study of the “molten globule” intermediate state in protein folding by a hydrophobic fluorescent probe. Biopolymers 1991, 31, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Bai, J.H.; Wang, H.J.; Zhou, H.M. Alkaline-induced unfolding and salt-induced folding of pig heart lactate dehydrogenase under high pH conditions. Int. J. Biol. Macromol. 1998, 23, 127–133. [Google Scholar] [CrossRef]
- Garber Cohen, I.P.; Castello, P.R.; Flecha, F.L.G. Ice-induced partial unfolding and aggregation of an integral membrane protein. Biochim. Biophys. Acta Biomembr. 2010, 1798, 2040–2047. [Google Scholar] [CrossRef] [Green Version]
- Izutsu, K.-I.; Yoshioka, S.; Terao, T. Stabilizing effect of amphiphilic excipients on the freeze-thawing and freeze-drying of lactate dehydrogenase. Biotechnol. Bioeng. 1994, 43, 1102–1107. [Google Scholar] [CrossRef]
- Anchordoquy, T.J.; Izutsu, K.I.; Randolph, T.W.; Carpenter, J.F. Maintenance of quaternary structure in the frozen state stabilizes lactate dehydrogenase during freeze-drying. Arch. Biochem. Biophys. 2001, 390, 35–41. [Google Scholar] [CrossRef]
- Zhang, T.; Zhou, H.M. Comparison of inactivation and unfolding of yeast alcohol dehydrogenase during denaturation in urea solutions. Int. J. Biol. Macromol. 1996, 19, 113–119. [Google Scholar] [CrossRef]
- He, B.; Bai, J.H.; Zhou, H.M. Comparison of inactivation and unfolding of yeast alcohol dehydrogenase during thermal denaturation. Int. J. Biochem. Cell Biol. 1997, 29, 1021–1028. [Google Scholar] [CrossRef]
- Roessl, U.; Leitgeb, S.; Nidetzky, B. Protein freeze concentration and micro-segregation analysed in a temperature-controlled freeze container. Biotechnol. Rep. 2015, 6, 108–111. [Google Scholar] [CrossRef] [Green Version]
- Kim, H.J.; Shim, H.E.; Lee, J.H.; Kang, Y.-C.; Hur, Y.B. Ice-binding protein derived from glaciozyma can improve the viability of cryopreserved mammalian cells. J. Microbiol. Biotechnol. 2015, 25, 1989–1996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, H.J.; Koo, B.W.; Kim, D.; Seo, Y.S.; Nam, Y.K. Effect of marine-derived ice-binding proteins on the cryopreservation of marine microalgae. Mar. Drugs 2017, 15, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hansen, T.N.; Smith, K.M.; Brockbank, K.G. Type I antifreeze protein attenuates cell recoveries following cryopreservation. Transpl. Proc. 1993, 25, 3182–3184. [Google Scholar]
- Shaw, J.M.; Ward, C.; Trounson, A.O. Evaluation of propanediol, ethylene glycol, sucrose and antifreeze proteins on the survival of slow-cooled mouse pronuclear and 4-cell embryos. Hum. Reprod. 1995, 10, 396–402. [Google Scholar] [CrossRef]
- Garner, J.; Harding, M.M. Design and synthesis of antifreeze glycoproteins and mimics. ChemBioChem 2010, 11, 2489–2498. [Google Scholar] [CrossRef]
- Trant, J.F.; Biggs, R.A.; Capicciotti, C.J.; Ben, R.N. Developing highly active small molecule ice recrystallization inhibitors based upon C-linked antifreeze glycoprotein analogues. RSC Adv. 2013, 3, 26005–26009. [Google Scholar] [CrossRef]
- Balcerzak, A.K.; Capicciotti, C.J.; Briard, J.G.; Ben, R.N. Designing ice recrystallization inhibitors: From antifreeze (glyco) proteins to small molecules. RSC Adv. 2014, 4, 42682–42696. [Google Scholar] [CrossRef]
- Capicciotti, C.J.; Kurach, J.D.R.; Turner, T.R.; Mancini, R.S.; Acker, J.P.; Ben, R.N. Small molecule ice recrystallization inhibitors enable freezing of human red blood cells with reduced glycerol concentrations. Sci. Rep. 2015, 5, 9692. [Google Scholar] [CrossRef] [Green Version]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, Y.H.; Kim, K.; Lee, J.H.; Kim, H.J. Protection of Alcohol Dehydrogenase against Freeze–Thaw Stress by Ice-Binding Proteins Is Proportional to Their Ice Recrystallization Inhibition Property. Mar. Drugs 2020, 18, 638. https://doi.org/10.3390/md18120638
Lee YH, Kim K, Lee JH, Kim HJ. Protection of Alcohol Dehydrogenase against Freeze–Thaw Stress by Ice-Binding Proteins Is Proportional to Their Ice Recrystallization Inhibition Property. Marine Drugs. 2020; 18(12):638. https://doi.org/10.3390/md18120638
Chicago/Turabian StyleLee, Young Hoon, Kitae Kim, Jun Hyuck Lee, and Hak Jun Kim. 2020. "Protection of Alcohol Dehydrogenase against Freeze–Thaw Stress by Ice-Binding Proteins Is Proportional to Their Ice Recrystallization Inhibition Property" Marine Drugs 18, no. 12: 638. https://doi.org/10.3390/md18120638
APA StyleLee, Y. H., Kim, K., Lee, J. H., & Kim, H. J. (2020). Protection of Alcohol Dehydrogenase against Freeze–Thaw Stress by Ice-Binding Proteins Is Proportional to Their Ice Recrystallization Inhibition Property. Marine Drugs, 18(12), 638. https://doi.org/10.3390/md18120638