The Reducing Effects of Pyrogallol-Phloroglucinol-6,6-Bieckol on High-Fat Diet-Induced Pyroptosis in Endothelial and Vascular Smooth Muscle Cells of Mice Aortas

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
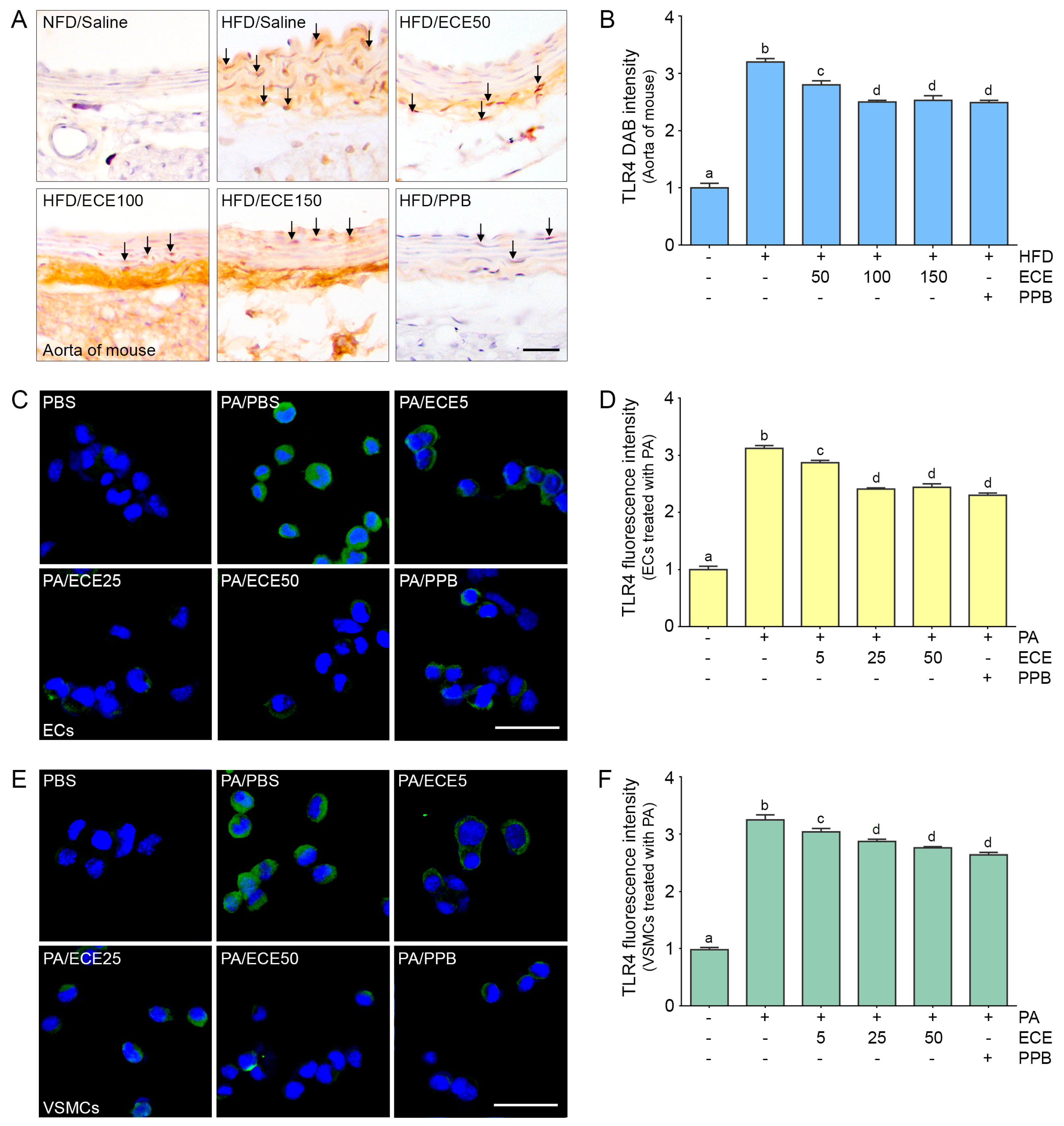
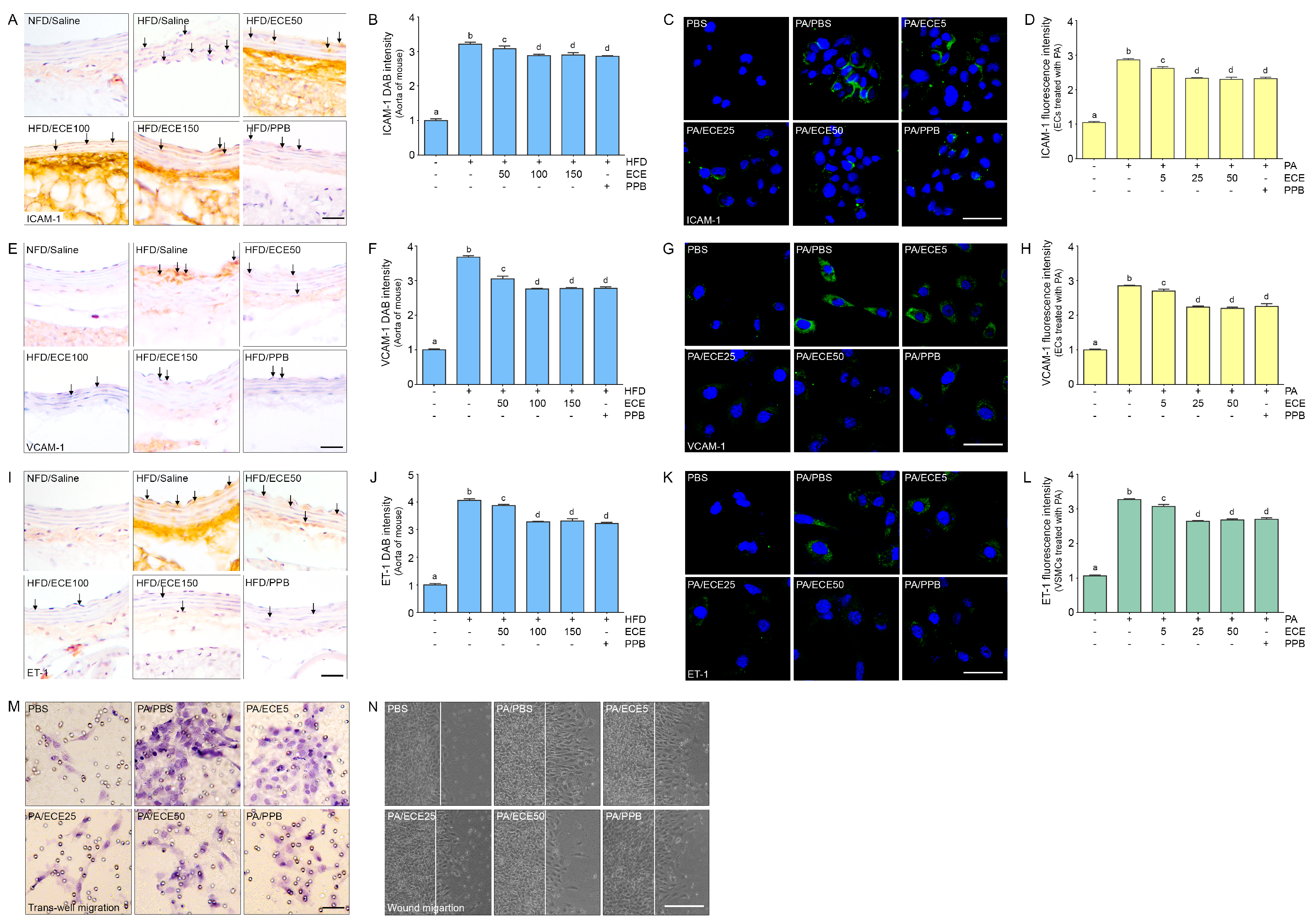
2.1. ECE and PPB Reduced the Expression of TLR4 Increased by HFD in the Aorta and in Palmitate-Treated ECs and VSMCs
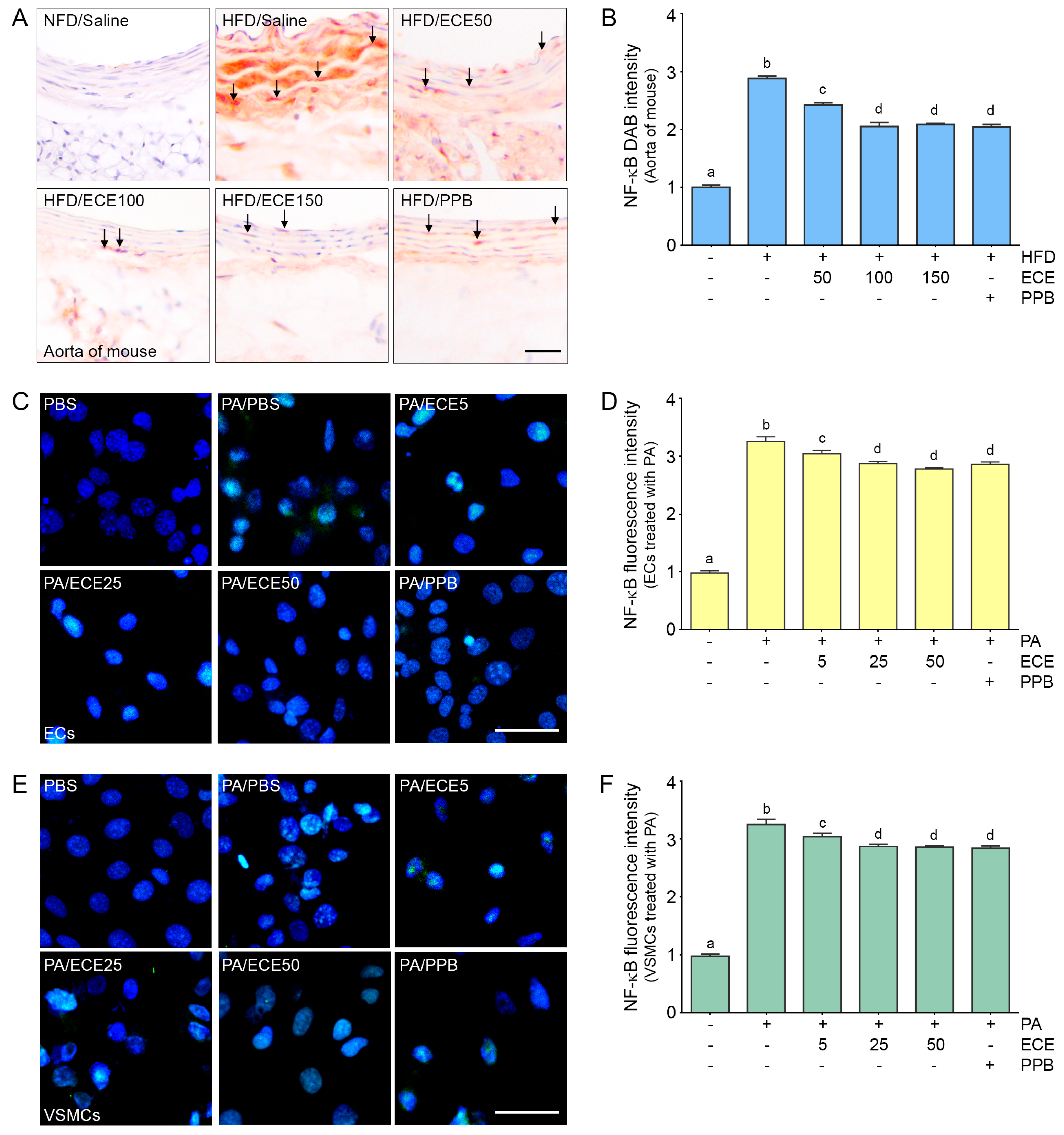
2.2. ECE and PPB Reduced the Expression of NF-κB Increased by HFD in the Aorta and in Palmitate-Treated ECs and VSMCs
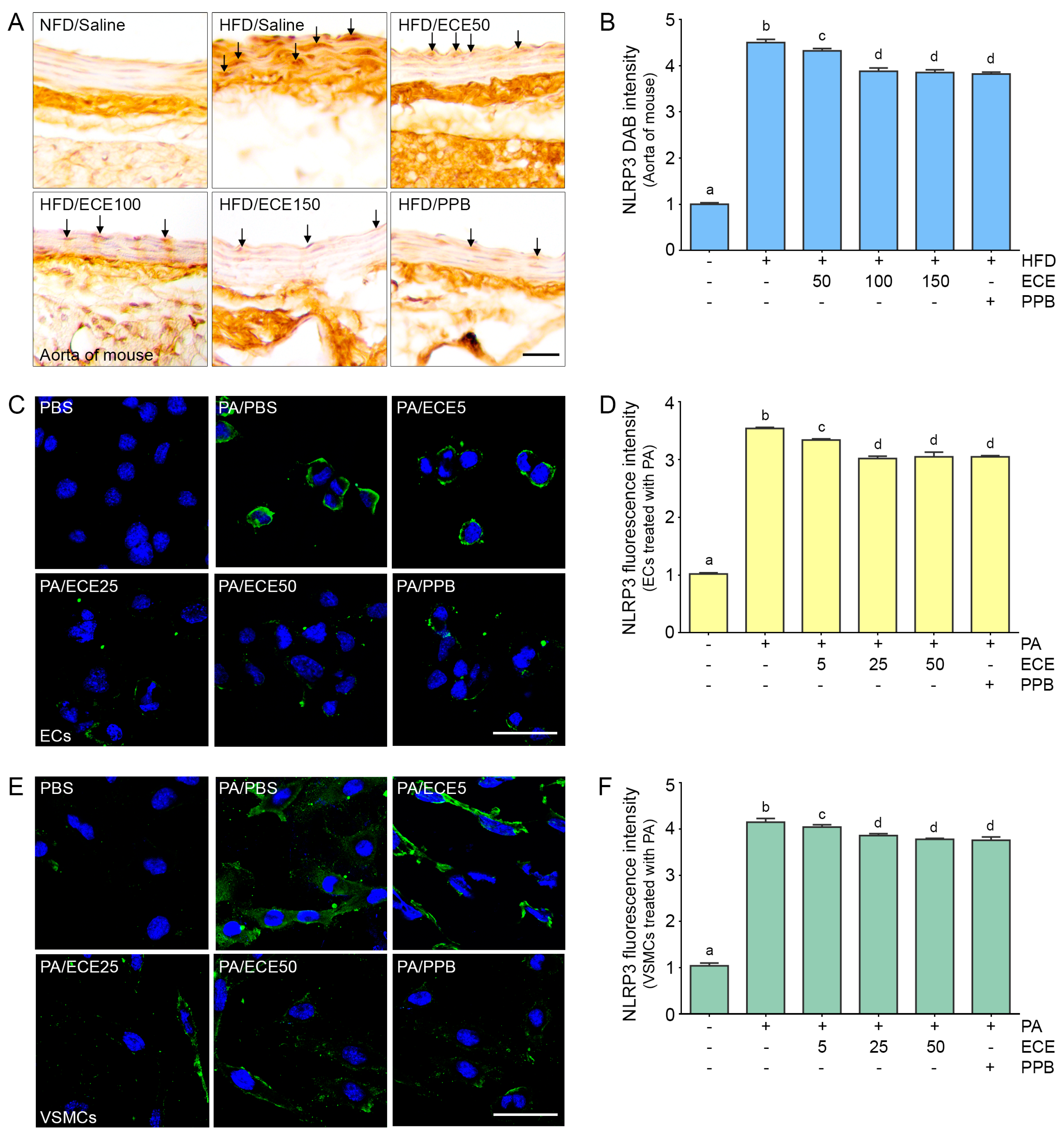
2.3. ECE and PPB Reduced the Expression of NLRP3 Increased by HFD in the Aorta and in the Palmitate-Treated ECs and VSMCs
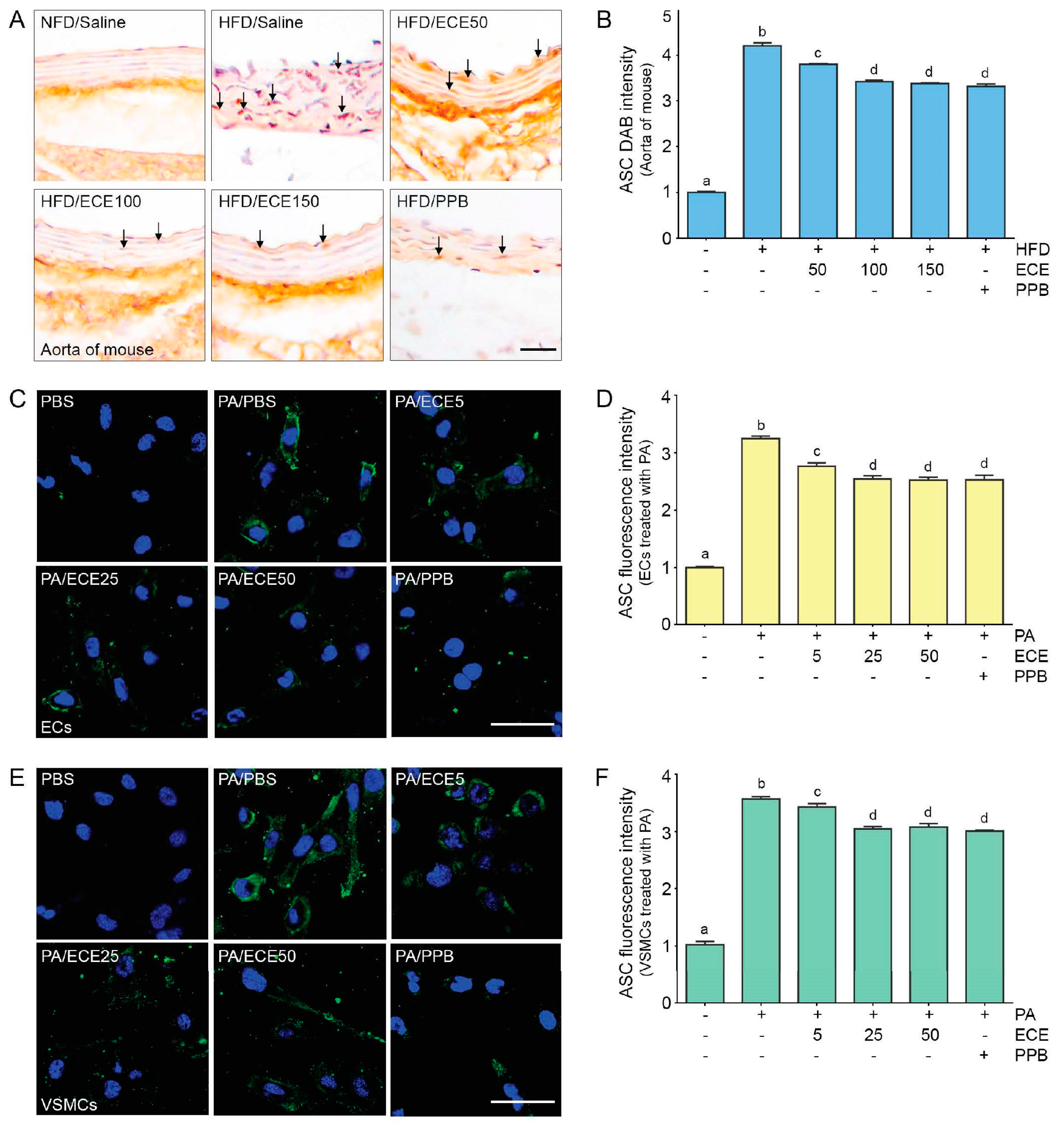
2.4. ECE and PPB Reduced the Expression of ASC, Which Was Increased by HFD in the Aorta and in Palmitate-Treated ECs and VSMCs
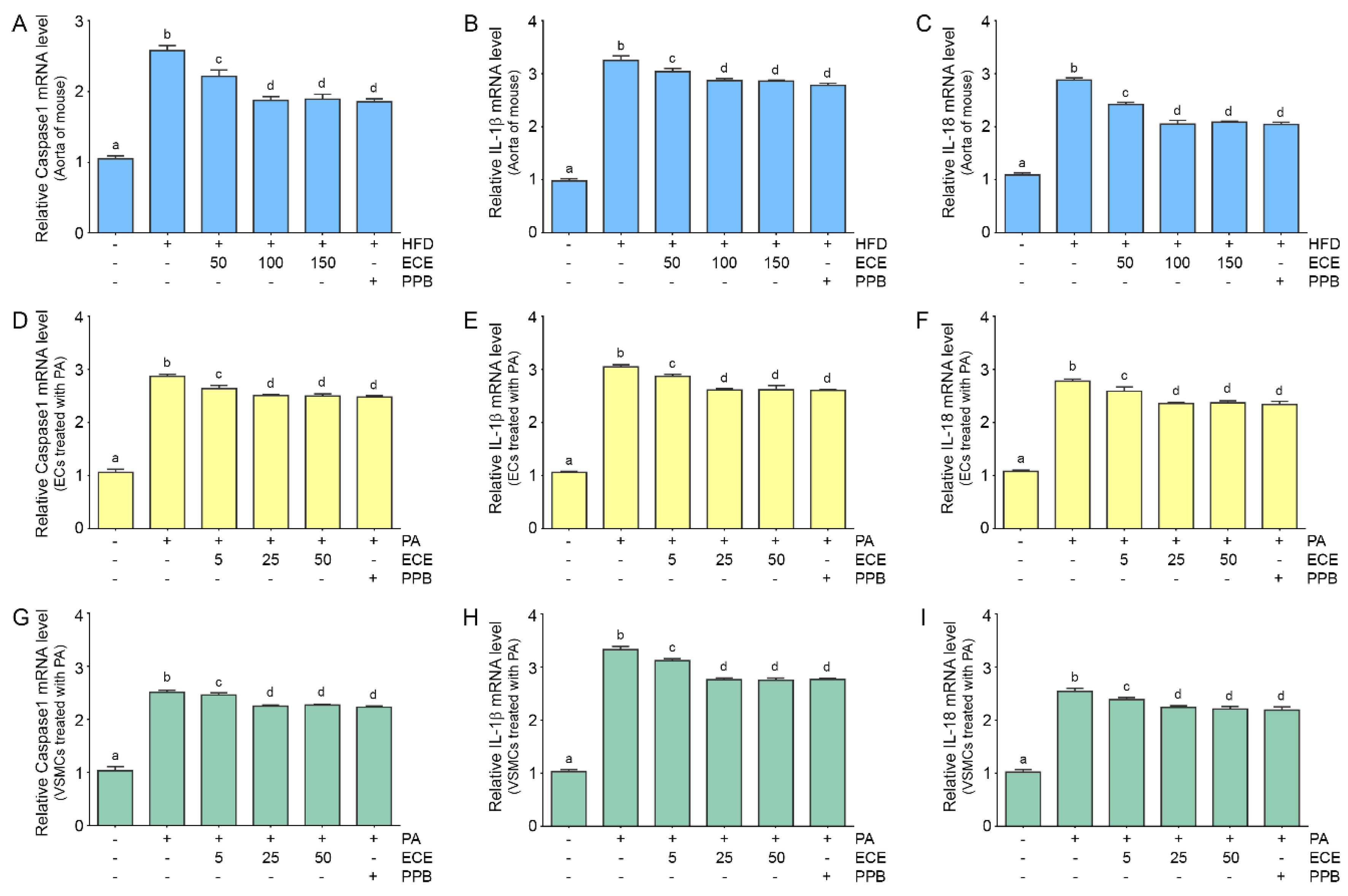
2.5. ECE and PPB Reduced Expressions of Caspase-1, IL-1β, and IL-18, Which Were Increased by HFD in the Aorta and in Palmitate-Treated ECs and VSMCs
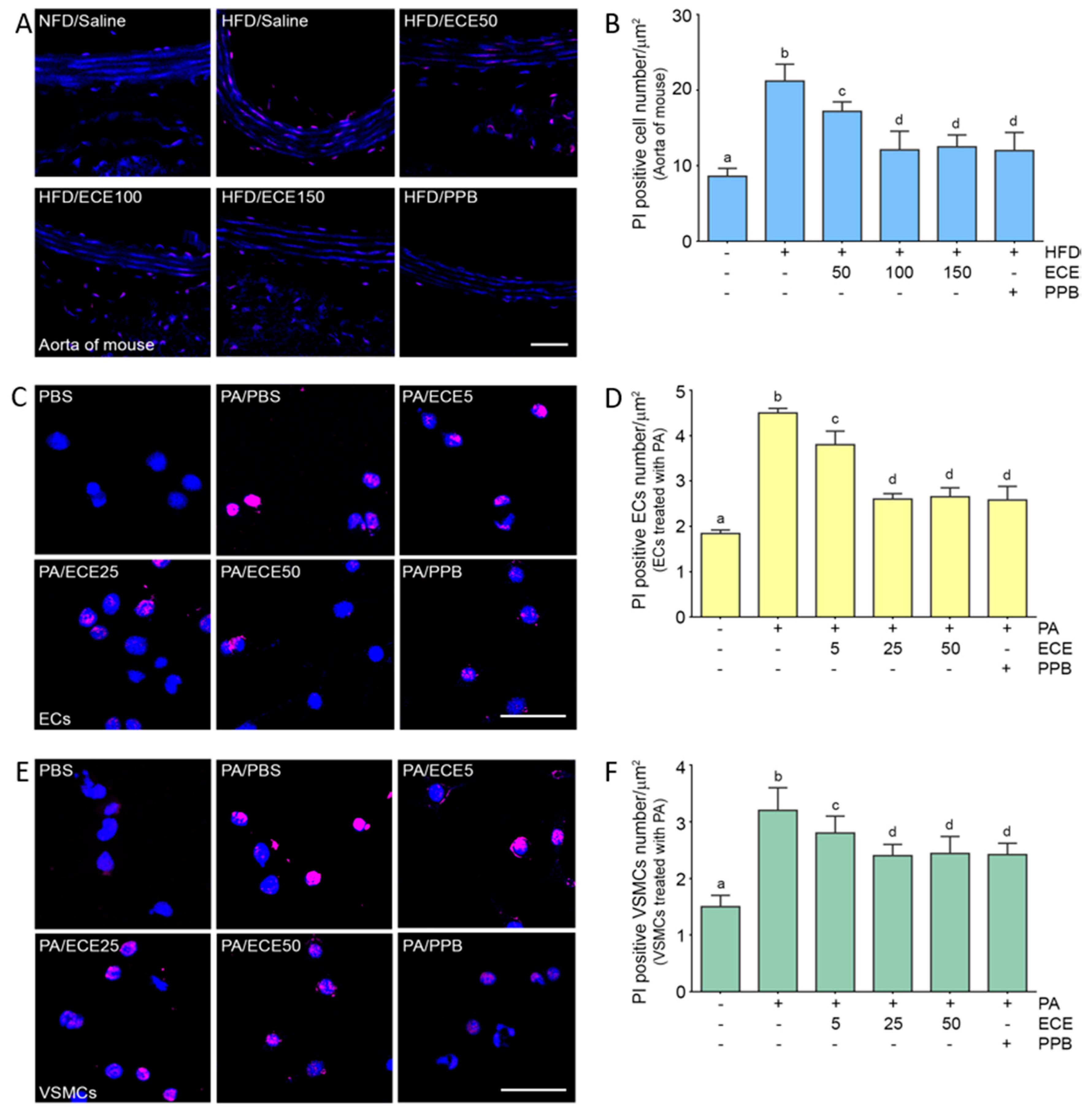
2.6. ECE and PPB Reduced Pyroptosis, Which Was Increased by HFD, in the Aorta and in Palmitate-Treated ECs and VSMCs
2.7. ECE and PPB Reduced Cell Dysfunction in the Aorta and in Palmitate-Treated ECs and VSMCs
3. Discussion
4. Materials and Methods
4.1. Ecklonia Cava Extract (ECE) and Pyrogallol-Phloroglucinol-6,6-Bieckol (PPB) Preparation
4.2. Diet Induced Obesity Animal Model
4.3. Cell Culture and Experimental Cell Models
4.4. RNA Extraction and cDNA Synthesis
4.5. Real-Time Reverse Transcription Polymerase Chain Reaction (qRT-PCR)
4.6. Immunohistochemistry (3,3-diaminobenzidine; DAB)
4.7. Immunocytochemistry
4.8. Trans-Well Migration Assay
4.9. Wound Migration Assay
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mestas, J.; Ley, K. Monocyte-Endothelial Cell Interactions in the Development of Atherosclerosis. Trends Cardiovasc. Med. 2008, 18, 228–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabas, I. Macrophage death and defective inflammation resolution in atherosclerosis. Nat. Rev. Immunol. 2010, 10, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.; Yan, C.; Zhang, L.; Li, Y.; Wan, Q. LncRNA ENST00113 promotes proliferation, survival, and migration by activating PI3K/Akt/mTOR signaling pathway in atherosclerosis. Medicine 2018, 97, e0473. [Google Scholar] [CrossRef]
- Savoia, C.; Schiffrin, E.L. Vascular inflammation in hypertension and diabetes: Molecular mechanisms and therapeutic interventions. Clin. Sci. 2007, 112, 375–384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ying, S.; Li, X.; Sha, X.; Xinyuan, L.; Li, Y.-F.; Shao, Y.; Mai, J.; Virtue, A.; Lopez-Pastrana, J.; Meng, S.; et al. Early Hyperlipidemia Promotes Endothelial Activation via a Caspase-1-Sirtuin 1 Pathway. Arter. Thromb. Vasc. Biol. 2015, 35, 804–816. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Yin, Y.; Wang, H. Vascular inflammation and atherogenesis are activated via receptors for PAMPs and suppressed by regulatory T cells. Drug Discov. Today Ther. Strat. 2008, 5, 125–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paone, S.; Baxter, A.A.; Hulett, M.D.; Poon, I.K.H. Endothelial cell apoptosis and the role of endothelial cell-derived extracellular vesicles in the progression of atherosclerosis. Cell. Mol. Life Sci. 2018, 76, 1093–1106. [Google Scholar] [CrossRef]
- Grebe, A.; Hoss, F.; Latz, E. NLRP3 Inflammasome and the IL-1 Pathway in Atherosclerosis. Circ. Res. 2018, 122, 1722–1740. [Google Scholar] [CrossRef]
- Hoseini, Z.; Sepahvand, F.; Rashidi, B.; Sahebkar, A.; Masoudifar, A.; Mirzaei, H. NLRP3 inflammasome: Its regulation and involvement in atherosclerosis. J. Cell. Physiol. 2018, 233, 2116–2132. [Google Scholar] [CrossRef] [PubMed]
- Zhaolin, Z.; Guohua, L.; Wu, S.; Wang, Z. Role of pyroptosis in cardiovascular disease. Cell Prolif. 2019, 52, e12563. [Google Scholar] [CrossRef] [Green Version]
- Sokolova, M.; Binge, L.E.; Alfsnes, K.; Olsen, M.B.; Eide, L.; KaasbΦll, O.J.; Attramadal, H.; Torp, M.K.; Fosshaug, L.E.; Rashidi, A.; et al. Palmitate promotes inflammatory responses and cellular senescence in cardiac fibroblasts. Biochim. Biophys. Acta Mol. Cell Biolology Lipids 2017, 1862, 234–245. [Google Scholar] [CrossRef]
- Yang, X.-F. Inflammasomes: Sensors of metabolic stresses for vascular inflammation. Front. Biosci. 2013, 18, 638. [Google Scholar] [CrossRef] [Green Version]
- Bauernfeind, F.G.; Horvath, G.; Stutz, A.; Alnemri, E.S.; MacDonald, K.; Speert, D.; Fernandes-Alnemri, T.; Wu, J.; Monks, B.G.; Fitzgerald, K.A.; et al. Cutting edge: NF-kB activating pattern recognition and cytokine receptors license NLRP3 inflammasome activation by regulating NLRP3 expression. J. Immunol. 2009, 183, 787–791. [Google Scholar] [CrossRef] [PubMed]
- Bauernfeind, F.; Bartok, E.; Rieger, A.; Franchi, L.; N´uñez, G.; Hornung, V. Cutting edge: Reactive oxygen species inhibitors block priming, but not activation, of the NLRP3 inflammasome. J. Immunol. 2011, 187, 613–617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afonina, I.S.; Zhong, Z.; Karin, M.; Beyaert, R. Limiting inflammation-the negative regulation of NF-κB and the NLRP3 inflammasome. Nat. Immunol. 2017, 18, 861–869. [Google Scholar] [CrossRef] [PubMed]
- Man, S.M.; Karki, R.; Kanneganti, T.-D. Molecular mechanisms and functions of pyroptosis, inflammatory caspases and inflammasomes in infectious diseases. Immunol. Rev. 2017, 277, 61–75. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Pastrana, J.; Ferrer, L.M.; Li, Y.F.; Xiong, X.; Xi, H.; Bueto, R.; Nelson, J.; Sha, X.; Li, X.; Cannella, A.L.; et al. Inhibition of caspase-1 activation in endothelial cells improves angiogenesis: A novel therapeutic potential for ischemia. J. Biol. Chem. 2015, 290, 17485–17494. [Google Scholar] [CrossRef] [Green Version]
- Son, M.; Oh, S.; Choi, J.; Jang, J.T.; Choi, C.H.; Park, K.Y.; Son, K.H.; Byun, K. The Phlorotannin-Rich Fraction of Ecklonia cava Extract Attenuated the Expressions of the Markers Related with Inflammation and Leptin Resistance in Adipose Tissue. Int. J. Endocrinol. 2020, 2020, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Son, M.; Oh, S.; Lee, H.S.; Chung, D.-M.; Jang, J.T.; Jeon, Y.-J.; Choi, C.H.; Park, K.Y.; Son, K.H.; Byun, K. Ecklonia Cava Extract Attenuates Endothelial Cell Dysfunction by Modulation of Inflammation and Brown Adipocyte Function in Perivascular Fat Tissue. Nutrients 2019, 11, 2795. [Google Scholar] [CrossRef] [Green Version]
- Oh, S.; Son, M.; Lee, H.S.; Kim, H.-S.; Jeon, Y.-J.; Byun, K. Protective Effect of Pyrogallol-Phloroglucinol-6,6-Bieckol from Ecklonia cava on Monocyte-Associated Vascular Dysfunction. Mar. Drugs 2018, 16, 441. [Google Scholar] [CrossRef] [Green Version]
- Geng, Y.; Ma, Q.; Liu, Y.-N.; Peng, N.; Yuan, F.-F.; Li, X.-G.; Li, M.; Wu, Y.-S.; Li, B.-L.; Song, W.-B.; et al. Heatstroke induces liver injury via IL-1β and HMGB1-induced pyroptosis. J. Hepatol. 2015, 63, 622–633. [Google Scholar] [CrossRef] [PubMed]
- Fernandesalnemri, T.; Wu, J.; Yu, J.-W.; Datta, P.K.; Miller, B.; Jankowski, W.; Rosenberg, S.A.; Zhang, J.; Alnemri, E.S. The pyroptosome: A supramolecular assembly of ASC dimers mediating inflammatory cell death via caspase-1 activation. Cell Death Differ. 2007, 14, 1590–1604. [Google Scholar] [CrossRef]
- Hu, Q.; Zhang, T.; Yi, L.; Zhou, X.; Mi, M. Dihydromyricetin inhibits NLRP3 inflammasome-dependent pyroptosis by activating the Nrf2 signaling pathway in vascular endothelial cells. BioFactors 2018, 44, 123–136. [Google Scholar] [CrossRef]
- Bochaton-Piallat, M.L.; Gabbiani, F.; Redard, M.; Desmoulière, A.; Gabbiani, G. Apoptosis participates in cellularity regulation during rat aortic intimal thickening. Am. J. Pathol. 1995, 146, 1059–1064. [Google Scholar] [PubMed]
- Anderson, T.J. Assessment and treatment of endothelial dysfunction in humans. J. Am. Coll. Cardiol. 1999, 34, 631–638. [Google Scholar] [CrossRef] [Green Version]
- Mensah, G.A. Healthy endothelium: The scientific basis for cardiovascular health promotion and chronic disease prevention. Vasc. Pharmacol. 2007, 46, 310–314. [Google Scholar] [CrossRef]
- Goligorsky, M.S. Endothelial cell dysfunction and nitric oxide synthase. Kidney Int. 2000, 58, 1360–1376. [Google Scholar] [CrossRef] [Green Version]
- Nakashima, Y.; Plump, A.S.; Raines, E.W.; Breslow, J.L.; Ross, R. ApoE-deficient mice develop lesions of all phases of atherosclerosis throughout the arterial tree. Arter. Thromb. A J. Vasc. Biol. 1994, 14, 133–140. [Google Scholar] [CrossRef] [Green Version]
- Komuro, I.; Kurihara, H.; Sugiyama, T.; Takaku, F.; Yazaki, Y. Endothelin stimulates c-fos and c-myc expression and proliferation of vascular smooth muscle cells. FEBS Lett. 1988, 238, 249–252. [Google Scholar] [CrossRef] [Green Version]
- Kohno, M.; Yokokawa, K.; Yasunari, K.; Kano, H.; Minami, M.; Yoshikawa, J. Effect of the Endothelin Family of Peptides on Human Coronary Artery Smooth-Muscle Cell Migration. J. Cardiovasc. Pharmacol. 1998, 31, S84–S89. [Google Scholar] [CrossRef] [PubMed]
- Ishida, N.; Tsujioka, K.; Tomoi, M.; Saida, K.; Mitsui, Y. Differential activities of two distinct endothelin family peptides on ileum and coronary artery. FEBS Lett. 1989, 247, 337–340. [Google Scholar] [CrossRef] [Green Version]
- Harris, A.K.; Hutchinson, J.R.; Sachidanandam, K.; Johnson, M.H.; Dorrance, A.M.; Stepp, D.W.; Fagan, S.C.; Ergul, A. Type 2 Diabetes Causes Remodeling of Cerebrovasculature via Differential Regulation of Matrix Metalloproteinases and Collagen Synthesis: Role of Endothelin-1. Diabetes 2005, 54, 2638–2644. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naito, S.; Shimizu, S.; Maeda, S.; Wang, J.; Paul, R.; Fagin, J.A. Ets-1 is an early response gene activated by ET-1 and PDGF-BB in vascular smooth muscle cells. Am. J. Physiol. Content 1998, 274, C472–C480. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Vita, J.; Ruiz-Ortega, M.; Rupérez, M.; Esteban, V.; Sanchez-Lopez, E.; Plaza, J.J.; Egido, J. Endothelin-1, via ETAReceptor and Independently of Transforming Growth Factor-β, Increases the Connective Tissue Growth Factor in Vascular Smooth Muscle Cells. Circ. Res. 2005, 97, 125–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lerman, A.; Webster, M.W.; Chesebro, J.H.; Edwards, W.D.; Wei, C.M.; Fuster, V.; Burnett, J.C., Jr. Circulating and tissue endothelin immunoreactivity in hypercholesterolemic pigs. Circulation 1993, 88, 2923–2928. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, T.; Miyauchi, T.; Iwasa, S.; Sakai, S.; Fan, J.; Nagata, M.; Goto, K.; Watanabe, T. Corresponding distributions of increased endothelin-B receptor expression and increased endothelin-1 expression in the aorta of apolipoprotein E-deficient mice with advanced atherosclerosis. Pathol. Int. 2000, 50, 929–936. [Google Scholar] [CrossRef]
- Iwasa, S.; Fan, J.; Shimokama, T.; Nagata, M.; Watanabe, T. Increased immunoreactivity of endothelin-1 and endothelin B receptor in human atherosclerotic lesions. A possible role in atherogenesis. Atherosclerosis 1999, 146, 93–100. [Google Scholar] [CrossRef]
- Winkles, J.; Alberts, G.; Brogi, E.; Libby, P. Endothelin-1 and Endothelin Receptor mRNA Expression in Normal and Atherosclerotic Human Arteries. Biochem. Biophys. Res. Commun. 1993, 191, 1081–1088. [Google Scholar] [CrossRef]
- Lerman, A.; Edwards, B.S.; Hallett, J.W.; Heublein, D.M.; Sandberg, S.M.; Burnett, J.C., Jr. Circulating and tissue endothelin immunoreactivity in advanced atherosclerosis. N. Engl. J. Med. 1991, 325, 997–1001. [Google Scholar] [CrossRef] [Green Version]
- Davies, M.G.; Hagen, P.O. Pathobiology of intimal hyperplasia. Br. J. Surg. 1994, 81, 1254–1269. [Google Scholar] [CrossRef]
- Son, M.; Oh, S.; Lee, H.S.; Ryu, B.; Jiang, Y.; Jang, J.T.; Jeon, Y.-J.; Byun, K. Pyrogallol-Phloroglucinol-6,6’-Bieckol from Ecklonia cava Improved Blood Circulation in Diet-Induced Obese and Diet-Induced Hypertension Mouse Models. Mar. Drugs 2019, 17, 272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, J.; Oh, S.; Son, M.; Byun, K. Pyrogallol-Phloroglucinol-6,6-Bieckol Alleviates Obesity and Systemic Inflammation in a Mouse Model by Reducing Expression of RAGE and RAGE Ligands. Mar. Drugs 2019, 17, 612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, S.; Son, M.; Choi, J.; Choi, C.H.; Park, K.Y.; Son, K.H.; Byun, K. Phlorotannins from Ecklonia cava Attenuates Palmitate-Induced Endoplasmic Reticulum Stress and Leptin Resistance in Hypothalamic Neurons. Mar. Drugs 2019, 17, 570. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, J.S.; Oh, A.R.; Lee, H.J.; Ahn, Y.H.; Cha, J.Y. Hepatic Elovl6 gene expression is regulated by the synergistic action of ChREBP and SREBP-1c. Biochem. Biophys. Res. Commun. 2016, 23, 478. [Google Scholar] [CrossRef]
- Kim, O.H.; Booth, C.J.; Choi, H.S.; Lee, J.; Kang, J.; Hur, J.; Jung, W.J.; Jung, Y.S.; Choi, H.J.; Kim, H.; et al. High-phytate/low-calcium diet is a risk factor for crystal nephropathies, renal phosphate wasting, and bone loss. eLife 2020, 9, 9. [Google Scholar] [CrossRef]
- Lee, E.H.; Itan, M.; Jang, J.; Gu, H.J.; Rozenberg, P.; Mingler, M.; Wen, T.; Yoon, J.; Park, S.Y.; Roh, J.Y.; et al. Eosinophils support adipocyte maturation and promote glucose tolerance in obesity. Sci. Rep. 2018, 2, 8. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oh, S.; Son, M.; Park, C.-H.; Jang, J.T.; Son, K.H.; Byun, K. The Reducing Effects of Pyrogallol-Phloroglucinol-6,6-Bieckol on High-Fat Diet-Induced Pyroptosis in Endothelial and Vascular Smooth Muscle Cells of Mice Aortas. Mar. Drugs 2020, 18, 648. https://doi.org/10.3390/md18120648
Oh S, Son M, Park C-H, Jang JT, Son KH, Byun K. The Reducing Effects of Pyrogallol-Phloroglucinol-6,6-Bieckol on High-Fat Diet-Induced Pyroptosis in Endothelial and Vascular Smooth Muscle Cells of Mice Aortas. Marine Drugs. 2020; 18(12):648. https://doi.org/10.3390/md18120648
Chicago/Turabian StyleOh, Seyeon, Myeongjoo Son, Chul-Hyun Park, Ji Tae Jang, Kuk Hui Son, and Kyunghee Byun. 2020. "The Reducing Effects of Pyrogallol-Phloroglucinol-6,6-Bieckol on High-Fat Diet-Induced Pyroptosis in Endothelial and Vascular Smooth Muscle Cells of Mice Aortas" Marine Drugs 18, no. 12: 648. https://doi.org/10.3390/md18120648
APA StyleOh, S., Son, M., Park, C. -H., Jang, J. T., Son, K. H., & Byun, K. (2020). The Reducing Effects of Pyrogallol-Phloroglucinol-6,6-Bieckol on High-Fat Diet-Induced Pyroptosis in Endothelial and Vascular Smooth Muscle Cells of Mice Aortas. Marine Drugs, 18(12), 648. https://doi.org/10.3390/md18120648