Mining the Metabolome and the Agricultural and Pharmaceutical Potential of Sea Foam-Derived Fungi

 , , and
, , and 
Abstract
:
1. Introduction
2. Results
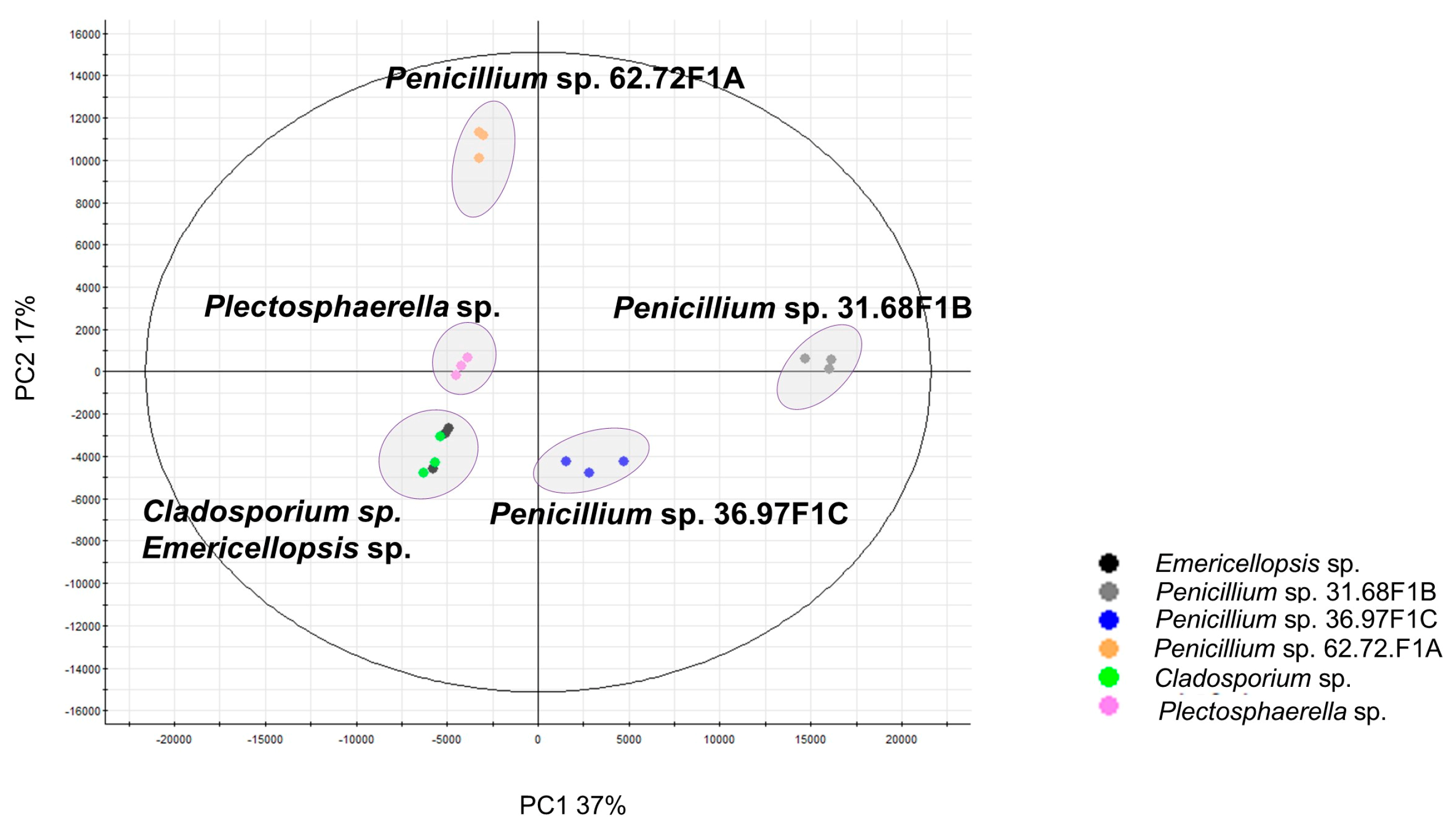
2.1. Selection of Isolates and Phylogenetic Analysis
2.2. Metabolomics
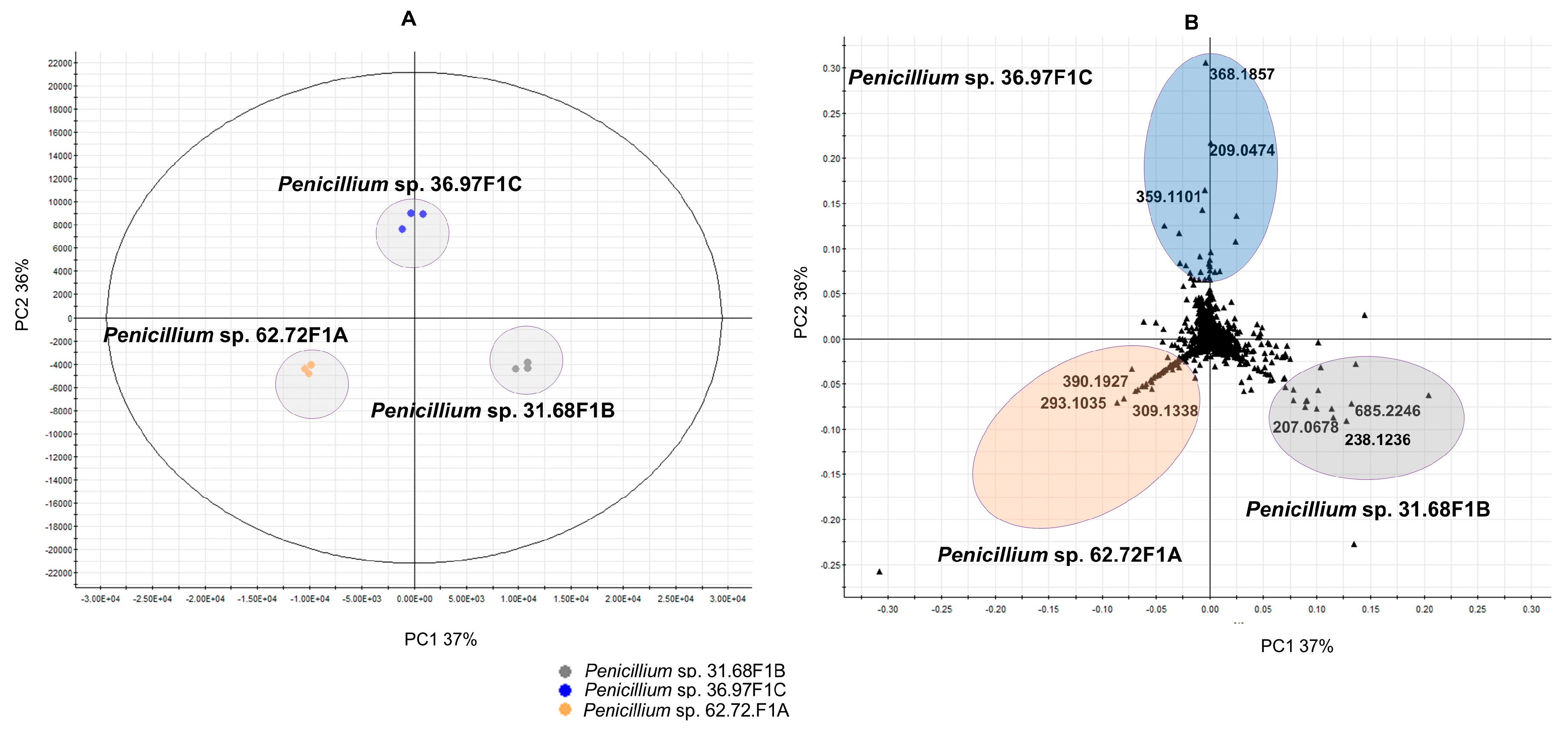
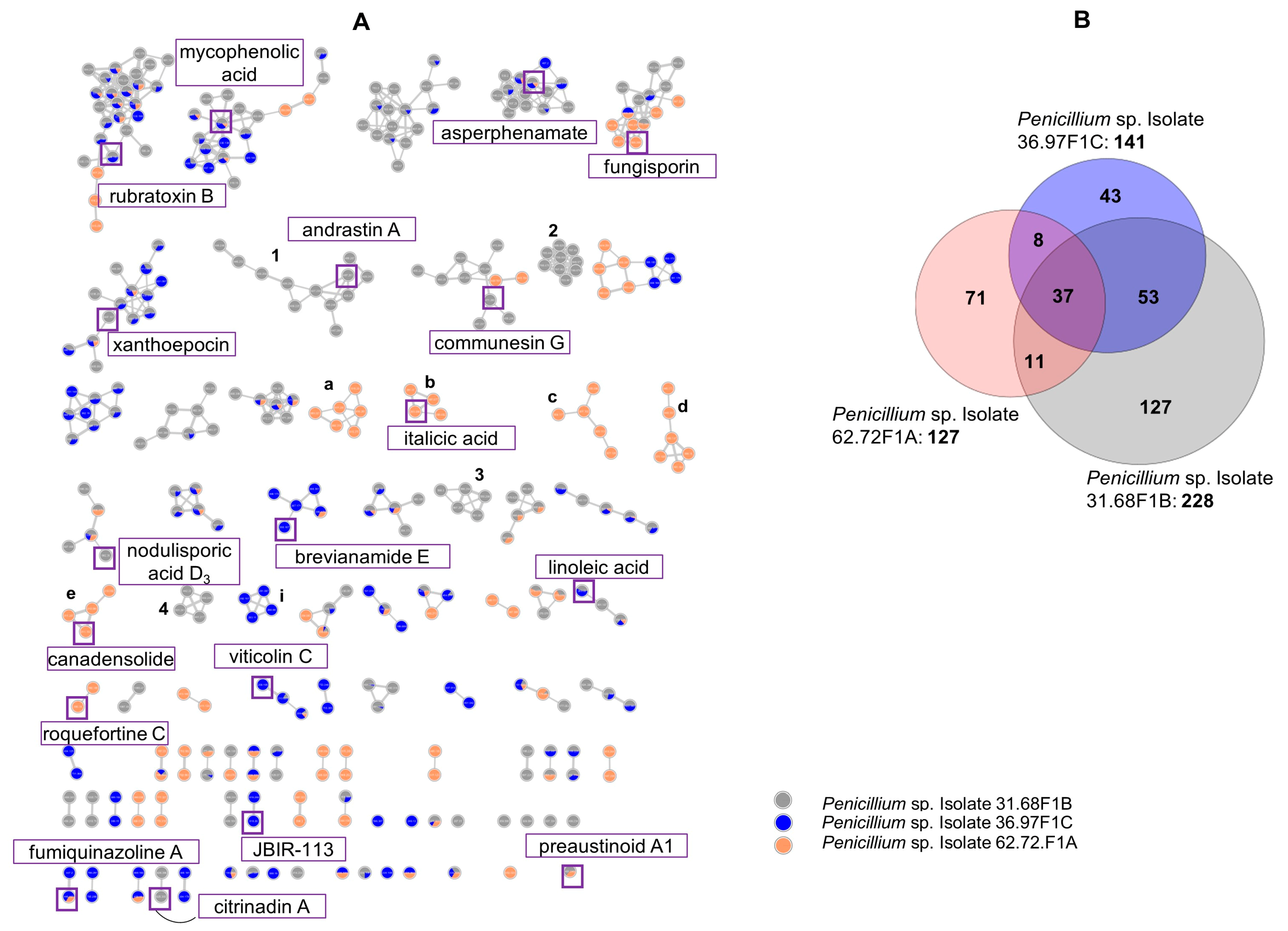
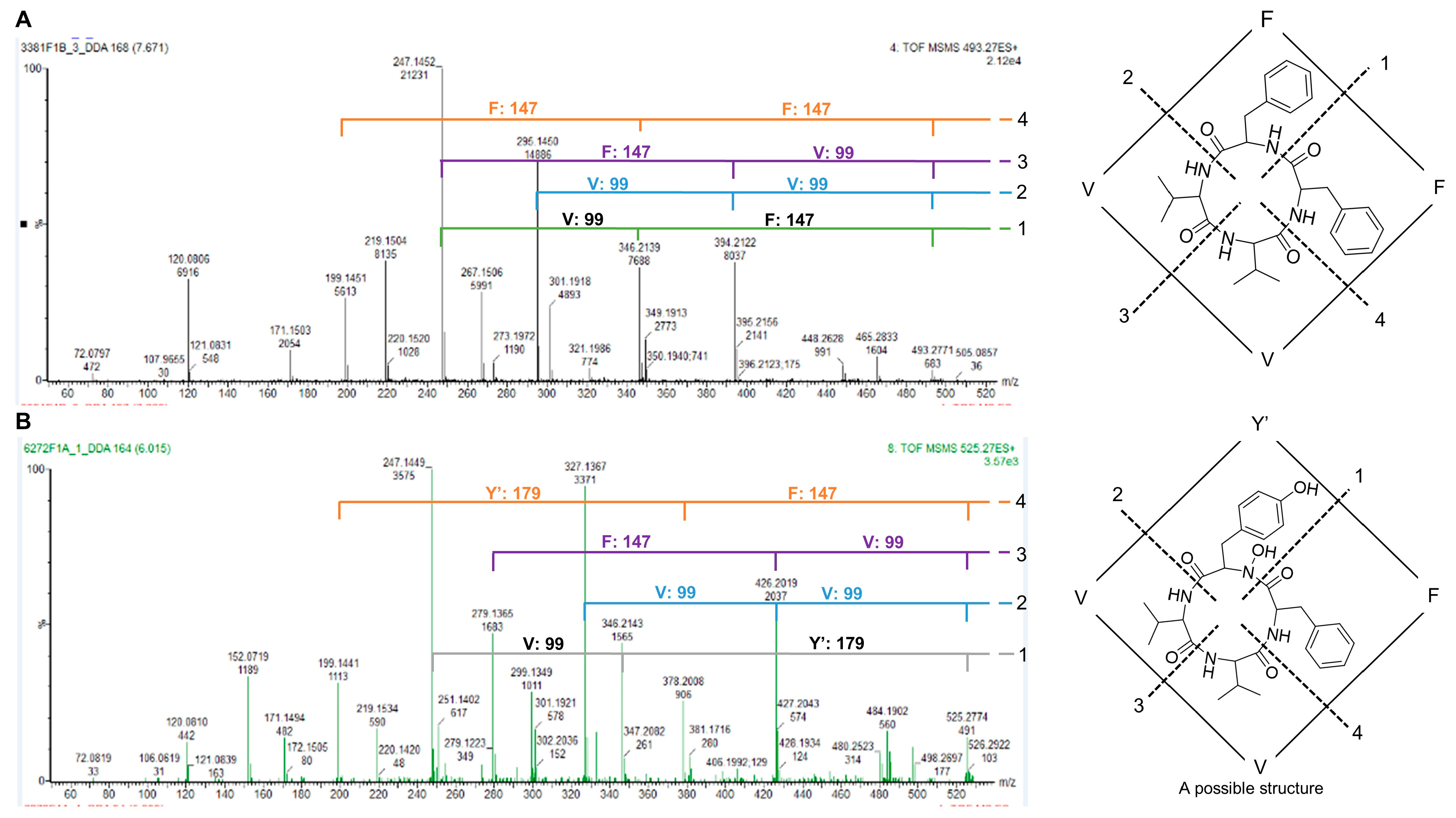
2.2.1. Molecular Network Analyses of the Penicillium Strains
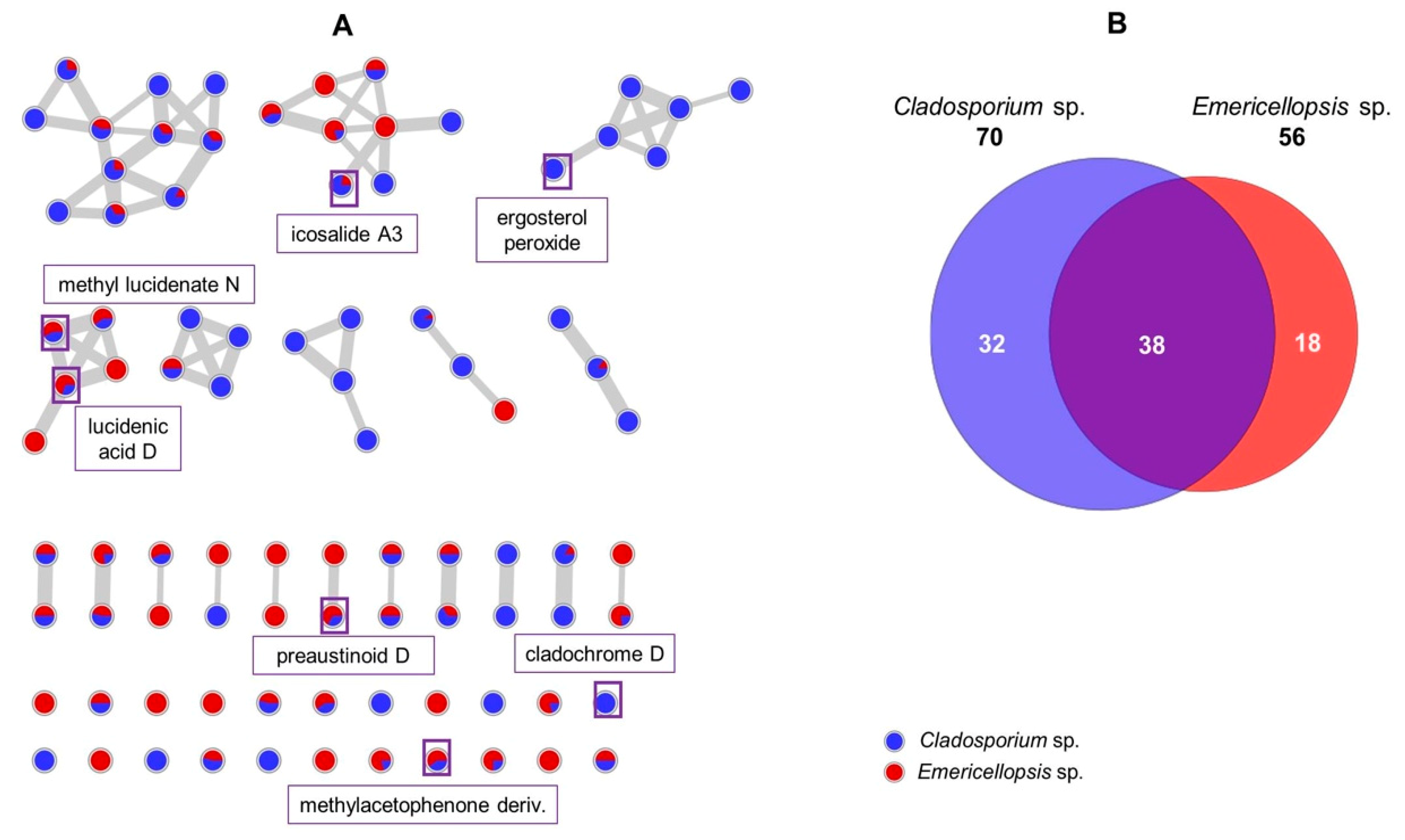
2.2.2. Molecular Network Analyses of the Cladosporium and Emericellopsis sp.
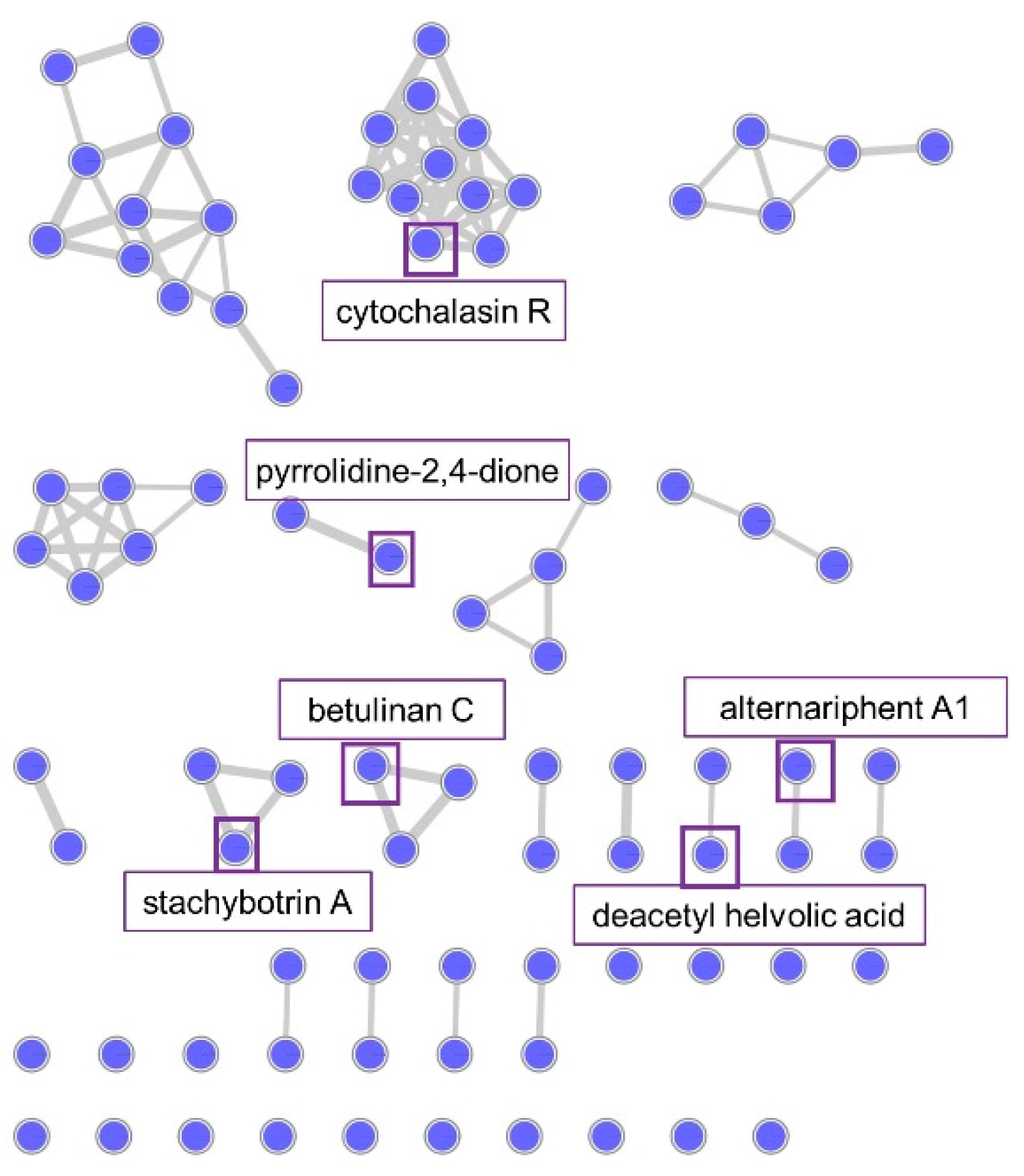
2.2.3. Molecular Network Analysis of Plectosphaerella sp.
2.3. Antimicrobial and Cytotoxic Activity
3. Discussion
4. Materials and Methods
4.1. Fungal Materials
4.2. Phylogenetic Analyses
4.3. Fermentation and Extraction
4.4. UPLC/ESI-QToF-MS/MS Analyses
4.5. Data Processing and Metabolomics
4.5.1. Molecular Networking
4.5.2. Statistical Analysis
4.6. Antimicrobial Activity
4.7. Anticancer Activity
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Koehn, R.D. Fungi isolated from sea foam collected at North Padre Island beaches. Southwest. Nat. 1982, 27, 17–21. [Google Scholar] [CrossRef]
- Wilson, M.I.; Robertson, L.D.; Daly, M.; Walton, S.A. Effects of visual cues on assessment of water quality. J. Environ. Psychol. 1995, 15, 53–63. [Google Scholar] [CrossRef]
- Schilling, K.; Zessner, M. Foam in the aquatic environment. Water Res. 2011, 45, 4355–4366. [Google Scholar] [CrossRef] [PubMed]
- Overy, D.P.; Berrue, F.; Correa, H.; Hanif, N.; Hay, K.; Lanteigne, M.; Mquilian, K.; Duffy, S.; Boland, P.; Jagannathan, R. Sea foam as a source of fungal inoculum for the isolation of biologically active natural products. Mycology 2014, 5, 130–144. [Google Scholar] [CrossRef]
- Eberlein, K.; Leal, M.T.; Hammer, K.D.; Hickel, W. Dissolved organic substances during a Phaeocystis pouchetii bloom in the German Bight (North Sea). Mar. Biol. 1985, 89, 311–316. [Google Scholar] [CrossRef]
- Ruzicka, K.; Gabriel, O.; Bletterie, U.; Winkler, S.; Zessner, M. Cause and effect relationship between foam formation and treated wastewater effluents in a transboundary river. Phys. Chem. Earth 2009, 34, 565–573. [Google Scholar] [CrossRef]
- Ettema, R.; Johnson, J.; Schaefer, J. Foam-initiated ice covers on small rivers and streams: an observation. Cold Reg. Sci. Technol. 1989, 16, 95–99. [Google Scholar] [CrossRef]
- Napolitano, G.E.; Richmond, J.E. Enrichment of biogenic lipids, hydrocarbons and PCBs in stream-surface foams. Environ. Toxicol. Chem. 1995, 14, 197–201. [Google Scholar] [CrossRef]
- Kirk Jr, P.W. Direct enumeration of marine arenicolous fungi. Mycologia 1983, 75, 670–682. [Google Scholar] [CrossRef]
- Kohlmeyer, J. Ecological observations on arenicolous marine fungi. Z. Allg. Mikrobiol. 1966, 6, 95–106. [Google Scholar] [CrossRef]
- Hill, C.; Eastoe, J. Foams: from nature to industry. Adv. Colloid Interface Sci. 2017, 247, 496–513. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oppong-Danquah, E.; Parrot, D.; Blümel, M.; Labes, A.; Tasdemir, D. Molecular networking-based metabolome and bioactivity analyses of marine-adapted fungi co-cultivated with phytopathogens. Front. Microbiol. 2018, 9, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Zhang, X.; Nong, X.; Wang, J.; Qi, S. Brevianamides and mycophenolic acid derivatives from the deep-Sea-derived fungus Penicillium brevicompactum DFFSCS025. Mar. Drugs 2017, 15, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ali, H.; Ries, M.I.; Lankhorst, P.P.; van der Hoeven, R.A.M.; Schouten, O.L.; Noga, M.; Hankemeier, T.; van Peij, N.N.M.E.; Bovenberg, R.A.L.; Vreeken, R.J.; et al. A non-canonical NRPS is involved in the synthesis of fungisporin and related hydrophobic cyclic tetrapeptides in Penicillium chrysogenum. PLoS ONE 2014, 9, e98212. [Google Scholar] [CrossRef]
- Nielsen, K.F.; Dalsgaard, P.W.; Smedsgaard, J.; Larsen, T.O. Andrastins A−D, Penicillium roqueforti metabolites consistently produced in blue-mold-ripened cheese. J. Agric. Food Chem. 2005, 53, 2908–2913. [Google Scholar] [CrossRef]
- dos Santos, R.M.G.; Rodrigues-Fo, E. Further meroterpenes produced by Penicillium sp., an endophyte obtained from Melia azedarach. Z. Naturforsch. C 2003, 58, 663–669. [Google Scholar] [CrossRef]
- Arai, K.; Miyajima, H.; Mushiroda, T.; Yamamoto, Y. Metabolites of Penicillium italicum WEHMER: isolation and structures of new metabolites including naturally occurring 4-ylidene-acyltetronic acids, italicinic acid and italicic acid. Chem. Pharm. Bull. 1989, 37, 3229–3235. [Google Scholar] [CrossRef] [Green Version]
- Igarashi, Y.; Kuwamori, Y.; Takagi, K.; Ando, T.; Fudou, R.; Furumai, T.; Oki, T. Xanthoepocin, a new antibiotic from Penicillium simplicissimum IFO5762. J. Antibiot. 2000, 53, 928–933. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.-D.; Yan, Z.; Zou, J.-H.; Wang, N.; Dai, J.-G. Rubratoxin C, a new nonadride derivative from an endophytic fungus Penicillium sp. F-14. Chin. Chem. Lett. 2014, 25, 1308–1310. [Google Scholar] [CrossRef]
- Sakata, K.; Masago, H.; Sakurai, A.; Takahashi, N. Isolation of aspirochlorine (=antibiotic A30641) possessing a novel dithiodiketopiperazine structure from Aspergillus flavus. Tetrahedron Lett. 1982, 23, 2095–2098. [Google Scholar] [CrossRef]
- Birch, A.J.; Wright, J.J. Studies in relation to biosynthesis—XLII: the structural elucidation and some aspects of the biosynthesis of the brevianamides-A and -E. Tetrahedron 1970, 26, 2329–2344. [Google Scholar] [CrossRef]
- Dalsgaard, P.W.; Blunt, J.W.; Munro, M.H.; Frisvad, J.C.; Christophersen, C. Communesins G and H, new alkaloids from the psychrotolerant fungus Penicillium rivulum. J. Nat. Prod. 2005, 68, 258–261. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liu, R.; Yang, J.; Li, H.; Zhou, F. Bioactive alkaloids of Aspergillus fumigatus, an endophytic fungus from Astragalus membranaceus. Chem. Nat. Compd. 2017, 53, 802–805. [Google Scholar] [CrossRef]
- Du, L.; Li, D.; Zhu, T.; Cai, S.; Wang, F.; Xiao, X.; Gu, Q. New alkaloids and diterpenes from a deep ocean sediment derived fungus Penicillium sp. Tetrahedron 2009, 65, 1033–1039. [Google Scholar] [CrossRef]
- Liu, Z.-G.; Bao, L.; Liu, H.-W.; Ren, J.-W.; Wang, W.-Z.; Wang, L.; Li, W.; Yin, W.-B. Chemical diversity from the Tibetan Plateau fungi Penicillium kongii and P. brasilianum. Mycology 2018, 9, 10–19. [Google Scholar] [CrossRef] [Green Version]
- Buckingham, J. Dictionary of Natural Products (on-line web edition). Available online: http://www.chemnetbase.com (accessed on 20 January 2020).
- Tsuda, M.; Kasai, Y.; Komatsu, K.; Sone, T.; Tanaka, M.; Mikami, Y.; Kobayashi, J.i. Citrinadin A, a novel pentacyclic alkaloid from marine-derived fungus Penicillium citrinum. Org. Lett. 2004, 6, 3087–3089. [Google Scholar] [CrossRef]
- Kawahara, T.; Takagi, M.; Shin-ya, K. Three new depsipeptides, JBIR-113, JBIR-114 and JBIR-115, isolated from a marine sponge-derived Penicillium sp. fS36. J. Antibiot. 2011, 65, 147–150. [Google Scholar] [CrossRef] [Green Version]
- Harrison, A.G.; Young, A.B.; Bleiholder, C.; Suhai, S.; Paizs, B. Scrambling of sequence information in collision-induced dissociation of peptides. J. Am. Chem. Soc. 2006, 128, 10364–10365. [Google Scholar] [CrossRef]
- Liu, W.-T.; Ng, J.; Meluzzi, D.; Bandeira, N.; Gutierrez, M.; Simmons, T.L.; Schultz, A.W.; Linington, R.G.; Moore, B.S.; Gerwick, W.H.; et al. Interpretation of tandem mass spectra obtained from cyclic nonribosomal peptides. Anal. Chem. 2009, 81, 4200–4209. [Google Scholar] [CrossRef] [Green Version]
- Bok, J.W.; Lermer, L.; Chilton, J.; Klingeman, H.G.; Towers, G.H.N. Antitumor sterols from the mycelia of Cordyceps sinensis. Phytochemistry 1999, 51, 891–898. [Google Scholar] [CrossRef] [Green Version]
- Arnone, A.; Assante, G.; Merlini, L.; Nasini, G. Structure and stereochemistry of cladochrome D and E, novel perylenequinone pigments from Cladosporium cladosporioides. Gazz. Chim. Ital. 1989, 119, 557–559. [Google Scholar] [CrossRef]
- Prabpai, S.; Wiyakrutta, S.; Sriubolmas, N.; Kongsaeree, P. Antimycobacterial dihydronaphthalenone from the endophytic fungus Nodulisporium sp. of Antidesma ghaesembilla. Phytochem. Lett. 2015, 13, 375–378. [Google Scholar] [CrossRef]
- Duan, R.; Zhou, H.; Yang, Y.; Li, H.; Dong, J.; Li, X.; Chen, G.; Zhao, L.; Ding, Z. Antimicrobial meroterpenoids from the endophytic fungus Penicillium sp. T2-8 associated with Gastrodia elata. Phytochem. Lett. 2016, 18, 197–201. [Google Scholar] [CrossRef]
- Kikuchi, T.; Matsuda, S.; Kadota, S.; Murai, Y.; Ogita, Z. Ganoderic acid D, E, F, and H and lucidenic acid D, E, and F, new triterpenoids from Ganoderma lucidum. Chem. Pharm. Bull. 1985, 33, 2624–2627. [Google Scholar] [CrossRef] [Green Version]
- Izawa, Y.; Hirose, T.; Shimizu, T.; Koyama, K.; Natori, S. Six new 10-pheynl-[11]cytochalasans, cytochalasins N - S from Phomopsis sp. Tetrahedron 1989, 45, 2323–2335. [Google Scholar] [CrossRef]
- Liu, K.; Wang, J.-L.; Wu, H.-B.; Wang, Q.; Bi, K.-L.; Song, Y.-F. A new pyranone from Lenzites betulina. Chem. Nat. Compd. 2012, 48, 780–781. [Google Scholar] [CrossRef]
- Lebrun, M.H.; Dutfoy, F.; Gaudemer, F.; Kunesch, G.; Gaudemer, A. Detection and quantification of the fungal phytotoxin tenuazonic acid produced by Pyricularia oryzae. Phytochemistry 1990, 29, 3777–3783. [Google Scholar] [CrossRef]
- Kamalov, L.S.; Aripova, S.F.; Isaev, M.I. Low-molecular-mass metabolites of fungi IV. the structures of stachybotrin A and stachybotral. Chem. Nat. Compd. 1999, 35, 82–85. [Google Scholar] [CrossRef]
- Chen, Y.; Chen, R.; Xu, J.; Tian, Y.; Xu, J.; Liu, Y. Two new altenusin/thiazole hybrids and a new benzothiazole derivative from the marine sponge-derived fungus Alternaria sp. SCSIOS02F49. Molecules 2018, 23, 2844. [Google Scholar] [CrossRef] [Green Version]
- Pang, X.; Lin, X.; Wang, P.; Zhou, X.; Yang, B.; Wang, J.; Liu, Y. Perylenequione derivatives with anticancer activities isolated from the marine sponge-derived fungus, Alternaria sp. SCSIO41014. Mar. Drugs 2018, 16, 280. [Google Scholar] [CrossRef] [Green Version]
- Kong, F.-D.; Huang, X.-L.; Ma, Q.-Y.; Xie, Q.-Y.; Wang, P.; Chen, P.-W.; Zhou, L.-M.; Yuan, J.-Z.; Dai, H.-F.; Luo, D.-Q.; et al. Helvolic acid derivatives with antibacterial activities against Streptococcus agalactiae from the marine-derived fungus Aspergillus fumigatus HNMF0047. J. Nat. Prod. 2018, 81, 1869–1876. [Google Scholar] [CrossRef] [PubMed]
- Fan, B.; Parrot, D.; Blümel, M.; Labes, A.; Tasdemir, D. Influence of OSMAC-based cultivation in metabolome and Anticancer activity of fungi associated with the brown Alga Fucus vesiculosus. Mar. Drugs 2019, 17, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vieira, G.; Purić, J.; Morão, L.; Dos Santos, J.; Inforsato, F.; Sette, L.; Ferreira, H.; Sass, D. Terrestrial and marine Antarctic fungi extracts active against Xanthomonas citri subsp. citri. Lett. Appl. Microbiol. 2018, 67, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Su, L.; Deng, H.; Niu, Y.-C. Phylogenetic analysis of Plectosphaerella species based on multi-locus DNA sequences and description of P. sinensis sp. nov. Mycol. Progress 2017, 16, 823–829. [Google Scholar] [CrossRef]
- Carr, G.; Tay, W.; Bottriell, H.; Andersen, S.K.; Mauk, A.G.; Andersen, R.J. Plectosphaeroic acids A, B, and C, Indoleamine 2,3-dioxygenase inhibitors produced in culture by a marine isolate of the fungus Plectosphaerella cucumerina. Org. Lett. 2009, 11, 2996–2999. [Google Scholar] [CrossRef]
- Overy, D.P.; Rämä, T.; Oosterhuis, R.; Walker, A.K.; Pang, K.-L. The neglected marine fungi, Sensu stricto, and their isolation for natural products’ discovery. Mar. Drugs 2019, 17, 42. [Google Scholar] [CrossRef] [Green Version]
- Bertrand, S.; Schumpp, O.; Bohni, N.; Bujard, A.; Azzollini, A.; Monod, M.; Gindro, K.; Wolfender, J.L. Detection of metabolite induction in fungal co-cultures on solid media by high-throughput differential ultra-high pressure liquid chromatography-time-of-flight mass spectrometry fingerprinting. J. Chromatogr. A 2013, 1292, 219–228. [Google Scholar] [CrossRef]
- Brondz, I.; Olsen, I. Microbial chemotaxonomy: chromatography, electrophoresis and relevant profiling techniques. J. Chromatogr. B Biomed. Sci. Appl. 1986, 379, 367–411. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Andersen, B.; Thrane, U. The use of secondary metabolite profiling in chemotaxonomy of filamentous fungi. Mycol. Res. 2008, 112, 231–240. [Google Scholar] [CrossRef]
- Yang, J.Y.; Sanchez, L.M.; Rath, C.M.; Liu, X.; Boudreau, P.D.; Bruns, N.; Glukhov, E.; Wodtke, A.; de Felicio, R.; Fenner, A.; et al. Molecular networking as a dereplication strategy. J. Nat. Prod. 2013, 76, 1686–1699. [Google Scholar] [CrossRef] [Green Version]
- da Silva, R.R.; Dorrestein, P.C.; Quinn, R.A. Illuminating the dark matter in metabolomics. PNAS 2015, 112, 12549–12550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratnaweera, P.B.; Williams, D.E.; de Silva, E.D.; Andersen, R.J. Antibacterial metabolites from the Sri Lankan demosponge-derived fungus, Aspergillus flavipes. Curr. Sci. 2016, 111, 1473–1479. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Houbraken, J.; Popma, S.; Samson, R.A. Two new Penicillium species Penicillium buchwaldii and Penicillium spathulatum, producing the anticancer compound asperphenamate. FEMS Microbiol. Lett. 2013, 339, 77–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, W.; Fan, A.; Wang, L.; Zhang, P.; Liu, Z.; An, Z.; Yin, W.-B. Asperphenamate biosynthesis reveals a novel two-module NRPS system to synthesize amino acid esters in fungi. Chem. Sci. 2018, 9, 2589–2594. [Google Scholar] [CrossRef] [Green Version]
- Bentley, R. Mycophenolic acid: A one hundred year odyssey from antibiotic to immunosuppressant. Chem. Rev. 2000, 100, 3801–3826. [Google Scholar] [CrossRef]
- Santini, A.; Mikušová, P.; Sulyok, M.; Krska, R.; Labuda, R.; Šrobárová, A. Penicillium strains isolated from Slovak grape berries taxonomy assessment by secondary metabolite profile. Mycotoxin Res. 2014, 30, 213–220. [Google Scholar] [CrossRef]
- Kim, H.Y.; Heo, D.Y.; Park, H.M.; Singh, D.; Lee, C.H. Metabolomic and transcriptomic comparison of solid-state and submerged fermentation of Penicillium expansum KACC 40815. PLoS ONE 2016, 11, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Rhee, Y.-H.; Jeong, S.-J.; Lee, H.-J.; Lee, H.-J.; Koh, W.; Jung, J.H.; Kim, S.-H.; Sung-Hoon, K. Inhibition of STAT3 signaling and induction of SHP1 mediate antiangiogenic and antitumor activities of ergosterol peroxide in U266 multiple myeloma cells. BMC Cancer 2012, 12, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.L.; Zhang, J.; Jiang, N.; Lu, Y.H.; Wang, L.; Xu, S.H.; Wang, W.; Zhang, G.F.; Xu, Q.; Ge, H.M.; et al. Immunosuppressive Polyketides from Mantis-associated Daldinia eschscholzii. J. Am. Chem. Soc. 2011, 133, 5931–5940. [Google Scholar] [CrossRef]
- Romano, S.; Jackson, A.S.; Patry, S.; Dobson, A.D.W. Extending the “One Strain Many Compounds” (OSMAC) principle to marine microorganisms. Mar. Drugs 2018, 16, 244. [Google Scholar] [CrossRef] [Green Version]
- Purves, K.; Macintyre, L.; Brennan, D.; Hreggviðsson, G.Ó.; Kuttner, E.; Ásgeirsdóttir, M.E.; Young, L.C.; Green, D.H.; Edrada-Ebel, R.; Duncan, K.R. Using molecular networking for microbial secondary metabolite bioprospecting. Metabolites 2016, 6, 2. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, L.; Ding, W.; Ma, Z. Induced production of cytochalasans in co-culture of marine fungus Aspergillus flavipes and actinomycete Streptomyces sp. Nat. Prod. Res. 2016, 30, 1718–1723. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Liu, Y.T.; Song, B.; Zhang, H.W.; Ding, G.; Liu, X.Z.; Zou, Z.M. Stereochemical determination of new cytochalasans from the plant endophytic fungus Trichoderma gamsii. Fitoterapia 2014, 96, 115–122. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.-A.; Ma, Y.-M.; Zhang, H.-C.; Liu, R. A new helvolic acid derivative from an endophytic Fusarium sp. of Ficus carica. Nat. Prod. Res. 2016, 30, 2407–2412. [Google Scholar] [CrossRef]
- Hall, T.; Biosciences, I.; Carlsbad, C. BioEdit: An important software for molecular biology. GERF Bull. Biosci. 2011, 2, 60–61. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Allen, F.; Pon, A.; Wilson, M.; Greiner, R.; Wishart, D. CFM-ID: a web server for annotation, spectrum prediction and metabolite identification from tandem mass spectra. Nucleic Acids Res. 2014, 42, 94–99. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Isolate | Fungal ID | Accession Number |
|---|---|---|---|
| 1 | B2F1B | Plectosphaerella sp. | MH791266.1 |
| 2 | 86F1C | Cladosporium sp. | MH791259.1 |
| 3 | 25.88F1C | Emericellopsis sp. | MH791280.1 |
| 4 | 36.97F1C | Penicillium sp. | MH791183.1 |
| 5 | 31.68F1B | Penicillium sp. | MH791236.1 |
| 6 | 62.72F1A | Penicillium sp. | MH791282.1 |
| Source | tR (min) | m/z | Mol. formula for M | [ppm] difference | Identification |
|---|---|---|---|---|---|
| Penicillium sp. 31.68F1B | 6 | 207.0678 [M + H]+ | C11H10O4 | 0.0 | 7-hydroxy-2-(hydroxymethyl)-5-methyl-4H-1-benzopyran-4-one |
| 8.2 | 238.1236 [M + H]+ | C16H15NO | 2.5 | n.d. | |
| 6 | 685.2246 [2M + H]+ | C13H18N4O7 | 0.6 | n.d. | |
| Penicillium sp. 36.97F1C | 2.5 | 209.0474 [M + H]+ | C10H8O5 | 1.9 | viticolin C |
| 4.8 | 368.1857 [M + H]+ | C21H25N3O3 | 0.5 | brevianamide E | |
| 4.7 | 359.1101 [M + Na]+ | C17H20O7 | 1.7 | 4-hydroxymycophenolic acid | |
| Penicillium sp. 62.72F1A | 3.8 | 309.1338 [M − H2O + H]+ | C16H22O7 | 1.5 | n.d. |
| 5 | 390.1927 [M + H]+ | C22H23N5O2 | 0.5 | tryhistatin | |
| 5.7 | 293.1035 [M + H]+ | C15H16O6 | 5.0 | italicic acid |
| Fungus | Plant Pathogens | Human Pathogens | |||||
|---|---|---|---|---|---|---|---|
| Pi | Mo | Bc | Efm | MRSA | Ca | Cn | |
| 31.68F1B | 4.3 | 2.2 | 13.8 | 15.0 | 31.0 | 6.3 | 9.2 |
| 36.97F1C | 31.6 | 12.9 | >100 | 37.8 | >100 | >100 | 26.9 |
| Positive control | 0.1 | 0.4 | 0.1 | 1.7 | 0.5 | 1.1 | 0.6 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oppong-Danquah, E.; Passaretti, C.; Chianese, O.; Blümel, M.; Tasdemir, D. Mining the Metabolome and the Agricultural and Pharmaceutical Potential of Sea Foam-Derived Fungi. Mar. Drugs 2020, 18, 128. https://doi.org/10.3390/md18020128
Oppong-Danquah E, Passaretti C, Chianese O, Blümel M, Tasdemir D. Mining the Metabolome and the Agricultural and Pharmaceutical Potential of Sea Foam-Derived Fungi. Marine Drugs. 2020; 18(2):128. https://doi.org/10.3390/md18020128
Chicago/Turabian StyleOppong-Danquah, Ernest, Cristina Passaretti, Orazio Chianese, Martina Blümel, and Deniz Tasdemir. 2020. "Mining the Metabolome and the Agricultural and Pharmaceutical Potential of Sea Foam-Derived Fungi" Marine Drugs 18, no. 2: 128. https://doi.org/10.3390/md18020128
APA StyleOppong-Danquah, E., Passaretti, C., Chianese, O., Blümel, M., & Tasdemir, D. (2020). Mining the Metabolome and the Agricultural and Pharmaceutical Potential of Sea Foam-Derived Fungi. Marine Drugs, 18(2), 128. https://doi.org/10.3390/md18020128