Secondary Metabolites from the Culture of the Marine-derived Fungus Paradendryphiella salina PC 362H and Evaluation of the Anticancer Activity of Its Metabolite Hyalodendrin

 , ,
, ,  ,
,  ,
, 
Abstract
:1. Introduction
2. Results and Discussion
2.1. Cytotoxic Activity of the Crude Extracts on MCF7, MCF7-Sh-WISP2 and 3T3-F442A Cell Lines
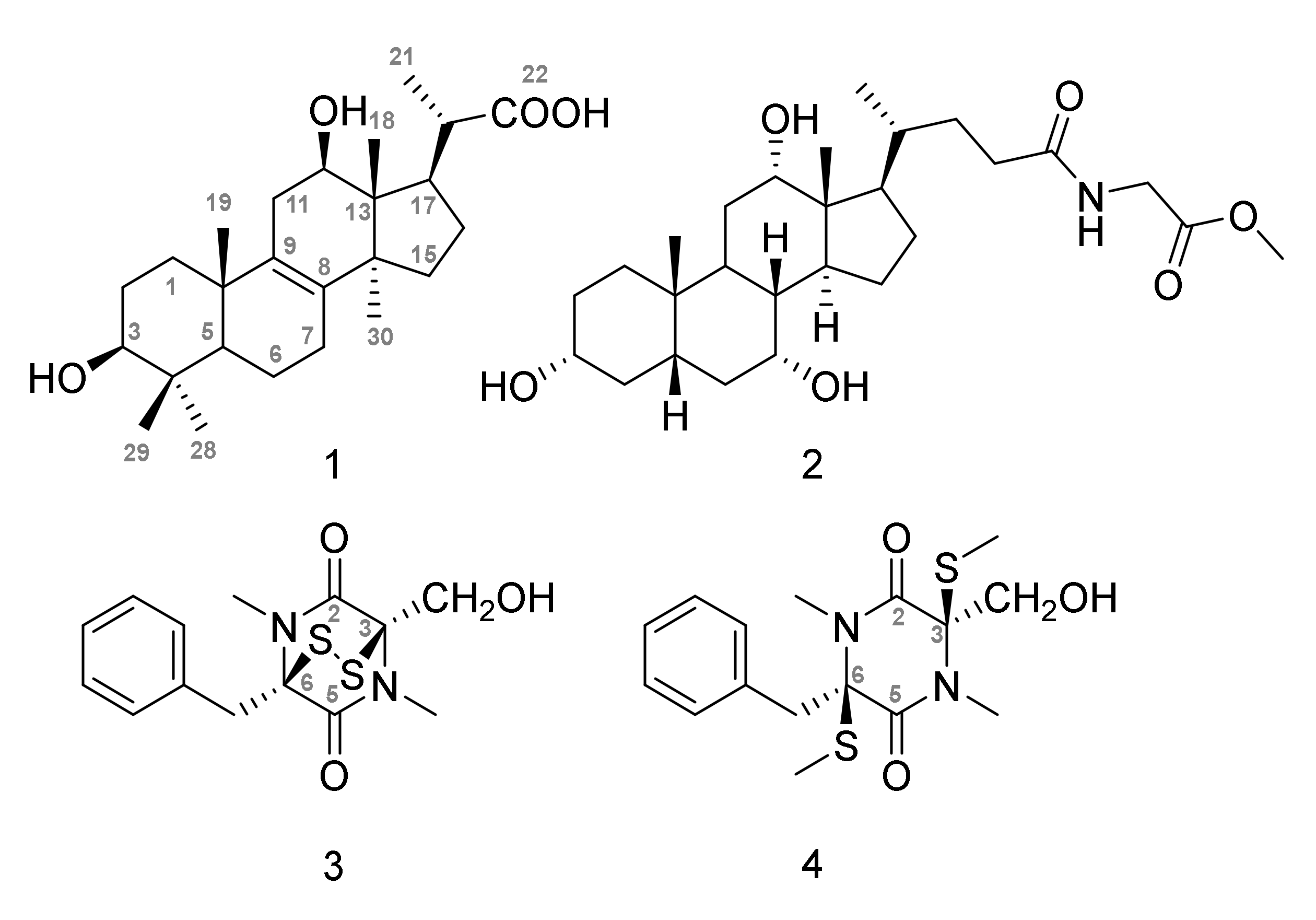
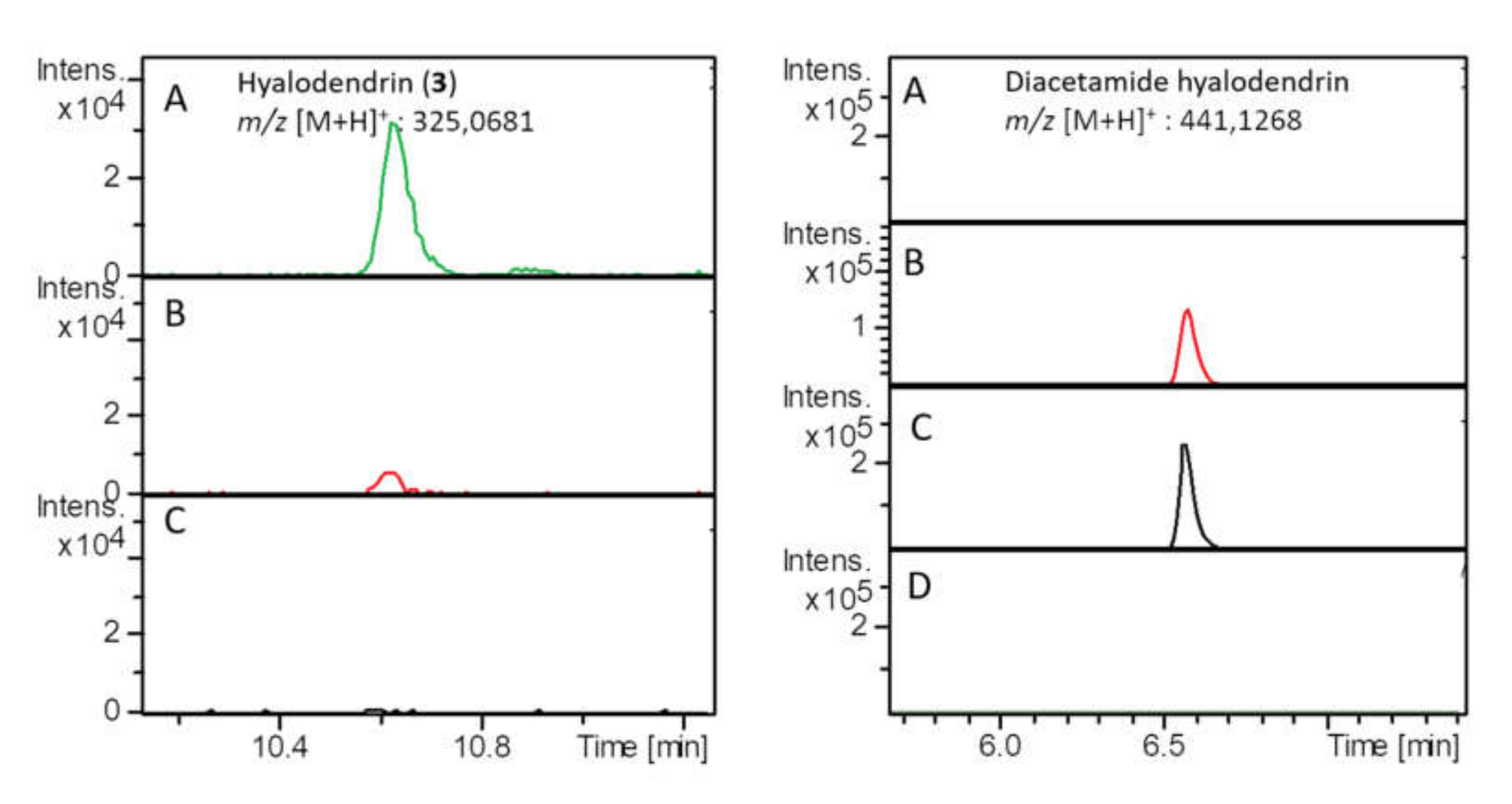
2.2. Chemical Investigation of Paradendryphiella Salina PC 362H
2.3. Cytotoxic Activity of 1–4 toward MCF7, MCF7-Sh-WISP2 and 3T3-F442A Cells
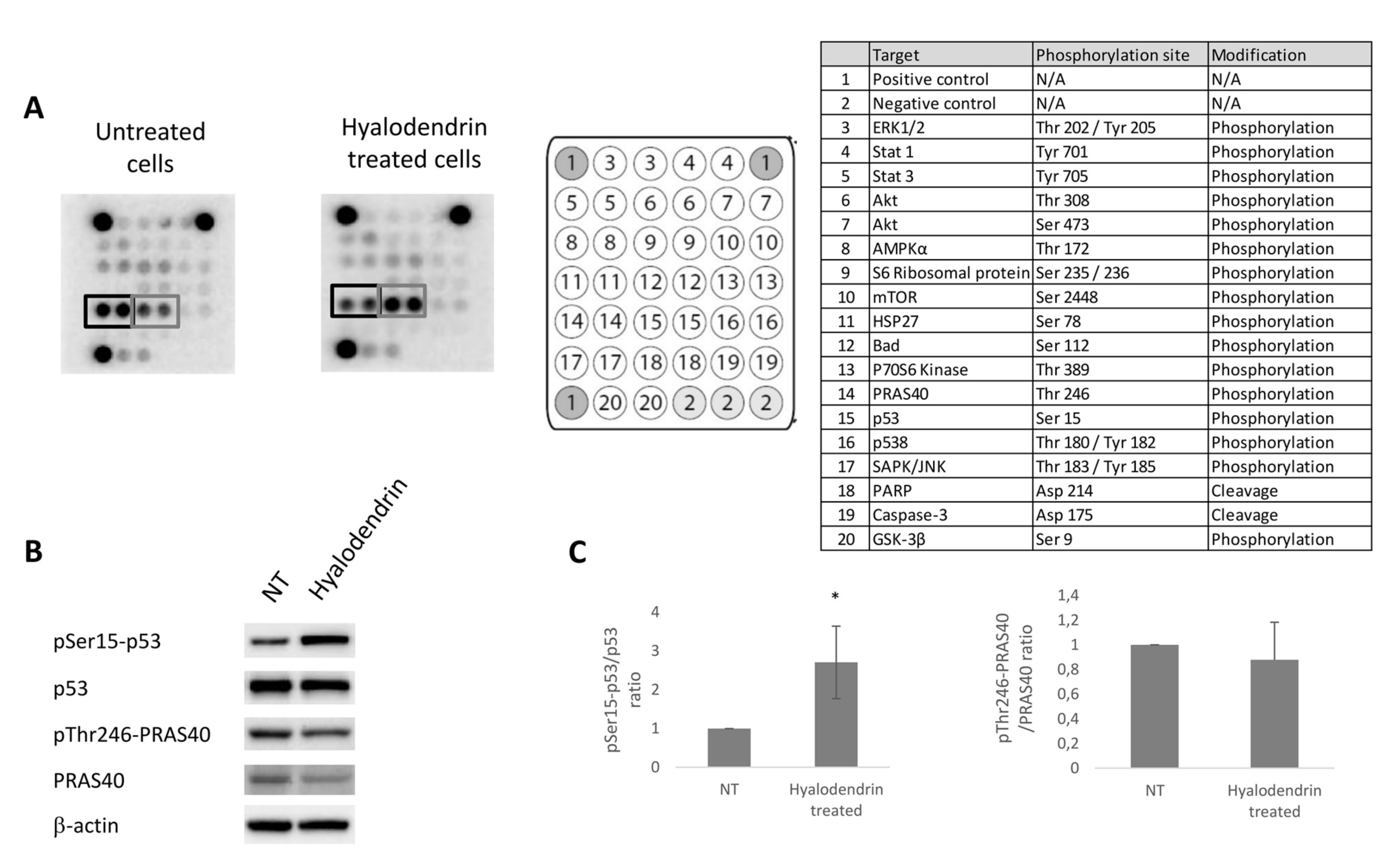
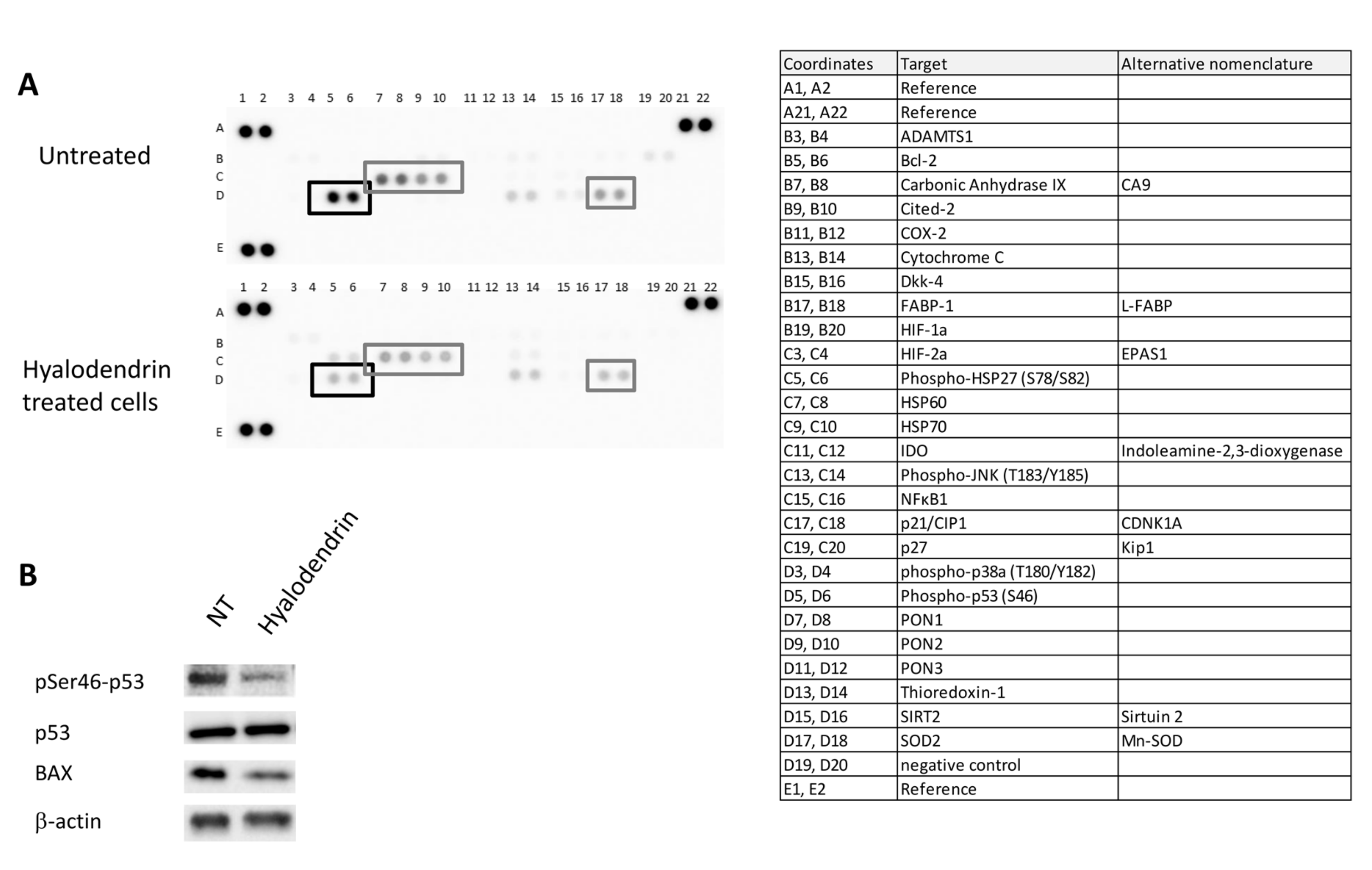
2.4. Cell Signaling Pathways Modulated Following Exposure to 3
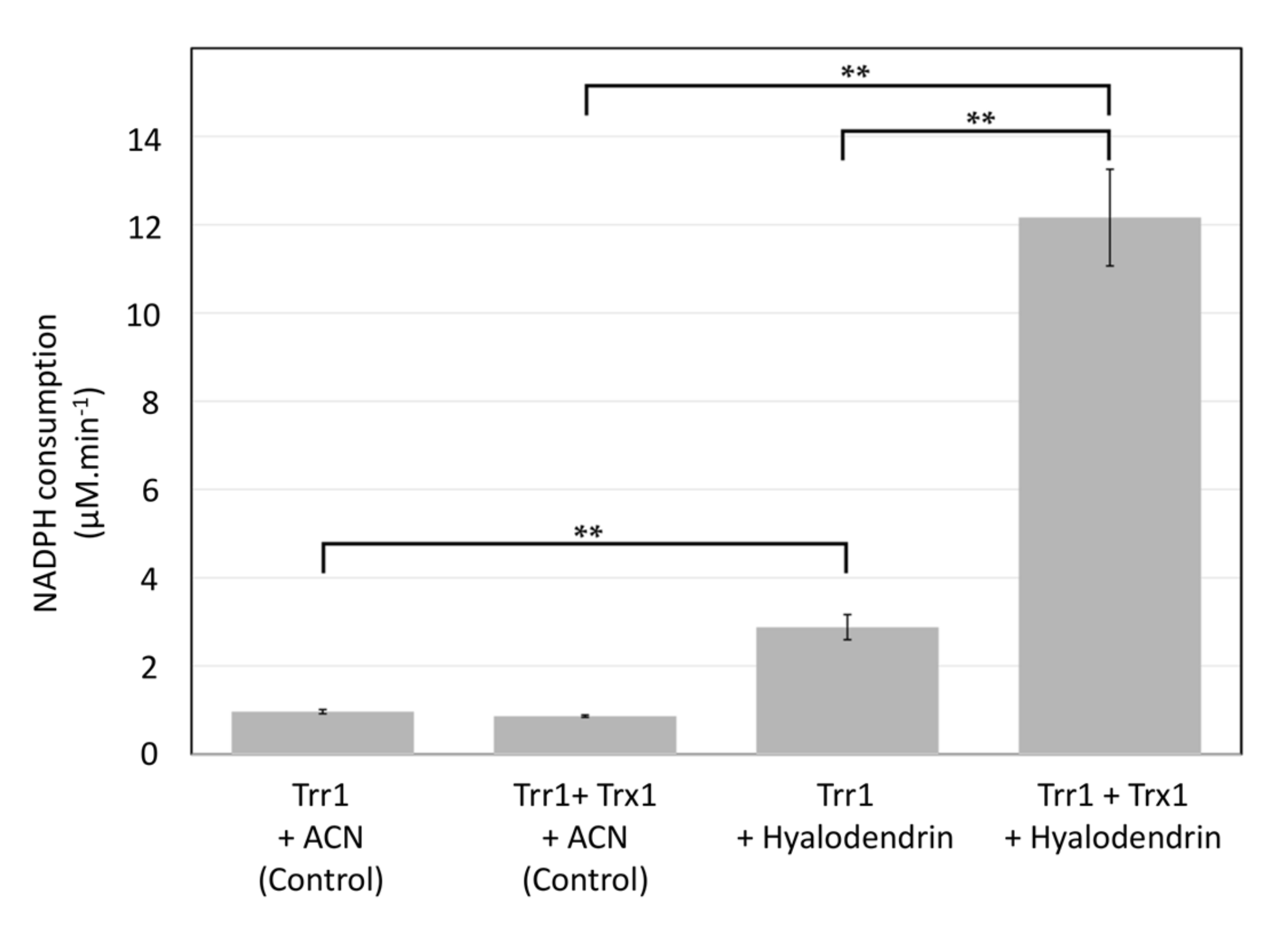
2.5. Compound 3 is Efficiently Reduced by the Thioredoxin System in Vitro
2.6. Compound 3 is Active Against a Panel of Cancer Cell Lines with Aggressive Phenotypes
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Crude Extracts Library
3.3. Fermentation, Extractions and Purification
3.4. Antibodies
3.5. Cells
3.6. Viability Assays
3.7. Compound 3 Concentrations
3.8. Intracellular Signaling Array
3.9. Human Cell Stress Array
3.10. Immunoblotting
3.11. In Vitro Reduction in Hyalodendrin (3) by the Thioredoxin System
3.12. High-resolution Accurate-Mass (HRMS) Mass Spectrometry
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Nowell, P.C. The clonal evolution of tumor cell populations. Science 1976, 194, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Greaves, M.; Maley, C.C. Clonal evolution in cancer. Nature 2012, 481, 306–313. [Google Scholar] [CrossRef] [PubMed]
- Escargueil, A.E.; Prado, S.; Dezaire, A.; Clairambault, J.; Larsen, A.K.; Soares, D.G. Genotype- or phenotype-targeting anticancer therapies? Lessons from tumor evolutionary biology. Curr. Pharm. Des. 2016, 22, 6625–6644. [Google Scholar] [CrossRef] [PubMed]
- Allred, D.C.; Mohsin, S.K.; Fuqua, S.A. Histological and biological evolution of human premalignant breast disease. Endocr. Relat. Cancer 2001, 8, 47–61. [Google Scholar] [CrossRef] [Green Version]
- Polyak, K. Breast cancer: Origins and evolution. J. Clin. Investig. 2007, 117, 3155–3163. [Google Scholar] [CrossRef]
- Radisky, D.C. Epithelial-mesenchymal transition. J. Cell Sci. 2005, 118, 4325–4326. [Google Scholar] [CrossRef] [Green Version]
- Thomson, S.; Buck, E.; Petti, F.; Griffin, G.; Brown, E.; Ramnarine, N.; Iwata, K.K.; Gibson, N.; Haley, J.D. Epithelial to mesenchymal transition is a determinant of sensitivity of non-small-cell lung carcinoma cell lines and xenografts to epidermal growth factor receptor inhibition. Cancer Res. 2005, 65, 9455–9462. [Google Scholar] [CrossRef] [Green Version]
- Akalay, I.; Janji, B.; Hasmim, M.; Noman, M.Z.; André, F.; De Cremoux, P.; Bertheau, P.; Badoual, C.; Vielh, P.; Larsen, A.K.; et al. Epithelial-to-mesenchymal transition and autophagy induction in breast carcinoma promote escape from T-cell-mediated lysis. Cancer Res. 2013, 73, 2418–2427. [Google Scholar] [CrossRef] [Green Version]
- Sequist, L.V.; Waltman, B.A.; Dias-Santagata, D.; Digumarthy, S.; Turke, A.B.; Fidias, P.; Bergethon, K.; Shaw, A.T.; Gettinger, S.; Cosper, A.K.; et al. Genotypic and histological evolution of lung cancers acquiring resistance to EGFR inhibitors. Sci. Transl. Med. 2011, 3, 75ra26. [Google Scholar] [CrossRef] [Green Version]
- Ayadi, M.; Bouygues, A.; Ouaret, D.; Ferrand, N.; Chouaib, S.; Thiery, J.P.; Muchardt, C.; Sabbah, M.; Larsen, A.K. Chronic chemotherapeutic stress promotes evolution of stemness and WNT/beta-catenin signaling in colorectal cancer cells: Implications for clinical use of WNT-signaling inhibitors. Oncotarget 2015, 6, 18518–18533. [Google Scholar] [CrossRef]
- Chua, K.N.; Sim, W.J.; Racine, V.; Lee, S.Y.; Goh, B.C.; Thiery, J.P. A cell-based small molecule screening method for identifying inhibitors of epithelial-mesenchymal transition in carcinoma. PLoS ONE 2012, 7, e33183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhang, W.; Qin, L. Mesenchymal-mode migration assay and antimetastatic drug screening with high-throughput microfluidic channel networks. Angew. Chem. Int. Ed. Engl. 2014, 53, 2344–2348. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- König, G.M.; Kehraus, S.; Seibert, S.F.; Abdel-Lateff, A.; Müller, D. Natural products from marine organisms and their associated microbes. Chembiochem 2006, 7, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Bugni, T.S.; Ireland, C.M. Marine-derived fungi: A chemically and biologically diverse group of microorganisms. Nat. Prod. Rep. 2004, 21, 143–163. [Google Scholar] [CrossRef]
- Vallet, M.; Strittmatter, M.; Murúa, P.; Lacoste, S.; Dupont, J.; Hubas, C.; Genta-Jouve, G.; Gachon, C.M.M.; Kim, G.H.; Prado, S. Chemically-Mediated interactions between macroalgae, their fungal endophytes, and protistan pathogens. Front. Microbiol. 2018, 9, 3161. [Google Scholar] [CrossRef] [Green Version]
- Tourneroche, A.; Lami, R.; Hubas, C.; Blanchet, E.; Vallet, M.; Escoubeyrou, K.; Paris, A.; Prado, S. Bacterial-fungal interactions in the kelp endomicrobiota drive autoinducer-2 quorum sensing. Front. Microbiol. 2019, 10, 1693. [Google Scholar] [CrossRef] [Green Version]
- Fritah, A.; Saucier, C.; De Wever, O.; Bracke, M.; Bièche, I.; Lidereau, R.; Gespach, C.; Drouot, S.; Redeuilh, G.; Sabbah, M. Role of WISP-2/CCN5 in the maintenance of a differentiated and noninvasive phenotype in human breast cancer cells. Mol. Cell. Biol. 2008, 28, 1114–1123. [Google Scholar] [CrossRef] [Green Version]
- Ferrand, N.; Gnanapragasam, A.; Dorothee, G.; Redeuilh, G.; Larsen, A.K.; Sabbah, M. Loss of WISP2/CCN5 in estrogen-dependent MCF7 human breast cancer cells promotes a stem-like cell phenotype. PLoS ONE 2014, 9, e87878. [Google Scholar] [CrossRef]
- Guerriero, A.; D’Ambrosio, M.; Cuomo, V.; Vanzanella, F.; Pietra, F. Dendryphiellin A, the first fungal trinoreremophilane. Isolation from the marine deuteromycete Dendryphiella salina (Sutherland) Pugh et Nicot. Helv. Chim. Acta 1988, 71, 57–61. [Google Scholar] [CrossRef]
- Guerriero, A.; D’Ambrosio, M.; Cuomo, V.; Vanzanella, F.; Pietra, F. Novel trinoreremophilanes (dendryphiellin B, C, and D), eremophilanes (dendryphiellin E, F, and G), and branched C9-carboxylic acids (dendryphiellic acid A and B) from the marine deuteromycete Dendryphiella salina (Sutherland) Pugh and Nicot. Helv. Chim. Acta 1989, 72, 438–446. [Google Scholar] [CrossRef]
- Guerriero, A.; Cuomo, V.; Vanzanella, F.; Pietra, F. A novel glyceryl ester (glyceryl dendryphiellate A), a trinor-eremophilane (dendryphiellin A1), and eremophilanes (dendryphiellin E1 and E2) from the marine deuteromycete Dendryphiella salina (SUTHERLAND) PUGHet NICOT. Helv. Chim. Acta 1990, 73, 2090–2096. [Google Scholar] [CrossRef]
- Tanaka, M.; Ohra, J.; Tsujtno, Y.; Fujimori, T.; Agob, H.; Tsuge, H. Dendryol A, B, C, and D, four new compounds produced by a weed pathogenic fungus Dendryphiella sp. Z. Naturforsch. C 1995, 50, 751–756. [Google Scholar] [CrossRef]
- Yoiprommarat, S.; Srichomthong, K.; Deelai, S.; Suetrong, S.; Sakayaroj, J.; Bunyapaiboonsri, T.; Unagul, P. Secondary metabolites of the marine fungus Paradendryphiellaarenariae BCC 17999. Bot. Mar. 2015, 58, 393–399. [Google Scholar] [CrossRef]
- Pil, G.; Won, H.; Shin, H. Bile acids from a marine sponge-associated fungus penicillium sp. J. Korean Magn. Reson. Soc. 2016, 20, 41–45. [Google Scholar] [CrossRef] [Green Version]
- Michel, K.H.; Chaney, M.O.; Jones, N.D.; Hoehn, M.M.; Nagarajan, R. Epipolythiopiperazinedione antibiotics from Penicillium turbatum. J. Antibiot. (Tokyo) 1974, 27, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.; Fenical, W. Isolation of gliovictin from the marine deuteromycete Asteromyces cruciatus. Phytochemistry 1987, 26, 3347. [Google Scholar] [CrossRef]
- Xue, C.Y.; Li, J.S.; Qi, H.; Xi, F.Y.; Xiang, W.S.; Wang, J.D.; Wang, X.J. Two new pentanorlanostane metabolites from a soil fungus Curvularia borreriae strain HS-FG-237. J. Antibiot. (Tokyo) 2013, 66, 735–737. [Google Scholar] [CrossRef]
- Strunz, G.M.; Kakushima, M.; Stillwell, M.A.; Heissner, C.J. Hyalodendrin: A new fungitoxic epidithiodioxopiperazine produced by a Hyalodendron species. J. Chem. Soc. Perkin Trans. 1 1973, 2600–2602. [Google Scholar] [CrossRef]
- Takeuchi, R.; Shimokawa, J.; Fukuyama, T. Development of a route to chiral epidithiodioxopiperazine moieties and application to the asymmetric synthesis of (+)-hyalodendrin. Chem. Sci. 2014, 5, 2003–2006. [Google Scholar] [CrossRef]
- Stillwell, M.A.; Magasi, L.P.; Strunz, G.M. Production, isolation, and antimicrobial activity of hyalodendrin, a new antibiotic produced by a species of Hyalodendron. Can. J. Microbiol. 1974, 20, 759–764. [Google Scholar] [CrossRef] [Green Version]
- Welch, T.R.; Williams, R.M. Epidithiodioxopiperazines. occurrence, synthesis and biogenesis. Nat. Prod. Rep. 2014, 31, 1376–1404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meek, D.W. Tumour suppression by p53: A role for the DNA damage response? Nat. Rev. Cancer 2009, 9, 714–723. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Harris, T.E.; Roth, R.A.; Lawrence, J.C. PRAS40 regulates mTORC1 kinase activity by functioning as a direct inhibitor of substrate binding. J. Biol. Chem. 2007, 282, 20036–20044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, D.; Guo, L.; Zhang, T.; Huang, L. PRAS40 signaling in tumor. Oncotarget 2017, 8, 69076–69085. [Google Scholar] [CrossRef] [Green Version]
- Escargueil, A.E.; Soares, D.G.; Salvador, M.; Larsen, A.K.; Henriques, J.A. What histone code for DNA repair? Mutat. Res. 2008, 658, 259–270. [Google Scholar] [CrossRef]
- Jänicke, R.U.; Sprengart, M.L.; Wati, M.R.; Porter, A.G. Caspase-3 is required for DNA fragmentation and morphological changes associated with apoptosis. J. Biol. Chem. 1998, 273, 9357–9360. [Google Scholar] [CrossRef] [Green Version]
- Mayo, L.D.; Seo, Y.R.; Jackson, M.W.; Smith, M.L.; Rivera Guzman, J.; Korgaonkar, C.K.; Donner, D.B. Phosphorylation of human p53 at serine 46 determines promoter selection and whether apoptosis is attenuated or amplified. J. Biol. Chem. 2005, 280, 25953–25959. [Google Scholar] [CrossRef] [Green Version]
- Smeenk, L.; van Heeringen, S.J.; Koeppel, M.; Gilbert, B.; Janssen-Megens, E.; Stunnenberg, H.G.; Lohrum, M. Role of p53 serine 46 in p53 target gene regulation. PLoS ONE 2011, 6, e17574. [Google Scholar] [CrossRef] [Green Version]
- Zylicz, M.; King, F.W.; Wawrzynow, A. Hsp70 interactions with the p53 tumour suppressor protein. EMBO J. 2001, 20, 4634–4638. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, J.C.; Dohi, T.; Kang, B.H.; Altieri, D.C. Hsp60 regulation of tumor cell apoptosis. J. Biol. Chem. 2008, 283, 5188–5194. [Google Scholar] [CrossRef] [Green Version]
- Liu, D.; Xu, Y. P53, oxidative stress, and aging. Antioxid. Redox Signal. 2011, 15, 1669–1678. [Google Scholar] [CrossRef]
- Hall, L.; Martinus, R.D. Hyperglycaemia and oxidative stress upregulate HSP60 & HSP70 expression in HeLa cells. Springerplus 2013, 2, 431. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Candas, D.; Li, J.J. MnSOD in oxidative stress response-potential regulation via mitochondrial protein influx. Antioxid. Redox Signal. 2014, 20, 1599–1617. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chai, C.L.; Waring, P. Redox sensitive epidithiodioxopiperazines in biological mechanisms of toxicity. Redox Rep. 2000, 5, 257–264. [Google Scholar] [CrossRef] [PubMed]
- Weindling, R.; Emerson, O.H. The isolation of a toxic substance from the culture filtrate of Trichoderma. Phytopathology 1936, 26, 1068–1070. [Google Scholar]
- Mason, J.W.; Kidd, J.G. Effects of gliotoxin and other sulfur-containing compounds on tumor cells in vitro; with observations on the mechanism of action of gliotoxin. J. Immunol. 1951, 66, 99–106. [Google Scholar] [PubMed]
- Eichner, R.D.; Waring, P.; Geue, A.M.; Braithwaite, A.W.; Müllbacher, A. Gliotoxin causes oxidative damage to plasmid and cellular DNA. J. Biol. Chem. 1988, 263, 3772–3777. [Google Scholar]
- Waring, P.; Beaver, J. Gliotoxin and related epipolythiodioxopiperazines. Gen. Pharmacol. 1996, 27, 1311–1316. [Google Scholar] [CrossRef]
- Zaffagnini, M.; Fermani, S.; Costa, A.; Lemaire, S.D.; Trost, P. Plant cytoplasmic GAPDH: Redox post-translational modifications and moonlighting properties. Front. Plant Sci. 2013, 4, 450. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.Y.; Zhang, F.; Hong, C.Q.; Giuliano, A.E.; Cui, X.J.; Zhou, G.J.; Zhang, G.J.; Cui, Y.K. Critical protein GAPDH and its regulatory mechanisms in cancer cells. Cancer Biol. Med. 2015, 12, 10–22. [Google Scholar] [CrossRef]
- Bedhomme, M.; Adamo, M.; Marchand, C.H.; Couturier, J.; Rouhier, N.; Lemaire, S.D.; Zaffagnini, M.; Trost, P. Glutathionylation of cytosolic glyceraldehyde-3-phosphate dehydrogenase from the model plant Arabidopsis thaliana is reversed by both glutaredoxins and thioredoxins in vitro. Biochem. J. 2012, 445, 337–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaffagnini, M.; Bedhomme, M.; Groni, H.; Marchand, C.H.; Puppo, C.; Gontero, B.; Cassier-Chauvat, C.; Decottignies, P.; Lemaire, S.D. Glutathionylation in the photosynthetic model organism Chlamydomonas reinhardtii: A proteomic survey. Mol. Cell. Proteom. 2012, 11, M111.014142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaffagnini, M.; Bedhomme, M.; Lemaire, S.D.; Trost, P. The emerging roles of protein glutathionylation in chloroplasts. Plant Sci. 2012, 185–186, 86–96. [Google Scholar] [CrossRef]
- Mahmood, D.F.; Abderrazak, A.; El Hadri, K.; Simmet, T.; Rouis, M. The thioredoxin system as a therapeutic target in human health and disease. Antioxid. Redox Signal. 2013, 19, 1266–1303. [Google Scholar] [CrossRef] [PubMed]
- Harris, I.S.; Treloar, A.E.; Inoue, S.; Sasaki, M.; Gorrini, C.; Lee, K.C.; Yung, K.Y.; Brenner, D.; Knobbe-Thomsen, C.B.; Cox, M.A.; et al. Glutathione and thioredoxin antioxidant pathways synergize to drive cancer initiation and progression. Cancer Cell 2015, 27, 211–222. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Li, X.; Han, X.; Liu, R.; Fang, J. Targeting the thioredoxin system for cancer therapy. Trends Pharmacol. Sci. 2017, 38, 794–808. [Google Scholar] [CrossRef]
- Srinivas, U.S.; Tan, B.W.Q.; Vellayappan, B.A.; Jeyasekharan, A.D. ROS and the DNA damage response in cancer. Redox Biol. 2018, 101084. [Google Scholar] [CrossRef]
- Biscardi, J.S.; Belsches, A.P.; Parsons, S.J. Characterization of human epidermal growth factor receptor and c-Src interactions in human breast tumor cells. Mol. Carcinog. 1998, 21, 261–272. [Google Scholar] [CrossRef]
- Herbst, R.S.; Langer, C.J. Epidermal growth factor receptors as a target for cancer treatment: The emerging role of IMC-C225 in the treatment of lung and head and neck cancers. Semin. Oncol. 2002, 29, 27–36. [Google Scholar] [CrossRef]
- Kester, D.R.I.W.; Duedall, I.W.; Connors, D.N.; Pytkowicz, R.M. Preparation of artificial seawater. Limnol. Oceanog. 1967, 12, 176–178. [Google Scholar] [CrossRef]
- Petitprez, A.; Poindessous, V.; Ouaret, D.; Regairaz, M.; Bastian, G.; Guérin, E.; Escargueil, A.E.; Larsen, A.K. Acquired irinotecan resistance is accompanied by stable modifications of cell cycle dynamics independent of MSI status. Int. J. Oncol. 2013, 42, 1644–1653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Pérez, M.E.; Zaffagnini, M.; Marchand, C.H.; Crespo, J.L.; Lemaire, S.D. The yeast autophagy protease Atg4 is regulated by thioredoxin. Autophagy 2014, 10, 1953–1964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardiner, D.M.; Waring, P.; Howlett, B.J. The epipolythiodioxopiperazine (ETP) class of fungal toxins: Distribution, mode of action, functions and biosynthesis. Microbiology 2005, 151, 1021–1032. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramsay, E.E.; Dilda, P.J. Glutathione S-conjugates as prodrugs to target drug-resistant tumors. Front. Pharmacol. 2014, 5, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 (µg/mL) | IC50 (µg/mL) | |||||||
|---|---|---|---|---|---|---|---|---|
| Strain Code | Culture Medium | MCF7 | MCF7-Sh- WISP2 | 3T3-F442A | Culture Medium | MCF7 | MCF7-Sh- WISP2 | 3T3-F442A |
| LD150F | MEA–ASW | 0.3 | 2.7 | 6.3 | Tubaki | 2.4 | 3.4 | 36.8 |
| AN325T | 1.5 | 0.4 | 3.7 | 1.8 | 1.2 | 3.8 | ||
| LD327H | 0.9 | 0.3 | 0.7 | 0.5 | 0.1 | 0.1 | ||
| SL332T | 4.2 | 3.0 | 3.3 | 61.0 | 111.2 | 50.7 | ||
| PC362H | 0.4 | 0.2 | 0.5 | 8.3 | 10.0 | 16.7 | ||
| SL469T | 88.2 | 22.8 | 37.6 | 2.3 | 2.8 | 4.7 | ||
| LD481H | 0.3 | 0.4 | 0.3 | 0.3 | 0.9 | 0.2 | ||
| Pos. | δC, Type | δH Mult. (J in Hz). | Pos. | δC, Type | δH Mult. (J in Hz) |
|---|---|---|---|---|---|
| 1 | 35.4, CH2 | (α) 1.14, m; (β) 1.60, m | 15 | 30.7, CH2 | (α) 1.09, m; (β) 1.58, m |
| 2 | 27.6, CH2 | 1.47, m | 16 | 25.9, CH2 | (α) 1.54, m, (β) 1.80, m |
| 3 | 76.7, CH | 2.99, dt (10.7, 5.1) | 17 | 48.1, CH | 1.91, m |
| 4 | 38.6, C | - | CH3-18 | 9.7, CH3 | 0.59, s |
| 5 | 50.0, CH | 0.93,d (1.9) | CH3-19 | 18.9, CH3 | 0.91, s |
| 6 | 17.8, CH2 | (β) * 1.40, m; (α) * 1.63, m | 20 | 41.3, CH | 2.36, m |
| 7 | 25.6, CH2 | 1.95, m | CH3-21 | 19.0, CH3 | 1.21, d (6.6) |
| 8 | 132.8, C | - | 22 | 178.9, C | - |
| 9 | 135.8, C | - | CH3-28 | 28.1, CH3 | 0.89, s |
| 10 | 36.3, C | - | CH3-29 | 15.8, CH3 | 0.69, s |
| 11 | 33.6, CH2 | (α) 2.38, m; (β) 1.68, m | CH3-30 | 23.7, CH3 | 0.84, s |
| 12 | 69.8, CH | 3.87, brt (7.5) | OH-3 | - | 4.31, d (5.1) |
| 13 | 48.3, C | - | OH-12 | - | 4.42, br s |
| 14 | 51.6, C | - |
| Cell Line | IC50 µg/mL (µM) | ||||
|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | Hydroxi Tamoxifen | |
| MCF7 | 20 µg/mL (50 µM) | > 25 µg/mL (> 50 µM) | 0.07 µg/mL (0.22 µM) | 15 µg/mL (42 µM) | 8.8 µM |
| MCF7-Sh-WISP2 | > 25 µg/mL (> 60 µM) | > 25 µg/mL (> 50 µM) | 0.046 µg/mL (0.14 µM) | 24 µg/mL (68 µM) | 11.6 µM |
| 3T3-F442A | > 25 µg/mL (> 60 µM) | > 25 µg/mL (> 50 µM) | 0.099 µg/mL (0.3 µM) | 92 µg/mL (26 µM) | 14.5 µM |
| Cell line | IC50 (nM) | Phenotypes | Relevant Genetic Status | MSS vs MSI | |
|---|---|---|---|---|---|
| Breast | MC7-Sh-WISP2 | 142.0 ± 3.0 | mesenchymal, invasive | ER-/PR-/HER2- | |
| MCF7 | 216.3 ± 6.0 | epithelial | ER+/PR+/HER2-, TP53 WT | ||
| MDA-MB-231 | 132.5 ± 13.4 | mesenchymal, invasive | ER-/PR-/HER2-, TP53 mutant (R280K) | ||
| Colon | SW48 | 149.0 ± 15.5 | mesenchymal, invasive | BRAF and KRAS WT, TP53 mutant (R248Y) | MSI |
| DLD1 | 40.0 ± 5.8 | epithelial | KRAS mutant (G13D), TP53 mutant (S241F) | MSI | |
| HT29 | 58.0 ± 13.7 | epithelial | BRAF mutant (V600E), TP53 mutant (R273H) | MSS | |
| HT29 5FU | 146.8 ± 10.2 | mesenchymal, invasive | ND | MSS | |
| HT29 oxa | 141.8 ± 2.6 | epithelial | ND | MSS | |
| HT29 SN-38 | 93.8 ± 13.6 | epithelial | ND | MSS | |
| HCT116 | 48.0 ± 9.3 | epithelial | KRAS mutant (G13D), TP53 WT | MSI | |
| HCT116 5FU | 72.0 ± 10.9 | mesenchymal, invasive | ND | MSI | |
| HCT116 oxa | 25.7 ± 4.2 | mesenchymal, invasive | ND | MSI | |
| HCT116 SN-38 | 43.8 ± 4.2 | mesenchymal, invasive | ND | MSI | |
| LS513 | 78.0 ± 9.3 | epithelial | KRAS mutant (G12D), TP53 WT | MSS | |
| LOVO | 73.4 ± 16.4 | epithelial | KRAS mutant (G13D; A14V), TP53 WT | MSI | |
| RKO | 74.3 ± 1.9 | mesenchymal, invasive | BRAF mutant (V600E), TP53 WT | MSI | |
| LS174T | 158.0 ± 13.0 | epithelial | KRAS mutant (G12D), TP53 WT | MSI | |
| SW480 | 163.7 ± 11.0 | mesenchymal, invasive | KRAS mutant (G12V), TP53 mutant (R273H) | MSS | |
| Cervix | HeLa | 69.0 ± 9.9 | epithelial | TP53 WT (HPV positive) | |
| Normal | 3T3F442A | 305.0 ± 4.2 | pre-adipocytic | ||
| C19 | 252.5 ± 20.5 | fibroblastic |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dezaire, A.; Marchand, C.H.; Vallet, M.; Ferrand, N.; Chaouch, S.; Mouray, E.; Larsen, A.K.; Sabbah, M.; Lemaire, S.D.; Prado, S.; et al. Secondary Metabolites from the Culture of the Marine-derived Fungus Paradendryphiella salina PC 362H and Evaluation of the Anticancer Activity of Its Metabolite Hyalodendrin. Mar. Drugs 2020, 18, 191. https://doi.org/10.3390/md18040191
Dezaire A, Marchand CH, Vallet M, Ferrand N, Chaouch S, Mouray E, Larsen AK, Sabbah M, Lemaire SD, Prado S, et al. Secondary Metabolites from the Culture of the Marine-derived Fungus Paradendryphiella salina PC 362H and Evaluation of the Anticancer Activity of Its Metabolite Hyalodendrin. Marine Drugs. 2020; 18(4):191. https://doi.org/10.3390/md18040191
Chicago/Turabian StyleDezaire, Ambre, Christophe H. Marchand, Marine Vallet, Nathalie Ferrand, Soraya Chaouch, Elisabeth Mouray, Annette K. Larsen, Michèle Sabbah, Stéphane D. Lemaire, Soizic Prado, and et al. 2020. "Secondary Metabolites from the Culture of the Marine-derived Fungus Paradendryphiella salina PC 362H and Evaluation of the Anticancer Activity of Its Metabolite Hyalodendrin" Marine Drugs 18, no. 4: 191. https://doi.org/10.3390/md18040191
APA StyleDezaire, A., Marchand, C. H., Vallet, M., Ferrand, N., Chaouch, S., Mouray, E., Larsen, A. K., Sabbah, M., Lemaire, S. D., Prado, S., & Escargueil, A. E. (2020). Secondary Metabolites from the Culture of the Marine-derived Fungus Paradendryphiella salina PC 362H and Evaluation of the Anticancer Activity of Its Metabolite Hyalodendrin. Marine Drugs, 18(4), 191. https://doi.org/10.3390/md18040191