The Effects of the Marine-Derived Polysaccharides Laminarin and Chitosan on Aspects of Colonic Health in Pigs Challenged with Dextran Sodium Sulphate


 , , , , , ,
, , , , , ,
Abstract
:1. Introduction
2. Results
2.1. Growth Performance and Faecal Scores
2.1.1. Growth Performance in the Pre-Challenge Period (d0-35)
2.1.2. Growth Performance and Faecal Scores in the DSS Challenge Period (d35-44)
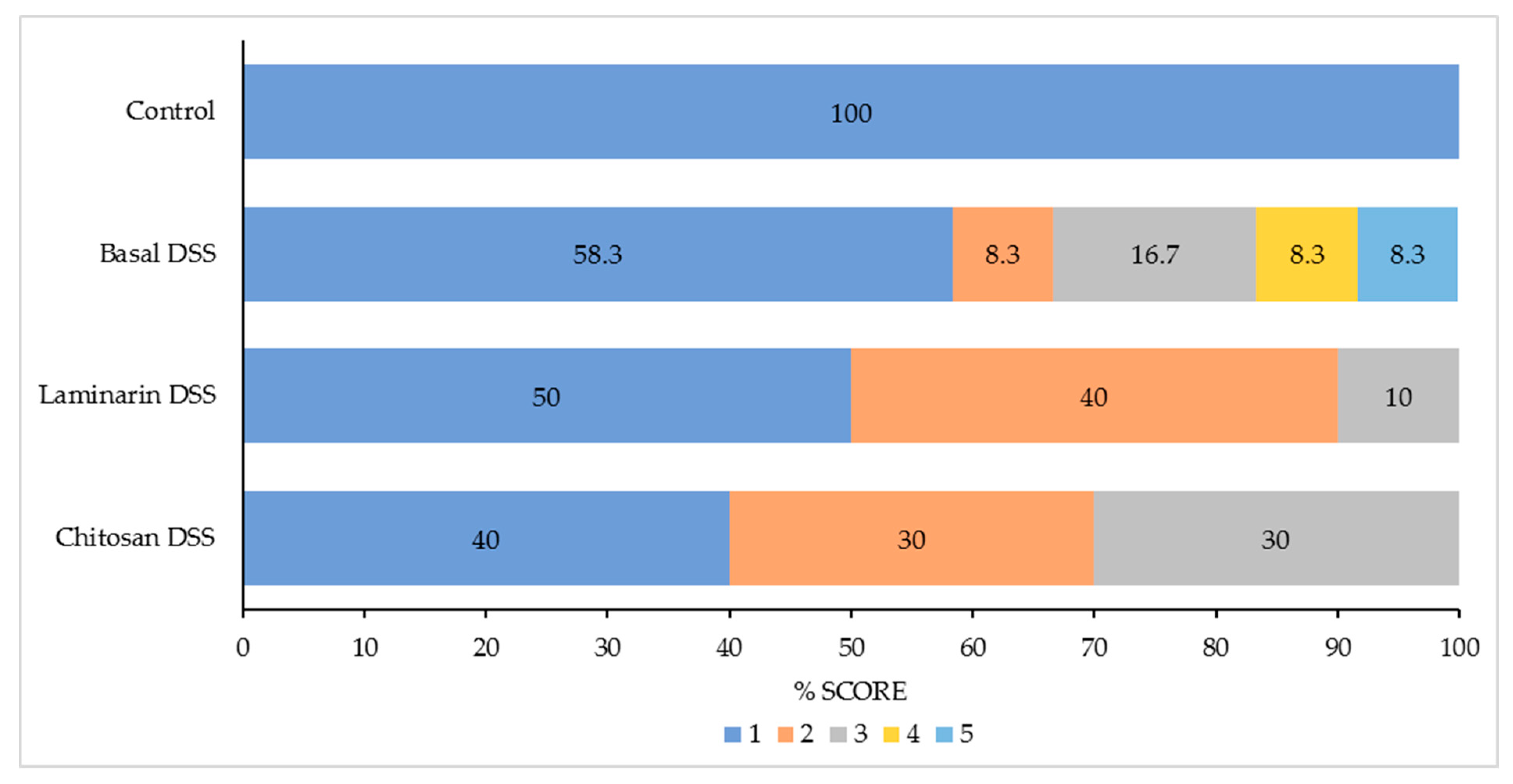
2.2. Colonic Histopathology
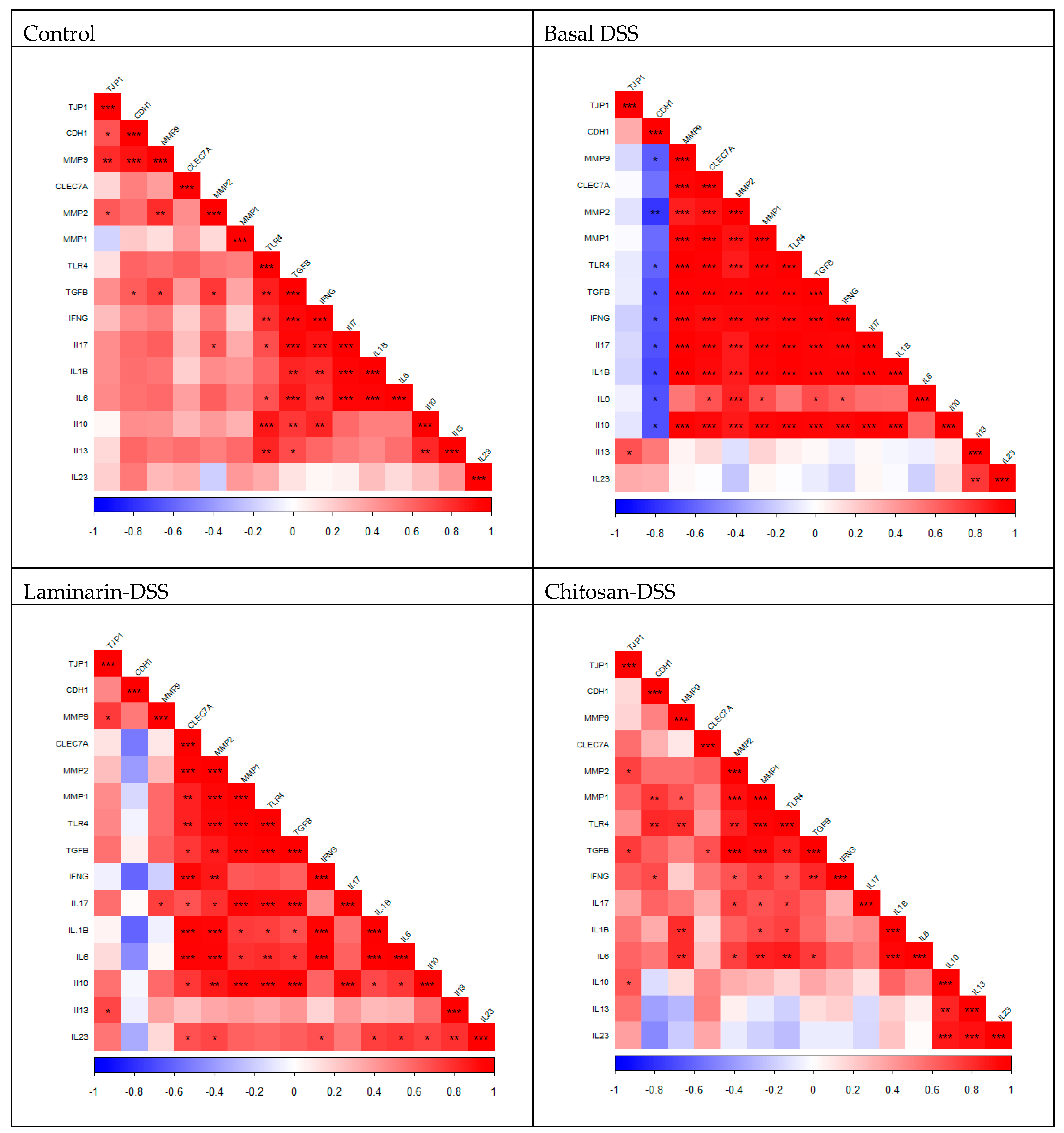
2.3. Colonic Gene Expression
Gene Expression Correlations
2.4. Volatile Fatty Acids (VFA)
2.5. Microbiology
2.5.1. Alpha Diversity Analysis
2.5.2. Differential Abundance Analysis of Bacterial Taxa in Colonic Digesta
Comparison of the Control and the Basal DSS Groups
Comparison of Basal DSS Group with the Laminarin and Chitosan DSS Groups
3. Discussion
4. Materials and Methods
4.1. Experimental Design and Diets
4.2. Housing and Animal Management
4.3. Induction of Experimental Colitis, Clinical Assessment of Colitis and Sample Collection
4.4. Feed Analysis
4.5. Colon Histopathology
4.6. Gene Expression Analysis
4.6.1. RNA Extraction and cDNA Synthesis
4.6.2. Quantitative Real-Time PCR (QPCR)
4.7. Volatile Fatty Acid (VFA) Analysis
4.8. Microbial Analysis
4.8.1. DNA Isolation and Quantification
4.8.2. PCR Amplicon Library Preparation
4.8.3. Bioinformatic Analyses of Pyrosequencing Reads
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stange, E.F.; Travis, S.; Vermeire, S.; Reinisch, W.; Geboes, K.; Barakauskiene, A.; Feakins, R.; Fléjou, J.; Herfarth, H.; Hommes, D.; et al. European evidence-based Consensus on the diagnosis and management of ulcerative colitis: Definitions and diagnosis. J. Crohns Coliti 2008, 2, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.J.; Kovacs-Nolan, J.A.; Yang, C.; Archbold, T.; Fan, M.Z.; Mine, Y. l-Tryptophan exhibits therapeutic function in a porcine model of dextran sodium sulfate (DSS)-induced colitis. J. Nutr. Biochem. 2010, 21, 468–475. [Google Scholar] [CrossRef] [PubMed]
- Yadav, V.; Varum, F.; Bravo, R.; Furrer, E.; Bojic, D.; Basit, A.W. Inflammatory bowel disease: Exploring gut pathophysiology for novel therapeutic targets. Transl. Res. 2016, 176, 38–68. [Google Scholar] [CrossRef] [PubMed]
- Head, K.A.; Jurenka, J.S. Inflammatory bowel disease Part 1: Ulcerative colitis-pathophysiology and conventional and alternative treatment options. Altern. Med. Rev. 2003, 8, 247–283. [Google Scholar]
- Kokkinidis, D.G.; Bosdelekidou, E.E.; Iliopoulou, S.M.; Kousoulis, A.A. Emerging treatments for ulcerative colitis: A systematic review. Scand. J. Gastroenterol. 2017, 52, 923–931. [Google Scholar] [CrossRef]
- Håkansson, Å.; Tormo-Badia, N.; Baridi, A.; Xu, J.; Molin, G.; Hagslätt, M.-L.; Karlsson, C.; Jeppsson, B.; Cilio, C.M.; Ahrné, S. Immunological alteration and changes of gut microbiota after dextran sulfate sodium (DSS) administration in mice. Clin. Exp. Med. 2014, 15, 107–120. [Google Scholar] [CrossRef] [Green Version]
- Ardizzone, S.; Porro, G.B.; Porro, P.G.B. Biologic Therapy for Inflammatory Bowel Disease. Drugs 2005, 65, 2253–2286. [Google Scholar] [CrossRef]
- Atreya, R.; Neurath, M.F. New therapeutic strategies for treatment of inflammatory bowel disease. Mucosal Immunol. 2008, 1, 175–182. [Google Scholar] [CrossRef]
- Faubion, W.A.; Loftus, E.V.; Harmsen, W.S.; Zinsmeister, A.R.; Sandborn, W.J. The natural history of corticosteroid therapy for inflammatory bowel disease: A population-based study. Gastroenterology 2001, 121, 255–260. [Google Scholar] [CrossRef] [Green Version]
- Bassaganya-Riera, J.; Hontecillas, R. CLA and n-3 PUFA differentially modulate clinical activity and colonic PPAR-responsive gene expression in a pig model of experimental IBD. Clin. Nutr. 2006, 25, 454–465. [Google Scholar] [CrossRef]
- Rioux, L.-E.; Turgeon, S.; Beaulieu, M. Characterization of polysaccharides extracted from brown seaweeds. Carbohydr. Polym. 2007, 69, 530–537. [Google Scholar] [CrossRef]
- Devillé, C.; Damas, J.; Forget, P.; Dandrifosse, G.; Peulen, O. Laminarin in the dietary fibre concept. J. Sci. Food Agric. 2004, 84, 1030–1038. [Google Scholar] [CrossRef]
- Walsh, A.M.; Sweeney, T.; O’Shea, C.; Doyle, D.N.; O’Doherty, J.V. Effect of dietary laminarin and fucoidan on selected microbiota, intestinal morphology and immune status of the newly weaned pig. Br. J. Nutr. 2013, 110, 1630–1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sweeney, T.; Collins, C.B.; Reilly, P.; Pierce, K.M.; Ryan, M.; O’Doherty, J.V. Effect of purified β-glucans derived from Laminaria digitata, Laminaria hyperborea and Saccharomyces cerevisiae on piglet performance, selected bacterial populations, volatile fatty acids and pro-inflammatory cytokines in the gastrointestinal tract of pigs. Br. J. Nutr. 2012, 108, 1226–1234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, M.; Collins, C.B.; O’Doherty, J.V.; Sweeney, T. Effects of dietary β-glucans supplementation on cytokine expression in porcine liver. J. Anim. Sci. 2012, 90, 40–42. [Google Scholar] [CrossRef]
- Rattigan, R.; Sweeney, T.; Maher, S.; Thornton, K.; Rajauria, G.; O’Doherty, J.V. Laminarin-rich extract improves growth performance, small intestinal morphology, gene expression of nutrient transporters and the large intestinal microbial composition of piglets during the critical post-weaning period. Br. J. Nutr. 2019, 123, 255–263. [Google Scholar] [CrossRef]
- Ryan, M.; Smith, A.; O’Doherty, J.V.; Bahar, B.; Reilly, P.; Sweeney, T. Effects of nutrient supplementation with laminarin derived from Laminaria hyperborea and Laminaria digitata on mucin gene expression in the porcine ileum. Livest. Sci. 2010, 133, 236–238. [Google Scholar] [CrossRef]
- Smith, A.G.; O’Doherty, J.V.; Reilly, P.; Ryan, M.; Bahar, B.; Sweeney, T. The effects of laminarin derived from Laminaria digitata on measurements of gut health: Selected bacterial populations, intestinal fermentation, mucin gene expression and cytokine gene expression in the pig. Br. J. Nutr. 2011, 105, 669–677. [Google Scholar] [CrossRef] [Green Version]
- Bouwhuis, M.A.; Sweeney, T.; Mukhopadhya, A.; Thornton, K.; McAlpine, P.O.; O’Doherty, J.V. Zinc methionine and laminarin have growth-enhancing properties in newly weaned pigs influencing both intestinal health and diarrhoea occurrence. J. Anim. Physiol. Anim. Nutr. 2016, 101, 1273–1285. [Google Scholar] [CrossRef]
- Ilium, L. Chitosan and Its Use as a Pharmaceutical Excipient. Pharm. Res. 1998, 15, 1326–1331. [Google Scholar] [CrossRef]
- Liu, G.; Chen, S.; Guan, G.; Tan, J.; Al-Dhabi, N.A.; Wang, H.; Duraipandiyan, V.; Fang, J. Chitosan Modulates Inflammatory Responses in Rats Infected with Enterotoxigenic Escherichia coli. Mediat. Inflamm. 2016, 2016, 7432845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chien, R.-C.; Yen, M.-T.; Mau, J. Antimicrobial and antitumor activities of chitosan from shiitake stipes, compared to commercial chitosan from crab shells. Carbohydr. Polym. 2016, 138, 259–264. [Google Scholar] [CrossRef]
- Wang, J.; Zhang, C.; Guo, C.; Li, X. Chitosan Ameliorates DSS-Induced Ulcerative Colitis Mice by Enhancing Intestinal Barrier Function and Improving Microflora. Int. J. Mol. Sci. 2019, 20, 5751. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yousef, M.; Pichyangkura, R.; Soodvilai, S.; Chatsudthipong, V.; Muanprasat, C. Chitosan oligosaccharide as potential therapy of inflammatory bowel disease: Therapeutic efficacy and possible mechanisms of action. Pharmacol. Res. 2012, 66, 66–79. [Google Scholar] [CrossRef] [PubMed]
- Azuma, K.; Osaki, T.; Kurozumi, S.; Kiyose, M.; Tsuka, T.; Murahata, Y.; Imagawa, T.; Itoh, N.; Minami, S.; Sato, K.; et al. Anti-inflammatory effects of orally administered glucosamine oligomer in an experimental model of inflammatory bowel disease. Carbohydr. Polym. 2015, 115, 448–456. [Google Scholar] [CrossRef] [PubMed]
- Young, D.; Ibuki, M.; Nakamori, T.; Fan, M.; Mine, Y. Soy-Derived Di- and Tripeptides Alleviate Colon and Ileum Inflammation in Pigs with Dextran Sodium Sulfate-Induced Colitis. J. Nutr. 2011, 142, 363–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, E.R.; Ullrey, D.E. The Pig as a Model for Human Nutrition. Annu. Rev. Nutr. 1987, 7, 361–382. [Google Scholar] [CrossRef]
- Gonzalez, L.M.; Moeser, A.; Blikslager, A. Porcine models of digestive disease: The future of large animal translational research. Transl. Res. 2015, 166, 12–27. [Google Scholar] [CrossRef] [Green Version]
- Eichele, D.D.; Kharbanda, K.K. Dextran sodium sulfate colitis murine model: An indispensable tool for advancing our understanding of inflammatory bowel diseases pathogenesis. World J. Gastroenterol. 2017, 23, 6016–6029. [Google Scholar] [CrossRef]
- Ibuki, M.; Fukui, K.; Kanatani, H.; Mine, Y. Anti-Inflammatory Effects of Mannanase-Hydrolyzed Copra Meal in a Porcine Model of Colitis. J. Vet. Med. Sci. 2014, 76, 645–651. [Google Scholar] [CrossRef] [Green Version]
- Marin, D.E.; Marin, D.E.; Rotar, M.C.; Ropota, M.; Taranu, I. Bioactive compounds from dietary whole grape seed meal improved colonic inflammation via inhibition of MAPKs and NF-kB signaling in pigs with DSS induced colitis. J. Funct. Foods 2020, 66, 103708. [Google Scholar] [CrossRef]
- Morgan, X.C.; Tickle, T.L.; Sokol, H.; Gevers, D.; Devaney, K.L.; Ward, D.V.; Reyes, J.A.; Shah, S.A.; Leleiko, N.; Snapper, S.B.; et al. Dysfunction of the intestinal microbiome in inflammatory bowel disease and treatment. Genome Biol. 2012, 13, R79. [Google Scholar] [CrossRef] [PubMed]
- Papa, E.; Docktor, M.; Smillie, C.; Weber, S.; Preheim, S.P.; Gevers, D.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Ingram, J.; et al. Non-Invasive Mapping of the Gastrointestinal Microbiota Identifies Children with Inflammatory Bowel Disease. PLoS ONE 2012, 7, e39242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comito, D.; Cascio, A.; Romano, C. Microbiota biodiversity in inflammatory bowel disease. Ital. J. Pediatr. 2014, 40, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuo, T.; Ng, S.C. The Gut Microbiota in the Pathogenesis and Therapeutics of Inflammatory Bowel Disease. Front. Microbiol. 2018, 9, 2247. [Google Scholar] [CrossRef]
- Shen, Z.; Zhu, C.; Quan, Y.; Yang, J.; Yuan, W.; Yang, Z.; Wu, S.; Luo, W.; Tan, B.; Wang, X. Insights into Roseburia intestinalis which alleviates experimental colitis pathology by inducing anti-inflammatory responses. J. Gastroenterol. Hepatol. 2018, 33, 1751–1760. [Google Scholar] [CrossRef]
- Bednorz, C.; Oelgeschläger, K.; Kinnemann, B.; Hartmann, S.; Neumann, K.; Pieper, R.; Bethe, A.; Semmler, T.; Tedin, K.; Schierack, P.; et al. The broader context of antibiotic resistance: Zinc feed supplementation of piglets increases the proportion of multi-resistant Escherichia coli in vivo. Int. J. Med. Microbiol. 2013, 303, 396–403. [Google Scholar] [CrossRef]
- Diether, N.E.; Willing, B. Microbial Fermentation of Dietary Protein: An Important Factor in Diet–Microbe–Host Interaction. Microorganisms 2019, 7, 19. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y. Fatty acids, inflammation and intestinal health in pigs. J. Anim. Sci. Biotechnol. 2015, 6, 41. [Google Scholar] [CrossRef] [Green Version]
- Besten, G.D.; Van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [Green Version]
- Litvak, Y.; Byndloss, M.X.; Tsolis, R.M.; Bäumler, A.J. Dysbiotic Proteobacteria expansion: A microbial signature of epithelial dysfunction. Curr. Opin. Microbiol. 2017, 39, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Berry, D.; Schwab, C.; Milinovich, G.J.; Reichert, J.; Ben Mahfoudh, K.; Decker, T.; Engel, M.; Hai, B.; Hainzl, E.; Heider, S.; et al. Phylotype-level 16S rRNA analysis reveals new bacterial indicators of health state in acute murine colitis. ISME J. 2012, 6, 2091–2106. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishna, B.S.; Jayakanthan, P.; Pugazhendhi, S.; Kabeerdoss, J. Alterations of mucosal microbiota in the colon of patients with inflammatory bowel disease revealed by real time polymerase chain reaction amplification of 16S ribosomal ribonucleic acid. Indian J. Med Res. 2015, 142, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Mylonaki, M.; Rayment, N.B.; Rampton, D.S.; Hudspith, B.N.; Brostoff, J. Molecular characterization of rectal mucosa-associated bacterial flora in inflammatory bowel disease. Inflamm. Bowel Dis. 2005, 11, 481–487. [Google Scholar] [CrossRef]
- Baumgart, M.; Dogan, B.; Rishniw, M.; Weitzman, G.; Bosworth, B.; Yantiss, R.; Orsi, R.H.; Wiedmann, M.; McDonough, P.; Kim, S.G.; et al. Culture independent analysis of ileal mucosa reveals a selective increase in invasive Escherichia coli of novel phylogeny relative to depletion of Clostridiales in Crohn’s disease involving the ileum. ISME J. 2007, 1, 403–418. [Google Scholar] [CrossRef] [Green Version]
- Darfeuille-Michaud, A.; Boudeau, J.; Bulois, P.; Christel, N.; Glasser, A.-L.; Barnich, N.; Bringer, M.-A.; Swidsinski, A.; Beaugerie, L.; Colombel, J.-F. High prevalence of adherent-invasive Escherichia coli associated with ileal mucosa in Crohn’s disease. Gastroenterology 2004, 127, 412–421. [Google Scholar] [CrossRef]
- Khan, S.; Imran, A.; Malik, A.; Chaudhary, A.A.; Rub, A.; Jan, A.T.; Syed, J.B.; Rolfo, C. Bacterial imbalance and gut pathologies: Association and contribution of E. coli in inflammatory bowel disease. Crit. Rev. Clin. Lab. Sci. 2018, 56, 1–17. [Google Scholar] [CrossRef]
- O’Shea, C.J.; O’Doherty, J.V.; Callanan, J.J.; Doyle, D.; Thornton, K.; Sweeney, T. The effect of algal polysaccharides laminarin and fucoidan on colonic pathology, cytokine gene expression and Enterobacteriaceae in a dextran sodium sulfate-challenged porcine model. J. Nutr. Sci. 2016, 5. [Google Scholar] [CrossRef] [Green Version]
- Vigors, S.; O’Doherty, J.V.; Rattigan, R.; McDonnell, M.J.; Rajauria, G.; Sweeney, T. Effect of a Laminarin Rich Macroalgal Extract on the Caecal and Colonic Microbiota in the Post-Weaned Pig. Mar. Drugs 2020, 18, 157. [Google Scholar] [CrossRef] [Green Version]
- O’Doherty, J.V.; McDonnell, P.; Figat, S. The effect of dietary laminarin and fucoidan in the diet of the weanling piglet on performance and selected faecal microbial populations. Livest. Sci. 2010, 134, 208–210. [Google Scholar] [CrossRef]
- Nishida, A.; Inoue, R.; Inatomi, O.; Bamba, S.; Naito, Y.; Andoh, A. Gut microbiota in the pathogenesis of inflammatory bowel disease. Clin. J. Gastroenterol. 2017, 11, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heller, F.; Florian, P.; Bojarski, C.; Richter, J.; Christ, M.; Hillenbrand, B.; Mankertz, J.; Gitter, A.H.; Bürgel, N.; Fromm, M. Interleukin-13 Is the Key Effector Th2 Cytokine in Ulcerative Colitis That Affects Epithelial Tight Junctions, Apoptosis, and Cell Restitution. Gastroenterology 2005, 129, 550–564. [Google Scholar] [CrossRef] [PubMed]
- Nemeth, Z.H.; Bogdanovski, D.A.; Barratt-Stopper, P.; Paglinco, S.R.; Antonioli, L.; Rolandelli, R.H. Crohn’s Disease and Ulcerative Colitis Show Unique Cytokine Profiles. Cureus 2017, 9, e1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirsattari, D.; Seyyedmajidi, M.; Zojaji, H.; Haghazali, M.; Orimi, P.G.; Shoushtarizadeh, T.; Almasi, S. The relation between the level of interleukin-23 with duration and severity of ulcerative colitis. Gastroenterol. Hepatol. Bed Bench 2012, 5, 49–53. [Google Scholar]
- Duvallet, E.; Semerano, L.; Assier, E.; Falgarone, G.; Boissier, M.-C. Interleukin-23: A key cytokine in inflammatory diseases. Ann. Med. 2011, 43, 503–511. [Google Scholar] [CrossRef]
- Wang, Y.-D.; Mao, J.-W. Expression of matrix metalloproteinase-1 and tumor necrosis factor-α in ulcerative colitis. World J. Gastroenterol. 2007, 13, 5926–5932. [Google Scholar] [CrossRef]
- O’shea, N.R.; Smith, A.M. Matrix Metalloproteases Role in Bowel Inflammation and Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2014, 20, 2379–2393. [Google Scholar] [CrossRef]
- Hu, S.; Wang, Y.; Wen, X.; Wang, L.; Jiang, Z.; Zheng, C. Effects of low-molecular-weight chitosan on the growth performance, intestinal morphology, barrier function, cytokine expression and antioxidant system of weaned piglets. BMC Vet. Res. 2018, 14, 215. [Google Scholar] [CrossRef] [Green Version]
- Sweeney, T.; O’Doherty, J.V. Marine macroalgal extracts to maintain gut homeostasis in the weaning piglet. Domest. Anim. Endocrinol. 2016, 56, S84–S89. [Google Scholar] [CrossRef]
- Fong, D.; Hoemann, C.D. Chitosan immunomodulatory properties: Perspectives on the impact of structural properties and dosage. Futur. Sci. OA 2018, 4, FSO225. [Google Scholar] [CrossRef] [Green Version]
- Jankowski, J.A.; Bedford, F.K.; Boulton, R.A.; Cruickshank, N.; Hall, C.; Elder, J.; Allan, R.; Forbes, A.; Kim, Y.S.; Wright, N.A.; et al. Alterations in classical cadherins associated with progression in ulcerative and Crohn’s colitis. Lab. Investig. 1998, 78, 1155–1167. [Google Scholar] [PubMed]
- Gassler, N.; Rohr, C.; Schneider, A.; Kartenbeck, J.; Bach, A.; Obermüller, N.; Otto, H.F.; Autschbach, F. Inflammatory bowel disease is associated with changes of enterocytic junctions. Am. J. Physiol. Liver Physiol. 2001, 281, G216–G228. [Google Scholar] [CrossRef] [PubMed]
- Kucharzik, T.; Walsh, S.V.; Chen, J.; Parkos, C.A.; Nusrat, A. Neutrophil Transmigration in Inflammatory Bowel Disease Is Associated with Differential Expression of Epithelial Intercellular Junction Proteins. Am. J. Pathol. 2001, 159, 2001–2009. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Shen, J.; Ran, Z. Epithelial–mesenchymal transition in Crohn’s disease. Mucosal Immunol. 2017, 11, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Castaneda, F.E.; Walia, B.; Vijay-Kumar, M.; Patel, N.R.; Roser, S.; Kolachala, V.L.; Rojas, M.; Wang, L.; Oprea, G.; Garg, P.; et al. Targeted Deletion of Metalloproteinase 9 Attenuates Experimental Colitis in Mice: Central Role of Epithelial-Derived MMP. Gastroenterology 2005, 129, 1991–2008. [Google Scholar] [CrossRef] [PubMed]
- Stokes, C.R. The development and role of microbial-host interactions in gut mucosal immune development. J. Anim. Sci. Biotechnol. 2017, 8, 12. [Google Scholar] [CrossRef] [Green Version]
- Pluske, J. Feed- and feed additives-related aspects of gut health and development in weanling pigs. J. Anim. Sci. Biotechnol. 2013, 4, 1. [Google Scholar] [CrossRef] [Green Version]
- Xiao, D.; Tang, Z.; Yin, Y.; Zhang, B.; Hu, X.; Feng, Z.; Wang, J. Effects of dietary administering chitosan on growth performance, jejunal morphology, jejunal mucosal sIgA, occluding, claudin-1 and TLR4 expression in weaned piglets challenged by enterotoxigenic Escherichia coli. Int. Immunopharmacol. 2013, 17, 670–676. [Google Scholar] [CrossRef]
- Garcia-Vaquero, M.; O’Doherty, J.V.; Tiwari, B.; Sweeney, T.; Rajauria, G. Enhancing the Extraction of Polysaccharides and Antioxidants from Macroalgae Using Sequential Hydrothermal-Assisted Extraction Followed by Ultrasound and Thermal Technologies. Mar. Drugs 2019, 17, 457. [Google Scholar] [CrossRef] [Green Version]
- NRC. Nutrient Requirements of Swine, 11th ed.; National Academies Press: Washington, DC, USA, 2012. [Google Scholar]
- Sauvant, D.; Perez, J.M.; Tran, G. Table of Composition and Nutritional Value of Feed Materials: Pigs, Poultry, Cattle, Sheep, Goats, Rabbits, Horses, Fish; Wageningen Academic Publishers: Wageningen, The Netherlands, 2004. [Google Scholar]
- Pierce, K.M.; Callan, J.J.; McCarthy, P.; O’Doherty, J.V. Performance of weanling pigs offered low or high lactose diets supplemented with avilamycin or inulin. Anim. Sci. 2005, 80, 313–318. [Google Scholar] [CrossRef]
- Van Soest, P.; Robertson, J.; Lewis, B. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Brown, C.; Baker, D.C.; Barker, I.K. Alimentary System. In Jubb, Kennedy, and Palmer’s Pathology of Domestic Animals, 5th ed.; Maxie, M., Ed.; Saunders Elsevie: Philadelphia, PA, USA, 2007; Volume 2, pp. 107–113. [Google Scholar]
- Cummins, E.; Seeballuck, F.; Keely, S.; Mangan, N.; Callanan, J.J.; Fallon, P.; Taylor, C.T. The Hydroxylase Inhibitor Dimethyloxalylglycine Is Protective in a Murine Model of Colitis. Gastroenterology 2008, 134, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Pierce, K.; Sweeney, T.; Callan, J.; Byrne, C.; McCarthy, P.; O’Doherty, J. The effect of inclusion of a high lactose supplement in finishing diets on nutrient digestibility, nitrogen excretion, volatile fatty acid concentrations and ammonia emission from boars. Anim. Feed Sci. Technol. 2006, 125, 45–60. [Google Scholar] [CrossRef]
- Cole, J.R.; Chai, B.; Farris, R.J.; Wang, Q.; Kulam, S.A.; McGarrell, D.M.; Garrity, G.M.; Tiedje, J.M. The Ribosomal Database Project (RDP-II): Sequences and tools for high-throughput rRNA analysis. Nucleic Acids Res. 2004, 33, D294–D296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reeder, J.; Knight, R. Rapidly denoising pyrosequencing amplicon reads by exploiting rank-abundance distributions. Nat. Methods 2010, 7, 668–669. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2012, 41, D590–D596. [Google Scholar] [CrossRef]
- Caporaso, J.G.; Bittinger, K.; Bushman, F.D.; DeSantis, T.Z.; Andersen, G.; Knight, R. PyNAST: A flexible tool for aligning sequences to a template alignment. Bioinformatics 2009, 26, 266–267. [Google Scholar] [CrossRef] [Green Version]
- McMurdie, P.; Holmes, S. phyloseq: An R Package for Reproducible Interactive Analysis and Graphics of Microbiome Census Data. PLoS ONE 2013, 8, e61217. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friendly, M. Corrgrams. Am. Stat. 2002, 56, 316–324. [Google Scholar] [CrossRef]
- Wei, T.; Simko, V. R Package “Corrplot”: Visualization of a Correlation Matrix, R package version 0.84; Available online: https://github.com/taiyun/corrplot (accessed on 15 April 2020).



{kind=link}
{kind=link}
| Treatment | SEM | |||
|---|---|---|---|---|
| Basal | Laminarin | Chitosan | ||
| Initial BW (kg) | 8.93 | 9.25 | 9.17 | 0.169 |
| BW d35 (kg) | 21.71 | 20.64 | 22.19 | 0.729 |
| ADG d0-35 (kg/day) | 0.360 | 0.326 | 0.370 | 0.016 |
| ADFI d0-35 (kg/day) | 0.743 a | 0.674 b | 0.705 ab | 0.020 |
| G:F d0-35 (kg/kg) | 0.743 | 0.674 | 0.705 | 0.020 |
| Treatment | SEM | Contrasts | ||||||
|---|---|---|---|---|---|---|---|---|
| Control | Basal DSS | Laminarin DSS | Chitosan DSS | 1 | 2 | 3 | ||
| Final BW (kg) | 26.35 | 24.47 | 24.76 | 24.77 | 0.382 | ** | NS | NS |
| ADG (kg/day) | 0.499 | 0.289 | 0.321 | 0.324 | 0.042 | ** | NS | NS |
| ADFI (kg/day) | 1.42 | 1.16 | 1.02 | 1.17 | 0.059 | ** | NS | NS |
| G:F (kg/kg) | 0.356 | 0.233 | 0.317 | 0.278 | 0.034 | * | NS | NS |
| FS | 2.14 | 2.43 | 2.76 | 2.83 | 0.066 | ** | *** | *** |
| Treatment | SEM | Contrasts | ||||||
|---|---|---|---|---|---|---|---|---|
| Control | Basal DSS | Laminarin DSS | Chitosan DSS | 1 | 2 | 3 | ||
| Pathology score | 1.0 | 2.0 | 1.7 | 1.9 | 0.330 | * | NS | NS |
| Goblet cell % | 14.9 | 27.02 | 23.12 | 22.54 | 2.72 | ** | NS | NS |
| Epithelial cell % | 85.1 | 72.99 | 76.88 | 77.46 | 2.72 | ** | NS | NS |
| Treatment | SEM | Contrasts | ||||||
|---|---|---|---|---|---|---|---|---|
| Control | Basal DSS | Laminarin DSS | Chitosan DSS | 1 | 2 | 3 | ||
| MMP1 | 0.245 | 2.142 | 0.857 | 2.371 | 0.646 | * | NS | NS |
| IL13 | 0.634 | 1.033 | 0.916 | 0.870 | 0.103 | * | NS | NS |
| IL23A | 0.487 | 0.975 | 1.052 | 0.740 | 0.153 | * | NS | NS |
| MMP2 | 0.476 | 1.113 | 0.945 | 1.079 | 0.268 | NS | NS | NS |
| MMP9 | 0.525 | 1.065 | 0.758 | 1.164 | 0.256 | NS | NS | NS |
| CLEC7A | 0.929 | 1.453 | 0.943 | 1.236 | 0.330 | NS | NS | NS |
| TLR4 | 0.904 | 1.046 | 1.554 | 1.359 | 0.350 | NS | NS | NS |
| TJP1 | 0.895 | 0.930 | 0.867 | 0.924 | 0.056 | NS | NS | NS |
| CDH1 | 0.891 | 1.122 | 1.017 | 1.022 | 0.112 | NS | NS | NS |
| MARVELD2 | 1.480 | 1.566 | 1.688 | 1.765 | 0.451 | NS | NS | NS |
| IL10 | 0.587 | 0.906 | 1.019 | 0.962 | 0.159 | NS | NS | NS |
| IL17A | 0.808 | 0.747 | 1.157 | 1.217 | 0.288 | NS | NS | NS |
| IL6 | 0.310 | 1.725 | 1.356 | 1.269 | 0.549 | NS | NS | NS |
| IL1B | 0.428 | 0.905 | 1.170 | 1.451 | 0.322 | NS | NS | NS |
| IFNG | 1.457 | 0.899 | 1.081 | 1.243 | 0.425 | NS | NS | NS |
| TGFB1 | 0.925 | 0.870 | 0.913 | 0.944 | 0.156 | NS | NS | NS |
| MUC4 | 1.393 | 0.780 | 0.931 | 1.337 | 0.247 | NS | NS | NS |
| Treatment | SEM | Contrasts | ||||||
|---|---|---|---|---|---|---|---|---|
| Control | Basal DSS | Laminarin DSS | Chitosan DSS | 1 | 2 | 3 | ||
| Total VFA(mmol/g digesta) | 152.58 | 169.00 | 158.29 | 169.83 | 6.910 | NS | NS | NS |
| Molar proportions | ||||||||
| Acetate | 0.64 | 0.57 | 0.62 | 0.58 | 0.014 | ** | * | NS |
| Propionate | 0.185 | 0.218 | 0.196 | 0.222 | 0.010 | * | NS | NS |
| Isobutyrate | 0.012 | 0.013 | 0.012 | 0.011 | 0.002 | NS | NS | NS |
| Butyrate | 0.111 | 0.123 | 0.110 | 0.124 | 0.006 | NS | NS | NS |
| Isovalerate | 0.012 | 0.014 | 0.012 | 0.012 | 0.002 | NS | NS | NS |
| Valerate | 0.039 | 0.061 | 0.051 | 0.054 | 0.006 | ** | NS | NS |
| Branched | 0.063 | 0.088 | 0.075 | 0.077 | 0.007 | ** | NS | NS |
| Measure of Diversity | Treatment | SEM | Contrasts a | |||||
|---|---|---|---|---|---|---|---|---|
| Control | Basal DSS | Laminarin DSS | Chitosan DSS | 1 | 2 | 3 | ||
| Observed | 195.80 | 128.17 | 163.75 | 160.80 | 7.13 | * | * | NS |
| Chao1 | 288.56 | 196.58 | 229.36 | 229.06 | 9.53 | * | NS | NS |
| ACE | 290.97 | 204.62 | 236.14 | 238.28 | 8.93 | * | NS | NS |
| Shannon | 3.74 | 3.12 | 3.42 | 3.31 | 0.09 | * | NS | NS |
| Simpson | 0.93 | 0.86 | 0.91 | 0.89 | 0.01 | * | NS | NS |
| InvSimpson | 18.78 | 12.91 | 15.34 | 12.23 | 1.52 | NS | NS | NS |
| Fisher | 51.93 | 33.54 | 41.63 | 41.44 | 2.05 | ** | NS | NS |
| Taxa | OTU | Treatment | SEM | Contrasts a | |||||
|---|---|---|---|---|---|---|---|---|---|
| Control | Basal DSS | Laminarin DSS | Chitosan DSS | 1 | 2 | 3 | |||
| P-Deferribacteres | AF059190.1.1451 | 0.07 | 0.78 | 0.27 | 0.90 | 0.120 | *** | NS | NS |
| P-Cyanobacteria | EU474510.1.1379 | 0.68 | 0 | 0.01 | 0.02 | 0.097 | *** | NS | NS |
| F-Lachnospiraceae | New.CleanUp.ReferenceOTU8 | 3.722 | 1.281 | 1.830 | 1.476 | 0.239 | * | NS | NS |
| F-GR-WP33-58 | AF371949.1.1454 | 0.851 | 0.000 | 0.000 | 0.014 | 0.094 | *** | NS | NS |
| F-Synergistaceae | JQ606918.1.1410 | 0.003 | 0.117 | 1.357 | 0.163 | 0.225 | * | NS | NS |
| F-Uncultured bacterium | DQ805702.1.1378 | 0.071 | 0.000 | 0.018 | 0.004 | 0.009 | *** | NS | NS |
| F-Clostridiales vadin BB60 group | HQ780759.1.1426 | 0.416 | 0.466 | 1.186 | 1.465 | 0.174 | NS | NS | * |
| G-Phascolarctobacterium | GQ358246.1.1466 | 1.011 | 0.000 | 0.178 | 0.097 | 0.115 | ** | NS | NS |
| G-[Eubacterium] ruminantium group | EU775472.1.1384 | 0.514 | 0.039 | 0.138 | 0.012 | 0.072 | ** | NS | NS |
| G-Oribacterium | EU461618.1.1389 | 0.283 | 0.004 | 0.000 | 0.012 | 0.025 | ** | NS | NS |
| G-Roseburia | EU466950.1.1385 | 1.324 | 0.115 | 0.179 | 0.243 | 0.097 | *** | NS | NS |
| G-Anaerovibrio | New.CleanUp.ReferenceOTU1 | 4.293 | 0.232 | 1.390 | 1.056 | 0.439 | *** | NS | NS |
| G-Bacteroides | KF842513.1.1417 | 0.000 | 0.413 | 0.455 | 0.210 | 0.081 | *** | NS | NS |
| G-Rikenellaceae RC9 gut group | New.CleanUp.ReferenceOTU14 | 1.406 | 0.238 | 0.585 | 0.268 | 0.165 | * | NS | NS |
| G-Prevotella 1 | GQ448219.1.1401 | 0.270 | 0.008 | 0.007 | 0.000 | 0.039 | * | NS | NS |
| G-Prevotella 7 | EU458732.1.1352 | 0.444 | 0.004 | 0.000 | 0.000 | 0.048 | * | NS | NS |
| G-Uncultured bacterium | EF445225.1.1490 | 0.382 | 0.012 | 0.045 | 0.057 | 0.047 | ** | NS | NS |
| G-Thalassospira | New.CleanUp.ReferenceOTU23 | 0.463 | 0.000 | 0.009 | 0.093 | 0.047 | ** | NS | NS |
| G-Escherichia-shigella | KF842027.1.1375 | 0.538 | 2.626 | 0.405 | 0.594 | 0.362 | ** | ** | NS |
| G-Campylobacter | AF550648.1.1562 | 0.019 | 0.283 | 0.054 | 0.104 | 0.050 | ** | NS | NS |
| G-Mucispirillum | AF059190.1.1451 | 0.074 | 0.799 | 0.273 | 0.913 | 0.122 | *** | NS | NS |
| G-Uncultured bacteria | HQ780759.1.1426 | 0.407 | 0.474 | 1.206 | 1.458 | 0.176 | NS | * | * |
| G-Uncultured bacterium | JQ184479.1.1344 | 0.542 | 0.004 | 0.008 | 0.022 | 0.067 | *** | NS | NS |
| G-Succinivibrio | KF843489.1.1384 | 3.745 | 1.117 | 9.575 | 1.261 | 1.078 | NS | * | NS |
| G-Succinivibrionaceae UCG-001 | HQ400282.1.1506 | 13.036 | 28.345 | 0.062 | 13.760 | 2.736 | NS | *** | NS |
| G-Olsenella | JQ188553.1.1334 | 0.099 | 0.174 | 0.000 | 0.012 | 0.029 | NS | * | NS |
| Ingredient | g/kg |
|---|---|
| Whey powder | 50.0 |
| Wheat | 380.0 |
| Barley | 234.0 |
| Soya bean meal | 170.0 |
| Full-fat soya bean | 120.0 |
| Soya oil | 10.0 |
| Vitamins and minerals b | 3.0 |
| Salt | 3.0 |
| Dicalcium phosphate | 13.0 |
| Limestone | 11.0 |
| Lysine HCL | 4.0 |
| DL-methionine | 1.5 |
| L-threonine | 1.5 |
| Chemical analysis | |
| Dry matter | 866.1 |
| Crude protein (N × 6.25) | 210.6 |
| Ash | 48.4 |
| Neutral detergent fibre | 115.1 |
| Digestible energy (MJ/kg) * | 14.5 |
| Lysine * | 14.5 |
| Methionine and cysteine * | 8.4 |
| Threonine * | 9.1 |
| Tryptophan * | 2.5 |
| Calcium * | 9.5 |
| Phosphorous * | 6.1 |
| Genes | Accession No. | Forward Primer (5’-3’) Reverse Primer (5’-3’) | Tm |
|---|---|---|---|
| Cytokines | |||
| IL23A | NM_001130236.1 | F:AGGGACTCAGGGACAACAGTC | 61.1 |
| R:GCGAAGGATCTTGAGGCGGAGAAGGAG | 68.42 | ||
| IL13 | NM_213803.1 | F:CCTGGAATCCCTCATCAACAT | 57.41 |
| R:AGGGCGCTCAGGATCCT | 59.66 | ||
| IL10 | NM_214041.1 | F:GCCTTCGGCCCAGTGAA | 57.6 |
| R:AGAGACCCGGTCAGCAACAA | 59.4 | ||
| IL6 | NM_214399.1 | F:GACAAAGCCACCACCCCTAA | 59.8 |
| R:CTCGTTCTGTGACTGCAGCTTATC | 62.7 | ||
| IFNG | NM_213948.1 | F:TCTAACCTAAGAAAGCGGAAGAGA | 61.1 |
| R:TTGCAGGCATGACAATTA | 61.5 | ||
| IL1B | NM_214055.1 | F:TTGAATTCGAGTCTGCCCTGT | 59.65 |
| R:CCCAGGAAGACGGGCTTT | 59.24 | ||
| TGFB1 | NM_214015.1 | F:AGGGCTACCATGCCAATTTCT | 59.71 |
| R:CGGGTTGTGCTGGTTGTACA | 60.82 | ||
| IL17A | NM_001005729.1 | F:CCCTGTCACTGCTGCTTCTG | 60.95 |
| R:TCATGATTCCCGCCTTCAC | 57.52 | ||
| Matrix metalloproteinase’s | |||
| MMP2 | NM_214192.2 | F:CTTCAAGGGCGCGTATTACC | 59.06 |
| R:GCCAGTCGGATTTGATGCTT | 58.9 | ||
| MMP9 | NM_001038004.1 | F:GGCTACAGCCTGTTCCTTGTG | 61.22 |
| R:GGCACGGTTGAGTGATCTAAGC | 61.56 | ||
| MMP1 | NM_001166229.1 | F:GGACCGTGCCATTGAGAA | 57.29 |
| R:CCTCGGAGACCTTGGTGAAC | 60.04 | ||
| Tight junctions | |||
| TJP1 | XM_005659811.1 | F:TGAGAGCCAACCATGTCTTGAA | 59.9 |
| R:CTCAGACCCTCTCTGTCT | 60.0 | ||
| CDH1 | NM_001163060.1 | F:GCCTGGCAACTGAGCTGACT | 62.74 |
| R:CCTCCCTCCTTCAGAATTTTC | 55.93 | ||
| MARVELD2 | NM_001243948.1 | F: CAGGCACCACGACGAAGTC | 61.02 |
| R: AAGCGTCTGGAAGGTTCTTACG | 60.61 | ||
| Mucin | |||
| MUC4 | XM_001926442.1 | F:GATGCCCTGGCCACAGAA | 63.3 |
| R:TGATTCAAGGTAGCATTCATTTGC | 62.4 | ||
| Pattern recognition receptors | |||
| TLR4 | NM_001293317.1 | F:TGCATGGAGCTGAATTTCTACAA | 58.6 |
| R:GATAATCCAGCACCTGCAGTTC | 59.9 | ||
| CLEC7A | NM_001145866.1 | F:GCACATCATTAGCTTCCTGGAA | 58.72 |
| R:GGAGCTGTCTATCTTCAGGAGA | 58.44 | ||
| Reference genes | |||
| PPIA | NM_214353.1 | F:CGGTCCTGGCATCTTGT | 59.67 |
| R:TGGCAGTGCAAATGAAAAACTG | 59.61 | ||
| ACTB | AY550069.1 | F:CAAATGCTTCTAGGCGGCGGACTGT | 60.9 |
| R:TCTCATTTTCTGCGCAAGTTAGG | 59.5 | ||
| Primer ID | Forward Primer 5’-3’ | Unique 6 Nucleotide Tag |
|---|---|---|
| PA-V4-1 | CCATCTCATCCCTGCGTGTCTCCGACTCAG | AGCAGC |
| PA-V4-2 | CCATCTCATCCCTGCGTGTCTCCGACTCAG | CTCAGC |
| PA-V4-3 | CCATCTCATCCCTGCGTGTCTCCGACTCAG | AGAGAG |
| PA-V4-4 | CCATCTCATCCCTGCGTGTCTCCGACTCAG | AGATGC |
| PA-V4-5 | CCATCTCATCCCTGCGTGTCTCCGACTCAG | AGCATG |
| PA-V4-6 | CCATCTCATCCCTGCGTGTCTCCGACTCAG | ATCATC |
| PA-V4-7 | CCATCTCATCCCTGCGTGTCTCCGACTCAG | ATCTGC |
| PA-V4-8 | CCATCTCATCCCTGCGTGTCTCCGACTCAG | ATGAGC |
| PA-V4-9 | CCATCTCATCCCTGCGTGTCTCCGACTCAG | ATGATG |
| PA-V4-10 | CCATCTCATCCCTGCGTGTCTCCGACTCAG | ATGCAG |
| PA-V4-11 | CCATCTCATCCCTGCGTGTCTCCGACTCAG | ATGCTC |
| PA-V4-12 | CCATCTCATCCCTGCGTGTCTCCGACTCAG | CAGAGC |
| PA-V4-13 | CCATCTCATCCCTGCGTGTCTCCGACTCAG | CAGATG |
| PA-V4-14 | CCATCTCATCCCTGCGTGTCTCCGACTCAG | CAGCAG |
| PA-V4-15 | CCATCTCATCCCTGCGTGTCTCCGACTCAG | CAGCTC |
| PA-V4-16 | CCATCTCATCCCTGCGTGTCTCCGACTCAG | CATCTG |
| PA-V4-17 | CCATCTCATCCCTGCGTGTCTCCGACTCAG | CATGAG |
| PA-V4-18 | CCATCTCATCCCTGCGTGTCTCCGACTCAG | CTCATG |
| PA-V4-19 | CCATCTCATCCCTGCGTGTCTCCGACTCAG | CTGATC |
| PA-V4-20 | CCATCTCATCCCTGCGTGTCTCCGACTCAG | CTGCTG |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rattigan, R.; O’Doherty, J.V.; Vigors, S.; Ryan, M.T.; Sebastiano, R.S.; Callanan, J.J.; Thornton, K.; Rajauria, G.; Margassery, L.M.; Dobson, A.D.W.; et al. The Effects of the Marine-Derived Polysaccharides Laminarin and Chitosan on Aspects of Colonic Health in Pigs Challenged with Dextran Sodium Sulphate. Mar. Drugs 2020, 18, 262. https://doi.org/10.3390/md18050262
Rattigan R, O’Doherty JV, Vigors S, Ryan MT, Sebastiano RS, Callanan JJ, Thornton K, Rajauria G, Margassery LM, Dobson ADW, et al. The Effects of the Marine-Derived Polysaccharides Laminarin and Chitosan on Aspects of Colonic Health in Pigs Challenged with Dextran Sodium Sulphate. Marine Drugs. 2020; 18(5):262. https://doi.org/10.3390/md18050262
Chicago/Turabian StyleRattigan, Ruth, John V. O’Doherty, Stafford Vigors, Marion T. Ryan, Rocco S. Sebastiano, John J. Callanan, Kevin Thornton, Gaurav Rajauria, Lekha M. Margassery, Alan D. W. Dobson, and et al. 2020. "The Effects of the Marine-Derived Polysaccharides Laminarin and Chitosan on Aspects of Colonic Health in Pigs Challenged with Dextran Sodium Sulphate" Marine Drugs 18, no. 5: 262. https://doi.org/10.3390/md18050262
APA StyleRattigan, R., O’Doherty, J. V., Vigors, S., Ryan, M. T., Sebastiano, R. S., Callanan, J. J., Thornton, K., Rajauria, G., Margassery, L. M., Dobson, A. D. W., O’Leary, N. D., & Sweeney, T. (2020). The Effects of the Marine-Derived Polysaccharides Laminarin and Chitosan on Aspects of Colonic Health in Pigs Challenged with Dextran Sodium Sulphate. Marine Drugs, 18(5), 262. https://doi.org/10.3390/md18050262