Litoralimycins A and B, New Cytotoxic Thiopeptides from Streptomonospora sp. M2




Abstract
:1. Introduction
2. Results
2.1. Screening
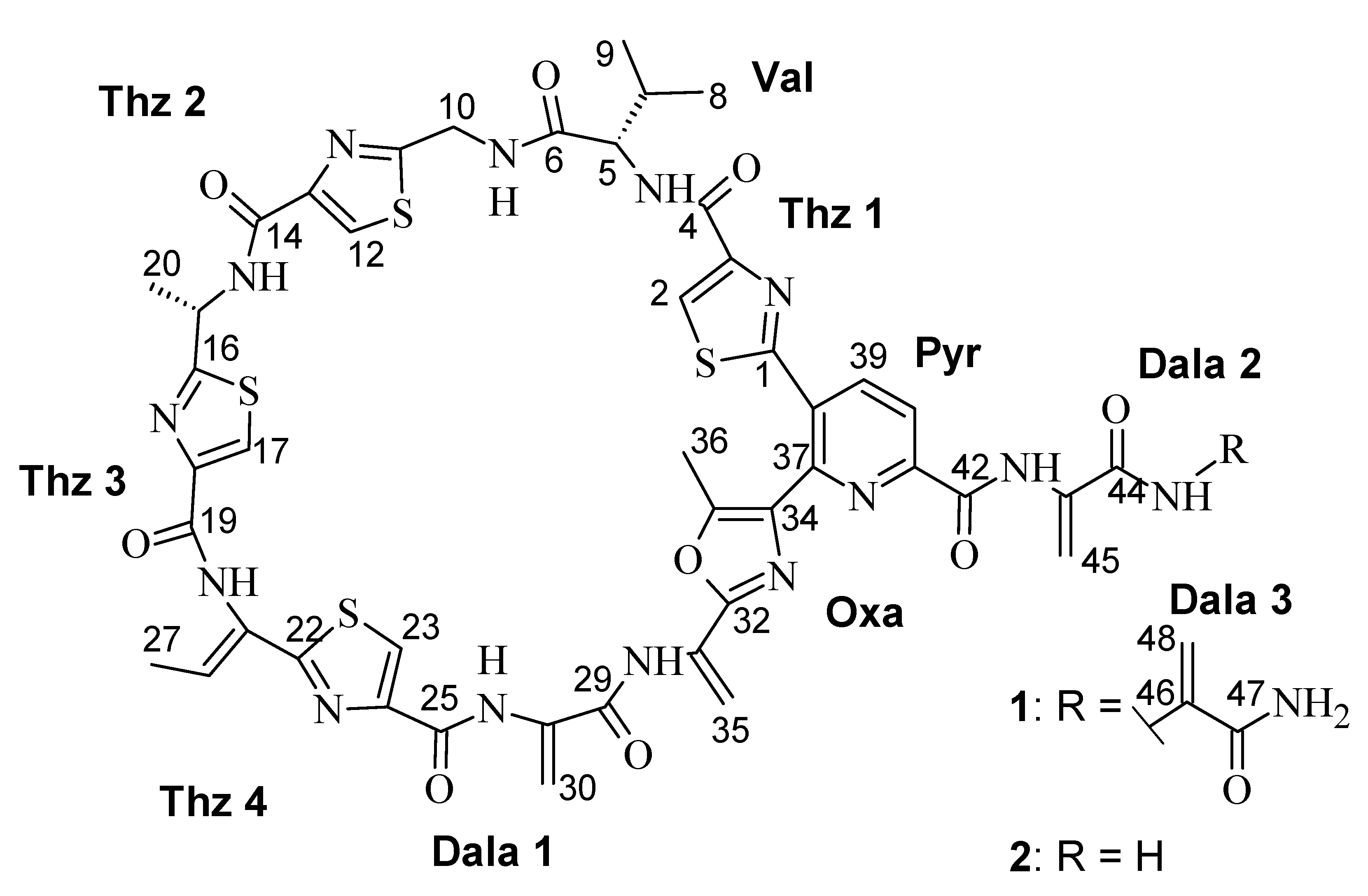
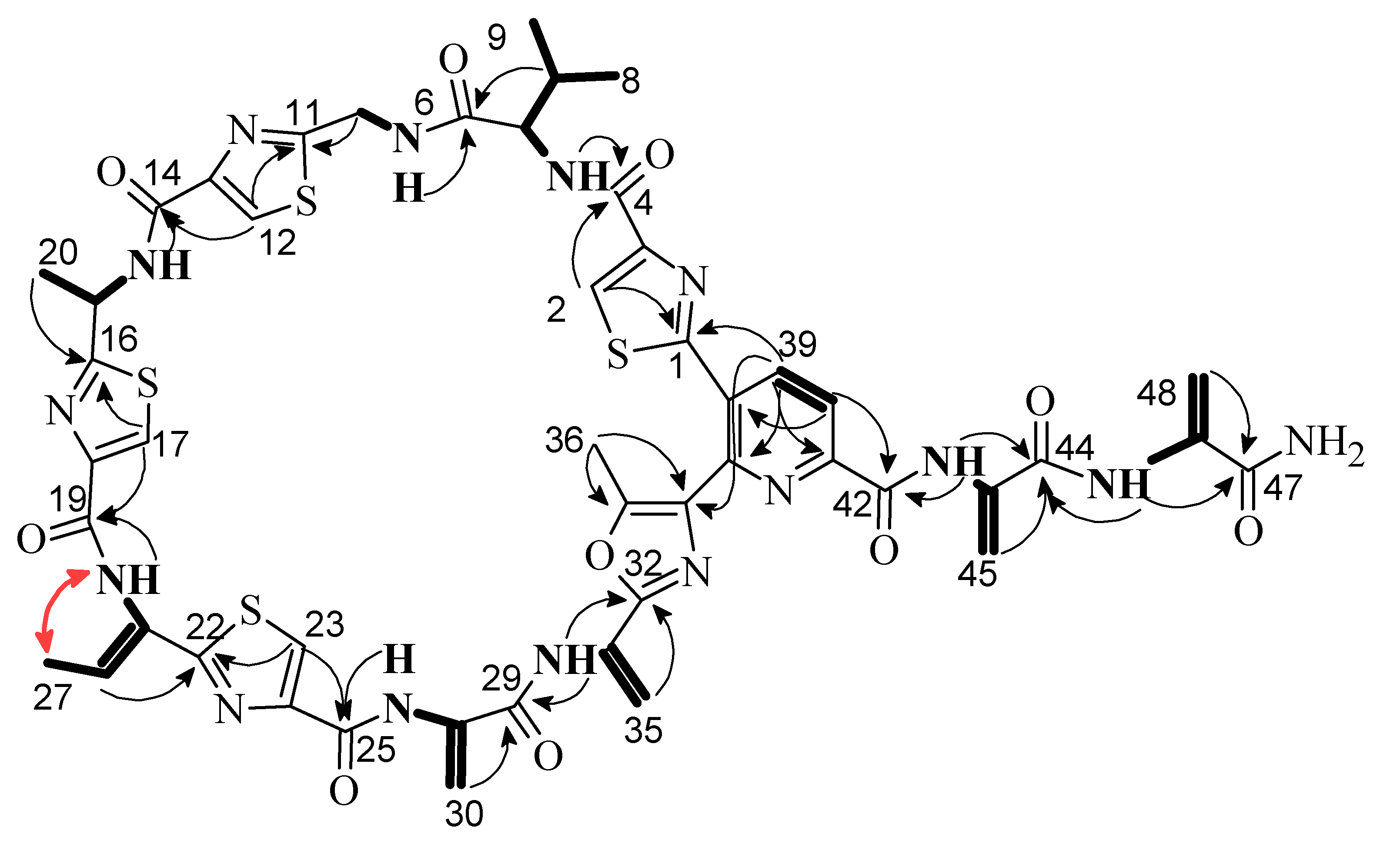
2.2. Structure Elucidation
2.3. Bioactivity
3. Discussion
4. Materials and Methods
4.1. General
4.2. Strain Maintenance
4.3. Fermentation, Extraction, and Isolation of Compounds
4.4. Ozonolysis, Hydrolysis and Marfey’s Derivatization with l-FDAA
4.5. Minimum Inhibitory Concentrations
4.6. Cytotoxicity Assay
4.7. HPLC Fractionation and Bioassays in 96-Well Plates
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Stadler, M.; Dersch, P. How to overcome the antibiotic crisis—Facts, challenges, technologies & future perspectives. Curr. Top Microbiol. Immunol. 2017, 398, 496. [Google Scholar]
- Genilloud, O. Actinomycetes: Still a source of novel antibiotics. Nat. Prod. Rep. 2017, 34, 1203–1232. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, K.; Gupta, R.K. Rare actinomycetes: A potential storehouse for novel antibiotics. Crit. Rev. Biotechnol. 2012, 32, 108–132. [Google Scholar] [CrossRef] [PubMed]
- Landwehr, W.; Wolf, C.; Wink, J. Actinobacteria and myxobacteria—Two of the most important bacterial resources for novel antibiotics. In How to Overcome the Antibiotic Crisis; Springer: Cham, Switzerland, 2016; pp. 273–302. [Google Scholar]
- Subramani, R.; Aalbersberg, W. Culturable rare Actinomycetes: Diversity, isolation and marine natural product discovery. Appl. Microbiol. Biotechnol. 2013, 97, 9291–9321. [Google Scholar] [CrossRef] [PubMed]
- Yun, B.-S.; Fujita, K.; Furihata, K.; Seto, H. Absolute stereochemistry and solution conformation of promothiocins. Tetrahedron 2001, 57, 9683–9687. [Google Scholar] [CrossRef]
- Marfey, P.; Ottensen, M. Determination of d-amino acids. I. Hydrolysis of DNP-l-amino acid methyl esters with carboxypeptidase-Y. Carlsberg Res. Commun. 1984, 49, 585–590. [Google Scholar] [CrossRef] [Green Version]
- Surup, F.; Halecker, S.; Nimtz, M.; Rodrigo, S.; Schulz, B.; Steinert, M.; Stadler, M. Hyfraxins A and B, cytotoxic ergostane-type steroid and lanostane triterpenoid glycosides from the invasive ash dieback ascomycete Hymenoscyphus fraxineus. Steroids 2018, 135, 92–97. [Google Scholar] [CrossRef] [PubMed]
- Shen, X.; Mustafa, M.; Chen, Y.; Cao, Y.; Gao, J. Natural thiopeptides as a privileged scaffold for drug discovery and therapeutic development. Med. Chem. Res. 2019, 28, 1063–1098. [Google Scholar] [CrossRef]
- Just-Baringo, X.; Albericio, F.; Álvarez, M. Thiopeptide antibiotics: Retrospective and recent advances. Mar. Drugs 2014, 12, 317–351. [Google Scholar] [CrossRef] [Green Version]
- Malcolmson, S.J.; Young, T.S.; Ruby, J.G.; Skewes-Cox, P.; Walsh, C.T. The posttranslational modification cascade to the thiopeptide berninamycin generates linear forms and altered macrocyclic scaffolds. Proc. Natl. Acad. Sci. USA 2013, 110, 8483–8488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lentzen, G.; Klinck, R.; Matassova, N.; Aboul-ela, F.; Murchie, A.I.H. Structural Basis for Contrasting Activities of Ribosome Binding Thiazole Antibiotics. Chem. Biol. 2003, 10, 769–778. [Google Scholar] [CrossRef] [Green Version]
- González Holgado, G.; Castro Rodríguez, J.; Cañedo Hernández, L.M.; Díaz, M.; Fernández-Abalos, J.M.; Trujillano, I.; Santamaría, R.I. Radamycin, a novel thiopeptide produced by Streptomyces sp. RSP9. I. Taxonomy, fermentation, isolation and biological activities. J. Antibiot. 2002, 55, 383–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro Rodríguez, J.; González Holgado, G.; Santamaría Sánchez, R.I.; Cañedo, L.M. Radamycin, a novel thiopeptide produced by Streptomyces sp. RSP9. II. Physico-chemical properties and structure determination. J. Antibiot. 2002, 55, 391–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chiu, M.L.; Folcher, M.; Katoh, T.; Puglia, A.M.; Vohradsky, J.; Yun, B.S.; Seto, H.; Thompson, C.J. Broad spectrum thiopeptide recognition specificity of the Streptomyces lividans TipAL protein and its role in regulating gene expression. J. Biol. Chem. 1999, 274, 20578–20586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegdem, N.S.; Sanders, D.A.; Rodriguez, R.; Balasubramanian, S. The transcription factor FOXM1 is a cellular target of the natural product thiostrepton. Nat. Chem. 2011, 3, 725–731. [Google Scholar] [CrossRef] [PubMed]
- Viehrig, K.; Surup, F.; Harmrolfs, K.; Jansen, R.; Kunze, B.; Müller, R. Concerted action of P450 plus helper protein to form the amino-hydroxy-piperidone moiety of the potent protease inhibitor Crocapeptin. J. Am. Chem. Soc. 2013, 135, 16885–16894. [Google Scholar] [CrossRef] [PubMed]
- Sandargo, B.; Michehl, M.; Praditya, D.; Steinmann, E.; Stadler, M.; Surup, F. Antiviral meroterpenoid rhodatin and sesquiterpenoids rhodocoranes A–E from the wrinkled peach mushroom, Rhodotus palmatus. Org. Lett. 2019, 21, 3286–3289. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Unit | Pos | δC | δH | Unit | Pos | δC | δH |
|---|---|---|---|---|---|---|---|
| Thz 1 | 1 | 163.6, C | Dala 1 | 28 | 133.7, C | ||
| 2 | 127.1, CH | 8.47, s | 28NH | NH | 9.70, br s | ||
| 3 | 149.3, C | 29 | 162.5, C | ||||
| 4 | 159.9, C | 30 | 104.5, CH2 | 5.74, br s | |||
| Val | 5 | 58.2, CH | 4.38, dd (9.3, 7.4) | 30 | 6.49, br s | ||
| 5NH | NH | 8.08, d (9.3) | Oxa | 31 | 129.0, C | ||
| 6 | 170.9, C | 31NH | NH | 9.78, br s | |||
| 5 | 30.6, CH | 2.11, m | 32 | 155.8, C | |||
| 8 | 19.3, CH3 | 0.91, d (6.7) | 33 | 150.0, C | |||
| 9 | 18.4, CH3 | 0.88, d (6.7) | 34 | 133.1, C | |||
| Thz 2 | 10 | 40.2, CH2 | 4.55, m | 35 | 109.5, CH2 | 5.64, br s | |
| 4.70, dd (16.0,6.4) | 5.43, br s | ||||||
| 10NH | NH | 8.95, t (6.4) | 36 | 11.4, CH3 | 2.60, s | ||
| 11 | 168.6, C | Pyr | 37 | 147.6, C | |||
| 12 | 125.3, CH | 8.25, s | 38 | 130.8, C | |||
| 13 | 148.6, C | 39 | 141.1, CH | 8.60, d (8.1) | |||
| 14 | 159.9, C | 40 | 121.1, CH | 8.25, d (8.1) | |||
| Thz 3 | 15 | 46.8, CH | 5.44, m | 41 | 149.0, C | ||
| 15NH | NH | 8.74, d (8.2) | 42 | 161.4, C | |||
| 16 | 173.5, C | Dala 2 | 43 | 134.0, C | |||
| 17 | 125.1, CH | 8.29, m | 43NH | NH | 10.5, br s | ||
| 18 | 148.7, C | 44 | 161.9, C | ||||
| 19 | 159.2, C | 45 | 104.0, CH2 | 5.82, br s | |||
| 20 | 20.5, CH3 | 1.55, d (6.9) | 6.61, br s | ||||
| Thz 4 | 21 | 129.2, C | Dala 3 | 46 | 135.3, C | ||
| 21NH | NH | 9.80, br s | 46NH | NH | 9.54, br s | ||
| 22 | 167.3, C | 47 | 165.1, C | ||||
| 23 | 125.0, CH | 8.31, s | 47NH | NH2 | 7.48, br s | ||
| 24 | 148.5, C | NH2 | 7.91, br s | ||||
| 25 | 158.8, C | 48 | 106.7, CH2 | 5.96, s | |||
| 26 | 128.3, CH | 6.52, d (6.9) | 5.67, s | ||||
| 27 | 14.1, CH3 | 1.79, d (6.9) |
| Compound | L929 | KB3.1 | MCF-7 | SKOV-3 | A431 | PC-3 |
|---|---|---|---|---|---|---|
| 1 | 2.9 | 2.6 | 1.0 | 28 | 0.8 | 31 |
| 2 | 24.0 | / | n.t. 1 | n.t. | n.t | n.t |
| epothilon B [8] | 0.00082 | 0.000065 | 0.000048 | 0.000095 | 0.000045 | 0.0001 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khodamoradi, S.; Stadler, M.; Wink, J.; Surup, F. Litoralimycins A and B, New Cytotoxic Thiopeptides from Streptomonospora sp. M2. Mar. Drugs 2020, 18, 280. https://doi.org/10.3390/md18060280
Khodamoradi S, Stadler M, Wink J, Surup F. Litoralimycins A and B, New Cytotoxic Thiopeptides from Streptomonospora sp. M2. Marine Drugs. 2020; 18(6):280. https://doi.org/10.3390/md18060280
Chicago/Turabian StyleKhodamoradi, Shadi, Marc Stadler, Joachim Wink, and Frank Surup. 2020. "Litoralimycins A and B, New Cytotoxic Thiopeptides from Streptomonospora sp. M2" Marine Drugs 18, no. 6: 280. https://doi.org/10.3390/md18060280
APA StyleKhodamoradi, S., Stadler, M., Wink, J., & Surup, F. (2020). Litoralimycins A and B, New Cytotoxic Thiopeptides from Streptomonospora sp. M2. Marine Drugs, 18(6), 280. https://doi.org/10.3390/md18060280