Neosaxitoxin Inhibits the Expression of Inflammation Markers of the M1 Phenotype in Macrophages

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. NeoSTX Does not Modify Cell Viability in Macrophages
2.2. Nav Characterization in Macrophages
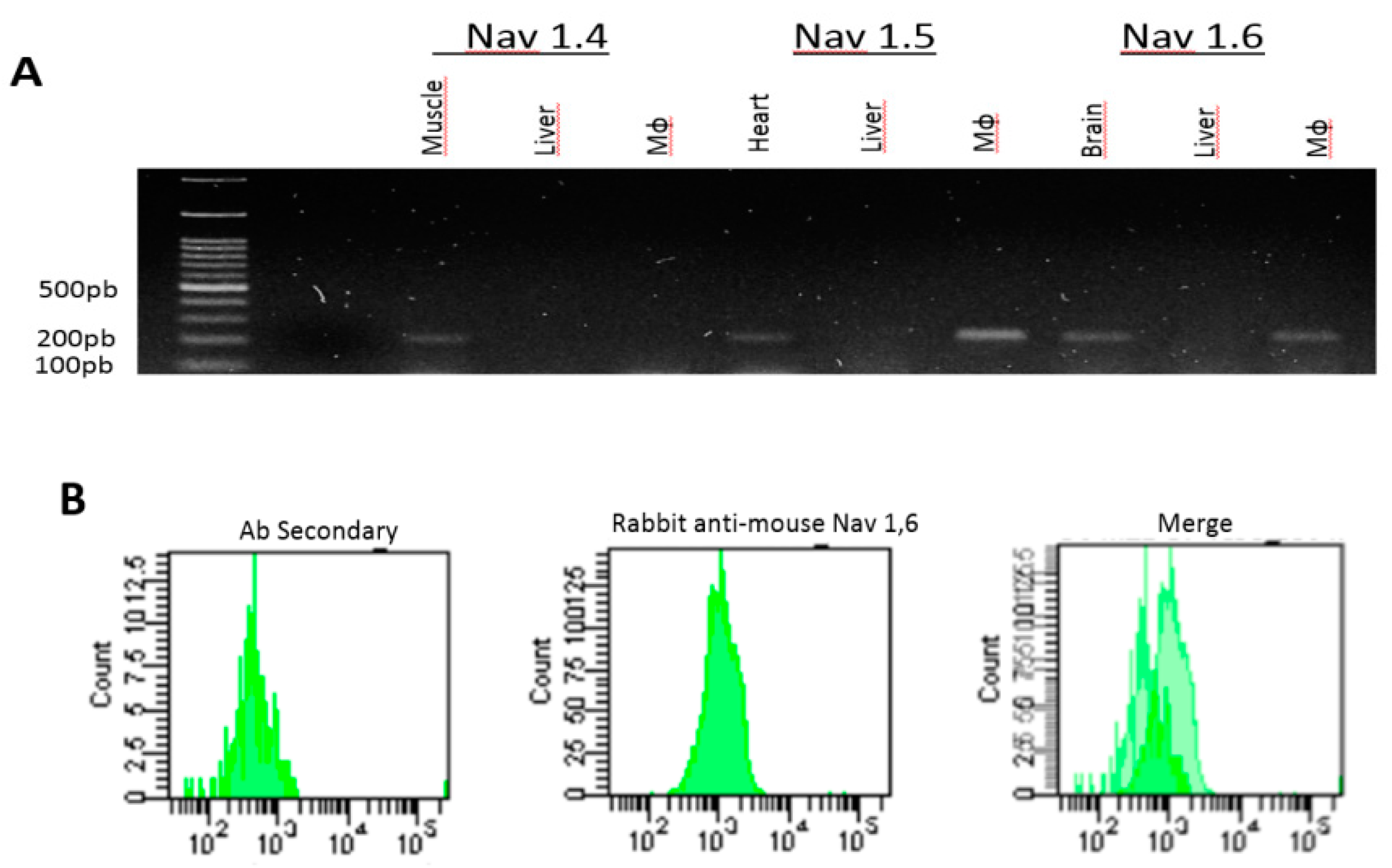
2.2.1. Equine PBMC Cells Express the Nav 1.5 and Nav 1.6 Isoforms
2.2.2. RAW 264.7 Cells Express the Isoform of the Nav 1.6 Channel
2.2.3. LPS Induces the Expression of Nav 1.6 but not of Nav 1.5
2.3. Macrophages Incorporate NeoSTX
2.4. NeoSTX Inhibits Polarization Towards the M1 Macrophage Phenotype
2.4.1. Effect of NeoSTX and Lidocaine on RAW 264.7 Cells
2.4.2. Effect of NeoSTX and Lidocaine in the Primary Culture
2.4.3. NeoSTX Inhibits Polarization towards the M1 Phenotype of Murine Macrophages
2.4.4. NeoSTX Inhibits Polarization towards the M1 Phenotype in the Primary Culture
2.5. NeoSTX Deactivates the M1 Macrophage Phenotype Generated in Response to a known M1 Macrophage Inducer
3. Discussion
3.1. Conclusions
3.2. Possible Application
4. Materials and Methods
4.1. Reagents and Chemicals
4.2. Obtaining Ex-Vivo Macrophages from Equines
4.2.1. Inclusion Criteria
4.2.2. Blood Samples
4.3. Cell Cultures
4.4. Nav Analysis
4.4.1. Primary Tissue Sample
4.4.2. RNA Extraction and Obtaining cDNA for RAW Cells and Equine Cells
4.4.3. RT-PCR
4.5. Cell Viability Tests
4.6. Nitric Oxide Test (NO)
4.7. ELISA Test
4.8. Development of Rabbit Polyclonal Antibodies against NeoSTX
4.9. Flow Cytometry
4.10. Confocal Microscopy
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Gordon, S.; Taylor, P.R. Monocyte and Macrophage Heterogeneity. Nat. Rev. Immunol. 2005, 5, 953–964. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, T.; Fong, C. The resolution of inflammation: Anti- inflammatory roles for NF-kB. Int. J. Biochem. Cell Biol. 2010, 42, 519–523. [Google Scholar] [CrossRef] [PubMed]
- Yona, S.; Jung, S. Monocytes: Subsets, origins, fate and functions. Curr. Opin. Hematol. 2010, 17, 53–59. [Google Scholar] [CrossRef] [PubMed]
- Strauss-Ayali, D.; Conrad, S.M.; Mosser, D. Monocyte subpopulations and their differentiation patterns during infection. J. Leukoc. Biol. 2007, 82, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Dudley, C. Anatomy of a discovery: M1 and M2 macrophages. Front. Immunol. 2015, 6, 1–12. [Google Scholar]
- Mantovani, A.; Locati, M. Orchestation of macrophage polarization. Blood 2009, 15, 3135–3136. [Google Scholar] [CrossRef] [PubMed]
- Biswas, S.; Mantovani, A. Orchestration of Metabolism by Macrophages. Cell Metab. 2012, 15, 432–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palomo, I.; Pereira, J.; Koening, C. Células y Órganos del Sistema Inmune. In Fundamentos de Inmunología Básica y Clínica; e-book Editorial Universidad de Talca: Talca, Chile, 2009; pp. 65–86. [Google Scholar]
- Duque, M.A.; Rojas, M. Revisión: Activación alternativa del macrófago: La diversidad en las respuestas de una célula de la inmunidad innata ante la complejidad de los eventos de su ambiente. Inmunología 2007, 26, 73–86. [Google Scholar] [CrossRef]
- Sica, A.; Mantovani, A. Macrophage plasticity and polarization: In vivo veritas. J. Clin. Investig. 2012, 3, 787–795. [Google Scholar] [CrossRef]
- Jimenez-Garcia, L.; Herranz, S.; Luque, A.; Hortelano, S. Critical role of p38 MAPK in IL-4-induced alternative activation of peritoneal macrophages. Eur. J. Immunol. 2015, 45, 273–286. [Google Scholar] [CrossRef]
- Zhao, X.N.; Li, Y.N.; Wang, Y.T. Interleukin-4 regulates macrophage polarization via the MAPK signaling pathway to protect against atherosclerosis 1. Genet. Mol. Res. 2016, 15, 15. [Google Scholar] [CrossRef] [PubMed]
- Saraiva, M.; O’Garra, A. The regulation of IL-10 production by immune cells. Nat. Rev. Immunol. 2010, 10, 170–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chao-Ying, L.; Juan-Ying, X.; Xiao-Yan, S.; Wei, H.; Ting-Yan, R.; Ping, X.; Jun-Li, D. M2-polarized tumor-associated macrophages promoted epithelial–mesenchymal transition in pancreatic cancer cells, partially through TLR4/IL-10 signaling pathway. Lab. Investig. 2013, 93, 844–854. [Google Scholar]
- Goldsmith, M.; Avni, D.; Levy-Rimler, G.; Mashiach, R.; Ernst, O.; Levi, M.; Webb, B.; Meijler, M.; Gray, N.; Rosen, H.; et al. Aceramide-1-phosphate analogue, PCERA-1, simultaneouly suppresses tumor necrosis factor-α and induces interleuki-10 production in activated macrophages. Immunology 2008, 127, 103–115. [Google Scholar] [CrossRef] [PubMed]
- Avni, D.; Philosoph, A.; Meijler, M.; Zor, T. The ceramide-1-phpsphate analogue PCERA-1 modulates tumor necrosis factor-α and interleukin-10 production in macrophages via the cAMP-PKA-CREB pathway in a GTP-dependent manner. Immunology 2009, 129, 375–385. [Google Scholar] [CrossRef] [PubMed]
- Schuelert, N.; McDougall, J. Involvement of Nav 1.8 sodium ion channels in the transduction of mechanical pain in a rodent model of osteoarthritis. Arthritis Res. Ther. 2012, 14, R5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luiz, A.P.; Wood, J.N. Sodium Channels in Pain and Cancer: New Therapeutic Opportunities. Adv. Pharmacol. 2016, 75, 153–178. [Google Scholar]
- Dib-Hajj, S.; Black, J.; Waxman, S. Voltage-Gated Sodium Channels: Therapeutic Targets for Pain. Pain Med. 2009, 10, 1260–1269. [Google Scholar] [CrossRef] [Green Version]
- Black, J.A.; Waxman, S.G. Noncanonicals rol of Voltage-Gate Sodium Channel. Neuronlogy 2013, 19, 532–542. [Google Scholar]
- Lee, P.; Tsai, P.; Huang, Y.; Huang, C. Inhibition of Toll-like Receptor-4, Nuclear Factor-kB and mitogen-activated protein kinase by Lignocaine may involve voltage-sensitive sodium channels. Clin. Exp. Pharmacol. Physiol. 2008, 35, 1052–1058. [Google Scholar] [CrossRef]
- Catterall, W.; Goldin, A.; Waxman, S. International Union of Pharmacology. XLVII. Nomenclature and Structure-Function Relationship of Voltage-Gated Sodium Channels. Pharmacol. Rev. 2005, 57, 397–409. [Google Scholar] [CrossRef] [PubMed]
- Scheuer, T. Regulation of Sodium Channel Activity by Phosphorylation. Semin. Cell Dev. Biol. 2011, 22, 160–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roselli, F.; Livrea, P.; Jirillo, E. Voltage-Gated Sodium Channel Blockers as Immunomodulators. Drug Discov. 2006, 1, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Carrithers, L.; Hulseberg, P.; Sandor, M.; Carrithers, M. Mycobacteria processing through organelle polarization and localized calcium oscillations. FEMS Immunol. Med. Microbiol. 2011, 63, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Feske, S.; Wulff, H.; Skolnik, E. Ion Channels in Innate and Adaptive Immunity. Annu. Rev. Immunol. 2015, 33, 291–353. [Google Scholar] [CrossRef] [Green Version]
- Chatelier, A.; Mercier, A.; Tremblier, B.; Thériault, O.; Moubarak, M.; Benamer, N.; Corbi, P.; Bois, P.; Chahine, M.; Faivre, J.F. A distinct de novo expression of Nav1.5 sodium channels in human atrial fibroblasts differentiated into myofibroblasts. J. Physiol. 2012, 590, 4307–4319. [Google Scholar] [CrossRef]
- Rathmell, J.P.; Strichartz, G.; Wanderer, J. Neosaxitoxin versus Traditional Local Anesthetics: Mechanism of Action and Sites of Notable Effect. Anesthesiology 2015, 123. [Google Scholar] [CrossRef]
- Lagos, N. Microalgal bloom: A global issue with negative impact in Chile. Biol. Res. 1998, 31, 375–386. [Google Scholar]
- Lagos, N. Clinical Applications of Paralytic Shellfish Poisoning Toxins. In Toxins and Biologically Active Compounds from Microalgae; CRC Press, Taylor & Francis Group: New York, NY, USA, 2014; Volume 2, pp. 309–329. [Google Scholar]
- Lobo, K.; Donado, C.; Cornelissen, L.; Kim, J.; Ortiz, R.; Peake, R.W.; Kellogg, M.; Alexander, M.E.; Zurakowski, D.; Kurgansky, K.E.; et al. A phase 1, dose-escalation, double-blind, block-randomized, controlled trial of safety and efficacy of neosaxitoxin alone and in combination with 0.2% bupivacaine, with and without epinephrine, for cutaneous anesthesia. Anesthesiology 2015, 123, 873–885. [Google Scholar] [CrossRef] [Green Version]
- Garrido, R.; Lagos, N.; Lattes, K.; Abedrapo, M.; Bocic, G.; Cuneo, A.; Chiong, H.; Jensen, C.; Azolas, R.; Henriquez, A.; et al. Gonyautoxin: New treatment for healing acute and chronic anal fissures. Dis. Colon Rectum 2005, 48, 335–340. [Google Scholar] [CrossRef]
- Garrido, R.; Lagos, N.; Lattes, K.; Azolas, C.G.; Bocic, G.; Cuneo, A.; Chiong, H.; Jensen, C.; Henríquez, A.; Fernández, C. The gonyautoxin 2/3 epimers reduces anal tone when injected in the anal sphincter of healthy adults. Biol Res. 2004, 37, 395–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lattes, K.; Venegas, P.; Lagos, N.; Lagos, M.; Pedraza, L.; Rodriguez-Navarro, A.J.; García, C. Local infiltration of gonyautoxin is safe and effective in treatment of chronic tensiontype headache. Neurol. Res. 2009, 31, 208–233. [Google Scholar] [CrossRef] [PubMed]
- Hinzpeter, J.; Barrientos, C.; Zamorano, A.; Martinez, A.; Palet, M.; Wulf, R.; Barahona, M.; Sepúlveda, J.M.; Guerra, M.; Bustamante, T.; et al. Gonyautoxins: First evidence in pain management in total knee arthroplasty. Toxicon 2016, 119, 180–185. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Navarro, A.J.; Lagos, M.; Figueroa, C.; Garcia, C.; Recabal, P.; Silva, P.; Iglesias, V.; Lagos, N. Potentiation of local anesthetic activity of neosaxitoxin with bupivacaine or epinephrine: Development of a long-acting pain blocker. Neurotox. Res. 2009, 4, 408–415. [Google Scholar] [CrossRef]
- Rodriguez-Navarro, A.J.; Lagos, N.; Lagos, M.; Braghetto, I.; Csendes, A.; Hamilton, J.; Berger, Z.; Wiedmaier, G.; Henriquez, A. Intrasphincteric neosaxitoxin injection: Evidence of lower esophageal sphincter relaxation in achalasia. Am. J. Gastroenterol. 2006, 11, 2667–2668. [Google Scholar] [CrossRef]
- Riquelme, G.; Sepúlveda, J.; Ghumgham, A.; Del Campo, M.; Montero, M.C.; Lagos, N. Neosaxitoxin, a Paralytic Shellfish Poison Toxin, effectively manages bucked shins pain, as a local long-action pain blocker in an equine model. Toxicon 2018, 141, 15–17. [Google Scholar] [CrossRef]
- Rampersad, S.N. Review Multiple Applications of Alamar Blue as an Indicator of Metabolic Function and Cellular Health in Cell Viability Bioassays. Sensors 2012, 12, 12347–12360. [Google Scholar] [CrossRef]
- Yuan, T.; Li, Z.; Li, X.; Yu, G.; Wang, N.; Yang, X. Lidocaine attenuates lipopolysaccharide-induced inflammatory responses in microglia. J. Surg. Res. 2014, 1, 150–162. [Google Scholar] [CrossRef]
- Huang, Y.; Tsai, P.; Kai, Y.; Yang, C.; Huang, C. Lidocaine Inhibition of Inducible Nitric Oxide Synthase and Cationic Amino Acid Transporter-2 Transcription in Activated Murine Macrophages May Involve Voltage-Sensitive Nav Channel. Anesth. Analg. 2006, 102, 1739–1744. [Google Scholar] [CrossRef]
- Carrithers, M.; Chatterjee, G.; Carrithers, L.; Tokmoulina, O.R.; Iheagwara, U.; Rahner, C.; Graham, M.; Waxman, S. Regulation of podosome Formation in Macrophages by a Splice Variant of the Sodium Channel SCN8A. J. Biol. Chem. 2009, 12, 8114–8126. [Google Scholar] [CrossRef] [Green Version]
- Carrithers, M.; Dib-Hajj, S.; Carrithers, L.; Tokmoulina, G.; Pypaert, M.; Jonas, E.; Waxman, E. Expression of the Voltage-Gated Sodium Channel NaV1.5 in the Macrophage Late Endosome Regulates Endosomal Acidification. J. Immunol. 2007, 178, 7822–7832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hossain, M.; Sonsalla, P.; Richardson, J. Coordinate rol of voltage-gate sodium channels and the Na+/H+ exchanger in sustaining microglial activation during inflammation. Toxicol. Appl. Pharmacol. 2013, 273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Craner, M.; Damarjian, T.; Liu, S.; Hains, B.; Lo, A.; Black, J.; Newcombe, J.; Cuzner, M.; Waxman, S. Sodium Channels Contribute to Microglia/Macrophage Activation and Function in EAE and MS. Glia 2005, 49, 220–229. [Google Scholar] [CrossRef] [PubMed]
- Holevinsky, K.; Nelson, D. Simultaneous Detection of Free Radical Release and Membrane Current during Phagocytosis. J. Biol. Chem. 1995, 14, 8328–8336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Black, J.; Liu, S.; Waxman, S. Sodium Channel Activity Modulates Multiple Functions in Microglia. Glia 2009, 57, 1072–1081. [Google Scholar] [CrossRef] [PubMed]
- Cestèle, S.; Catterall, W. Molecular mechanisms of neurotoxin action on voltage-gated sodium channels. Biochimie 2000, 82, 883–892. [Google Scholar] [CrossRef]
- Perez-Reyes, E. Molecular Physiology of Low-Voltage-Activated T-type Calcium Channels. Physiol. Rev. 2003, 83, 117–161. [Google Scholar] [CrossRef] [Green Version]
- Catterall, W.; Swanson, T. Structural Basis for Pharmacology of Voltage-Gated Sodium and Calcium Channels. Mol. Pharmacol. 2015, 88, 141–150. [Google Scholar] [CrossRef]
- Fozzard, H.; Lipkind, G. The Tetrodotoxin Binding Site Is within the Outer Vestibule of the Sodium Channel. Mar. Drugs 2010, 8, 219–234. [Google Scholar] [CrossRef] [Green Version]
- Tak, P.; Firestein, G. NF-kB: A key role in inflammatory diseases. J. Clin. Investig. 2001, 1, 7–11. [Google Scholar] [CrossRef]
- Venable, A.S.; Henning, A.L.; Prado, E.A.; McFarlin, B.K. Using Image-Based Flow Cytometry with a FISH-Based Flow RNA Assay to Simultaneously Detect Intracellular TNF-α Protein and mRNA in Monocytes Following LPS Stimulation. In Book Imaging Flow Cytometry: Methods and Protocols, Methods in Molecular Biology; Barteneva, N.S., Vorobjev, I.A., Eds.; Springer Science + Business Media: New York, NY, USA, 2016; pp. 139–151. [Google Scholar]
- Jung, G.; Lee, J.; Rhim, H.; Oh, T.; Yune, T. An increase in Volta-Gate Sodium Channel Current Elicits Microglial Activation Followed Inflammatory Responses In Vitro and In vivo After Spinal Cord Injury. Glia 2013, 61, 1807–1821. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Kintner, D.; Chanana, V.; Algharabli, J.; Chen, X.; Gao, Y.; Chen, J.; Ferrezzano, P.; Olson, J.; Sun, D. Activation of microglia depends on Na+/H+ exchange-mediated H+ homeostasis. J. Neurosci. 2010, 45, 15210–15220. [Google Scholar] [CrossRef] [PubMed]
- Ramírez-Bello, V.; Sevcik, C.; Peigneur, S.; Tytgat, J. Macrophage alteration induced by inflamatory toxins isolated from Tityusdiscrepans scorpion venom. The rol of Na+/Ca2+ exchangers. Toxicon 2014, 82, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Staiano, R.; Granata, F.; Secondo, A.; Petraroli, A.; Loffredo, S.; Annunziato, L.; Triggiani, M.; Marone, G. Human Macrophages and Monocytes Express Functional Na+/Ca2+ Exchangers 1 and 3. Eur. J. Immunol. 2009, 39, 1405–1418. [Google Scholar] [CrossRef] [PubMed]
- Jackson, K.; Stottb, J.; Horohovc, D.; Watson, J. IL-4 induced CD23 (FceRII) up-regulation in equine peripheral blood mononuclear cells and pulmonary alveolar macrophages. Vet. Immunol. Immunopathol. 2004, 101, 243–250. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.; Livak, K. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 6, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xing, R.; Liu, S.; Qin, Y.; Li, K.; Yu, H.; Li, P. Hydroxypropyltrimethyl ammonium chloride chitosan activates RAW 264.7 macrophages through the MAPK and JAK-STAT signaling pathways. Carbohydr. Polym. 2018, 205, 401–409. [Google Scholar] [CrossRef] [PubMed]
- Torres, E.; Vaquero, C.; Nicholson, L.; Sack, M.; Stöger, E.; Drossard, J.; Christou, P.; Fischer, R.; Perrin, Y. Rice cell culture as an alternative production system for functional diagnostic and therapeutic antibodies. Transgenic Res. 1999, 8, 441–449. [Google Scholar] [CrossRef] [PubMed]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montero, M.C.; del Campo, M.; Bono, M.; Simon, M.V.; Guerrero, J.; Lagos, N. Neosaxitoxin Inhibits the Expression of Inflammation Markers of the M1 Phenotype in Macrophages. Mar. Drugs 2020, 18, 283. https://doi.org/10.3390/md18060283
Montero MC, del Campo M, Bono M, Simon MV, Guerrero J, Lagos N. Neosaxitoxin Inhibits the Expression of Inflammation Markers of the M1 Phenotype in Macrophages. Marine Drugs. 2020; 18(6):283. https://doi.org/10.3390/md18060283
Chicago/Turabian StyleMontero, M. Cecilia, Miguel del Campo, M. Bono, M. Valeska Simon, Julia Guerrero, and Néstor Lagos. 2020. "Neosaxitoxin Inhibits the Expression of Inflammation Markers of the M1 Phenotype in Macrophages" Marine Drugs 18, no. 6: 283. https://doi.org/10.3390/md18060283
APA StyleMontero, M. C., del Campo, M., Bono, M., Simon, M. V., Guerrero, J., & Lagos, N. (2020). Neosaxitoxin Inhibits the Expression of Inflammation Markers of the M1 Phenotype in Macrophages. Marine Drugs, 18(6), 283. https://doi.org/10.3390/md18060283