Exploring the Microalga Euglena cantabrica by Pressurized Liquid Extraction to Obtain Bioactive Compounds

 ,
,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
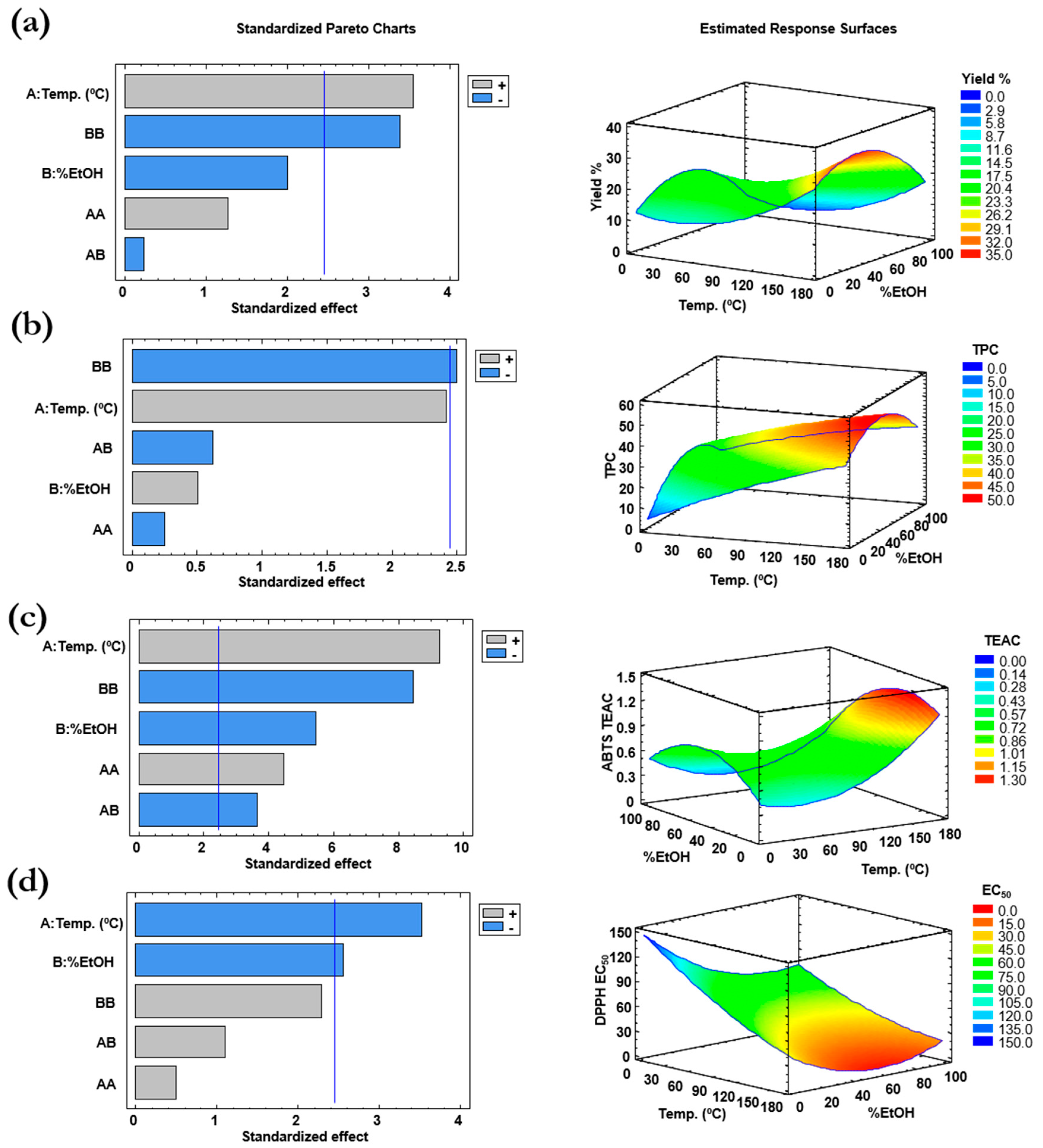
2.1. Design of Experiments (DoE) for Pressurized Liquid Extraction (PLE)
2.2. Chemical Characterization
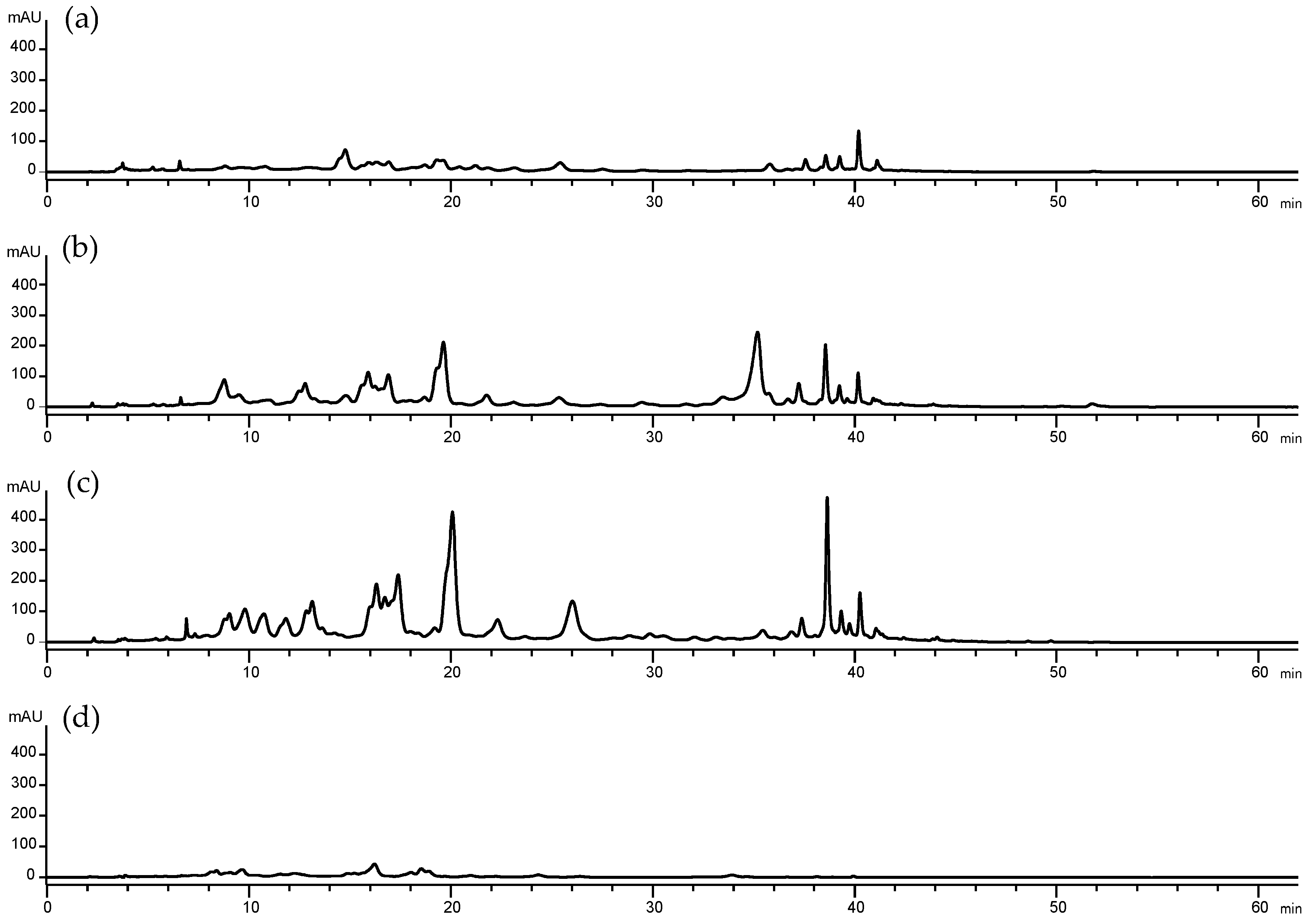
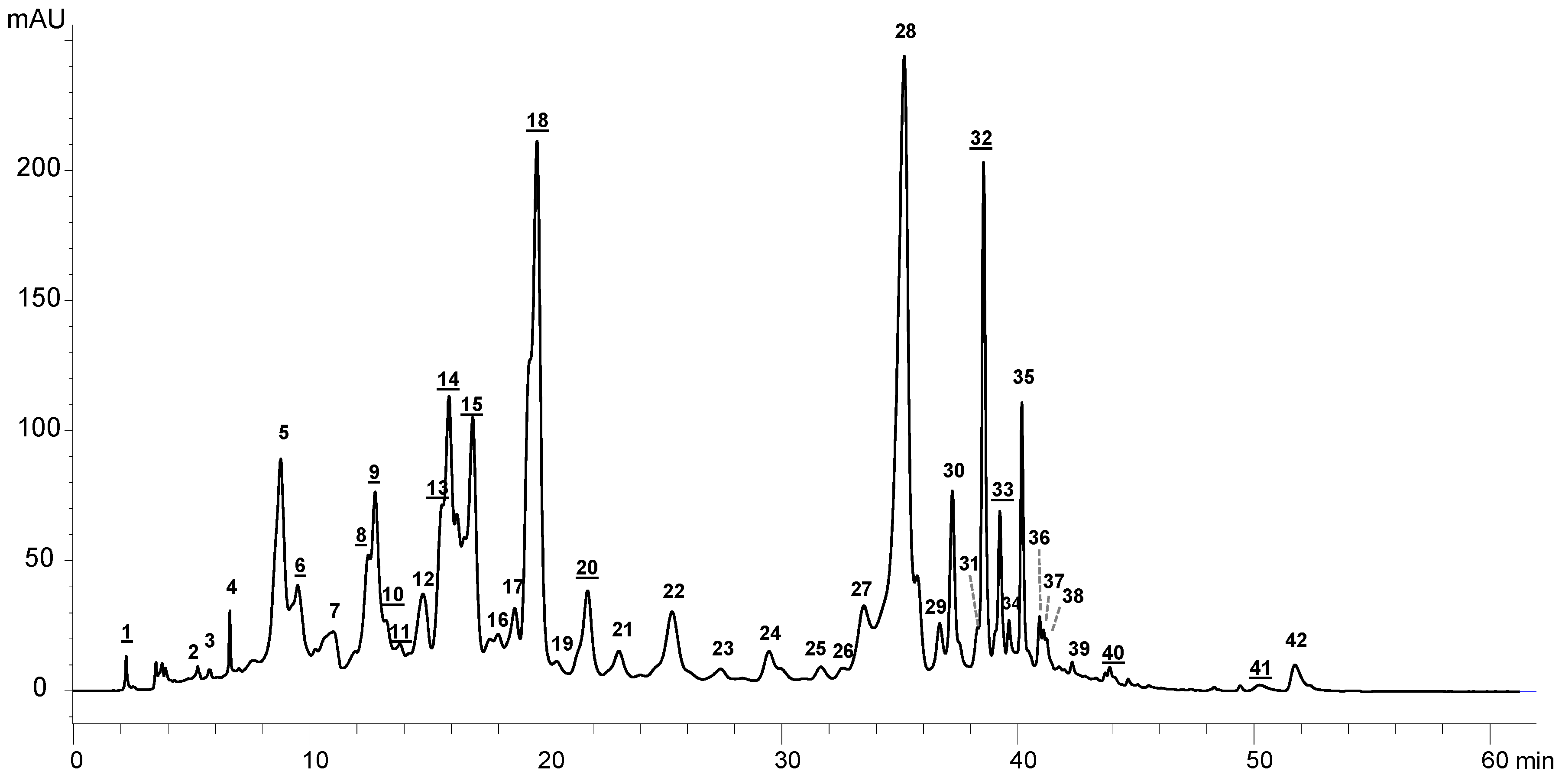
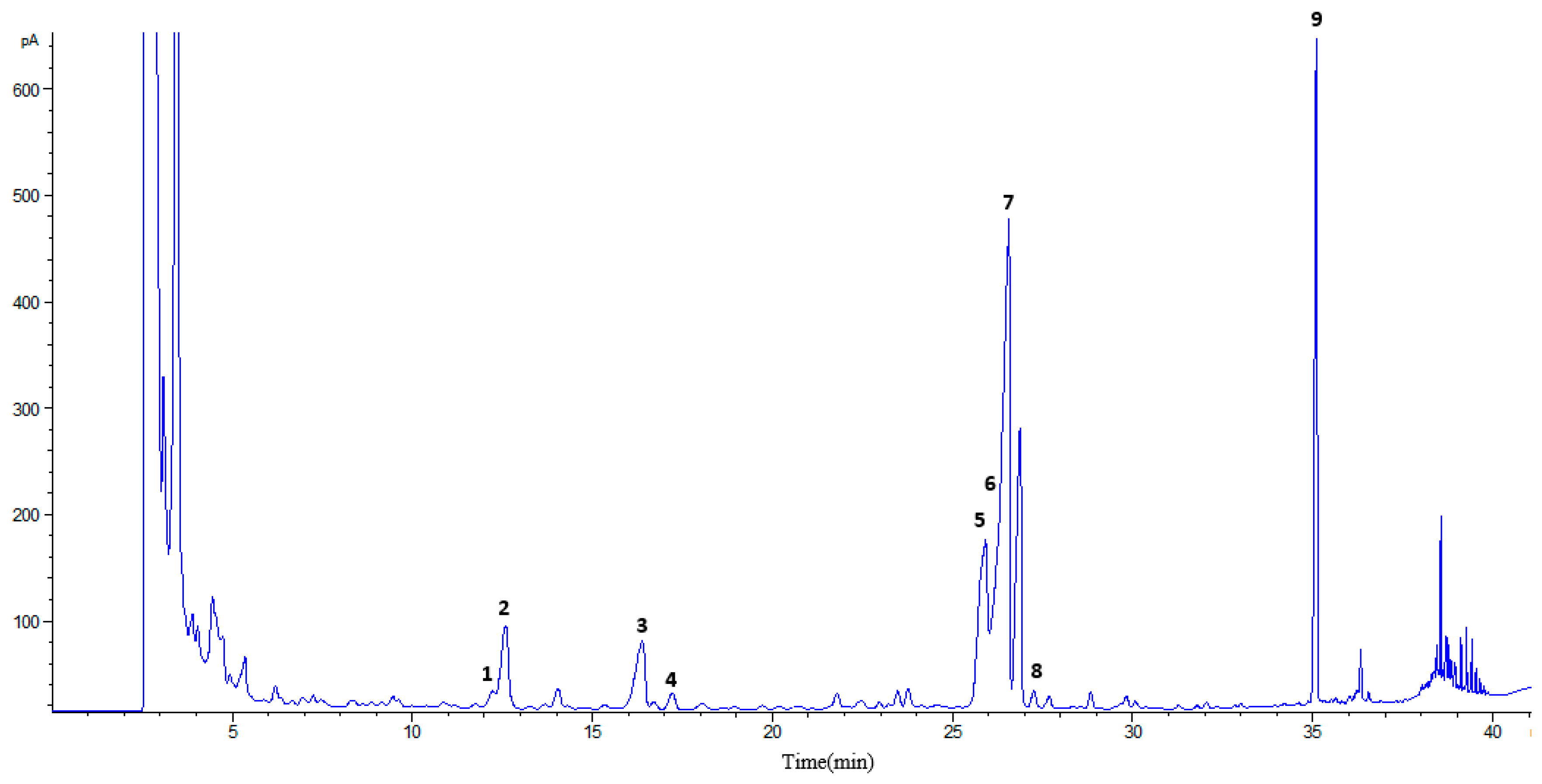
2.2.1. Analysis of Pigments by Liquid Chromatography-diode Array Detection Followed by Atmospheric Pressure Chemical Ionization Tandem Mass Spectrometry (HPLC-DAD-APCI-MS/MS)
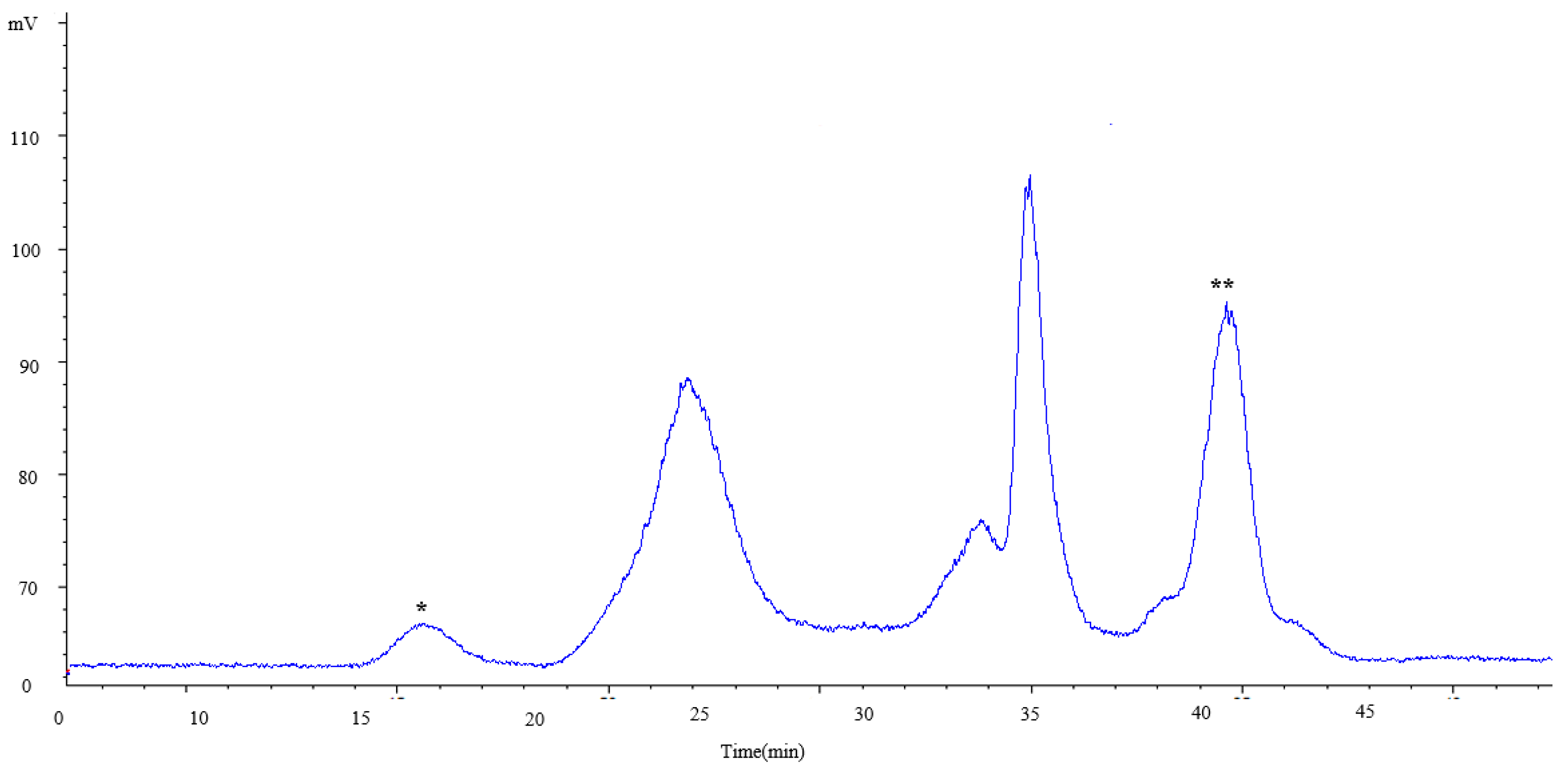
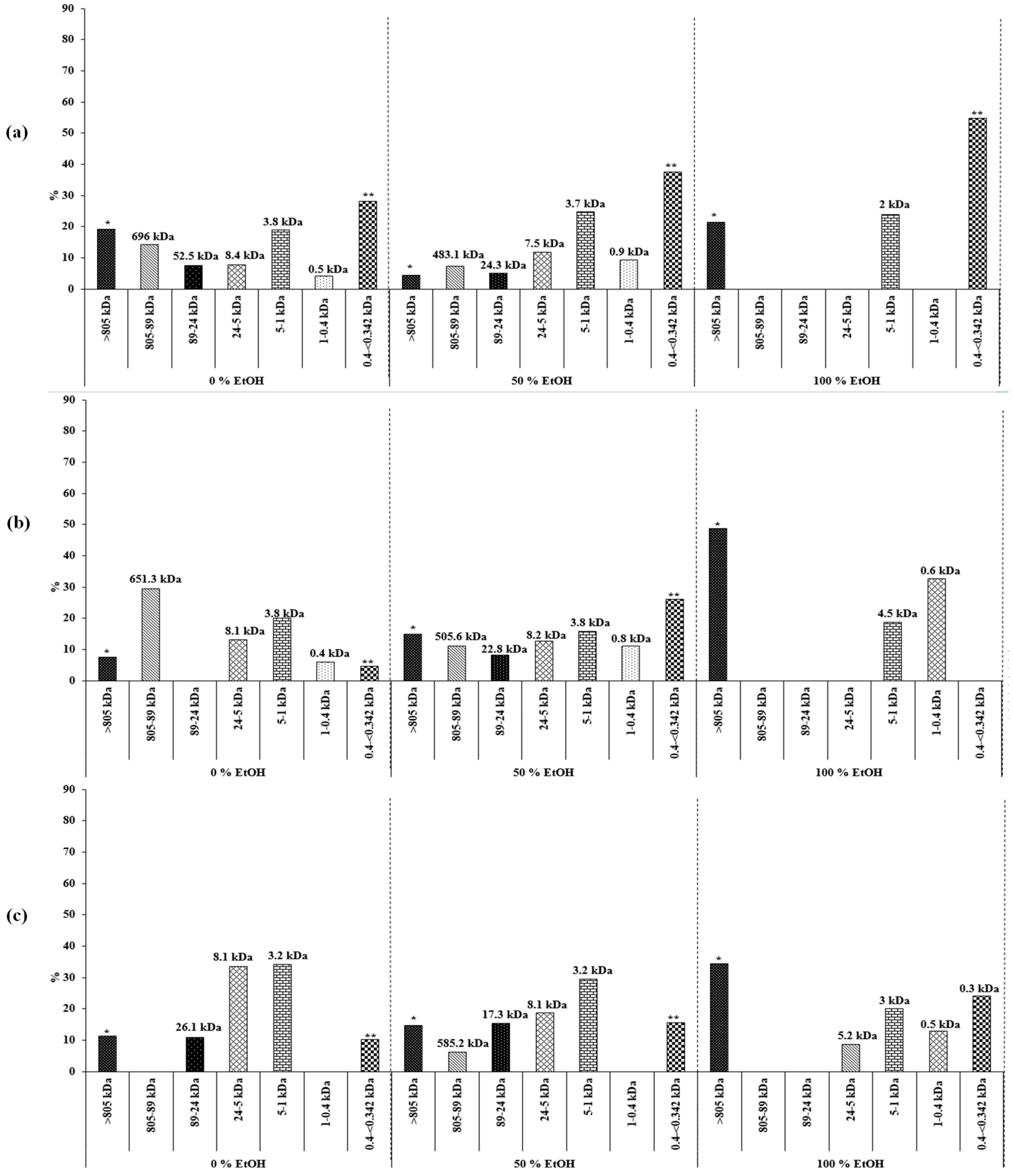
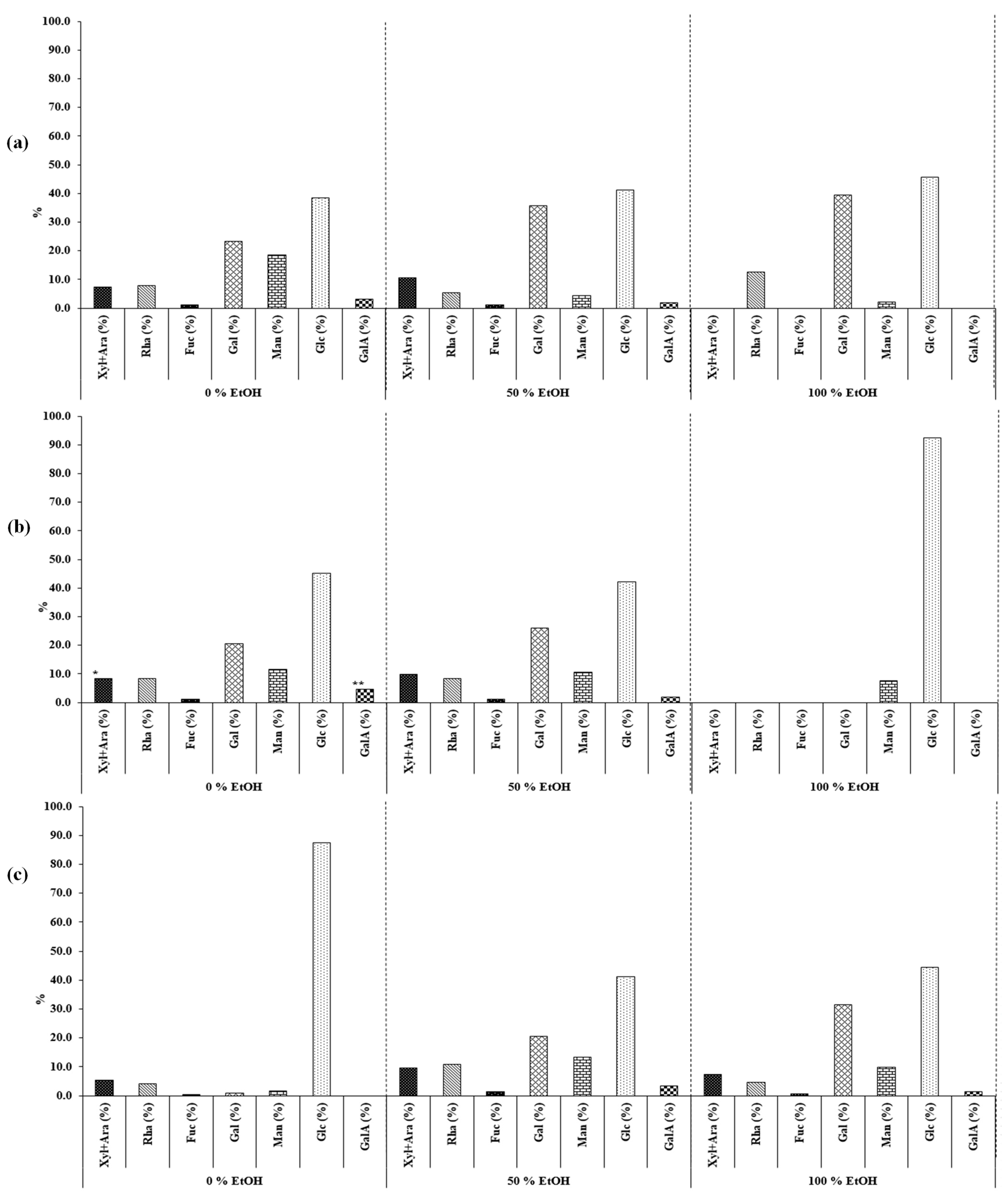
2.2.2. Characterization of Carbohydrates
3. Materials and Methods
3.1. Strain, Culture Conditions, and Breaking of the Cell Wall
3.2. Pressurized Liquid Extraction
3.3. Chemical Characterization and Evaluation of the Antioxidant Capacity
3.3.1. Total Phenols Content (TPC): Folin-Ciocalteau Test
3.3.2. Antioxidant Capacity: ABTS Test
3.3.3. Total Lipids
3.3.4. Carotenoids and Chlorophylls
3.3.5. Carbohydrates
Estimation of Molecular Weight (Mw)
Determination of Monomeric Composition
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Allen, E.J.; Allen, E.J.; Nelson, E.W. On the artificial culture of marine plankton organisms. J. Mar. Biol. Assoc. U.K. 1910, 8, 421–474. [Google Scholar] [CrossRef] [Green Version]
- Herrero, M.; Sánchez-Camargo, A.P.; Cifuentes, A.; Ibáñez, E. Plants, seaweeds, microalgae and food by-products as natural sources of functional ingredients obtained using pressurized liquid extraction and supercritical fluid extraction. TRAC-Trends Anal. Chem. 2015, 71, 26–38. [Google Scholar] [CrossRef]
- Mobin, S.; Alam, F. Some Promising Microalgal Species for Commercial Applications: A review. Energy Procedia 2017, 110, 510–517. [Google Scholar] [CrossRef]
- Watanabe, T.; Shimada, R.; Matsuyama, A.; Yoshida, E.; Suzuki, K. Antitumor activity of the β-glucan paramylon from Euglena against preneoplastic colonic aberrant crypt foci in mice. Food Funct. 2013, 4, 1685–1690. [Google Scholar] [CrossRef] [PubMed]
- Kottuparambil, S.; Thankamony, R.L.; Agusti, S. Euglena as a potential natural source of value-added metabolites. A review. Algal Res. 2019, 37, 154–159. [Google Scholar] [CrossRef]
- Shin, W.; Triemer, R.E. Phylogenetic analysis of the genus Euglena (Euglenophyceae) with particular reference to the type species Euglena viridis. J. Phycol. 2004, 40, 759–771. [Google Scholar] [CrossRef]
- Kosmala, S.; Karnkowska-Ishikawa, A.; Milanowski, R.; Kwiatowski, J.; Zakry’s, B. Phylogeny and systematics of Euglena (Euglenaceae) species with axial, stellate chloroplasts based on morphological and molecular data-new taxa, emended diagnoses, and epitypifications. J. Phycol. 2009, 45, 464–481. [Google Scholar] [CrossRef]
- Kim, J.I.; Linton, E.W.; Shin, W. Taxon-rich multigene phylogeny of the photosynthetic euglenoids (Euglenophyceae). Front. Ecol. Evol. 2015, 3, 98. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, K.; Arashida, R.; Marukawa, Y.; Yoshida, E.; Takeda, T.; Nakano, Y.; Konno, N.; Takahashi, M. B-1,3-glucanase, polynucleotide, recombinant vector, transformant, production method for B-1,3- glucanase, enzyme preparation, and production method for paramylon having reduced molecular weight. U.S. Patent US9644193B2, 9 May 2017. [Google Scholar]
- Borowitzka, M.A. Biology of Microalgae in Health and Disease Prevention; Academic Press: Cambridge, MA, USA, 2018; pp. 23–72. [Google Scholar]
- Deli, J.; Gonda, S.; Nagy, L.Z.; Szabó, I.; Gulyás-Fekete, G.; Agócs, A.; Marton, K.; Vasas, G. Carotenoid composition of three bloom-forming algae species. Food Res. Int. 2014, 65, 215–223. [Google Scholar] [CrossRef]
- Jerez-Martel, I.; García-Poza, S.; Rodríguez-Martel, G.; Rico, M.; Afonso-Olivares, C.; Gómez-Pinchetti, J.L. Phenolic Profile and Antioxidant Activity of Crude Extracts from Microalgae and Cyanobacteria Strains. J. Food Qual. 2017, 2924508. [Google Scholar] [CrossRef] [Green Version]
- Grosso, C.; Valentão, P.; Ferreres, F.; Andrade, P.B. Alternative and efficient extraction methods for marine-derived compounds. Mar. Drugs 2015, 13, 3182–3230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrero, M.; Castro-Puyana, M.; Mendiola, J.A.; Ibáñez, E. Compressed fluids for the extraction of bioactive compounds. TRAC-Trends Anal. Chem. 2013, 43, 67–83. [Google Scholar] [CrossRef]
- Herrero, M.; Jaime, L.; Martín-Álvarez, P.J.; Cifuentes, A.; Ibáñez, E. Optimization of the extraction of antioxidants from Dunaliella salina microalga by pressurized liquids. J. Agric. Food Chem. 2006, 54, 5597–5603. [Google Scholar] [CrossRef] [PubMed]
- Castro-Puyana, M.; Herrero, M.; Urreta, I.; Mendiola, J.A.; Cifuentes, A.; Ibáñez, E.; Suárez-Alvarez, S. Optimization of clean extraction methods to isolate carotenoids from the microalga Neochloris oleoabundans and subsequent chemical characterization using liquid chromatography tandem mass spectrometry. Anal. Bioanal. Chem. 2013, 405, 4607–4616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert-López, B.; Barranco, A.; Herrero, M.; Cifuentes, A.; Ibáñez, E. Development of new green processes for the recovery of bioactives from Phaeodactylum tricornutum. Food Res. Int. 2017, 99, 1056–1065. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zainan, N.H.; Thiruvenkadam, S.; Danquah, M.K.; Harun, R. Biochemical analysis and potential applications of aqueous and solid products generated from subcritical water extraction of microalgae Chlorella pyrenoidosa biomass. J. Appl. Phycol. 2020, 32, 111–126. [Google Scholar] [CrossRef]
- Golmakani, M.T.; Mendiola, J.A.; Rezaei, K.; Ibáñez, E. Pressurized limonene as an alternative bio-solvent for the extraction of lipids from marine microorganisms. J. Supercrit. Fluid. 2014, 92, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Gilbert-López, B.; Mendiola, J.A.; Fontecha, J.; van den Broek, L.A.; Sijtsma, L.; Cifuentes, A.; Ibáñez, E. Downstream processing of Isochrysis galbana: A step towards microalgal biorefinery. Green Chem. 2015, 17, 4599–4609. [Google Scholar] [CrossRef] [Green Version]
- Gilbert-López, B.; Mendiola, J.A.; van den Broek, L.A.M.; Houweling-Tan, B.; Cifuentes, A.; Herrero, M.; Ibáñez, E. Green compressed fluid technologies for downstream processing of Scenedesmus obliquus in a biorefinery approach. Algal Res. 2017, 24, 111–121. [Google Scholar] [CrossRef] [Green Version]
- Britton, G.; Liaaen-Jensen, S. Carotenoids Handbook; Pfander, H., Ed.; Birkhäuser: Basel, Switzerland, 2004. [Google Scholar]
- Huang, S.C.; Hung, C.F.; Wu, W.B.; Chen, B.H. Determination of chlorophylls and their derivatives in Gynostemma pentaphyllum Makino by liquid chromatography–mass spectrometry. J. Pharm. Biomed. Anal. 2008, 48, 105–112. [Google Scholar] [CrossRef]
- Chen, K.; Ríos, J.J.; Roca, M.; Pérez-Gálvez, A. Development of an accurate and high-throughput methodology for structural comprehension of chlorophylls derivatives. (II) Dephytylated derivatives. J. Chromatogr. A 2015, 1412, 90–99. [Google Scholar] [CrossRef] [Green Version]
- Viera, I.; Roca, M.; Perez-Galvez, A. Mass spectrometry of non-allomerized chlorophylls a and b derivatives from plants. Curr. Org. Chem. 2018, 22, 842–876. [Google Scholar] [CrossRef] [Green Version]
- Takaichi, S. Carotenoids in algae: Distributions, biosyntheses and functions. Mar. Drugs 2011, 9, 1101–1118. [Google Scholar] [CrossRef] [PubMed]
- Kato, S.; Soshino, M.; Takaichi, S.; Ishikawa, T.; Nagata, N.; Asahina, M.; Shinomura, T. Suppression of the phytoene synthase gene (EgcrtB) alters carotenoid content and intracellular structure of Euglena gracilis. BMC Plant Biol. 2017, 17, 125. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, A.; Sugimoto, R.; Suzuki, K.; Shirakata, Y.; Hashiguchi, T.; Yoshida, C.; Nakano, Y. Anti-fibrotic activity of Euglena gracilis and paramylon in a mouse model of non-alcoholic steatohepatitis. Food Sci. Nutr. 2018, 7, 139–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gissibl, A.; Care, A.; Parker, L.M.; Iqbal, S.; Hobba, G.; Nevalainen, H.; Sunna, A. Microwave pretreatment of paramylon enhances the enzymatic production of soluble β-1,3-glucans with immunostimulatory activity. Carbohydr. Polym. 2018, 196, 339–347. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, E.C.; Kuhaudomlarp, S.; Rejzek, M.; Fangel, J.U.; Alagesan, K.; Kolarich, D.; Willats, W.G.T.; Field, R.A. Exploring the Glycans of Euglena gracilis. Biology 2017, 6, 45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koşar, M.; Dorman, H.J.D.; Hiltunen, R. Effect of an acid treatment on the phytochemical and antioxidant characteristics of extracts from selected Lamiaceae species. Food Chem. 2005, 91, 525–533. [Google Scholar]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT-Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Nabet, N.; Gilbert-López, B.; Madani, K.; Herrero, M.; Ibáñez, E.; Mendiola, J.A. Optimization of microwave-assisted extraction recovery of bioactive compounds from Origanum glandulosum and Thymus fontanesii. Industrial Crops Products 2020, 129, 395–404. [Google Scholar] [CrossRef]
- Axelsson, M.; Gentili, F. A single-step method for rapid extraction of total lipids from green microalgae. PLoS ONE 2004, 9, e89643. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz-Almagro, N.; Valadez-Carmona, L.; Mendiola, J.A.; Ibáñez, E.; Villamiel, M. Structural characterisation of pectin obtained from cacao pod husk. Comparison of conventional and subcritical water extraction. Carbohydr. Polym. 2019, 217, 69–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment # | T, °C | %EtOH (v/v) | Extraction Yield, %(m/m) | TPC, mg GAE/g Extract | ABTS, mmol TE/g Extract | DPPH, EC50 (µg/mL) |
|---|---|---|---|---|---|---|
| Center (a) | 110 | 50 | 21.6 ± 1.1 | 45.6 ± 6.9 | 0.80 ± 0.02 | 26.8 ± 3.9 |
| 2 | 180 | 50 | 39.1 | 38.3 | 1.26 | 27.6 |
| 3 | 110 | 0 | 9.9 | 16.8 | 0.46 | 66.2 |
| 4 | 40 | 0 | 15.9 | 14.5 | 0.39 | 132.2 |
| 7 | 180 | 100 | 12.1 | 39.4 | 0.55 | 7.5 |
| 9 | 180 | 0 | 27.3 | 46.0 | 1.28 | 29.7 |
| 10 | 110 | 100 | 11.6 | 30.1 | 0.32 | 33.2 |
| 11 | 40 | 100 | 3.2 | 19.5 | 0.22 | 67.8 |
| 12 | 40 | 50 | 12.9 | 34.3 | 0.73 | 30.0 |
| Peak # | Rt (min) | Identification | Absorbance Max (nm) | Parent m/z | Fragments m/z |
|---|---|---|---|---|---|
| 1 | 2.2 | Carotenoid | 272, 424(sh), 448, 474 | ||
| 2 | 5.3 | Chlorophyll | 404, 660 | ||
| 3 | 5.8 | Chlorophyll | 402, 662 | ||
| 4 | 6.6 | Chlorophyll | 408, 660 | ||
| 5 | 8.8 | Chlorophyll | 438, 654 | ||
| 6 | 9.5 | Carotenoid | 402(sh), 422, 446 | 583.6 | 491.6 |
| 7 | 11.0 | Pheophorbide a | 408, 666 | 593.5 | 533.5, 451.5 |
| 8 | 12.5 | Carotenoid | 426(sh), 448 | ||
| 9 | 12.8 | Carotenoid | 426(sh), 448 | ||
| 10 | 13.2 | Carotenoid | 422, 446(sh) | 594.6 | 494.4 |
| 11 | 13.8 | Carotenoid | 450, 472(sh) | ||
| 12 | 14.8 | Pyropheophorbide b | 438, 654 | 549.4 | 521.5 |
| 13 | 15.6 | Carotenoid | 444, 472 | ||
| 14 | 15.9 | Carotenoid | 446, 468 | ||
| 15 | 16.9 | Carotenoid | 408(sh), 430, 456 | 506.9 | 268.4 |
| 16 | 18.0 | Pyropheophorbide a | 412, 666 | 535.5 | |
| 17 | 18.7 | Not identified | |||
| 18 | 19.6 | Diatoxanthin | 450, 478 | 567.5 | 549.7 |
| 19 | 20.5 | Chlorophyll | 418, 660 | 889.7 | 611.3, 551.5 |
| 20 | 21.8 | Carotenoid | 426(sh), 450, 478 | 557.7 | 291.4 |
| 21 | 23.1 | Chlorophyll | 418, 660 | 959.8 | |
| 22 | 25.4 | Hydroxypheophytin b | 438, 472(sh), 654 | 901.6 | 873.8, 623.5 |
| 23 | 27.4 | Chlorophyll | 418, 664 | 887.6 | 869.6, 609.5 |
| 24 | 29.5 | Hydroxypheophytin a | 404, 666 | 887.8 | 869.8, 609.5 |
| 25 | 31.7 | Chlorophyll | 416, 664 | 852.1, 887.8 | |
| 26 | 32.6 | Chlorophyll | 434, 660 | 823.8 | 805.8, 567.5 |
| 27 | 33.5 | Pheophytin b and b’ | 436, 654 | 885.8 | 607.4, 547.3 |
| 28 | 35.2 | 436, 654 | 885.8 | 607.5, 547.4 | |
| 29 | 36.7 | Pheophytin a and a’ | 408, 666 | 871.9 | 593.5, 533.5 |
| 30 | 37.2 | 408, 666 | 871.9 | 593.5, 533.5 | |
| 31 | 38.3 | Chlorophyll | 418, 664 | 871.9 | 856 |
| 32 | 38.6 | Carotenoid | 426(sh), 450, 478 | ||
| 33 | 39.2 | Carotenoid | 420(sh), 444, 472 | 603.6 | 265.3 |
| 34 | 39.6 | Chlorophyll | 418, 664 | 955.9 | 676.6, 616.6 |
| 35 | 40.2 | Pyropheophytin b | 436, 654 | 827.5 | 549.5 |
| 36 | 40.9 | Not identified | 466, 492(sh), 662 | ||
| 37 | 41.1 | Pyropheophytin a and a’ | 410, 668 | 813.9 | 535.5 |
| 38 | 41.2 | 410, 668 | 813.9 | 535.5 | |
| 39 | 42.3 | Chlorophyll | 420, 664 | ||
| 40 | 43.9 | Carotenoid | 438(sh), 470, 502 | 857.9 | 591.5, 441.6 |
| 41 | 49.4 | Carotenoid | 468(sh), 494, 528 | ||
| 42 | 51.7 | Not identified | 517.5 | 499.4, 488.5 |
| Temp. | Solvent | Molecular weights of the carbohydrate fractions | ||||||
| >805 kDa | 804–89 kDa | 89–24 kDa | 24–5 kDa | 5–1 kDa | 1-0.4 kDa | 0.39–<0.342 kDa | ||
| 40 ˚C | 0% EtOH | >805 (18%) | 696 (14%) | 52.5 (8%) | 8.4 (8%) | 3.8 (19%) | 0.5 (4%) | <0.342 (29%) |
| 50% EtOH | >805 (4%) | 483.1 (8%) | 24.3 (5%) | 7.5 (12%) | 3.7 (24%) | 0.9 (9%) | <0.342 (38%) | |
| 100% EtOH | >805 (21%) | - | - | - | 2 (24%) | - | <0.342 (55%) | |
| 110 ˚C | 0% EtOH | >805 (9%) | 651.3 (32%) | - | 8.1 (17%) | 3.8 (25%) | 0.4 (8%) | <0.342 (7%) |
| 50% EtOH | >805 (15%) | 505.6 (11%) | 22.8 (9%) | 8.2 (12%) | 3.8 (16%) | 0.8 (12%) | <0.342 (25%) | |
| 100% EtOH | >805 (49%) | - | - | - | 4.5 (19%) | 0.6 (32%) | - | |
| 180 ˚C | 0% EtOH | >805 (11%) | - | 26.1 (10%) | 8.1 (35%) | 3.2 (35%) | - | <0.342 (9%) |
| 50% EtOH | >805 (14%) | 585.2 (5%) | 17.3 (16%) | 8.1 (19%) | 3.2 (30%) | - | <0.342 (16%) | |
| 100% EtOH | >805 (36%) | - | - | 5.2 (9%) | 3 (21%) | 0.5 (12%) | <0.342 (22%) | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muñóz-Almagro, N.; Gilbert-López, B.; M. Carmen, P.-R.; García-Fernandez, Y.; Almeida, C.; Villamiel, M.; Mendiola, J.A.; Ibáñez, E. Exploring the Microalga Euglena cantabrica by Pressurized Liquid Extraction to Obtain Bioactive Compounds. Mar. Drugs 2020, 18, 308. https://doi.org/10.3390/md18060308
Muñóz-Almagro N, Gilbert-López B, M. Carmen P-R, García-Fernandez Y, Almeida C, Villamiel M, Mendiola JA, Ibáñez E. Exploring the Microalga Euglena cantabrica by Pressurized Liquid Extraction to Obtain Bioactive Compounds. Marine Drugs. 2020; 18(6):308. https://doi.org/10.3390/md18060308
Chicago/Turabian StyleMuñóz-Almagro, Nerea, Bienvenida Gilbert-López, Pozuelo-Rollón M. Carmen, Yolanda García-Fernandez, Carlos Almeida, Mar Villamiel, Jose A. Mendiola, and Elena Ibáñez. 2020. "Exploring the Microalga Euglena cantabrica by Pressurized Liquid Extraction to Obtain Bioactive Compounds" Marine Drugs 18, no. 6: 308. https://doi.org/10.3390/md18060308
APA StyleMuñóz-Almagro, N., Gilbert-López, B., M. Carmen, P. -R., García-Fernandez, Y., Almeida, C., Villamiel, M., Mendiola, J. A., & Ibáñez, E. (2020). Exploring the Microalga Euglena cantabrica by Pressurized Liquid Extraction to Obtain Bioactive Compounds. Marine Drugs, 18(6), 308. https://doi.org/10.3390/md18060308