Chemical Diversity and Biological Activities of Meroterpenoids from Marine Derived-Fungi: A Comprehensive Update




Abstract
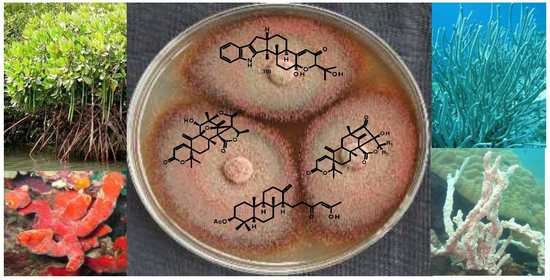
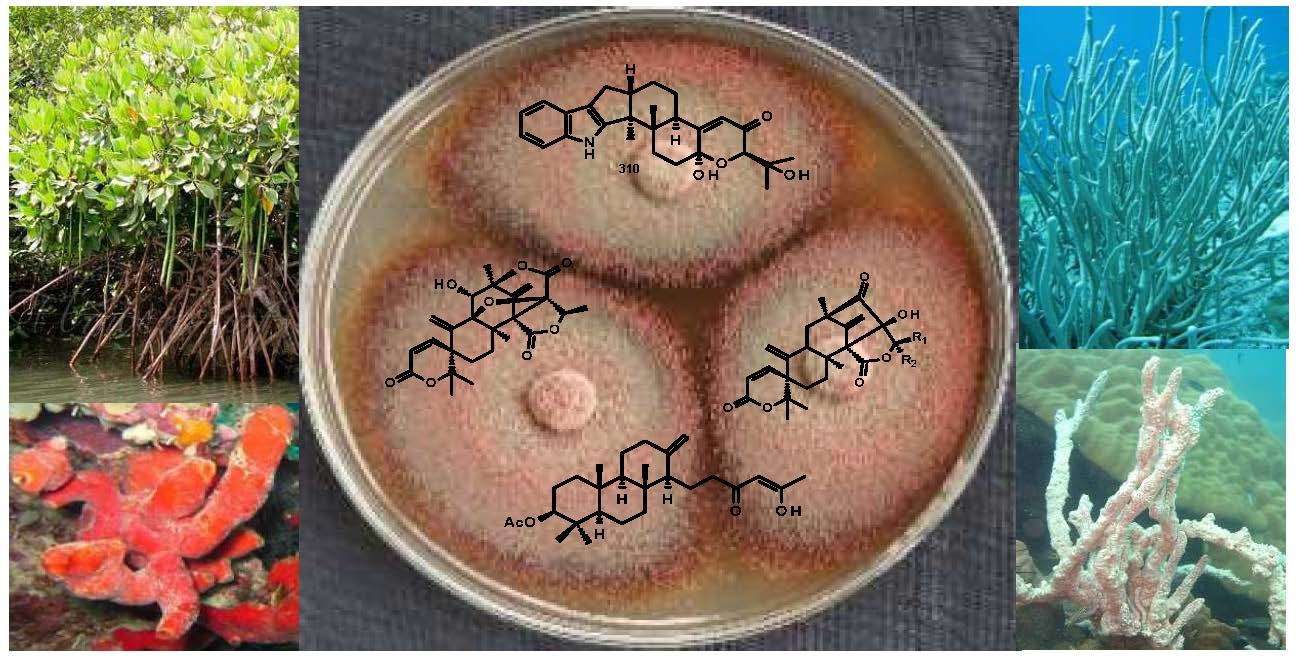
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Chemistry and Biology of Meroterpenoids Isolated from Marine-Derived Fungi
2.1. Merohemiterpenoids
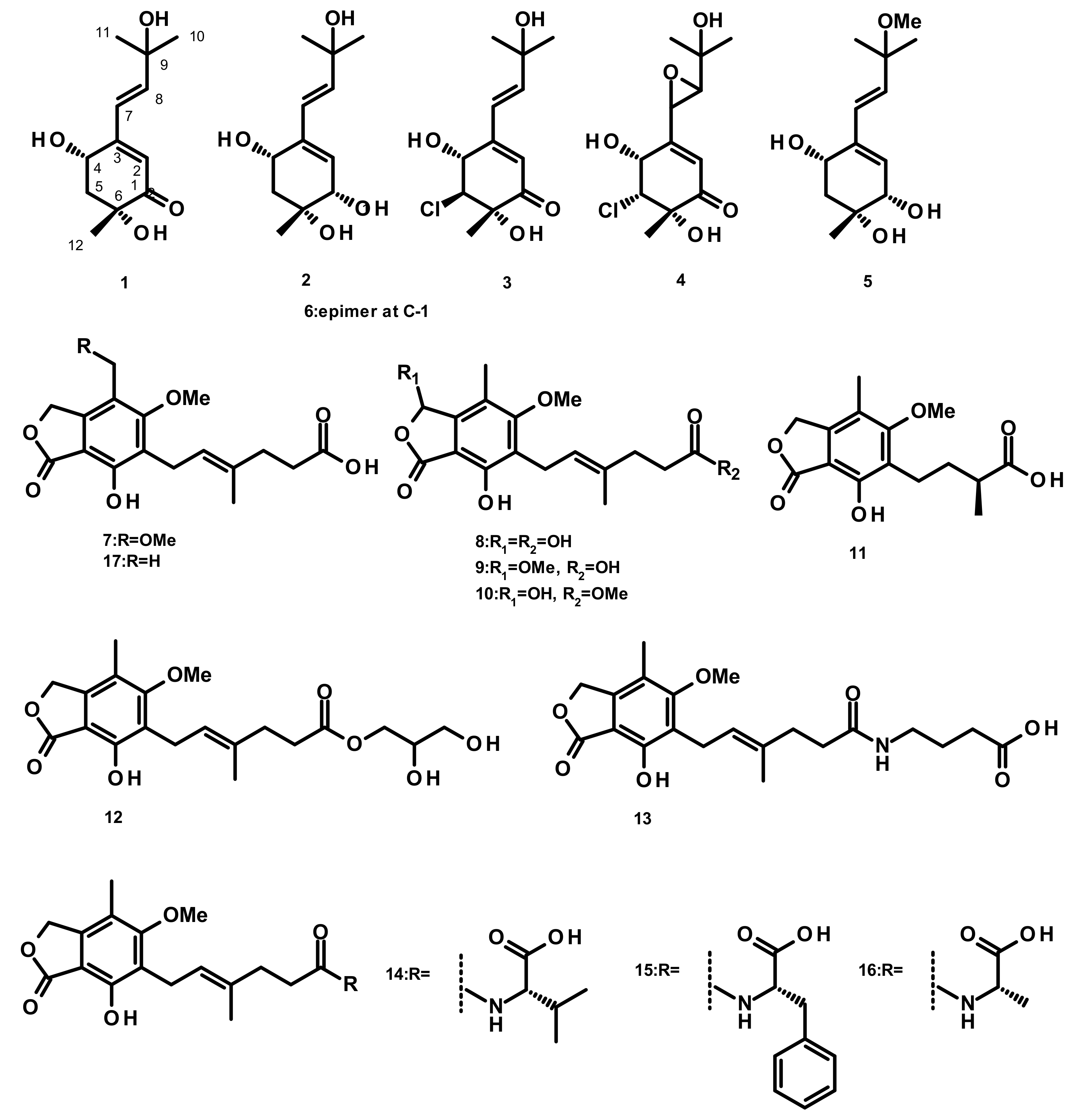
2.1.1. Merohemiterpenoids Containing Acyclic Hemiterpenes (Figure 1)
2.1.2. Merohemiterpenes Containing Cyclic Hemiterpenes (Figure 2)
2.2. Meromonoterpenoids (Figure 3)
2.3. Merosesquiterpenoids
2.3.1. Merosesquiterpenenoids Containing Acyclic Sesquiterpenes (Figures 4 and 5)
2.3.2. Meroterpenoids Containing Monocyclic Sesquiterpenes (Figure 6)
2.3.3. Meroterpenoids Containing Bicyclic Sesquiterpenes
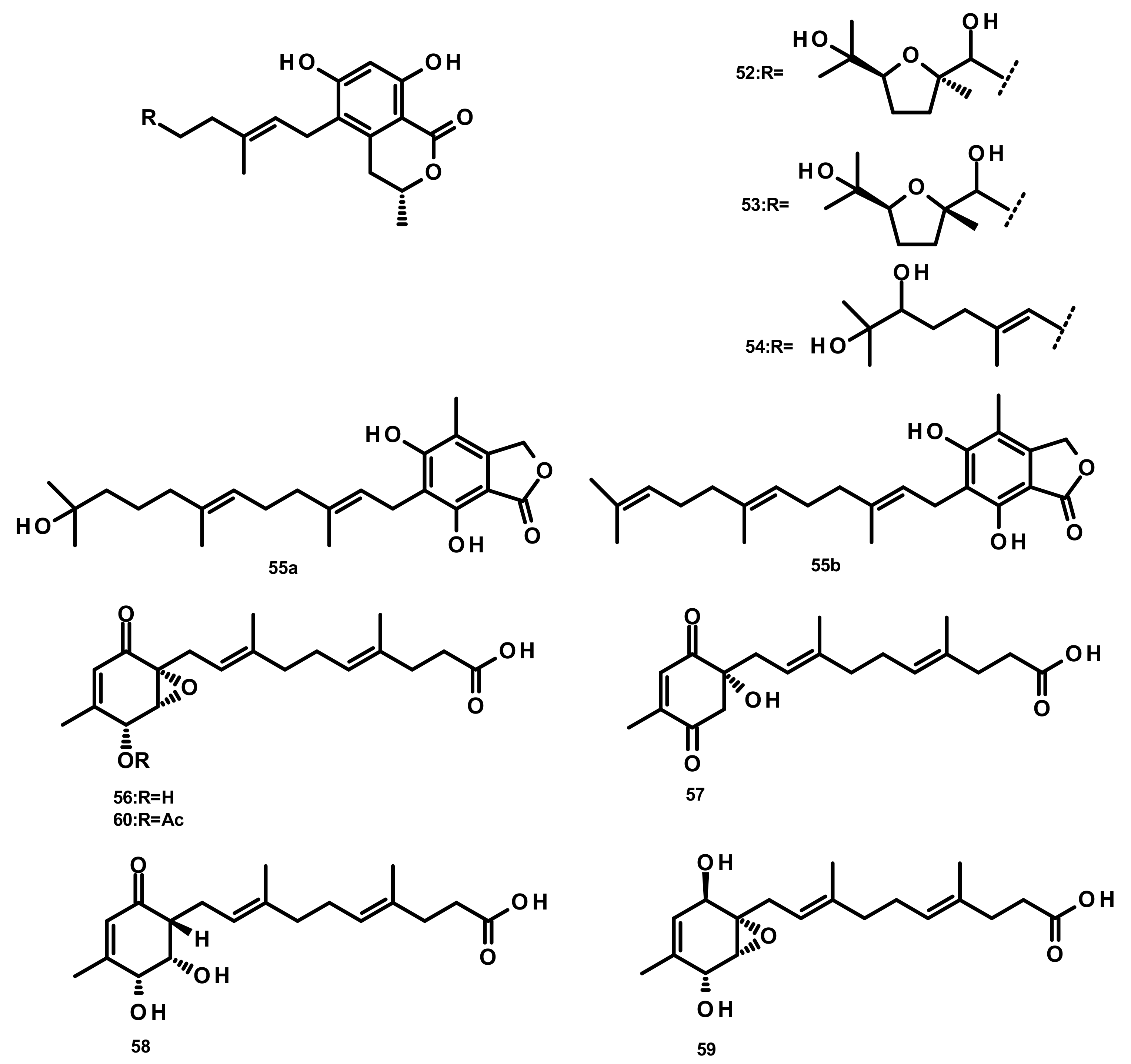
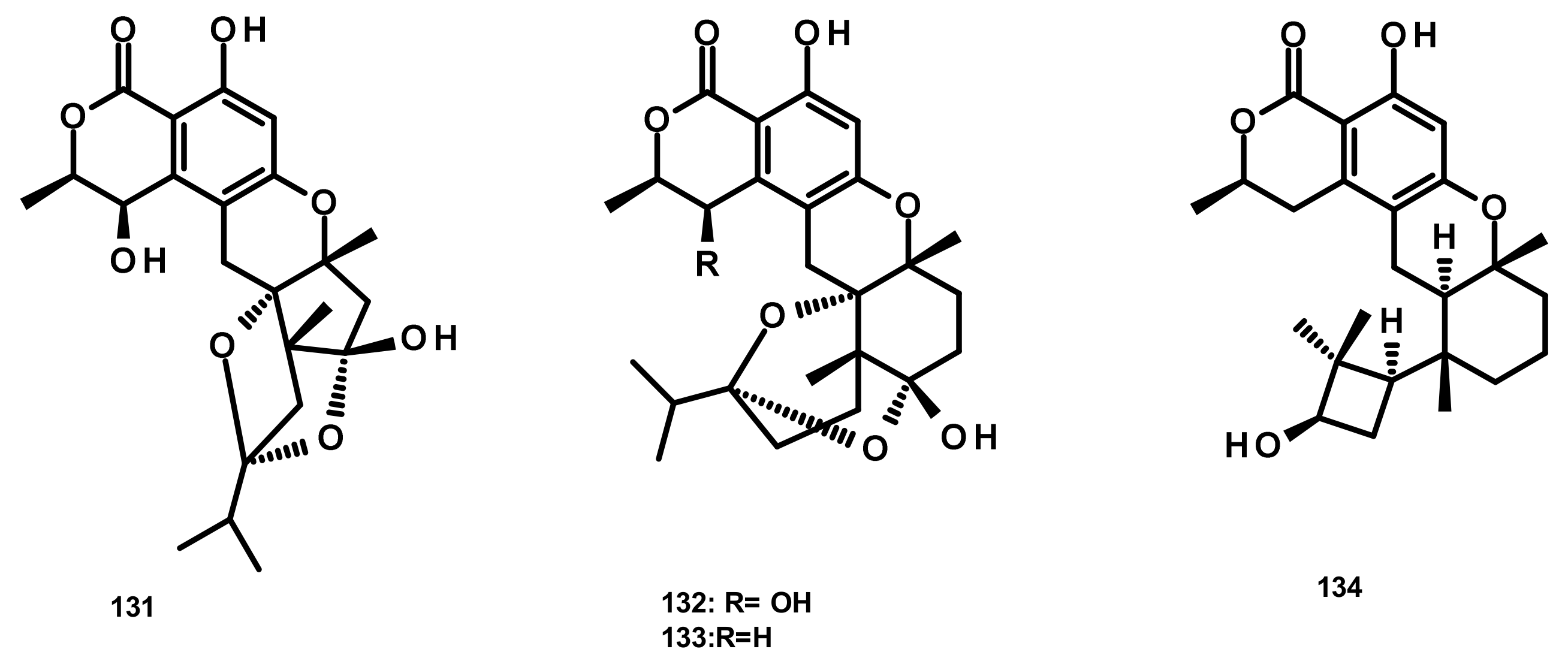
Drimane Sesquiterpenes Linked to a 2-pyrone (Figure 7)
Drimane Sesquiterpenes Linked to a Phenyl 2-pyrone (Figure 8)
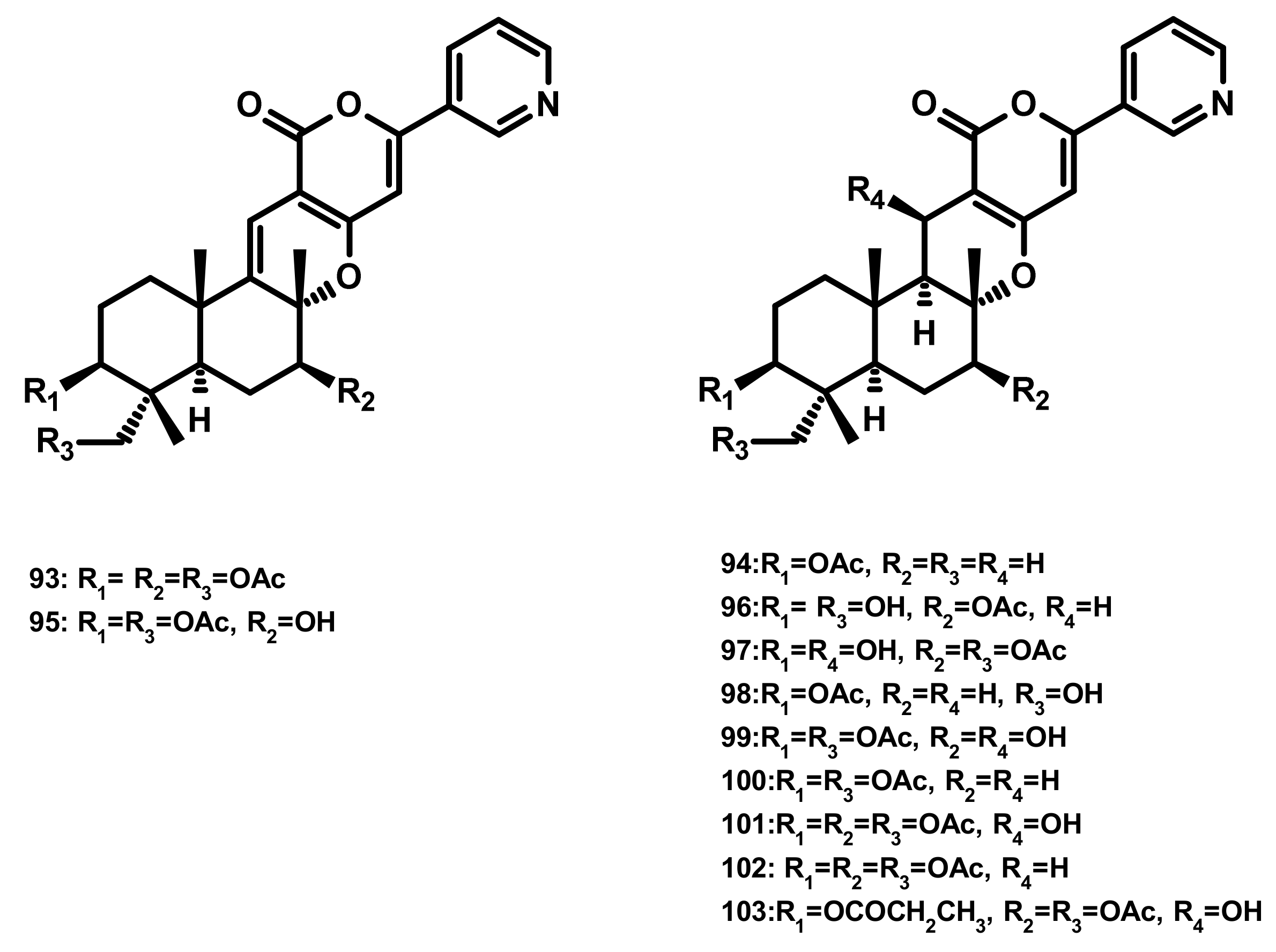
Pyripyropenes (Figure 9)
Drimane Sesquiterpenes Linked to a 4-pyrone (Figure 10)
Drimane Sesquiterpenes Linked to a Cyclohexanone Derivative by a Methylene Bridge (Microphorin-Related Compounds) (Figure 11)
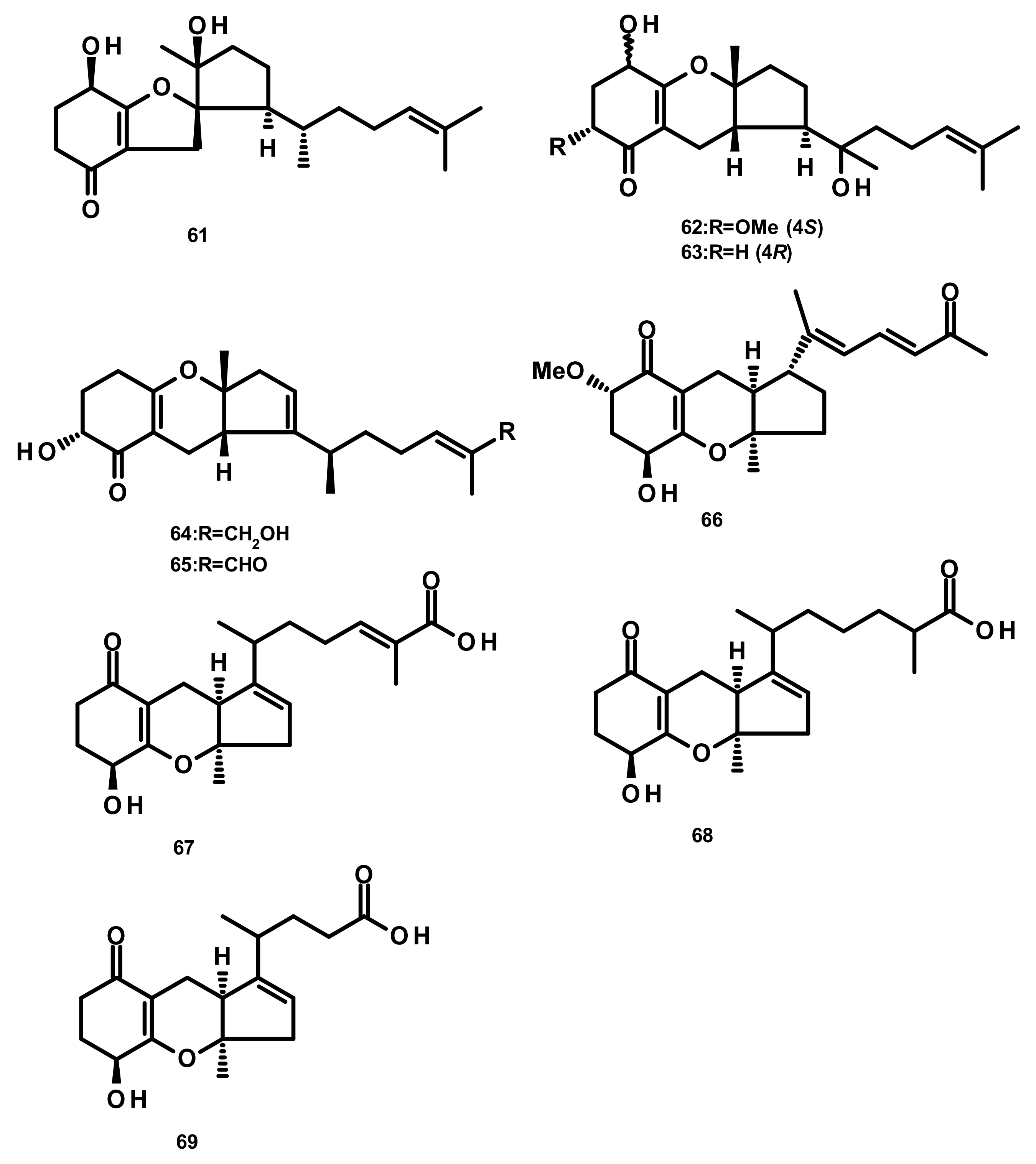
Drimane Sesquiterpenes Linked to Spirobezopyran Derivatives (Figure 12)
Drimane Sesquiterpenes Linked to Isochromone Derivatives (Figures 13 and 14)
Drimane Sesquiterpenes Linked to 5-Methylorsellinic Acid
Austalides (Figure 15)
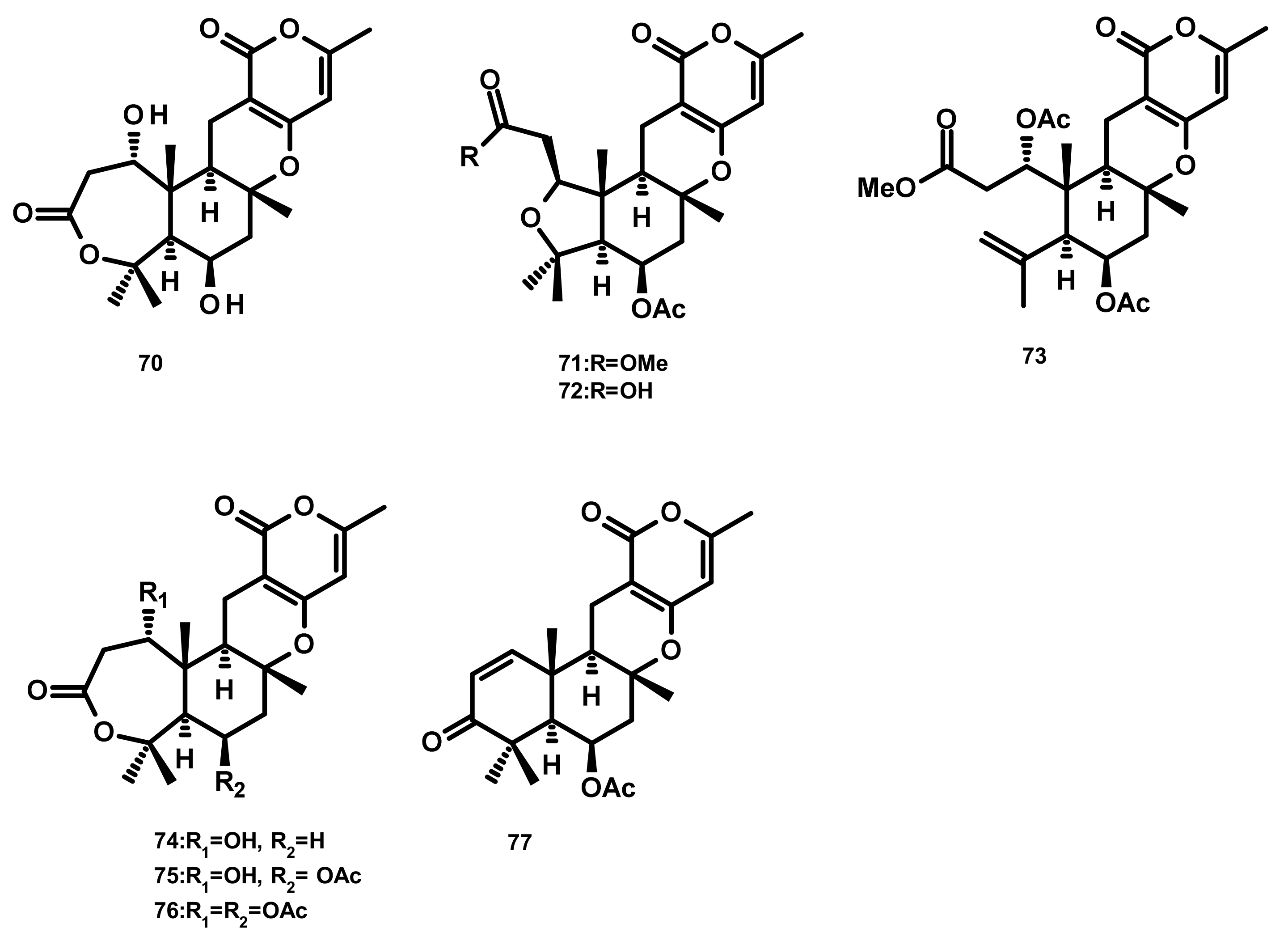
Drimane Sesquiterpenes Linked to 3,5-Methylorsellinic Acid
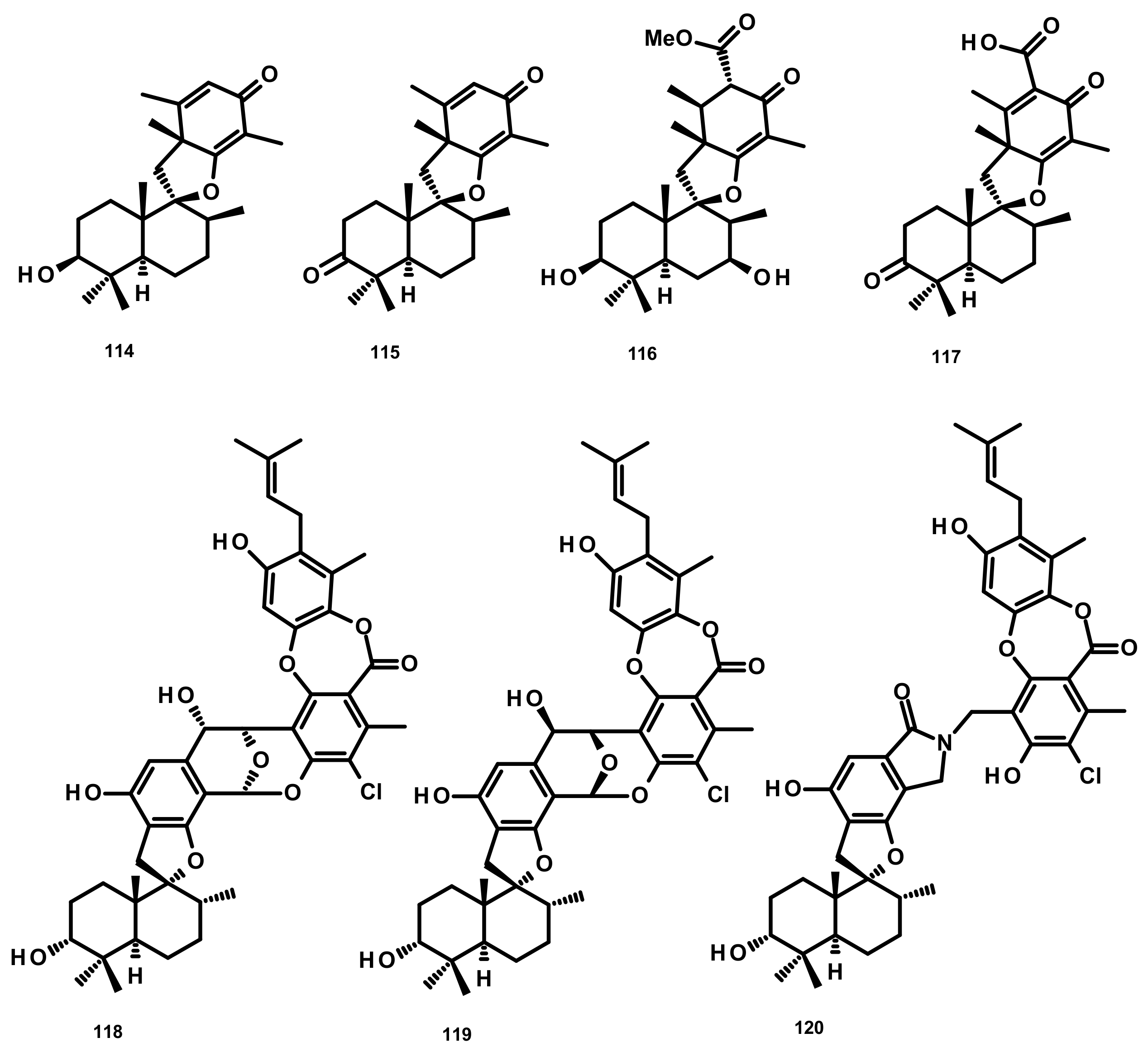
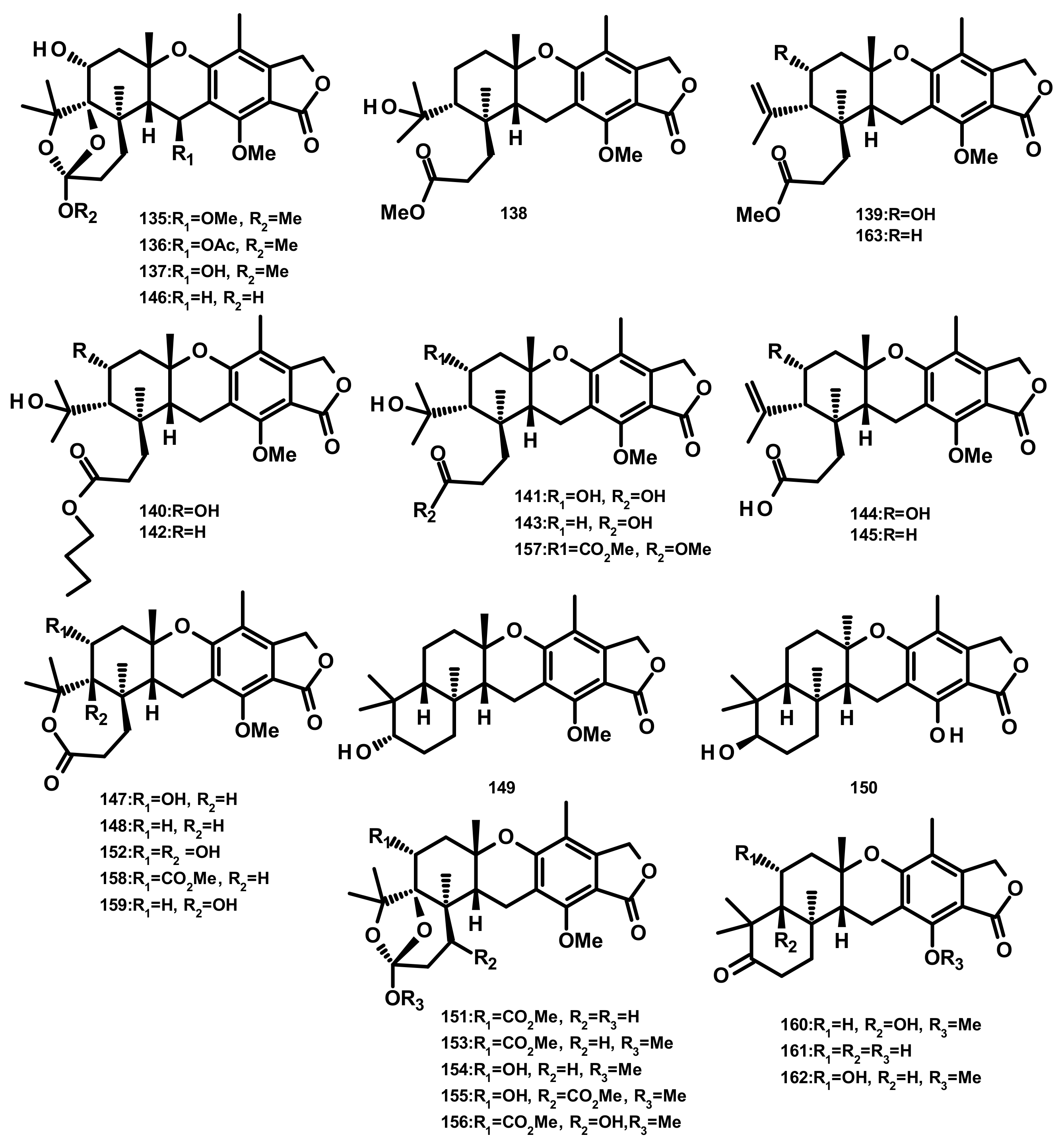
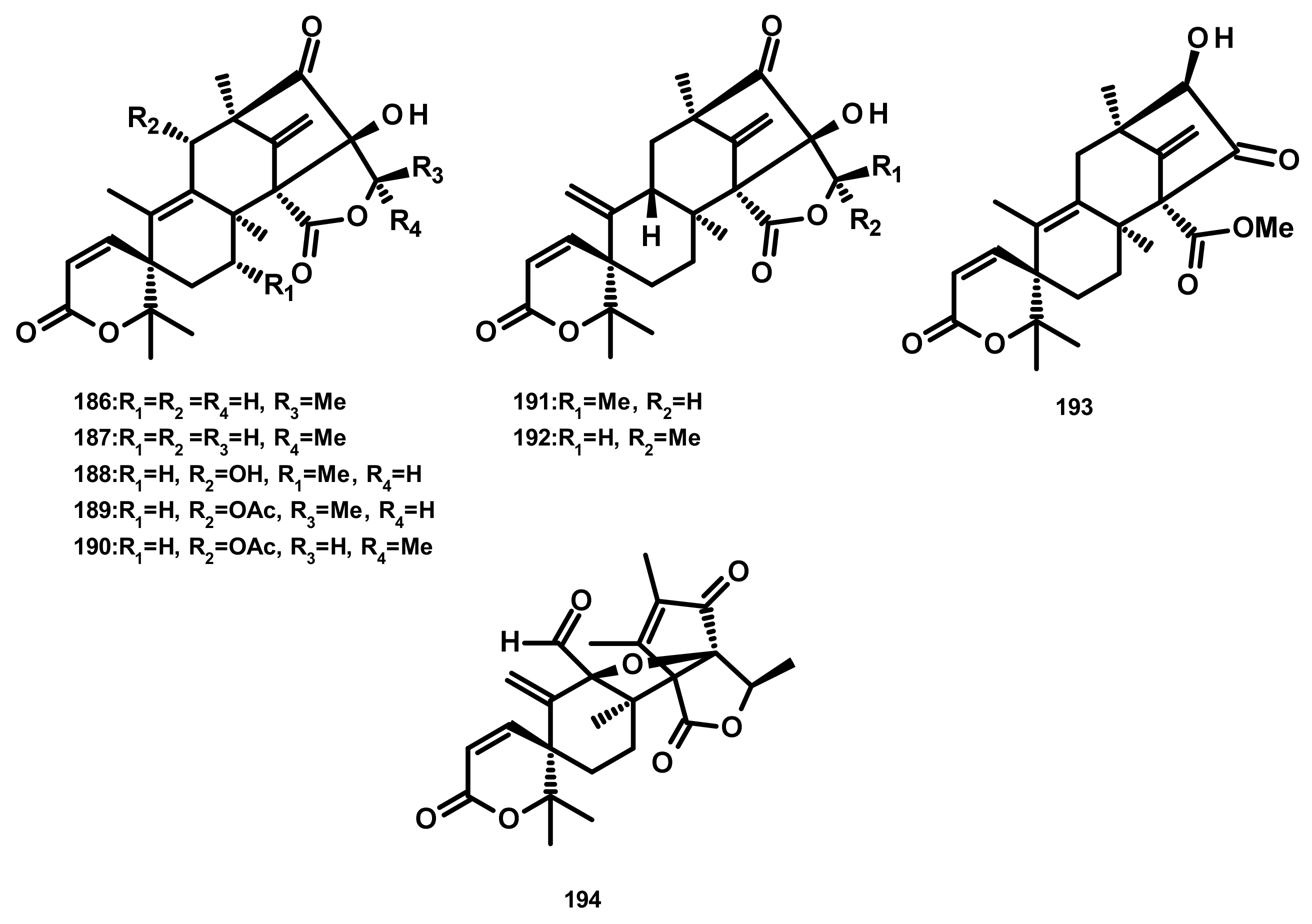
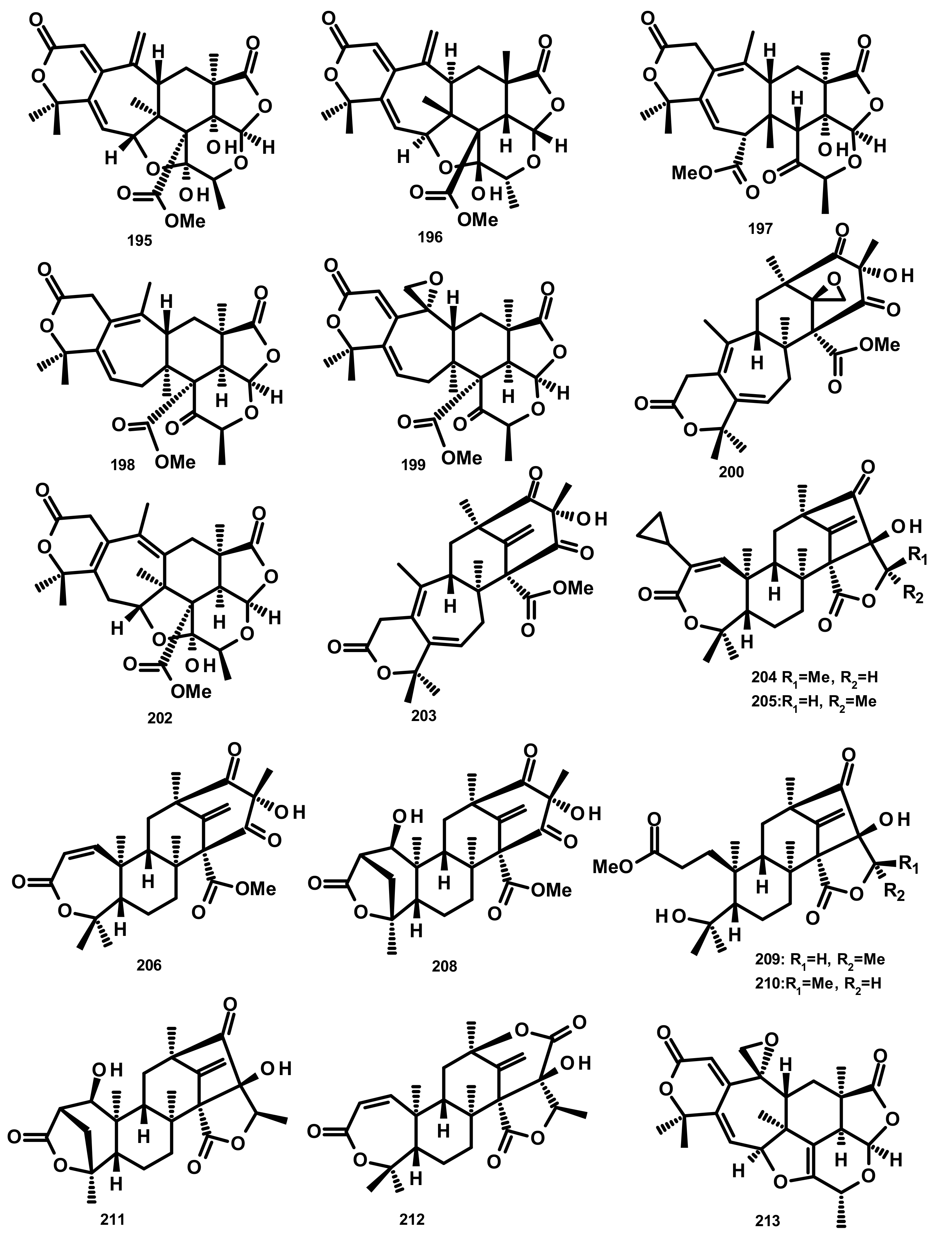
Austinoids and Related Compounds (Figures 16–18)
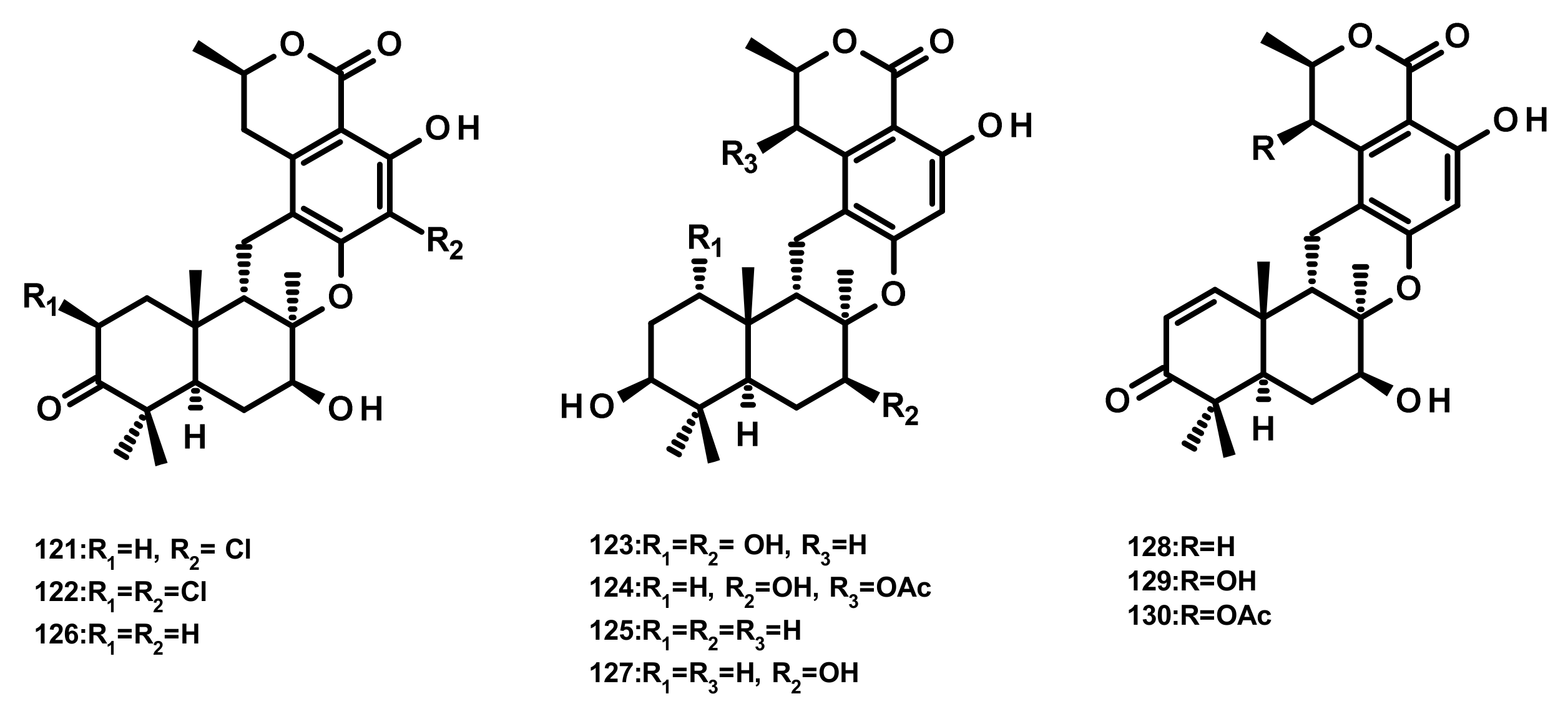
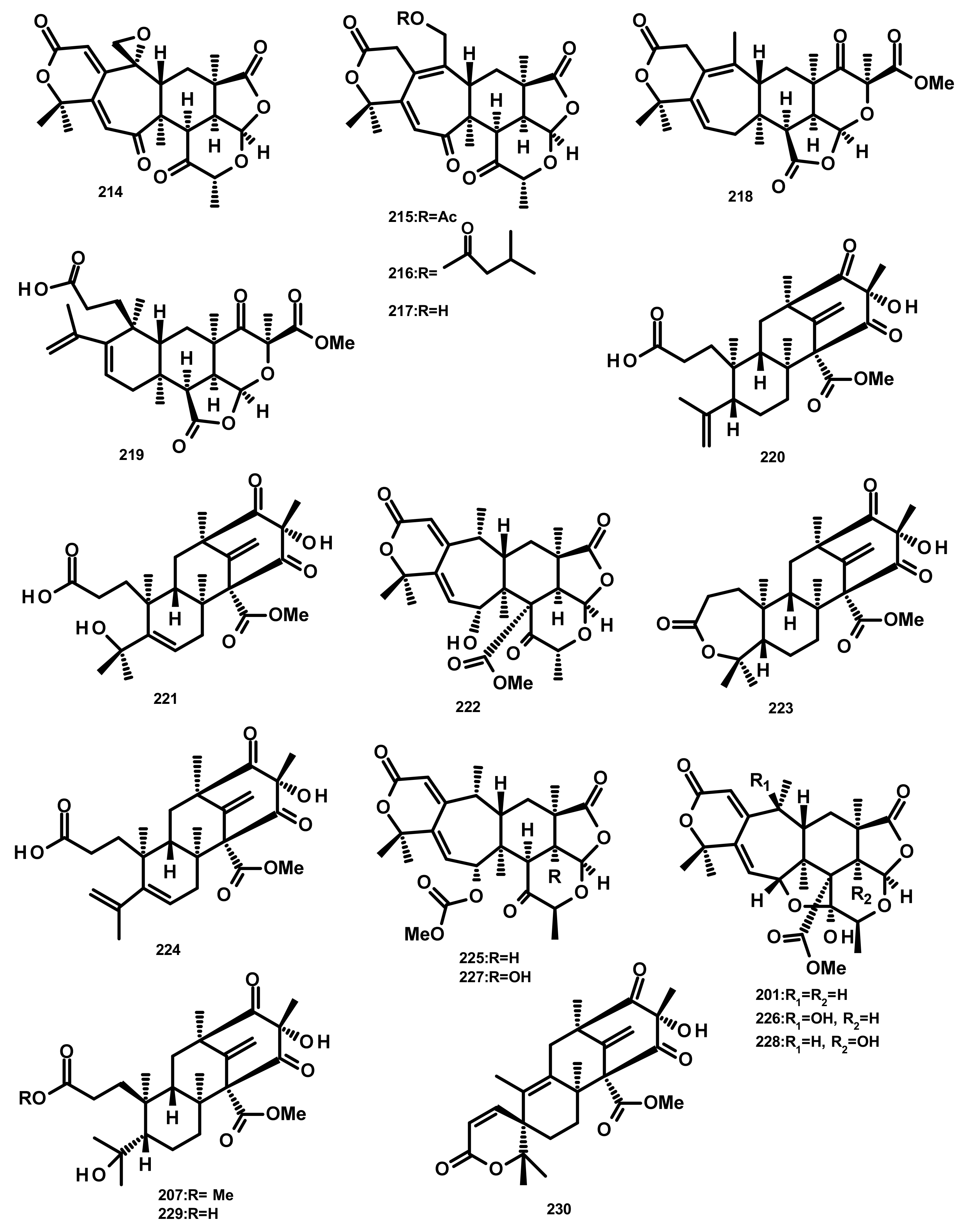
Preaustinoids and Related Compounds (Figures 19 and 20)
Terretonins and Related Compounds (Figure 21)
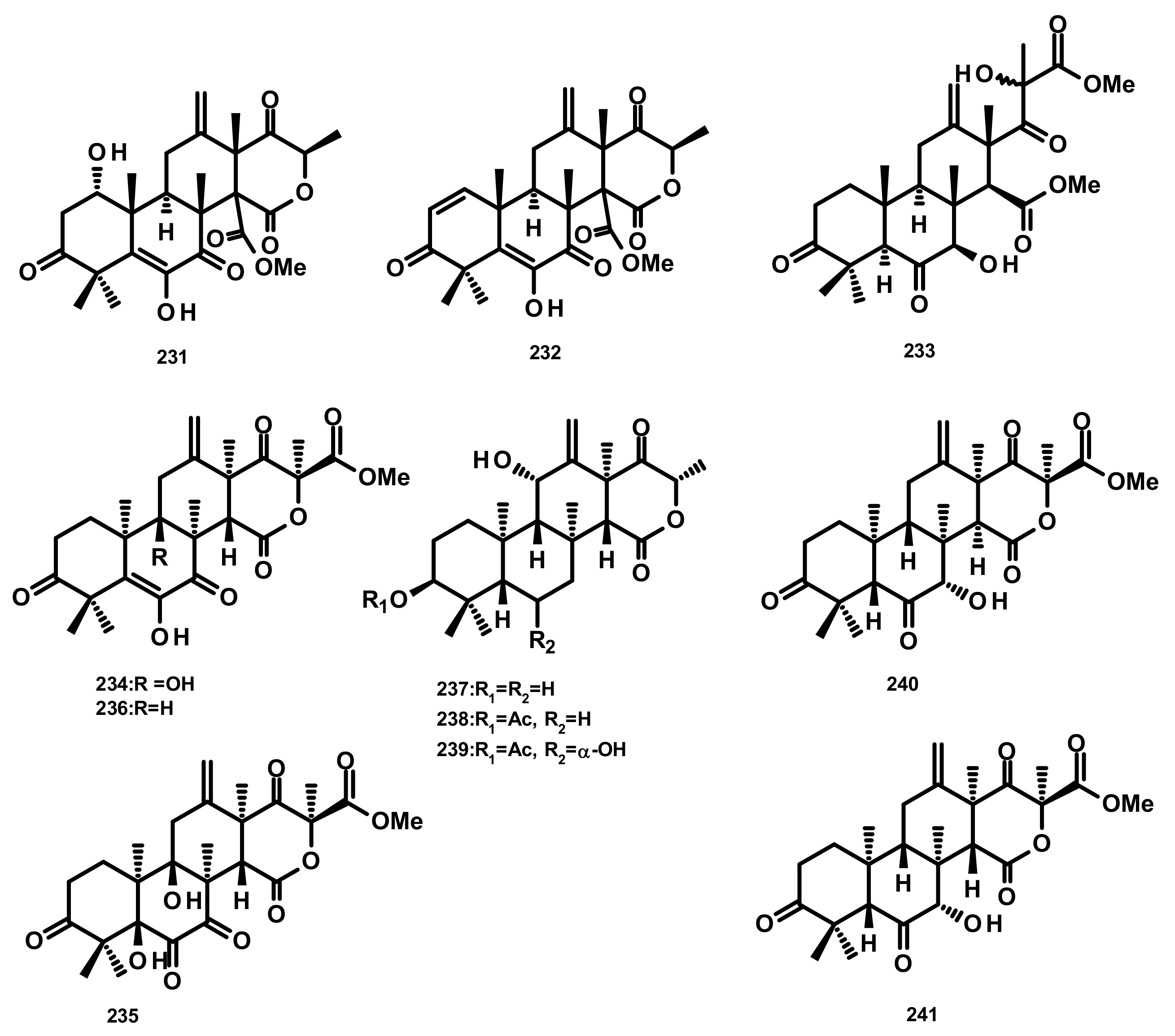
Andrastins and Related Compounds (Figure 22)
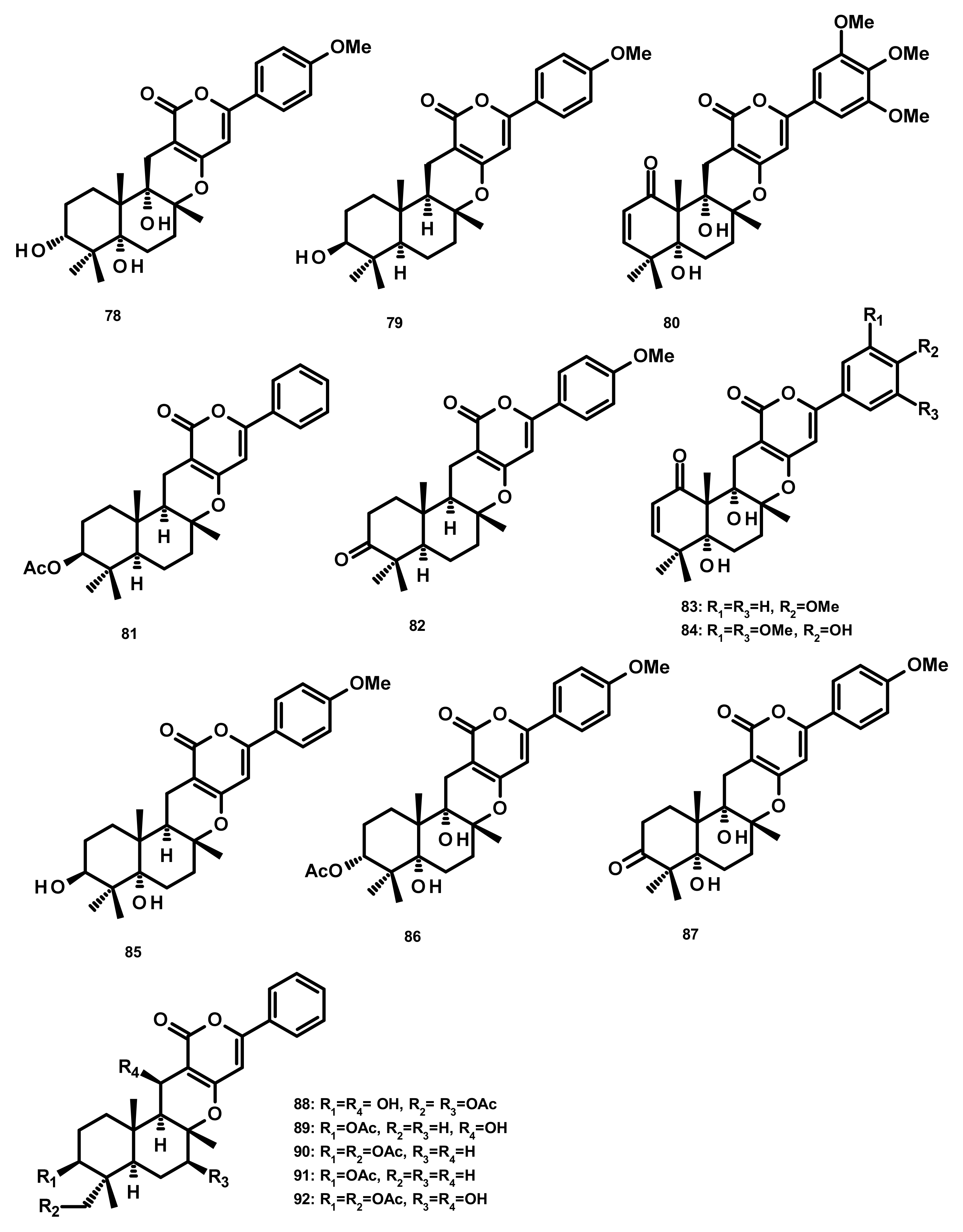
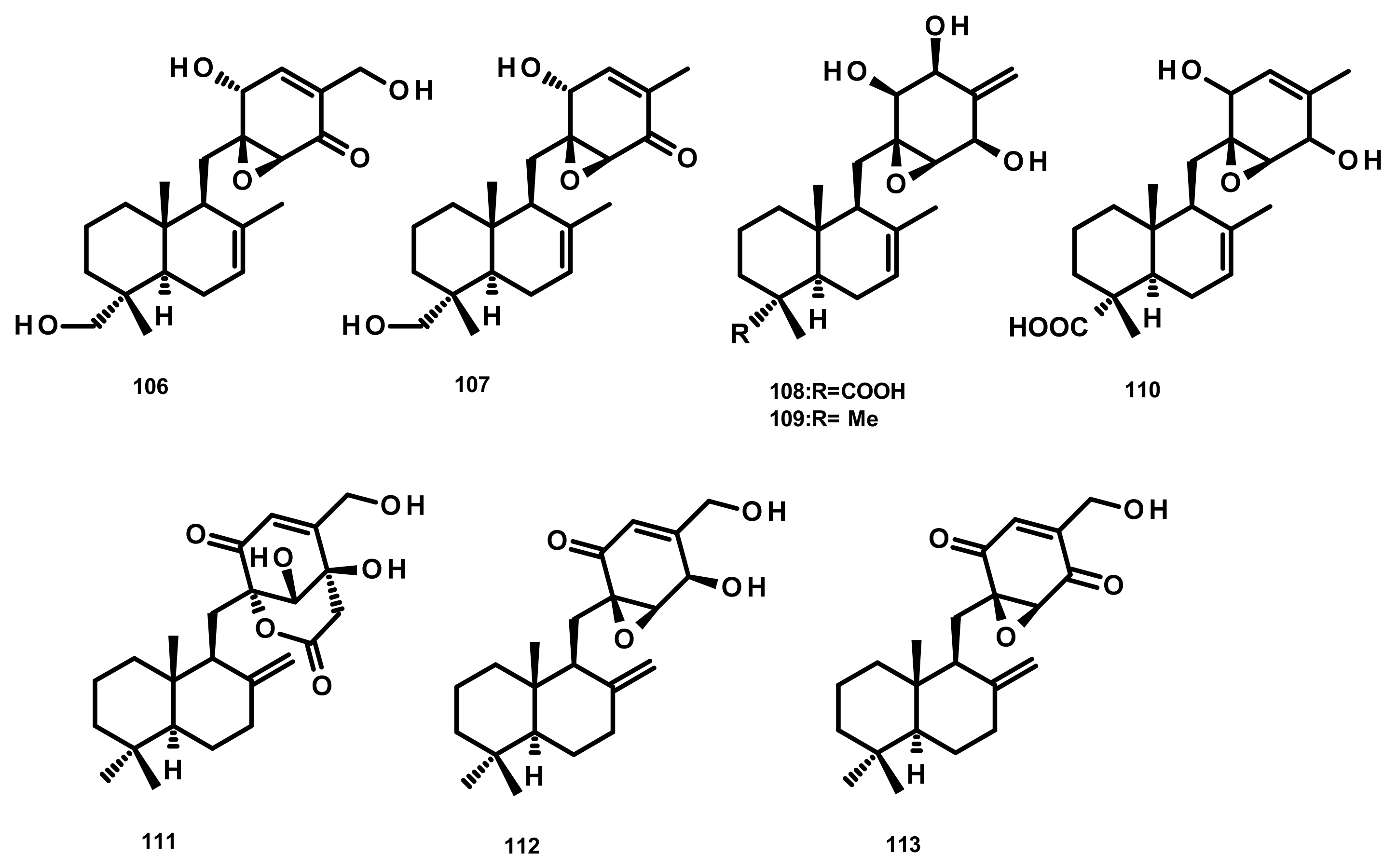
Drimane Sesquiterpenes Linked to Rearranged 3,5-dimethylorsellinic Acid (Figure 23)
Rearranged Drimane Sesquiterpenes Linked to an Isochromone (Figure 23)
2.4. Meroditerpenoids
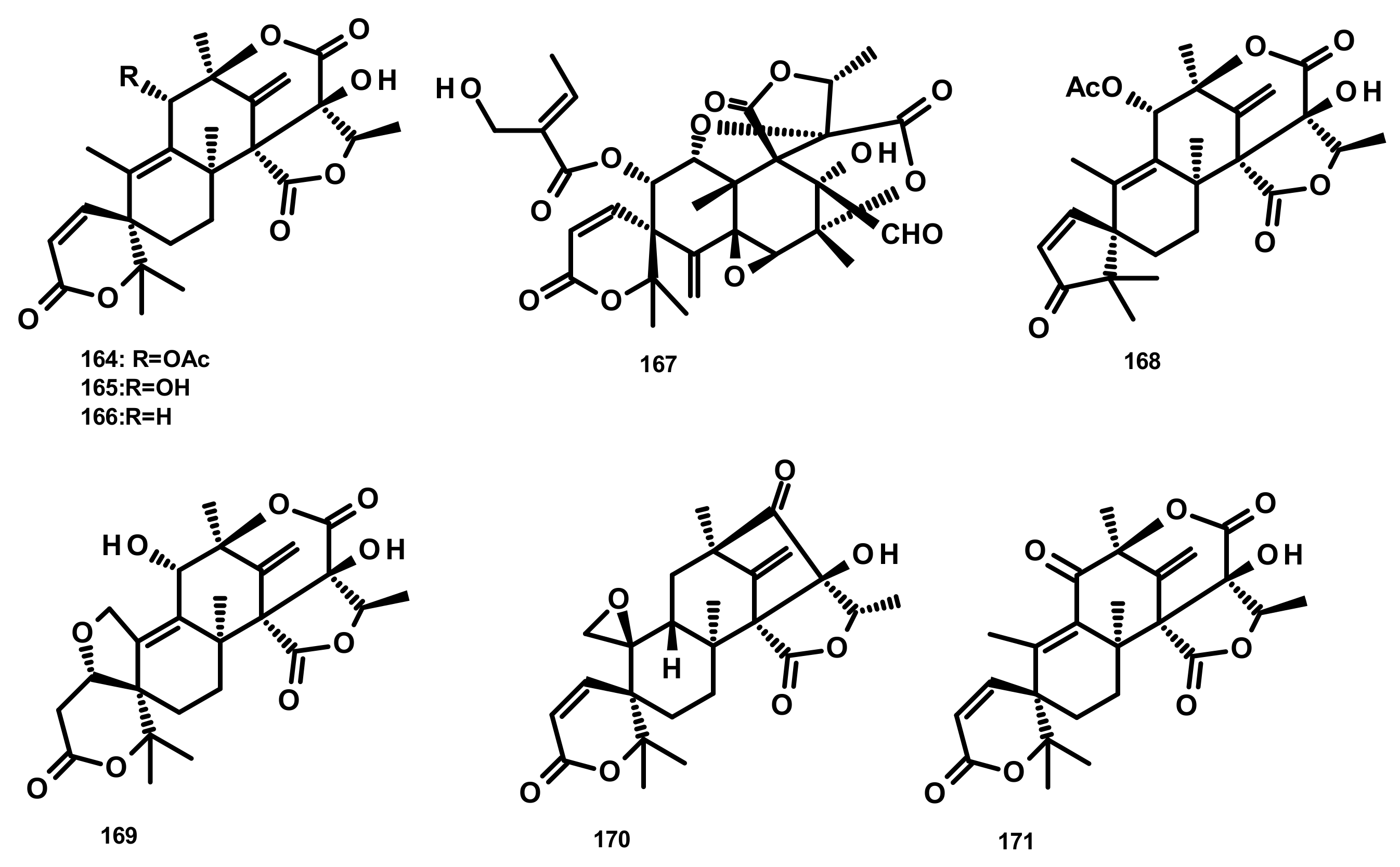
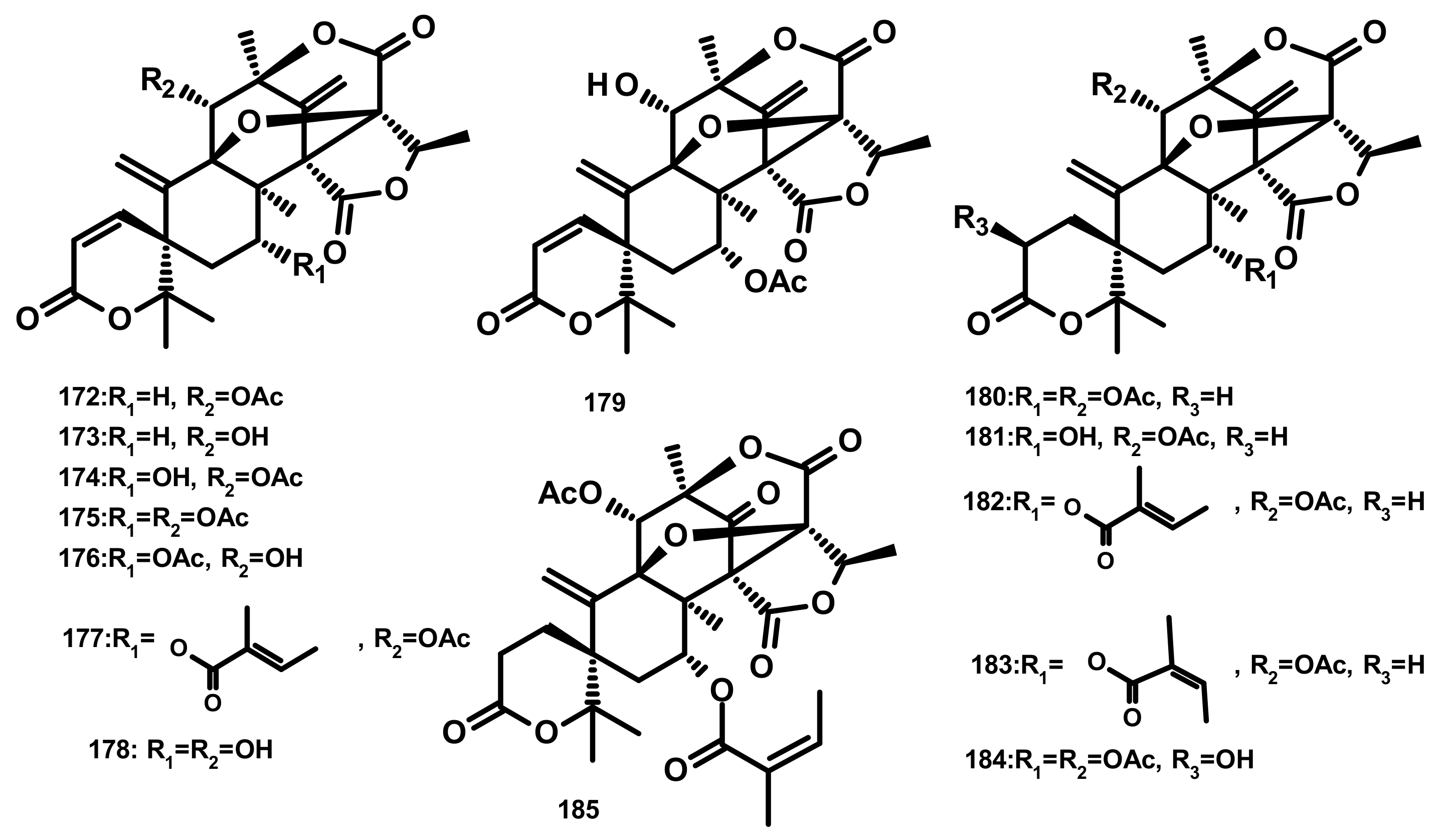
2.4.1. Diterpenes Linked to 3,5-Dimethylorsellinic Acid (Figure 24)
2.4.2. Diterpenes Linked to Polyketides (Figure 25)
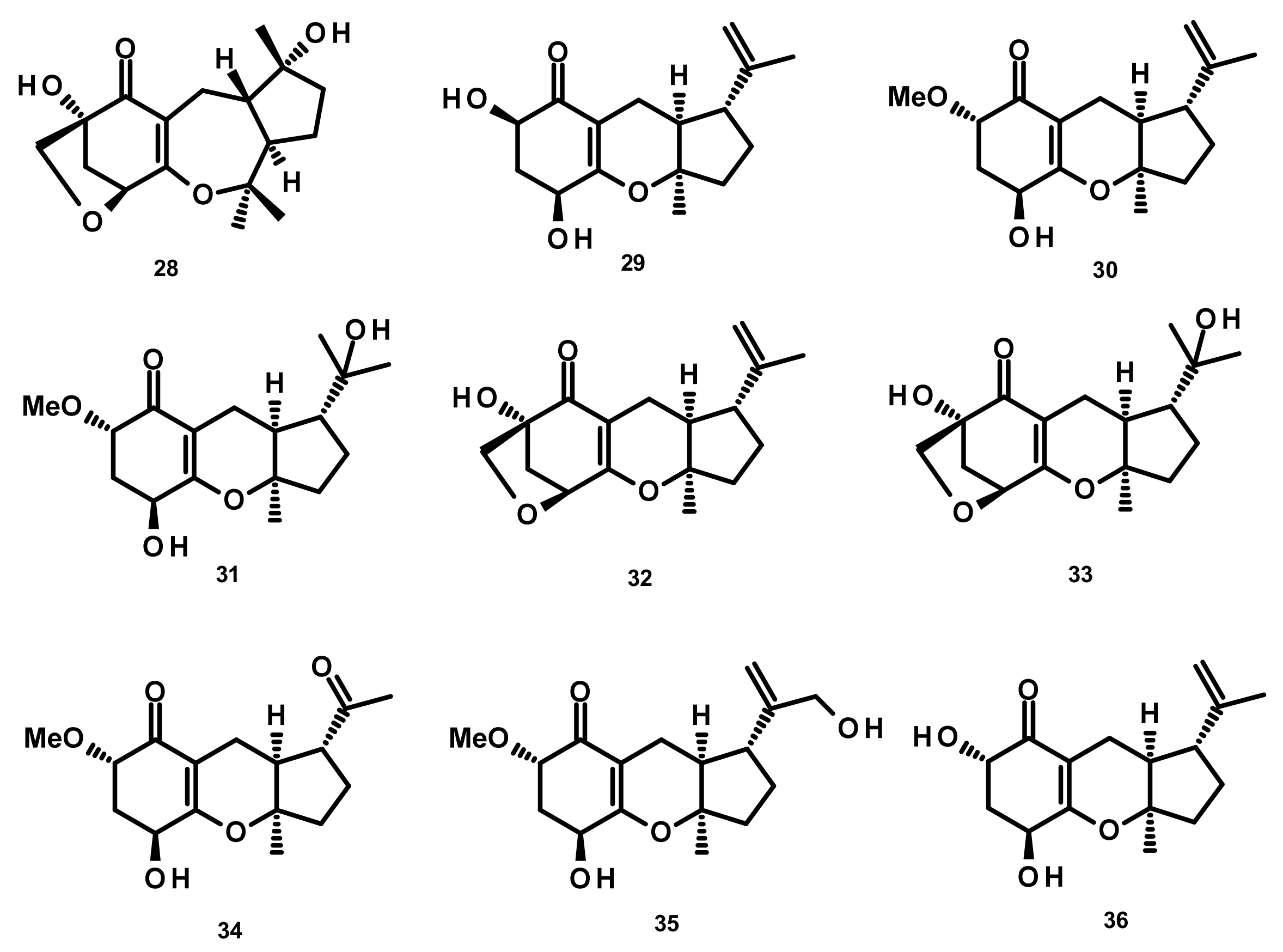
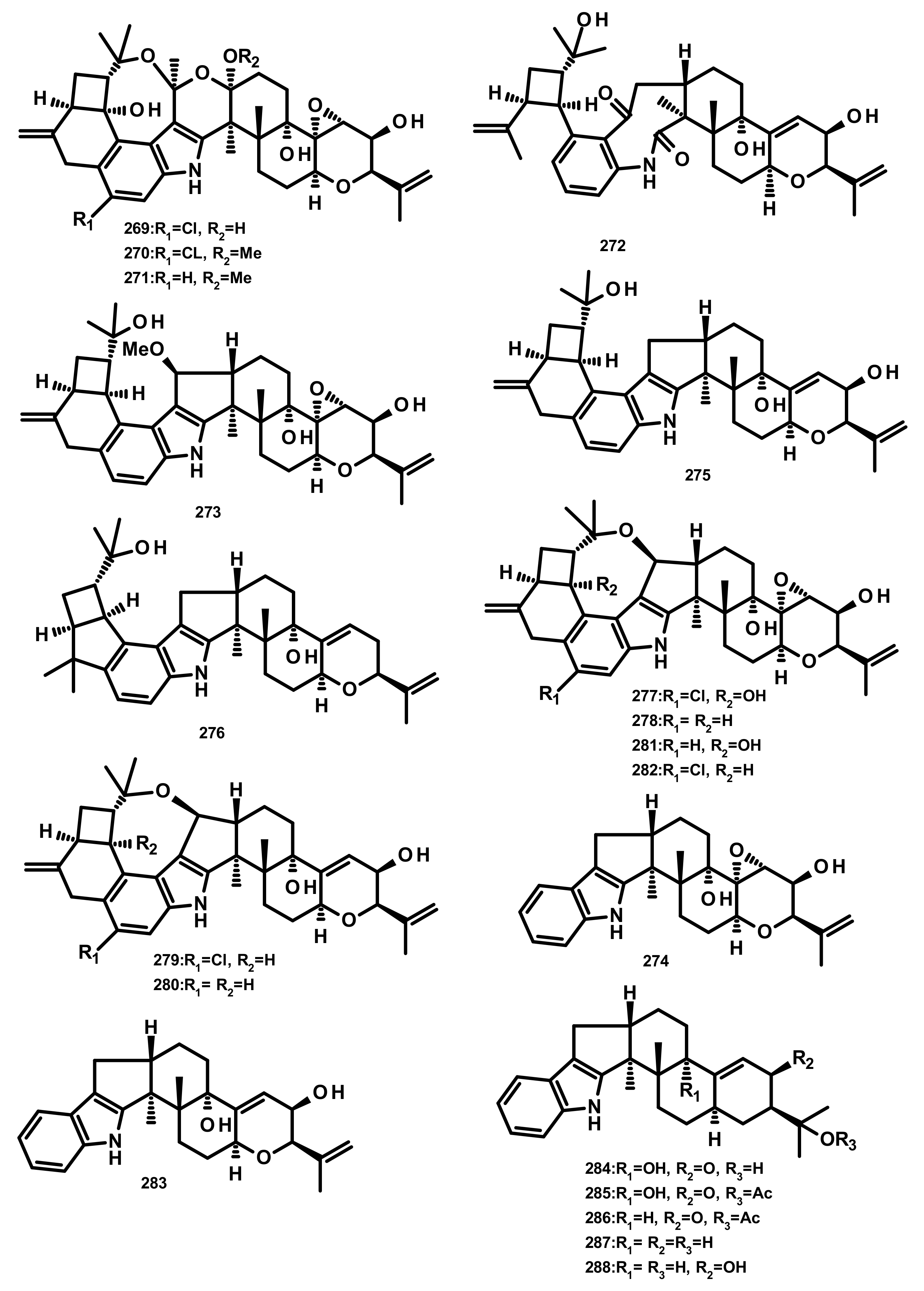
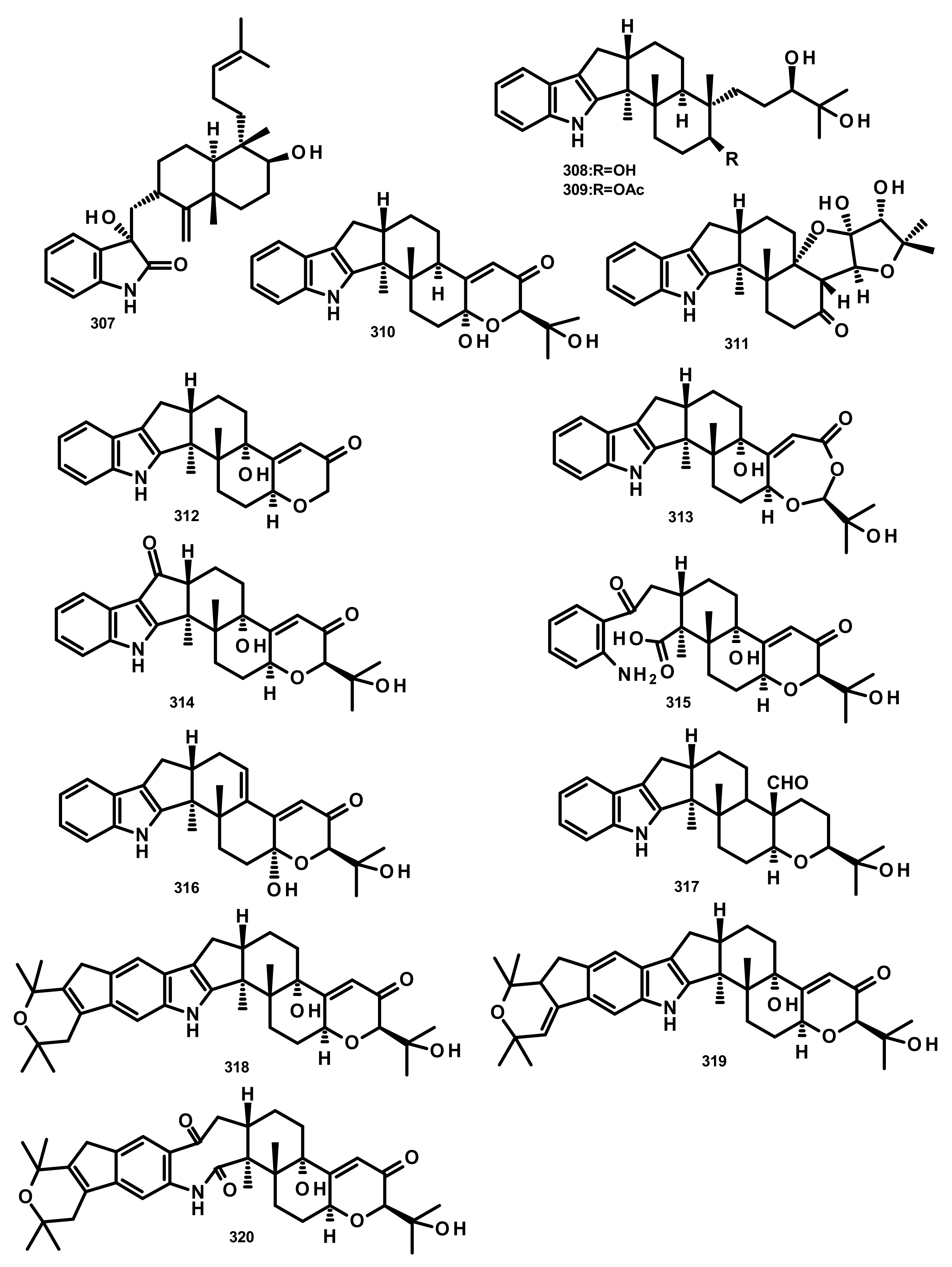
2.4.3. Indole Diterpenoids (Figure 26–28)
3. Conclusions and Prospects
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Matsuda, Y.; Abe, I. Biosynthesis of fungal meroterpenoids. Nat. Prod. Rep. 2016, 33, 26–53. [Google Scholar] [CrossRef] [Green Version]
- Peng, X.; Qiu, M. Meroterpenoids from Ganoderma species: A Review of Last Five Years. Nat. Prod. Bioprospect. 2018, 8, 137–149. [Google Scholar] [CrossRef] [Green Version]
- Menna, M.; Imperatore, C.; D’Aniello, F.; Aiello, A. Meroterpenes from marine invertebrates: Structures, occurrence, and ecological implications. Mar. Drugs 2013, 11, 1602–1643. [Google Scholar] [CrossRef] [Green Version]
- Geris, R.; Simpson, T.J. Meroterpenoids produced by fungi. Nat. Prod. Rep. 2009, 26, 1063–1094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; Legako, A.G.; Espindola, A.P.D.M.; MacMillan, J.B. Erythrolic acids A–E, meroterpenoids from a marine-derived Erythrobacter sp. J. Org. Chem. 2012, 77, 3401–3407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Y.; MacMillan, J. Biologically active meroterpenoids from Erythrobacter sp. Planta Med. 2012, 78, PI2. [Google Scholar] [CrossRef]
- Bo, G. Giuseppe Brotzu and the discovery of cephalosporins. Clin. Microbiol. Infect. 2000, 6, 6–8. [Google Scholar] [CrossRef] [Green Version]
- Fenical, W.; Jensen, P.R. Marine microorganisms: A new biomedical resource. In Marine Biotechnology; Attaway, D.H., Zaborsky, O.R., Eds.; Plenum Press: New York, NY, USA, 1993; Volume 1, pp. 419–457. ISBN 978-1-4899-2391-2. [Google Scholar]
- Bugni, T.S.; Ireland, C.M. Marine-derived fungi: A chemically and biologically diverse group of microorganisms. Nat. Prod. Rep. 2004, 21, 143–163. [Google Scholar] [CrossRef] [PubMed]
- Rateb, M.E.; Ebel, R. Secondary metabolites of fungi from marine habitats. Nat. Prod. Rep. 2011, 28, 290–344. [Google Scholar] [CrossRef]
- Petersen, L.-E.; Kellermann, M.Y.; Schupp, P.J. Secondary metabolites of marine microbes: From natural products chemistry to chemical ecology. In YOUMARES 9—The Oceans: Our Research, Our Future; Jungblut, S., Liebich, V., Bode-Dalby, M., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 159–180. ISBN 987-3-030-20388-7. [Google Scholar]
- Fisch, K.M.; Bohm, V.; Wright, A.D.; Konig, G.M. Antioxidative meroterpenoids from the brown alga Cystoseira crinita. J. Nat. Prod. 2003, 66, 968–975. [Google Scholar] [CrossRef]
- Cueto, M.; MacMillan, J.B.; Jensen, P.R.; Fenical, W. Tropolactones A-D, four meroterpenoids from a marine-derived fungus of the genus Aspergillus. Phytochemistry 2006, 67, 1826–1831. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.Z.; Cai, X.F.; Dat, N.T.; Hong, S.S.; Han, A.R.; Seo, E.K.; Hwang, B.Y.; Nan, J.X.; Lee, D.; Lee, J.J. Bisbakuchiols A and B, novel dimeric meroterpenoids from Psoralea corylifolia. Tetehedon Lett. 2007, 48, 8861–8864. [Google Scholar] [CrossRef]
- Jagels, A.; Hövelmann, Y.; Zielinski, A.; Esselen, M.; Köhler, J.; Hübner, F.; Humpf, H.-U. Stachybotrychromenes A–C: Novel cytotoxic meroterpenoids from Stachybotrys sp. Mycotoxin Res. 2018, 34, 179–185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanokmedhakul, K.; Kanokmedhakul, S.; Suwannatrai, R.; Soytong, K.; Prabpai, S.; Kongsaeree, P. Bioactive meroterpenoids and alkaloids from the fungus Eurotium chevalieri. Tetrahedron 2011, 67, 5461–5468. [Google Scholar] [CrossRef]
- Mamemura, T.; Tanaka, N.; Shibazaki, A.; Gonoi, T.; Kobayashi, J. Yojironins A-D, meroterpenoids and prenylated acylphloroglucinols from Hypericum yojiroanum. Tetrahedron Lett. 2011, 52, 3575–3578. [Google Scholar] [CrossRef]
- Peng, J.X.; Zhang, X.M.; Wang, W.; Zhu, T.J.; Gu, Q.Q.; Li, D.H. Austalides S-U, New meroterpenoids from the sponge-derived fungus Aspergillus aureolatus HDN14-107. Mar. Drugs 2016, 14. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.; Shi, Y.T.; Wu, X.D.; Wang, K.W.; Huang, S.H.; Sun, H.X.; Dickschat, J.S.; Wu, B. Talaromyolides A-D and talaromytin: Polycyclic meroterpenoids from the fungus Talaromyces sp. CX11. Org. Lett. 2019, 21, 6539–6542. [Google Scholar] [CrossRef]
- Chen, S.H.; Ding, M.; Liu, W.Y.; Huang, X.S.; Liu, Z.M.; Lu, Y.J.; Liu, H.J.; She, Z.G. Anti-inflammatory meroterpenoids from the mangrove endophytic fungus Talaromyces amestolkiae YX1. Phytochemistry 2018, 146, 8–15. [Google Scholar] [CrossRef]
- Long, Y.H.; Cui, H.; Liu, X.L.; Xiao, Z.E.; Wen, S.T.; She, Z.G.; Huang, X.S. Acetylcholinesterase inhibitory meroterpenoid from a mangrove endophytic fungus Aspergillus sp 16-5c. Molecules 2017, 22. [Google Scholar] [CrossRef] [Green Version]
- Fraser, J.A.; Lambert, L.K.; Pierens, G.K.; Bernhardt, P.V.; Garson, M.J. Secondary metabolites of the sponge-derived fungus Acremonium persicinum. J. Nat. Prod. 2013, 76, 1432–1440. [Google Scholar] [CrossRef]
- Zhang, Q.; Yang, B.; Li, F.; Liu, M.; Lin, S.; Wang, J.; Xue, Y.; Zhu, H.; Sun, W.; Hu, Z. Mycophenolic acid derivatives with immunosuppressive activity from the coral-derived fungus Penicillium bialowiezense. Mar. Drugs 2018, 16, 230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elissawy, A.M.; Ebada, S.S.; Ashour, M.L.; Ozkaya, F.C.; Ebrahim, W.; Singab, A.B.; Proksch, P. Spiroarthrinols A and B, two novel meroterpenoids isolated from the sponge-derived fungus Arthrinium sp. Phytochem. Lett. 2017, 20, 246–251. [Google Scholar] [CrossRef]
- Alves, A.J.S.; Pereira, J.A.; Dethoup, T.; Cravo, S.; Mistry, S.; Silva, A.M.S.; Pinto, M.M.M.; Kijjoa, A. A new meroterpene, a new benzofuran derivative and other constituents from cultures of the marine sponge-associated fungus Acremonium persicinum KUFA 1007 and their anticholinesterase activities. Mar. Drugs 2019, 17, 379. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mei, W.-L.; Zheng, B.; Zhao, Y.-X.; Zhong, H.-M.; Chen, X.-L.W.; Zeng, Y.-B.; Dong, W.-H.; Huang, J.-L.; Proksch, P.; Dai, H.-F. Meroterpenes from endophytic fungus A1 of mangrove plant Scyphiphora hydrophyllacea. Mar. Drugs 2012, 10, 1993–2001. [Google Scholar] [CrossRef] [PubMed]
- Bai, Z.Q.; Lin, X.P.; Wang, J.F.; Zhou, X.F.; Liu, J.; Yang, B.; Yang, X.W.; Liao, S.R.; Wang, L.S.; Liu, Y.H. New meroterpenoids from the endophytic fungus Aspergillus flavipes AIL8 derived from the mangrove plant Acanthus ilicifolius. Mar. Drugs 2015, 13, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.J.; Wu, G.W.; Zhu, T.J.; Kurtan, T.; Mandi, A.; Jiao, J.Y.; Li, J.; Qi, X.; Gu, Q.Q.; Li, D.H. Meroterpenoids with diverse ring systems from the sponge-associated fungus Alternaria sp. JJY-32. J. Nat. Prod. 2013, 76, 1946–1957. [Google Scholar] [CrossRef]
- Sun, X.H.; Kong, X.L.; Gao, H.Q.; Zhu, T.J.; Wu, G.W.; Gu, Q.Q.; Li, D.H. Two new meroterpenoids produced by the endophytic fungus Penicillium sp SXH-65. Arch. Pharm. Res. 2014, 37, 978–982. [Google Scholar] [CrossRef]
- He, W.J.; Zhou, X.J.; Qin, X.C.; Mai, Y.X.; Lin, X.P.; Liao, S.R.; Yang, B.; Zhang, T.Y.; Tu, Z.C.; Wang, J.F. Quinone/hydroquinone meroterpenoids with antitubercular and cytotoxic activities produced by the sponge-derived fungus Gliomastix sp. ZSDS1-F7. Nat. Prod. Res. 2017, 31, 604–609. [Google Scholar] [CrossRef]
- Kong, F.D.; Ma, Q.Y.; Huang, S.Z.; Wang, P.; Wang, J.F.; Zhou, L.M.; Yuan, J.Z.; Dai, H.F.; Zhao, Y.X. Chrodrimanins K-N and related meroterpenoids from the fungus Penicillium sp. SCS-KFD09 isolated from a marine worm, Sipunculus nudus. J. Nat. Prod. 2017, 80, 1039–1047. [Google Scholar] [CrossRef]
- Wang, W.; Lee, J.; Kim, K.-J.; Sung, Y.; Park, K.-H.; Oh, E.; Park, C.; Son, Y.-J.; Kang, H. Austalides, osteoclast differentiation inhibitors from a marine-derived strain of the fungus Penicillium rudallense. J. Nat. Prod. 2019, 82, 3083–3088. [Google Scholar] [CrossRef]
- Cheng, Z.; Xu, W.; Liu, L.; Li, S.; Yuan, W.; Luo, Z.; Zhang, J.; Cheng, Y.; Li, Q. Peniginsengins B–E, new farnesylcyclohexenones from the deep sea-derived fungus Penicillium sp. YPGA11. Mar. Drugs 2018, 16, 358. [Google Scholar] [CrossRef] [Green Version]
- Fang, S.-T.; Miao, F.-P.; Liu, X.-H.; Song, Y.-P.; Ji, N.-Y. Two new tricycloalternarene acids from the marine-derived fungus Alternaria alternata ICD5-11. Phytochem. Lett. 2018, 23, 185–188. [Google Scholar] [CrossRef]
- Yurchenko, A.N.; Smetanina, O.F.; Kalinovsky, A.I.; Pivkin, M.V.; Dmitrenok, P.S.; Kuznetsova, T.A. A new meroterpenoid from the marine fungus Aspergillus versicolor (Vuill.) Tirab. Russ. Chem. Bull. 2010, 59, 852–856. [Google Scholar] [CrossRef]
- Li, H.Q.; Sun, W.G.; Deng, M.Y.; Qi, C.X.; Chen, C.M.; Zhu, H.C.; Luo, Z.W.; Wang, J.P.; Xue, Y.B.; Zhang, Y.H. Asperversins A and B, two novel meroterpenoids with an unusual 5/6/6/6 ring from the marine-derived fungus Aspergillus versicolor. Mar. Drugs 2018, 16, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.; Sun, X.; Ding, B.; Lin, M.; Liu, L.; Huang, H.; She, Z. A New anti-acetylcholinesterase α-pyrone meroterpene, arigsugacin I, from mangrove endophytic fungus Penicillium sp. sk5GW1L of Kandelia candel. Planta Med. 2013, 79, 1572–1575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prompanya, C.; Dethoup, T.; Bessa, L.J.; Pinto, M.M.M.; Gales, L.; Costa, P.M.; Silva, A.M.S.; Kijjoa, A. New isocoumarin derivatives and meroterpenoids from the marine sponge-associated fungus Aspergillus similanensis sp. nov. KUFA 0013. Mar. Drugs 2014, 12, 5160–5173. [Google Scholar] [CrossRef] [PubMed]
- Ding, B.; Wang, Z.Y.; Huang, X.S.; Liu, Y.Y.; Chen, W.R.; She, Z.G. Bioactive α-pyrone meroterpenoids from mangrove endophytic fungus Penicillium sp. Nat. Prod. Res. 2016, 30, 2805–2812. [Google Scholar] [CrossRef]
- Ding, Z.; Zhang, L.; Fu, J.; Che, Q.; Li, D.; Gu, Q.; Zhu, T. Phenylpyropenes E and F: New meroterpenes from the marine-derived fungus Penicillium concentricum ZLQ-69. J. Antibiot. 2015, 68, 748–751. [Google Scholar] [CrossRef]
- Prompanya, C.; Fernandes, C.; Cravo, S.; Pinto, M.M.M.; Dethoup, T.; Silva, A.M.S.; Kijjoa, A. A new cyclic hexapeptide and a new isocoumarin derivative from the marine sponge-associated fungus Aspergillus similanensis KUFA 0013. Mar. Drugs 2015, 13, 1432–1450. [Google Scholar] [CrossRef] [Green Version]
- Cao, Q.-X.; Wei, J.-H.; Deng, R.; Feng, G.-K.; Zhu, X.-F.; Lan, W.-J.; Li, H.-J. Two new pyripyropenes from the marine fungus Fusarium lateritium 2016F18-1. Chem. Biodivers. 2017, 14. [Google Scholar] [CrossRef]
- Lan, W.-J.; Fu, S.-J.; Xu, M.-Y.; Liang, W.-L.; Lam, C.-K.; Zhong, G.-H.; Xu, J.; Yang, D.-P.; Li, H.-J. Five new cytotoxic metabolites from the marine fungus Neosartorya pseudofischeri. Mar. Drugs 2016, 14, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liao, L.J.; Lee, J.H.; You, M.J.; Choi, T.J.; Park, W.; Lee, S.K.; Oh, D.C.; Oh, K.B.; Shin, J. Penicillipyrones A and B, meroterpenoids from a marine-derived Penicillium sp. fungus. J. Nat. Prod. 2014, 77, 406–410. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, I.E.; Gross, H.; Pontius, A.; Kehraus, S.; Krick, A.; Kelter, G.; Maier, A.; Fiebig, H.-H.; König, G.M. Epoxyphomalin A and B, prenylated polyketides with potent cytotoxicity from the marine-derived fungus Phoma sp. Org. Lett. 2009, 11, 5014–5017. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, I.E.; Kehraus, S.; Krick, A.; König, G.M.; Kelter, G.; Maier, A.; Fiebig, H.-H.; Kalesse, M.; Malek, N.P.; Gross, H. Mode of action of epoxyphomalins A and B and characterization of related metabolites from the marine-derived fungus Paraconiothyrium sp. J. Nat. Prod. 2010, 73, 2053–2056. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Li, X.M.; Liu, Y.; Zhang, P.; Wang, J.N.; Wang, B.G. Chermesins A-D: Meroterpenoids with a drimane-type spirosesquiterpene skeleton from the marine algal-derived endophytic fungus Penicillium chermesinum EN-480. J. Nat. Prod. 2016, 79, 806–811. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.; Li, Y.; Li, X.D.; Cheng, Z.B.; Huang, J.; Proksch, P.; Lin, W.H. Chartarolides A-C, novel meroterpenoids with antitumor activities. Tetrahedron Lett. 2017, 58, 1826–1829. [Google Scholar] [CrossRef]
- Zhou, Y.; Mándi, A.; Debbab, A.; Wray, V.; Schulz, B.; Müller, W.E.G.; Lin, W.; Proksch, P.; Kurtán, T.; Aly, A.H. New austalides from the sponge-associated fungus Aspergillus sp. Eur. J. Org. Chem. 2011, 6009–6019. [Google Scholar] [CrossRef] [Green Version]
- Zhuravleva, O.I.; Sobolevskaya, M.P.; Leshchenko, E.V.; Kirichuk, N.N.; Denisenko, V.A.; Dmitrenok, P.S.; Dyshlovoy, S.A.; Zakharenko, A.M.; Kim, N.Y.; Afiyatullov, S.S. Meroterpenoids from the alga-derived fungi Penicillium thomii Maire and Penicillium lividum Westling. J. Nat. Prod. 2014, 77, 1390–1395. [Google Scholar] [CrossRef]
- Huang, G.L.; Zhou, X.M.; Bai, M.; Liu, Y.X.; Zhao, Y.L.; Luo, Y.P.; Niu, Y.Y.; Zheng, C.J.; Chen, G.Y. Dihydroisocoumarins from the mangrove-derived fungus Penicillium citrinum. Mar. Drugs 2016, 14, 177. [Google Scholar] [CrossRef] [Green Version]
- Arunpanichlert, J.; Rukachaisirikul, V.; Phongpaichit, S.; Supaphon, O.; Sakayaroj, J. Meroterpenoid, isocoumarin, and phenol derivatives from the seagrass-derived fungus Pestalotiopsis sp. PSU-ES 194. Tetrahedron 2015, 71, 882–888. [Google Scholar] [CrossRef]
- Liu, Z.M.; Liu, H.J.; Chen, Y.; She, Z.G. A new anti-inflammatory meroterpenoid from the fungus Aspergillus terreus H010. Nat. Prod. Res. 2018, 32, 2652–2656. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Quang, T.H.; Yoon, C.S.; Kim, H.J.; Sohn, J.H.; Oh, H. Furanoaustinol and 7-acetoxydehydroaustinol: New meroterpenoids from a marine-derived fungal strain Penicillium sp. SF-5497. J. Antibiot. 2018, 71, 557–563. [Google Scholar] [CrossRef] [PubMed]
- Bai, M.; Zheng, C.J.; Huang, G.L.; Mei, R.Q.; Wang, B.; Luo, Y.P.; Zheng, C.; Niu, Z.G.; Chen, G.Y. Bioactive meroterpenoids and isocoumarins from the mangrove derived fungus Penicillium sp. TGM112. J. Nat. Prod. 2019, 82, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.-Y.; You, M.J.; Oh, D.-C.; Oh, K.-B.; Shin, J. New meroterpenoids from a Penicillium sp. Fungus. Nat. Prod. Sci. 2018, 24, 253–258. [Google Scholar] [CrossRef]
- Wen, H.; Yang, X.; Liu, Q.; Li, S.; Li, Q.; Zang, Y.; Chen, C.; Wang, J.; Zhu, H.; Zhang, Y. Structurally diverse meroterpenoids from a marine-derived Aspergillus sp. fungus. J. Nat. Prod. 2020, 83, 99–104. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.P.; Wu, Y.F.; Yuan, B.C.; Liu, D.; Zhu, K.; Huang, J.; Proksch, P.; Lin, W.H. DMOA-based meroterpenoids with diverse scaffolds from the sponge-associated fungus Penicillium brasilianum. Tetrahedron 2019, 75, 2193–2205. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, X.-M.; Shang, Z.; Li, C.-S.; Ji, N.-Y.; Wang, B.-G. Meroterpenoid and diphenyl ether derivatives from Penicillium sp. MA-37, a fungus isolated from marine mangrove rhizospheric soil. J. Nat. Prod. 2012, 75, 1888–1895. [Google Scholar] [CrossRef]
- Li, J.; Yang, X.; Lin, Y.; Yuan, J.; Lu, Y.; Zhu, X.; Li, J.; Li, M.; Lin, Y.; He, J. Meroterpenes and azaphilones from marine mangrove endophytic fungus Penicillium 303#. Fitoterapia 2014, 97, 241–246. [Google Scholar] [CrossRef]
- Hoang, T.P.T.; Roullier, C.; Boumard, M.C.; du Pont, T.R.; Nazih, H.; Gallard, J.F.; Pouchus, Y.F.; Beniddir, M.A.; Grovel, O. Metabolomics-driven discovery of meroterpenoids from a mussel-derived Penicillium ubiquetum. J. Nat. Prod. 2018, 81, 2501–2511. [Google Scholar] [CrossRef]
- Zhang, J.P.; Yuan, B.C.; Liu, D.; Gao, S.; Proksch, P.; Lin, W.H. Brasilianoids A-F, new meroterpenoids from the sponge-associated fungus Penicillium brasilianum. Front. Chem. 2018, 6, 314. [Google Scholar] [CrossRef] [Green Version]
- Gu, B.B.; Wu, W.; Liu, L.Y.; Tang, J.; Zeng, Y.J.; Wang, S.P.; Sun, F.; Li, L.; Yang, F.; Lin, H.W. 3,5-Dimethylorsellinic acid derived meroterpenoids from Eupenicillium sp. 6A-9, a Fungus Isolated from the marine sponge Plakortis simplex. Eur. J. Org. Chem. 2018, 48–59. [Google Scholar] [CrossRef]
- Park, J.-S.; Quang, T.H.; Thi Thanh Ngan, N.; Sohn, J.H.; Oh, H. New preaustinoids from a marine-derived fungal strain Penicillium sp. SF-5497 and their inhibitory effects against PTP1B activity. J. Antibiot. 2019, 72, 629–633. [Google Scholar] [CrossRef] [PubMed]
- López-Gresa, M.P.; Cabedo, N.; González-Mas, M.C.; Ciavatta, M.L.; Avila, C.; Primo, J. Terretonins E and F, inhibitors of the mitochondrial respiratory chain from the marine-derived fungus Aspergillus insuetus. J. Nat. Prod. 2009, 72, 1348–1351. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, T.; Kurihara, Y.; Kanamoto, A.; Tomoda, H. Terretonin G, a new sesterterpenoid antibiotic from marine-derived Aspergillus sp. OPMF00272. J. Antibiot. 2014, 67, 593–595. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.L.; Li, X.M.; Li, X.; Yang, S.Q.; Wang, B.G. Structure, absolute configuration and biological evaluation of polyoxygenated meroterpenoids from the marine algal-derived Aspergillus terreus EN-539. Phytochem. Lett. 2019, 32, 138–142. [Google Scholar] [CrossRef]
- Hamed, A.; Abdel-Razek, A.S.; Omran, D.A.; El-Metwally, M.M.; El-Hosari, D.G.; Frese, M.; Soliman, H.S.M.; Sewald, N.; Shaaban, M. Terretonin O: A new meroterpenoid from Aspergillus terreus. Nat. Prod. Res. 2020, 34, 965–974. [Google Scholar] [CrossRef]
- Wu, C.-J.; Cui, X.; Xiong, B.; Yang, M.-S.; Zhang, Y.-X.; Liu, X.-M. Terretonin D1, a new meroterpenoid from marine-derived Aspergillus terreus ML-44. Nat. Prod. Res. 2019, 33, 2262–2265. [Google Scholar] [CrossRef]
- Cheng, Z.B.; Xu, W.; Wang, Y.Y.; Bai, S.Y.; Liu, L.J.; Luo, Z.H.; Yuan, W.J.; Li, Q. Two new meroterpenoids and two new monoterpenoids from the deep sea-derived fungus Penicillium sp. YPGA11. Fitoterapia 2019, 133, 120–124. [Google Scholar] [CrossRef]
- Xie, C.-L.; Xia, J.-M.; Lin, T.; Lin, Y.-J.; Lin, Y.-K.; Xia, M.-L.; Chen, H.-F.; Luo, Z.-H.; Shao, Z.-Z.; Yang, X.-W. Andrastone A From the deep-sea-derived fungus Penicillium allii-sativi acts as an inducer of caspase and RXRα-dependent apoptosis. Front. Chem. 2019, 7, 692. [Google Scholar] [CrossRef]
- Li, H.-L.; Xu, R.; Li, X.-M.; Yang, S.-Q.; Meng, L.-H.; Wang, B.-G. Simpterpenoid A, a meroterpenoid with a highly functionalized cyclohexadiene moiety featuring gem-propane-1,2-dione and methylformate groups, from the mangrove-derived Penicillium simplicissimum MA-332. Org. Lett. 2018, 20, 1465–1468. [Google Scholar] [CrossRef]
- Wang, Y.Z.; Qi, S.; Zhan, Y.; Zhang, N.W.; Wu, A.A.; Gui, F.; Guo, K.; Yang, Y.R.; Cao, S.G.; Hu, Z.Y. Aspertetranones A-D, putative meroterpenoids from the marine algal-associated fungus Aspergillus sp. ZL0-1b14. J. Nat. Prod. 2015, 78, 2405–2410. [Google Scholar] [CrossRef]
- Qi, C.X.; Qiao, Y.B.; Gao, W.X.; Liu, M.T.; Zhou, Q.; Chen, C.M.; Lai, Y.J.; Xue, Y.B.; Zhang, J.W.; Li, D.Y. New 3,5-dimethylorsellinic acid-based meroterpenoids with BACE1 and AchE inhibitory activities from Aspergillus terreus. Org. Biomol. Chem. 2018, 16, 9046–9052. [Google Scholar] [CrossRef]
- Eamvijarn, A.; Gomes, N.M.; Dethoup, T.; Buaruang, J.; Manoch, L.; Silva, A.; Pedro, M.; Marini, I.; Roussis, V.; Kijjoa, A. Bioactive meroditerpenes and indole alkaloids from the soil fungus Neosartorya fischeri (KUFC 6344), and the marine-derived fungi Neosartorya laciniosa (KUFC 7896) and Neosartorya tsunodae (KUFC 9213). Tetrahedron 2013, 69, 8583–8591. [Google Scholar] [CrossRef]
- Gomes, N.M.; Bessa, L.J.; Buttachon, S.; Costa, P.M.; Buaruang, J.; Dethoup, T.; Silva, A.M.S.; Kijjoa, A. Antibacterial and antibiofilm activities of tryptoquivalines and meroditerpenes isolated from the marine-derived fungi Neosartorya paulistensis, N. laciniosa, N. tsunodae, and the soil fungi N. fischeri and N. siamensis. Mar. Drugs 2014, 12, 822–839. [Google Scholar] [CrossRef] [PubMed]
- Zin, W.W.M.; Buttachon, S.; Buaruang, J.; Gales, L.; Pereira, J.A.; Pinto, M.M.M.; Silva, A.M.S.; Kijjoa, A. A new meroditerpene and a new tryptoquivaline analog from the algicolous fungus Neosartorya takakii KUFC 7898. Mar. Drugs 2015, 13, 3776–3790. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prata-Sena, M.; Ramos, A.A.; Buttachon, S.; Castro-Carvalho, B.; Marques, P.; Dethoup, T.; Kijjoa, A.; Rocha, E. Cytotoxic activity of secondary metabolites from marine-derived fungus Neosartorya siamensis in human cancer cells. Phytother. Res. 2016, 30, 1862–1871. [Google Scholar] [CrossRef]
- Hu, Y.; Yang, M.; Zhao, J.; Liao, Z.; Qi, J.; Wang, X.; Jiang, W.; Xia, X. A Meroterpenoid isolated from the fungus Aspergillus sp. Nat. Prod. Comm. 2019, 14, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Gao, S.-S.; Li, X.-M.; Williams, K.; Proksch, P.; Ji, N.-Y.; Wang, B.-G. Rhizovarins A-F, indole-diterpenes from the mangrove-derived endophytic fungus Mucor irregularis QEN-189. J. Nat. Prod. 2016, 79, 2066–2074. [Google Scholar] [CrossRef]
- Sun, K.; Li, Y.; Guo, L.; Wang, Y.; Liu, P.; Zhu, W. Indole diterpenoids and isocoumarin from the fungus, Aspergillus flavus, isolated from the prawn, Penaeus vannamei. Mar. Drugs 2014, 12, 3970–3981. [Google Scholar] [CrossRef] [Green Version]
- Chea, S.; Zhan, Y. Isolation of novel indole diterpenes and dihydrodibenzofuran from the marine fungus Aspergillus sp. J. Pharm. Bioresour. 2014, 11, 39–45. [Google Scholar] [CrossRef]
- Hu, X.-Y.; Meng, L.-H.; Li, X.; Yang, S.-Q.; Li, X.-M.; Wang, B.-G. Three new indole diterpenoids from the sea-anemone-derived fungus Penicillium sp. AS-79. Mar. Drugs 2017, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Xu, W.; Li, S.; Chen, M.; Cheng, Y.; Yuan, W.; Cheng, Z.; Li, Q. Penicindopene A, a new indole diterpene from the deep-sea fungus Penicillium sp. YPCMAC1. Nat. Prod. Res. 2019, 33, 2988–2994. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.-C.; Xu, L.-L.; Yang, R.-Y.; Yang, M.-Y.; Hu, L.-D.; Zhu, H.-J.; Cao, F. Anti-Vibrio indole-diterpenoids and C-25 epimeric steroids from the marine-derived fungus Penicillium janthinellum. Front. Chem. 2019, 7, 80. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L.-M.; Kong, F.-D.; Fan, P.; Ma, Q.-Y.; Xie, Q.-Y.; Li, J.-H.; Zheng, H.-Z.; Zheng, Z.-H.; Yuan, J.-Z.; Dai, H.-F. Indole-diterpenoids with protein tyrosine phosphatase inhibitory activities from the marine-derived fungus Penicillium sp. KFD28. J. Nat. Prod. 2019, 82, 2638–2644. [Google Scholar] [CrossRef] [PubMed]




























© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Demerdash, A.; Kumla, D.; Kijjoa, A. Chemical Diversity and Biological Activities of Meroterpenoids from Marine Derived-Fungi: A Comprehensive Update. Mar. Drugs 2020, 18, 317. https://doi.org/10.3390/md18060317
El-Demerdash A, Kumla D, Kijjoa A. Chemical Diversity and Biological Activities of Meroterpenoids from Marine Derived-Fungi: A Comprehensive Update. Marine Drugs. 2020; 18(6):317. https://doi.org/10.3390/md18060317
Chicago/Turabian StyleEl-Demerdash, Amr, Decha Kumla, and Anake Kijjoa. 2020. "Chemical Diversity and Biological Activities of Meroterpenoids from Marine Derived-Fungi: A Comprehensive Update" Marine Drugs 18, no. 6: 317. https://doi.org/10.3390/md18060317
APA StyleEl-Demerdash, A., Kumla, D., & Kijjoa, A. (2020). Chemical Diversity and Biological Activities of Meroterpenoids from Marine Derived-Fungi: A Comprehensive Update. Marine Drugs, 18(6), 317. https://doi.org/10.3390/md18060317