In Vitro and In Vivo Assessment of the Efficacy of Bromoageliferin, an Alkaloid Isolated from the Sponge Agelas dilatata, against Pseudomonas aeruginosa

 ,
,  ,
,  ,
,  ,
, 
Abstract
:1. Introduction
2. Results and Discussion
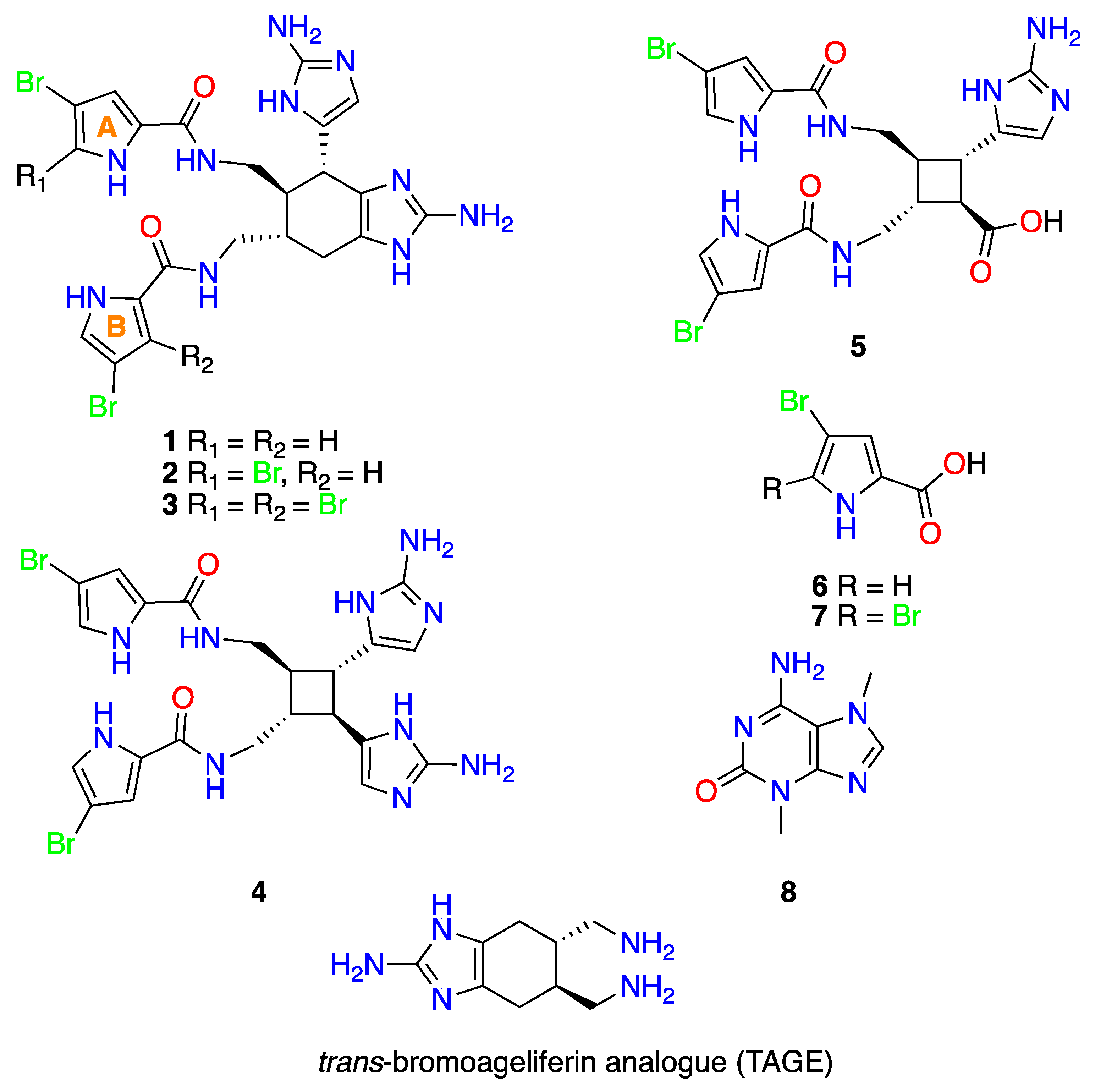
2.1. Extraction, Isolation and Structural Elucidation
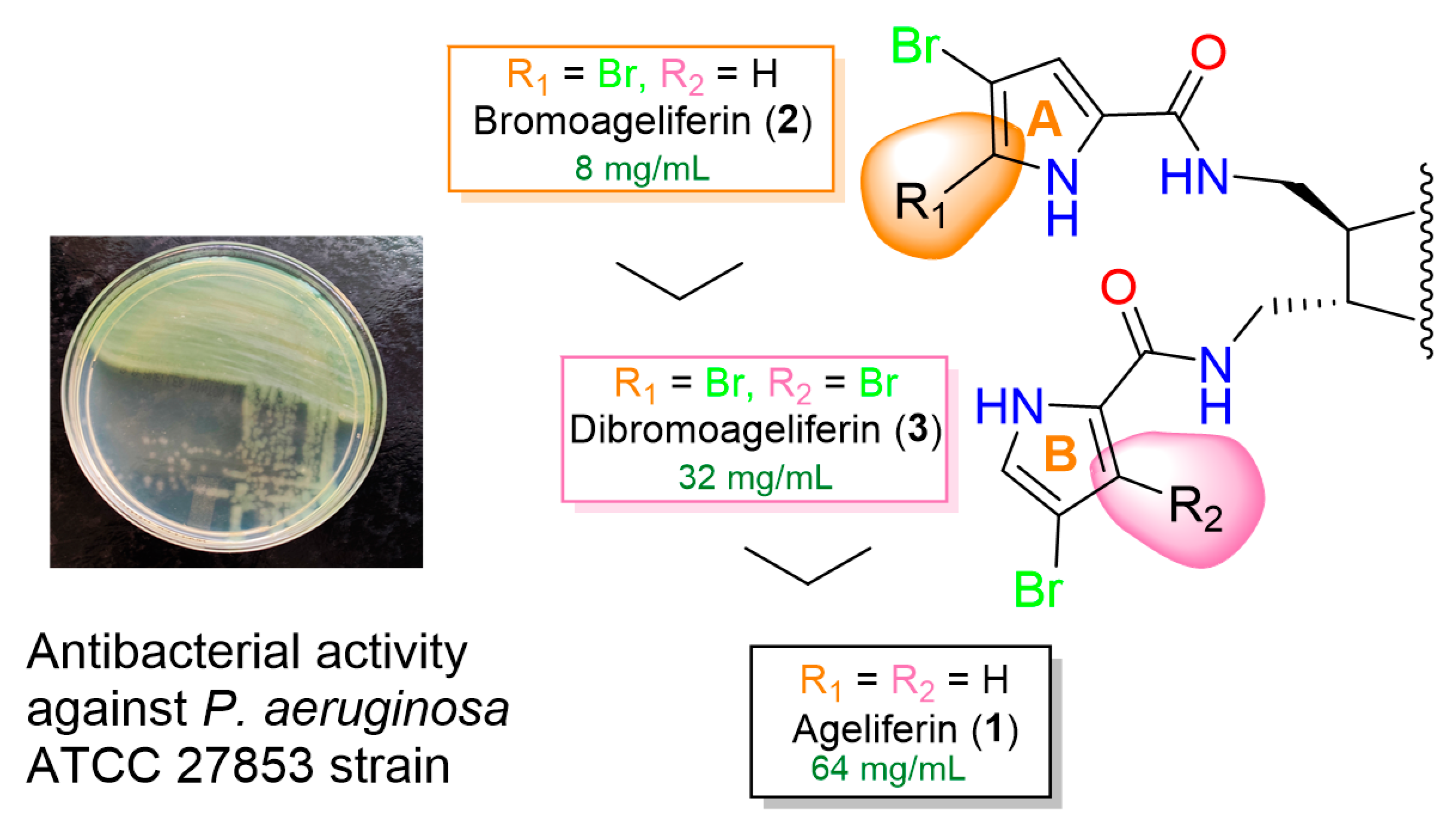
2.2. Antimicrobial Susceptibility Testing
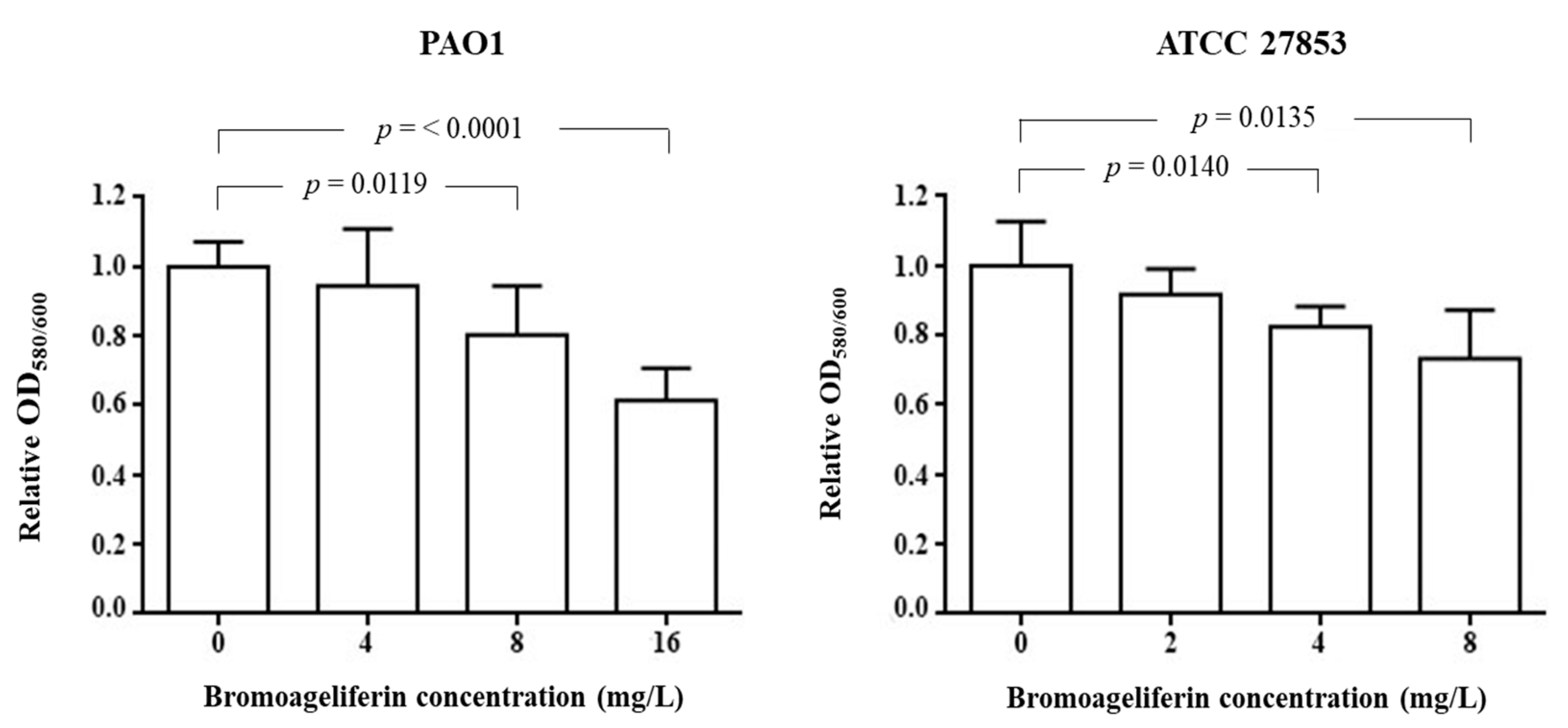
2.3. Analysis of Biofilm Biomass Inhibition
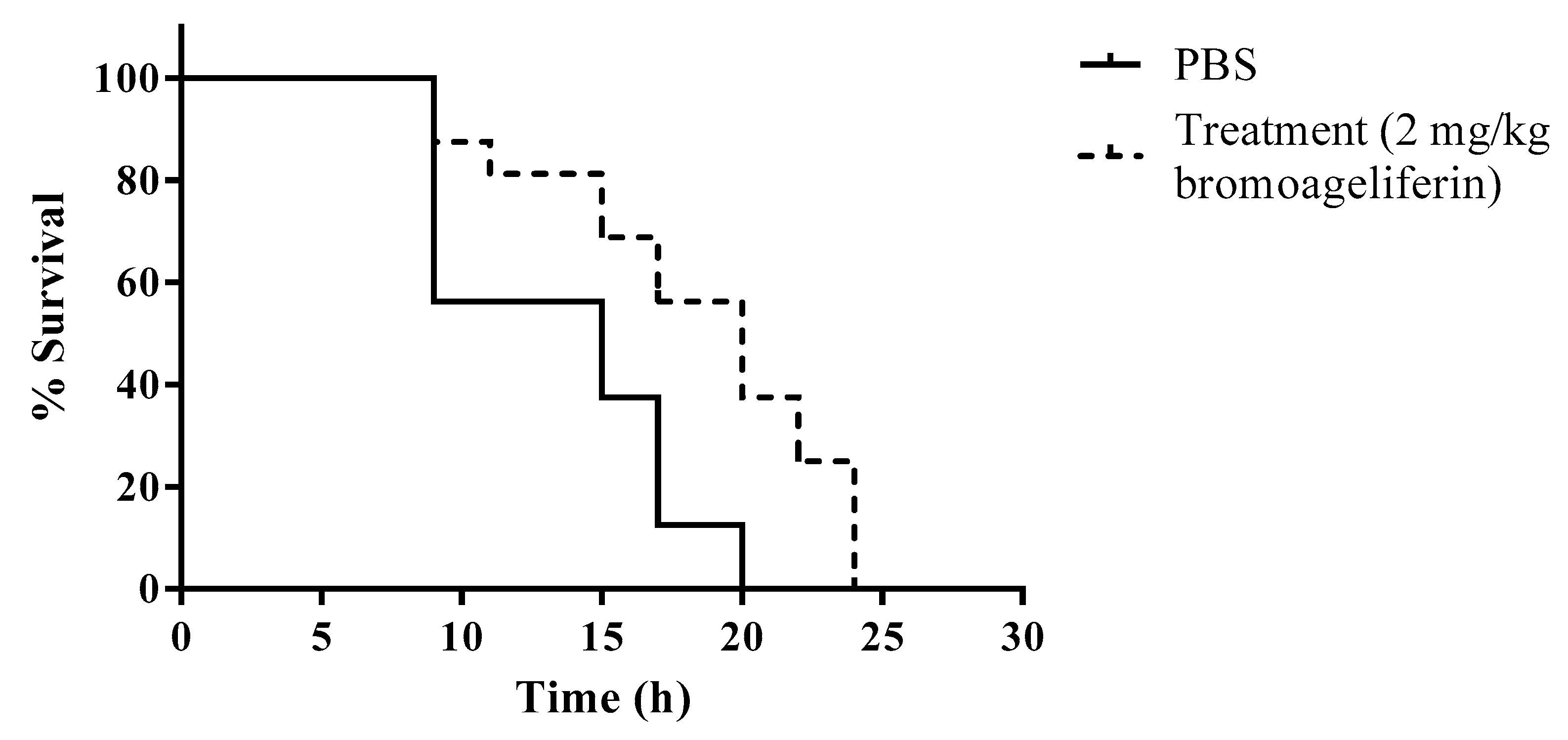
2.4. In Vivo Efficacy of Bromoageliferin against P. aeruginosa
2.5. Additional Reported Activities for 1–8
3. Materials and Methods
3.1. General Experimental Chemical Procedures
3.2. Sponge Collection
3.3. Extraction and Isolation
3.4. Structural Characterization
- Ageliferin (1). +15.1° (c 0.2, CH3OH); 1H and 13C NMR see SM; (−)-HRESIMS m/z 617.0380 [M−H]− (calcd. for C22H2379Br2N10O2, 617.0378).
- Bromoageliferin (2). +9.1° (c 0.2, CH3OH); 1H and 13C NMR see SM; (−)-HRESIMS m/z 694.9482 [M−H]− (calcd. for C22H2279Br3N10O2, 694.9483).
- Dibromoageliferin (3). +4.0° (c 0.2, CH3OH); 1H and 13C NMR see SM; (−)-HRESIMS m/z 772.8582 [M−H]− (calcd. for C22H2179Br4N10O2, 772.8588).
- Sceptrin (4). −13.8° (c 0.2, CH3OH); 1H and 13C NMR see SM; (+)-HRESIMS m/z 619.0531 [M−H]+ (calcd. for C22H2579Br2N10O2, 619.0523).
- Nakamuric acid (5). − 9.5° (c 0.2, CH3OH); 1H and 13C NMR see SM; (−)-HRESIMS m/z 579.9945 [M−H]− (calcd. for C20H2079Br2N7O4, 579.9949).
- 4-bromo-1H-pyrrole-2-carboxylic acid (6). 1H and 13C NMR see SM; (−)-HRESIMS m/z 187.9353 [M−H]− (calcd. for C5H379BrNO2, 187.9353).
- 4,5-dibromopyrrole-2- carboxylic acid (7). 1H and 13C NMR see SM; (−)-HRESIMS m/z 265.8455 [M−H]− (calcd. for C5H279Br2NO2, 265.8458).
- 3,7-dimethylisoguanine (8). 1H and 13C see SM; (+)−HRESIMS m/z 180.0881 [M+H]+ (calcd. for C7H10N5O, 180.0880).
3.5. Bacterial Strains and Culture Media
3.6. Microdilution Method: Minimum Inhibitory Concentration
3.7. Biofilm Inhibition Assay
3.8. Galleria Mellonella Treatment
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dolgin, E. Sequencing of superbugs seen as key to combating their spread. Nat. Med. 2010, 16, 1054. [Google Scholar] [CrossRef]
- Dadgostar, P. Antimicrobial resistance: Implications and costs. Infect. Drug Resist. 2019, 12, 3903–3910. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization (WHO). Antimicrobial Resistance-Global Report on Surveillance. 2014. Available online: http://apps.who.int/iris/bitstream/10665/112642/1/9789241564748_eng.pdf (accessed on 23 May 2014).
- Donlan, R.M.; Costerton, J.W. Biofilms: Survival mechanisms of clinically relevant microorganisms. Clin. Microbiol. Rev. 2002, 15, 167–193. [Google Scholar] [CrossRef] [Green Version]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [Green Version]
- Jiménez, C. Marine Natural Products in Medicinal Chemistry. ACS Med. Chem. Lett. 2018, 9, 959–961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindel, T. Chemistry and Biology of the Pyrrole–Imidazole Alkaloids. Alkaloids Chem. Biol. 2017, 77, 117–219. [Google Scholar] [CrossRef]
- Zhang, H.; Dong, M.; Chen, J.; Wang, H.; Tenney, K.; Crews, P. Bioactive secondary metabolites from the marine sponge genus Agelas. Mar. Drugs 2017, 15, 351. [Google Scholar] [CrossRef] [Green Version]
- Sharma, G.M.; Burkholder, P.R. Structure of dibromophakellin, a new bromine-containing alkaloid from the marine sponge Phakellia flabellata. J. Chem. Soc. D Chem. Commun. 1971, 151–152. [Google Scholar] [CrossRef]
- Rane, R.; Sahu, N.; Shah, C.; Karpoormath, R. Marine Bromopyrrole Alkaloids: Synthesis and Diverse Medicinal Applications. Curr. Top. Med. Chem. 2014, 14, 253–273. [Google Scholar] [CrossRef]
- Forte, B.; Malgesini, B.; Piutti, C.; Quartieri, F.; Scolaro, A.; Papeo, G. A submarine journey: The pyrrole-imidazole alkaloids. Mar. Drugs 2009, 7, 705–753. [Google Scholar] [CrossRef] [Green Version]
- Santafé, G.; Paz, V.; Rodríguez, J.; Jiménez, C. Novel cytotoxic oxygenated C29 sterols from the Colombian marine sponge Polymastia tenax. J. Nat. Prod. 2002, 65, 1161–1164. [Google Scholar] [CrossRef] [PubMed]
- Anta, C.; González, N.; Santafé, G.; Rodríguez, J.; Jiménez, C. New xenia diterpenoids from the Indonesian soft coral Xenia sp. J. Nat. Prod. 2002, 65, 766–768. [Google Scholar] [CrossRef] [PubMed]
- Pech-Puch, D.; Rodríguez, J.; Cautain, B.; Sandoval-Castro, C.A.; Jiménez, C. Cytotoxic Furanoditerpenes from the Sponge Spongia tubulifera Collected in the Mexican Caribbean. Mar. Drugs 2019, 17, 416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pech-Puch, D.; Pérez-Povedano, M.; Lenis-Rojas, O.A.; Rodríguez, J.; Jiménez, C. Marine Natural Products from the Yucatan Peninsula. Mar. Drugs 2020, 18, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hochmuth, T.; Niederkrüger, H.; Gernert, C.; Siegl, A.; Taudien, S.; Platzer, M.; Crews, P.; Hentschel, U.; Piel, J. Linking chemical and microbial diversity in marine sponges: Possible role for poribacteria as producers of methyl-branched fatty acids. ChemBioChem 2010, 11, 2572–2578. [Google Scholar] [CrossRef]
- Parra-Velandia, F.J.; Zea, S.; Van Soest, R.W.M. Reef sponges of the Genus Agelas (Porifera: Desmospongiae) from the Greater Caribbean. Zootaxa 2014, 3794, 301–343. [Google Scholar] [CrossRef]
- Shrivastava, S.R.; Shrivastava, P.S.; Ramasamy, J. World Health Organization releases global priority list of antibiotic-resistant bacteria to guide research, discovery, and development of new antibiotics. J. Med. Soc. 2018, 32, 76–77. [Google Scholar] [CrossRef]
- Jiménez, C.; Crews, P. Mauritamide A and accompanying oroidin alkaloids from the sponge Agelas mauritiana. Tetrahedron Lett. 1994, 35, 1375–1378. [Google Scholar] [CrossRef]
- Rinehart, K.L. Biologically Active Marine Natural Products. Pure Appl. Chem. 1989, 61, 525–528. [Google Scholar] [CrossRef]
- Kobayashi, J.; Tsuda, M.; Murayama, T.; Nakamura, H.; Ohizumi, Y.; Ishibashi, M.; Iwamura, M.; Ohta, T.; Nozoe, S. Ageliferins, potent actomyosin ATPase activators from the Okinawan marine sponge Agelas sp. Tetrahedron 1990, 46, 5579–5586. [Google Scholar] [CrossRef]
- Keifen, P.A.; Schwartz, R.E.; Koker, M.E.S.; Hughes, R.G.; Rittschof, D.; Rinehart, K.L. Bioactive Bromopyrrole Metabolites from the Caribbean Sponge Agelas conifera. J. Org. Chem. 1991, 56, 2965–2975. [Google Scholar] [CrossRef]
- Walker, R.P.; John Faulkner, D.; Van Engen, D.; Clardy, J. Sceptrin, an Antimicrobial Agent from the Sponge Agelas sceptrum. J. Am. Chem. Soc. 1981, 103, 6772–6773. [Google Scholar] [CrossRef]
- Eder, C.; Proksch, P.; Wray, V.; Van Soest, R.W.M.; Ferdinandus, E.; Pattisina, L.A. Sudarsono New bromopyrrole alkaloids from the Indopacific sponge Agelas nakamurai. J. Nat. Prod. 1999, 62, 1295–1297. [Google Scholar] [CrossRef] [PubMed]
- Chanas, B.; Pawlik, J.R.; Lindel, T.; Fenical, W. Chemical defense of the Caribbean sponge Agelas clathrodes (Schmidt). J. Exp. Mar. Bio. Ecol. 1997, 208, 185–196. [Google Scholar] [CrossRef]
- Forenza, S.; Minale, L.; Riccio, R.; Fattorusso, E. New bromo-pyrrole derivatives from the sponge Agelas oroides. J. Chem. Soc. D Chem. Commun. 1971, 1129–1130. [Google Scholar] [CrossRef]
- Cafieri, F.; Fattorusso, E.; Mangoni, A.; Taglialatela-scafati, O. Longamide and 3,7-dimethylisoguanine, Two Novel Alkaloids from the Marine Sponge Agelas longissima. Tetrahedron Lett. 1995, 36, 7893–7896. [Google Scholar] [CrossRef]
- Endo, T.; Tsuda, M.; Okada, T.; Mitsuhashi, S.; Shima, H.; Kikuchi, K.; Mikami, Y.; Fromont, J.; Kobayashi, J. Nagelamides A-H, new dimeric bromopyrrole alkaloids from marine sponge Agelas species. J. Nat. Prod. 2004, 67, 1262–1267. [Google Scholar] [CrossRef]
- Melander, R.J.; Liu, H.B.; Stephens, M.D.; Bewley, C.A.; Melander, C. Marine sponge alkaloids as a source of anti-bacterial adjuvants. Bioorganic Med. Chem. Lett. 2016, 26, 5863–5866. [Google Scholar] [CrossRef] [Green Version]
- Yamada, A.; Kitamura, H.; Yamaguchi, K.; Fukuzawa, S.; Kamijima, C.; Yazawa, K.; Kuramoto, M.; Wang, G.Y.S.; Fujitani, Y.; Uemura, D. Development of Chemical Substances Regulating Biofilm Formation. Bull. Chem. Soc. Jpn. 1997, 70, 3061–3069. [Google Scholar] [CrossRef]
- Mohammed, R.; Peng, J.; Kelly, M.; Hamann, M.T. Cyclic heptapeptides from the Jamaican sponge Stylissa caribica. J. Nat. Prod. 2006, 69, 1739–1744. [Google Scholar] [CrossRef] [Green Version]
- Chu, M.J.; Tang, X.L.; Qin, G.F.; Sun, Y.T.; Li, L.; de Voogd, N.J.; Li, P.L.; Li, G.Q. Pyrrole Derivatives and Diterpene Alkaloids from the South China Sea Sponge Agelas nakamurai. Chem. Biodivers. 2017, 14, e1600446. [Google Scholar] [CrossRef] [PubMed]
- Barrow, R.A.; Capon, R.J. Brominated pyrrole carboxylic acids from an australian marine sponge, Axinella sp. Nat. Prod. Lett. 1993, 1, 243–250. [Google Scholar] [CrossRef]
- König, G.M.; Wright, A.D.; Linden, A. Antiplasmodial and cytotoxic metabolites from the maltese sponge Agelas oroides. Planta Med. 1998, 64, 443–447. [Google Scholar] [CrossRef] [PubMed]
- Hassan, W.; Elkhayat, E.S.; Edrada, R.A.; Ebel, R.; Proksch, P. New Bromopyrrole Alkaloids from the Marine Sponges Axinella damicornis and Stylissa flabelliformis. Nat. Prod. Commun. 2007, 2, 1149–1154. [Google Scholar] [CrossRef]
- Abdjul, D.B.; Yamazaki, H.; Kanno, S.I.; Tomizawa, A.; Rotinsulu, H.; Wewengkang, D.S.; Sumilat, D.A.; Ukai, K.; Kapojos, M.M.; Namikoshi, M. An anti-mycobacterial bisfunctionalized sphingolipid and new bromopyrrole alkaloid from the Indonesian marine sponge Agelas sp. J. Nat. Med. 2017, 71, 531–536. [Google Scholar] [CrossRef]
- Tasdemir, D.; Topaloglu, B.; Perozzo, R.; Brun, R.; O’Neill, R.; Carballeira, N.M.; Zhang, X.; Tonge, P.J.; Linden, A.; Rüedi, P. Marine natural products from the Turkish sponge Agelas oroides that inhibit the enoyl reductases from Plasmodium falciparum, Mycobacterium tuberculosis and Escherichia coli. Bioorganic Med. Chem. 2007, 15, 6834–6845. [Google Scholar] [CrossRef]
- Pénez, N.; Culioli, G.; Pérez, T.; Briand, J.F.; Thomas, O.P.; Blache, Y. Antifouling properties of simple indole and purine alkaloids from the Mediterranean gorgonian Paramuricea clavata. J. Nat. Prod. 2011, 74, 2304–2308. [Google Scholar] [CrossRef]
- Huigens, R.W.; Richards, J.J.; Parise, G.; Ballard, T.E.; Zeng, W.; Deora, R.; Melander, C. Inhibition of Pseudomonas aeruginosa biofilm formation with bromoageliferin analogues. J. Am. Chem. Soc. 2007, 129, 6966–6967. [Google Scholar] [CrossRef]
- Huigens, R.W.; Ma, L.; Gambino, C.; Moeller, P.D.R.; Basso, A.; Cavanagh, J.; Wozniak, D.J.; Melander, C. Control of bacterial biofilms with marine alkaloid derivatives. Mol. Biosyst. 2008, 4, 614–621. [Google Scholar] [CrossRef] [PubMed]
- Yeagley, A.A.; Su, Z.; McCullough, K.D.; Worthington, R.J.; Melander, C. N-Substituted 2-aminoimidazole inhibitors of MRSA biofilm formation accessed through direct 1,3-bis(tert-butoxycarbonyl)guanidine cyclization. Org. Biomol. Chem. 2013, 11, 130–137. [Google Scholar] [CrossRef]
- Majik, M.S.; Tilvi, S.; Mascarenhas, S.; Kumar, V.; Chatterjee, A.; Banerjee, M. Construction and screening of 2-aryl benzimidazole library identifies a new antifouling and antifungal agent. RSC Adv. 2014, 4, 28259–28264. [Google Scholar] [CrossRef]
- Wright, C.J.; Wu, H.; Melander, R.J.; Melander, C.; Lamont, R.J. Disruption of heterotypic community development by Porphyromonas gingivalis with small molecule inhibitors. Mol. Oral Microbiol. 2014, 29, 185–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harding, C.R.; Schroeder, G.N.; Collins, J.W.; Frankel, G. Use of Galleria mellonella as a model organism to study Legionella pneumophila infection. J. Vis. Exp. 2013, e50964. [Google Scholar] [CrossRef] [Green Version]
- Champion, O.L.; Wagley, S.; Titball, R.W. Galleria mellonella as a model host for microbiological and toxin research. Virulence 2016, 7, 840–845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vassas, A.; Bourdy, G.; Paillard, J.J.; Lavayre, J.; Païs, M.; Quirion, J.C.; Debitus, C. Naturally occurring somatostatin and vasoactive intestinal peptide inhibitors. Isolation of alkaloids from two marine sponges. Planta Med. 1996, 62, 28–30. [Google Scholar] [CrossRef]
- Ortlepp, S.; Sjögren, M.; Dahlström, M.; Weber, H.; Ebel, R.; Edrada, R.A.; Thoms, C.; Schupp, P.; Bohlin, L.; Proksch, P. Antifouling activity of bromotyrosine-derived sponge metabolites and synthetic analogues. Mar. Biotechnol. 2007, 9, 776–785. [Google Scholar] [CrossRef]
- Bickmeyer, U.; Drechsler, C.; Köck, M.; Assmann, M. Brominated pyrrole alkaloids from marine Agelas sponges reduce depolarization-induced cellular calcium elevation. Toxicon 2004, 44, 45–51. [Google Scholar] [CrossRef]
- Assmann, M.; Lichte, E.; Pawlik, J.R.; Köck, M. Chemical defenses of the Caribbean sponges Agelas wiedenmayeri and Agelas conifera. Mar. Ecol. Prog. Ser. 2000, 207, 255–262. [Google Scholar] [CrossRef] [Green Version]
- Regalado, E.L.; Laguna, A.; Mendiola, J.; Thomas, O.P.; Nogueiras, C. Bromopyrrole alkaloids from the Caribbean sponge Agelas cerebrum. Quim. Nova 2011, 34, 289–291. [Google Scholar] [CrossRef] [Green Version]
- Peng, J.; Shen, X.; El Sayed, K.A.; Dunbar, D.C.; Perry, T.L.; Wilkins, S.P.; Hamann, M.T.; Bobzin, S.; Huesing, J.; Camp, R.; et al. Marine natural products as prototype agrochemical agents. J. Agric. Food Chem. 2003, 51, 2246–2252. [Google Scholar] [CrossRef] [Green Version]
- Cipres, A.; O’Malley, D.P.; Li, K.; Finlay, D.; Baran, P.S.; Vuori, K. Sceptrin, a marine natural compound, inhibits cell motility in a variety of cancer cell lines. ACS Chem. Biol. 2010, 5, 195–202. [Google Scholar] [CrossRef] [PubMed]
- Scala, F.; Fattorusso, E.; Menna, M.; Taglialatela-Scafati, O.; Tierney, M.; Kaiser, M.; Tasdemir, D. Bromopyrrole alkaloids as lead compounds against protozoan parasites. Mar. Drugs 2010, 8, 2162–2174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cafieri, F.; Carnuccio, R.; Fattorusso, E.; Taglialatela-Scafati, O.; Vallefuoco, T. Anti-histaminic activity of bromopyrrole alkaloids isolated from Caribbean Agelas sponges. Bioorganic Med. Chem. Lett. 1997, 7, 2283–2288. [Google Scholar] [CrossRef]
- Rosa, R.; Silva, W.; de Motta, G.E.; Rodríguez, A.D.; Morales, J.J.; Ortiz, M. Anti-Muscarinic Activity of a Family of C11N5 Compounds Isolated from Agelas Sponges. Experientia 1992, 48, 885–887. [Google Scholar] [CrossRef]
- Rodríguez, A.D.; Lear, M.J.; La Clair, J.J. Identification of the binding of sceptrin to MreB via a bidirectional affinity protocol. J. Am. Chem. Soc. 2008, 130, 7256–7258. [Google Scholar] [CrossRef] [PubMed]
- Tilvi, S.; Moriou, C.; Martin, M.T.; Gallard, J.F.; Sorres, J.; Patel, K.; Petek, S.; Debitus, C.; Ermolenko, L.; Al-Mourabit, A. Agelastatin E, agelastatin F, and benzosceptrin C from the marine sponge Agelas dendromorpha. J. Nat. Prod. 2010, 73, 720–723. [Google Scholar] [CrossRef]
- Muñoz, J.; Köck, M. Hybrid Pyrrole-Imidazole Alkaloids from the Sponge Agelas sceptrum. J. Nat. Prod. 2016, 79, 434–437. [Google Scholar] [CrossRef]
- Lindel, T.; Hoffmann, H.; Hochgürtel, M.; Pawlik, J.R. Structure-activity relationship of inhibition of fish feeding by sponge- derived and synthetic pyrrole-imidazole alkaloids. J. Chem. Ecol. 2000, 26, 1477–1496. [Google Scholar] [CrossRef]
- Bickmeyer, U.; Assmann, M.; Köck, M.; Schütt, C. A secondary metabolite, 4,5-dibromopyrrole-2-carboxylic acid, from marine sponges of the genus Agelas alters cellular calcium signals. Environ. Toxicol. Pharmacol. 2005, 19, 423–427. [Google Scholar] [CrossRef] [Green Version]
- Gunasekera, S.P.; Cranick, S.; Longley, R.E. Immunosuppressive compounds from a deep water marine sponge, Agelas flabelliformis. J. Nat. Prod. 1989, 52, 757–761. [Google Scholar] [CrossRef]
- Ponasik, J.A.; Conova, S.; Kinghorn, D.; Kinney, W.A.; Rittschof, D.; Ganem, B. Pseudoceratidine, a marine natural product with antifouling activity: Synthetic and biological studies. Tetrahedron 1998, 54, 6977–6986. [Google Scholar] [CrossRef]
- Orhan, I.E.; Ozcelik, B.; Konuklugil, B.; Putz, A.; Kaban, U.G.; Proksch, P. Bioactivity Screening of the Selected Turkish Marine Sponges and Three Compounds from Agelas oroides. Rec. Nat. Prod. 2012, 6, 356–367. [Google Scholar]
- Bou, G.; Oliver, A.; Martinez-Beltran, J. OXA-24, a novel class D beta-lactamase with carbapenemase activity in an Acinetobacter baumannii clinical strain. Antimicrob. Agents Chemother. 2000, 44, 1556–1561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gato, E.; Álvarez-Fraga, L.; Vallejo, J.A.; Rumbo-Feal, S.; Martínez-Guitián, M.; Beceiro, A.; Poza, M.; Bou, G.; Pérez, A. Draft Genome Sequences of Two Epidemic OXA-48-Producing Klebsiella pneumoniae Clinical Strains Isolated during a Large Outbreak in Spain. Genome Announc. 2018, 6, e00026-18. [Google Scholar] [CrossRef] [Green Version]
- Perez, A.; Gato, E.; Perez-Llarena, J.; Fernandez-Cuenca, F.; Gude, M.J.; Oviano, M.; Pachon, M.E.; Garnacho, J.; Gonzalez, V.; Pascual, A.; et al. High incidence of MDR and XDR Pseudomonas aeruginosa isolates obtained from patients with ventilator-associated pneumonia in Greece, Italy and Spain as part of the MagicBullet clinical trial. J. Antimicrob. Chemother. 2019, 74, 1244–1252. [Google Scholar] [CrossRef]
- Weinstein, M.P. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria that Grow Aerobically; National Committee for Clinical Laboratory Standards: Wayne, PA, USA, 2018. [Google Scholar]
- Martínez-Guitián, M.; Vázquez-Ucha, J.C.; Álvarez-Fraga, L.; Conde-Pérez, K.; Bou, G.; Poza, M.; Beceiro, A. Antisense inhibition of lpxB gene expression in Acinetobacter baumannii by peptide-PNA conjugates and synergy with colistin. J. Antimicrob. Chemother. 2020, 75, 51–59. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Acinetobacter baumannii | Klebsiella pneumoniae | Pseudomonas aeruginosa | |||
|---|---|---|---|---|---|---|
| ATCC 17978 | RYC 52763/97 | ATCC 700603 | KP 1803 | ATCC 27853 | PAO1 | |
| Ageliferin (1) | ≥128 | ≥128 | 64 | 64 | 64 | 64 |
| Bromoageliferin (2) | ≥128 | ≥128 | ≥128 | ≥128 | 8 | 32 |
| Dibromoageliferin (3) | ≥128 | 64 | 64 | 64 | 32 | 32 |
| Sceptrin (4) | ≥128 | ≥128 | 64 | 64 | 128 | 128 |
| Nakamuric acid (5) | ≥128 | ≥128 | ≥128 | ≥128 | ≥128 | ≥128 |
| 4-Bromo-1H-pyrrole-2-carboxylic acid (6) | ≥128 | ≥128 | ≥128 | ≥128 | 64 | ≥128 |
| 4,5-Dibromopyrrole-2- carboxylic acid (7) | ≥128 | ≥128 | 64 | ≥128 | 64 | ≥128 |
| 3,7-Dimethylisoguanine (8) | ≥128 | ≥128 | 64 | 64 | 64 | 128 |
| Imipenem (control) | 0.5 | 16 | 0.25 | 2 | 2 | 2 |
| Bacterial Strain | Description | Source/References |
|---|---|---|
| A. baumannii | ||
| ATCC 17978 | Reference strain, completely sequenced. | ATCC a |
| RYC 52763/97 | Clinical isolate from respiratory tract. | Outbreak in Ramón y Cajal Hospital, Madrid, Spain [64] |
| K. pneumoniae | ||
| ATCC 700603 | Reference strain, completely sequenced. | ATCC |
| KP 1803 | Clinical isolate from urinary tract, completely sequenced. | Outbreak in A Coruña Hospital, Spain [65] |
| P. aeruginosa | ||
| ATCC 27853 | Reference strain, completely sequenced. | ATCC |
| PAO1 | Reference strain, completely sequenced. | ATCC |
| 29-200 SV | Clinical isolate from respiratory tract. | MagicBullet clinical trial [66] |
| 30-127 VI | Clinical isolate from respiratory tract. | MagicBullet clinical trial [66] |
| 30-223 SV | Clinical isolate from respiratory tract. | MagicBullet clinical trial [66] |
| 30-230 SV | Clinical isolate from respiratory tract. | MagicBullet clinical trial [66] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pech-Puch, D.; Pérez-Povedano, M.; Martinez-Guitian, M.; Lasarte-Monterrubio, C.; Vázquez-Ucha, J.C.; Bou, G.; Rodríguez, J.; Beceiro, A.; Jimenez, C. In Vitro and In Vivo Assessment of the Efficacy of Bromoageliferin, an Alkaloid Isolated from the Sponge Agelas dilatata, against Pseudomonas aeruginosa. Mar. Drugs 2020, 18, 326. https://doi.org/10.3390/md18060326
Pech-Puch D, Pérez-Povedano M, Martinez-Guitian M, Lasarte-Monterrubio C, Vázquez-Ucha JC, Bou G, Rodríguez J, Beceiro A, Jimenez C. In Vitro and In Vivo Assessment of the Efficacy of Bromoageliferin, an Alkaloid Isolated from the Sponge Agelas dilatata, against Pseudomonas aeruginosa. Marine Drugs. 2020; 18(6):326. https://doi.org/10.3390/md18060326
Chicago/Turabian StylePech-Puch, Dawrin, Mar Pérez-Povedano, Marta Martinez-Guitian, Cristina Lasarte-Monterrubio, Juan Carlos Vázquez-Ucha, Germán Bou, Jaime Rodríguez, Alejandro Beceiro, and Carlos Jimenez. 2020. "In Vitro and In Vivo Assessment of the Efficacy of Bromoageliferin, an Alkaloid Isolated from the Sponge Agelas dilatata, against Pseudomonas aeruginosa" Marine Drugs 18, no. 6: 326. https://doi.org/10.3390/md18060326
APA StylePech-Puch, D., Pérez-Povedano, M., Martinez-Guitian, M., Lasarte-Monterrubio, C., Vázquez-Ucha, J. C., Bou, G., Rodríguez, J., Beceiro, A., & Jimenez, C. (2020). In Vitro and In Vivo Assessment of the Efficacy of Bromoageliferin, an Alkaloid Isolated from the Sponge Agelas dilatata, against Pseudomonas aeruginosa. Marine Drugs, 18(6), 326. https://doi.org/10.3390/md18060326