Anticancer Compounds Derived from Marine Diatoms



Abstract
:1. Introduction
2. Characteristics
3. Anticancer Compounds from Diatoms
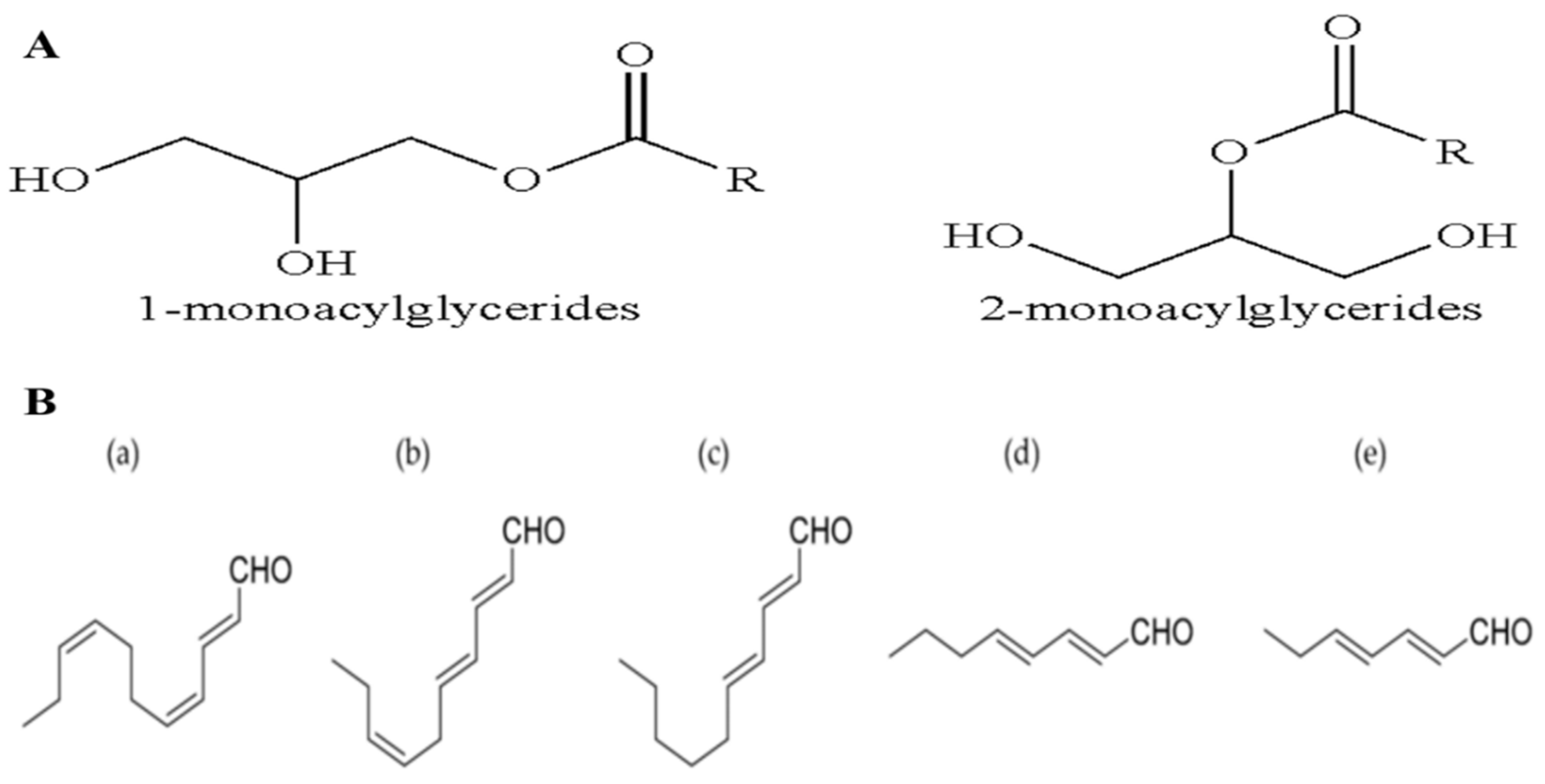
3.1. Monoacylglycerides (MAGs)
3.2. Oxylipins (OXLs)
3.3. Chrysolaminaran Polysaccharide
3.4. Fucoxanthin
3.5. Fatty Alcohol Ester (Nonyl 8-Acetoxy-6-Methyloctanoate, NAMO)
3.6. Adenosine and the Metabolites
3.7. Stigmasterol
3.8. Marennine
3.9. Haslene (Hasla-6(17),9,13,23-Tetraene) Lipid
3.10. Diatom Extracts
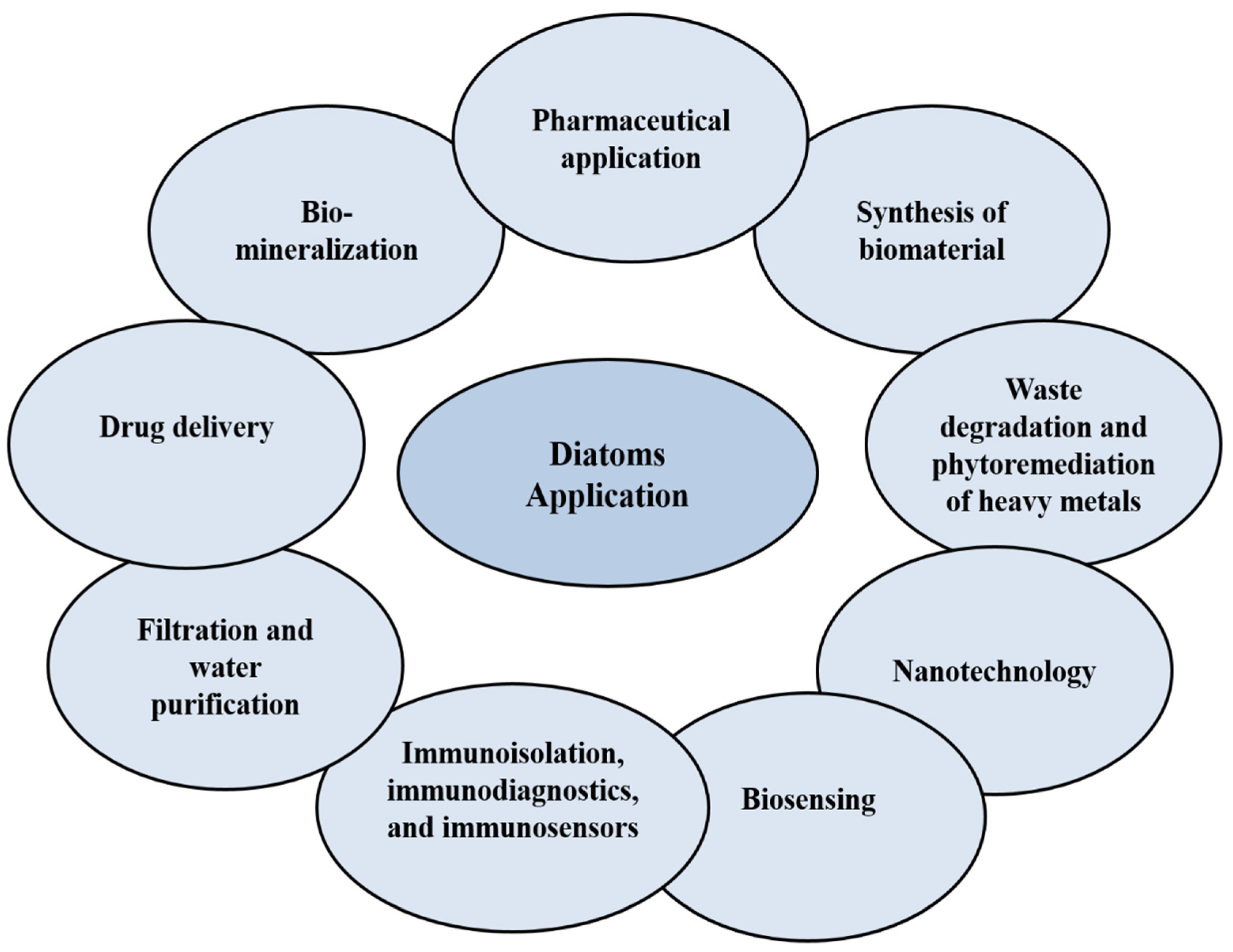
4. Diatoms for Drug Delivery Systems
4.1. Diatom-Based Nanoparticles (DNPs)
4.2. Diatoms-based Antibody
4.3. Diatoms-Based Vitamin B12
5. Conclusions and Future Work
Funding
Acknowledgments
Conflicts of Interest
References
- Cotas, J.; Marques, V.; Afonso, M.B.; Rodrigues, C.M.P.; Pereira, L. Antitumour potential of Gigartina pistillata Carrageenans against colorectal cancer stem cell-enriched tumourspheres. Mar. Drugs 2020, 18, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez Andrade, K.; Lauritano, C.; Romano, G.; Ianora, A. Marine microalgae with anti-cancer properties. Mar. Drugs 2018, 16, 165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samarakoon, K.W.; Ko, J.Y.; Lee, J.H.; Kwon, O.N.; Kim, S.W.; Jeon, Y.J. Apoptotic anticancer activity of a novel fatty alcohol ester isolated from cultured marine diatom, Phaeodactylum tricornutum. J. Funct. Foods 2014, 6, 231–240. [Google Scholar] [CrossRef]
- El-hack, M.E.A.; Abdelnour, S.; Alagawany, M.; Abdo, M.; Sakr, M.A.; Khafaga, A.F.; Mahgoub, A.; Elnesr, S.S.; Gebriel, M.G. Microalgae in modern cancer therapy: Current knowledge. Biomed. Pharmacother. 2019, 111, 42–50. [Google Scholar] [CrossRef]
- Folmer, F.; Jaspars, M.; Dicato, M.; Diederich, M. Photosynthetic marine organisms as a source of anticancer compounds. Phytochem. Rev. 2010, 9, 557–579. [Google Scholar] [CrossRef]
- Smyrniotopoulos, V.; Tomaz, A.C.D.A.; Souza, M.D.F.V.D.; Cunha, E.V.L.D.; Kiss, R.; Mathieu, V.; Ioannou, E.; Roussis, V. Halogenated diterpenes with In vitro antitumor activity from the red alga Sphaerococcus coronopifolius. Mar. Drugs 2020, 18, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dias, D.A.; Urban, S.; Roessner, U. A historical overview of natural products in drug discovery. Metabolites 2012, 2, 303–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, S.Y.; Liao, L.; Park, S.H.; Kim, W.K.; Shin, J.; Lee, S.K. Antitumor activity of Asperphenin A, a lipopeptidyl benzophenone from marine-derived Aspergillus sp. fungus, by inhibiting tubulin polymerization in colon cancer cells. Mar. Drugs 2020, 18, 110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lauritano, C.; Andersen, J.H.; Hansen, E.; Albrigtsen, M.; Escalera, L.; Esposito, F.; Helland, K.; Hanssen, K.Ø.; Romano, G.; Ianora, A. Bioactivity screening of microalgae for antioxidant, anti-inflammatory, anticancer, anti-diabetes, and antibacterial activities. Front. Mar. Sci. 2016, 3, 68. [Google Scholar] [CrossRef] [Green Version]
- Delasoie, J.; Rossier, J.; Haeni, L.; Rothen-Rutishauser, B.; Zobi, F. Slow-targeted release of a ruthenium anticancer agent from vitamin B12 functionalized marine diatom microalgae. Dalt. Trans. 2018, 47, 17221–17232. [Google Scholar] [CrossRef] [PubMed]
- Sansone, C.; Braca, A.; Ercolesi, E.; Romano, G.; Palumbo, A.; Casotti, R.; Francone, M.; Ianora, A. Diatom-derived polyunsaturated aldehydes activate cell death in human cancer cell lines but not normal cells. PLoS ONE 2014, 9, e101220. [Google Scholar] [CrossRef] [Green Version]
- Xia, S.; Gao, B.; Li, A.; Xiong, J.; Ao, Z.; Zhang, C. Preliminary characterization, antioxidant properties and production of chrysolaminarin from marine diatom Odontella aurita. Mar. Drugs 2014, 12, 4883–4897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dolatabadi, J.E.N.; de la Guardia, M. Applications of diatoms and silica nanotechnology in biosensing, drug and gene delivery, and formation of complex metal nanostructures. TrAC Trends Anal. Chem. 2011, 30, 1538–1548. [Google Scholar] [CrossRef]
- Bozarth, A.; Maier, U.G.; Zauner, S. Diatoms in biotechnology: Modern tools and applications. Appl. Microbiol. Biotechnol. 2009, 82, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Kuczynska, P.; Jemiola-Rzeminska, M.; Strzalka, K. Photosynthetic pigments in diatoms. Mar. Drugs 2015, 13, 5847–5881. [Google Scholar] [CrossRef] [PubMed]
- Jamali, A.A.; Akbari, F.; Ghorakhlu, M.M.; De Guardia, M.; Khosroushahi, A.Y. Applications of diatoms as potential microalgae in nanobiotechnology. BioImpacts 2012, 2, 83–89. [Google Scholar] [PubMed]
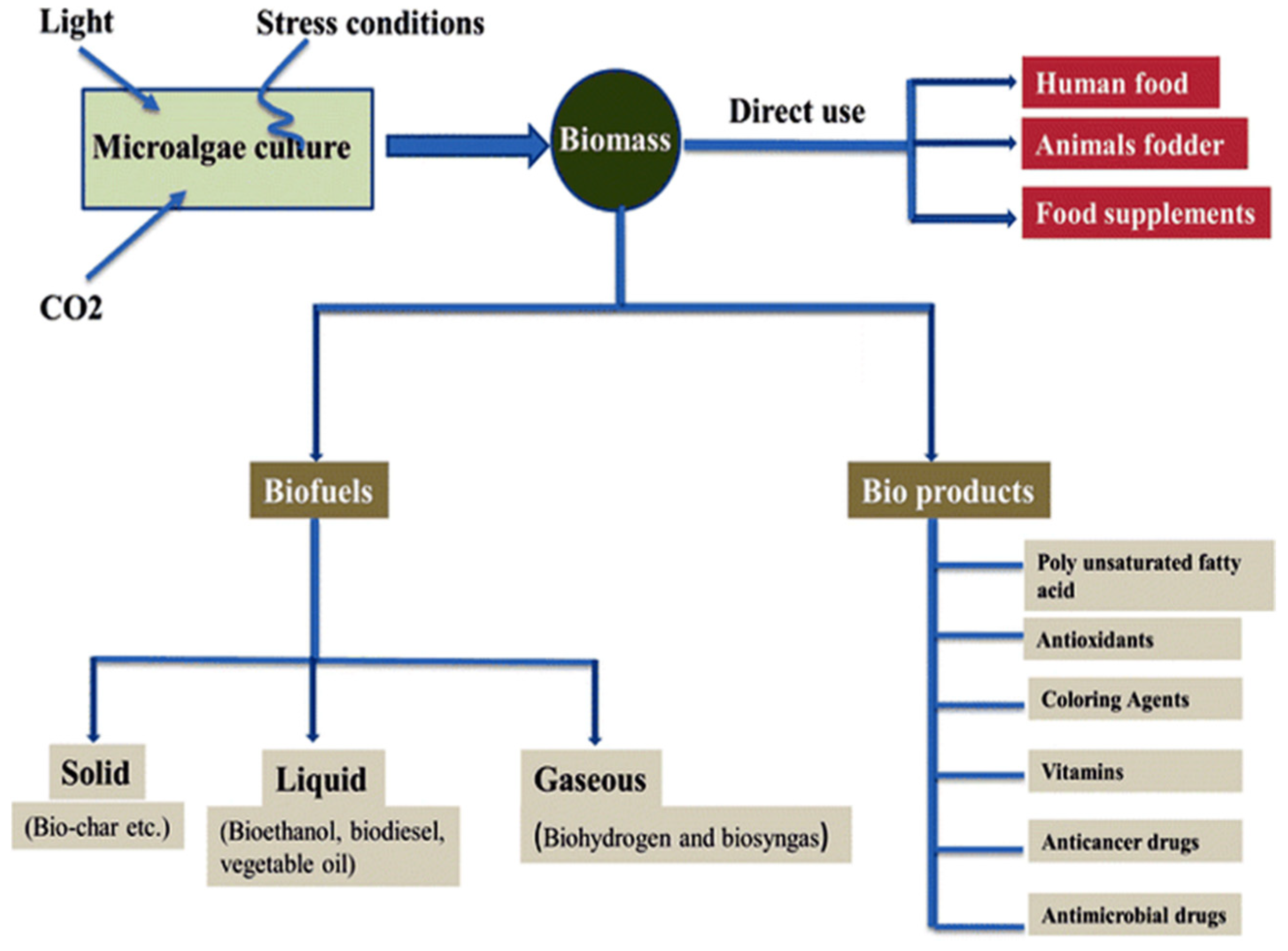
- Khan, M.I.; Shin, J.H.; Kim, J.D. The promising future of microalgae: Current status, challenges, and optimization of a sustainable and renewable industry for biofuels, feed, and other products. Microb. Cell Fact. 2018, 17, 36. [Google Scholar] [CrossRef]
- Kusaikin, M.I.; Ermakova, S.P.; Shevchenko, N.M.; Isakov, V.V.; Gorshkov, A.G.; Vereshchagin, A.L.; Grachev, M.A.; Zvyagintseva, T.N. Structural characteristics and antitumor activity of a new chrysolaminaran from the diatom alga Synedra acus. Chem. Nat. Compd. 2010, 46, 1–4. [Google Scholar] [CrossRef]
- Lopez, P.J.; Descle’s, J.; Allen, A.E.; Bowler, C. Prospects in diatom research. Curr. Opin. Biotechnol. 2005, 16, 180–186. [Google Scholar] [CrossRef]
- Medarević, D.P.; Lošić, D.; Ibrić, S.R. Diatoms—Nature materials with great potential for bioapplications. Hem. Ind. 2015, 70, 613–627. [Google Scholar] [CrossRef] [Green Version]
- Kuppusamy, P.; Soundharrajan, I.; Srigopalram, S.; Yusoff, M.M.; Maniam, G.P.; Govindan, N.; Choi, K.C. Potential pharmaceutical and biomedical applications of diatoms microalgae—An overview. Indian J. Geo. Mar. Sci. 2017, 46, 663–667. [Google Scholar]
- Rasmussen, R.S.; Morrissey, M.T. Marine biotechnology for production of food ingredients. Adv. Food Nutr. Res. 2007, 52, 237–292. [Google Scholar]
- Cheng, J.; Li, Y.; Liang, J.; Gao, Y.; Wang, P. Morphological variability and genetic diversity in five species of Skeletonema (Bacillariophyta). Prog. Nat. Sci. 2008, 18, 1345–1355. [Google Scholar] [CrossRef]
- Borowitzka, M.A. Biology of microalgae. In Microalgae in Health and Disease Prevention; Academic Press: Cambridge, MA, USA, 2018; pp. 23–72. [Google Scholar]
- Palenzuela, J.M.T.; Vilas, L.G.; Bellas, F.M.; Garet, E.; González-Fernández, Á.; Spyrakos, E. Pseudo-nitzschia blooms in a coastal upwelling system: Remote sensing detection, toxicity and environmental variables. Water 2019, 11, 1954. [Google Scholar] [CrossRef] [Green Version]
- Narang, A.S.; Desai, D.S. Anticancer Drug Development Unique Aspects of Pharmaceutical Development; Springer: New York, NY, USA, 2009; pp. 49–92. [Google Scholar]
- Hussein, H.A.; Mohamad, H.; Ghazaly, M.M.; Laith, A.A.; Abdullah, M.A. Cytotoxic effects of Tetraselmis suecica chloroform extracts with silver nanoparticle co-application on MCF-7, 4 T1, and Vero cell lines. J. Appl. Phycol. 2020, 32, 127–143. [Google Scholar] [CrossRef]
- Miceli, M.; Cutignano, A.; Conte, M.; Ummarino, R.; Romanelli, A.; Ruvo, M.; Leone, M.; Mercurio, F.A.; Doti, N.; Manzo, E.; et al. Monoacylglycerides from the diatom Skeletonema marinoi induce selective cell death in cancer cells. Mar. Drugs 2019, 17, 625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khaddaj-mallat, R.; Morin, C.; Rousseau, E. Novel n-3 PUFA monoacylglycerides of pharmacological and medicinal interest: Anti-inflammatory and anti-proliferative effects. Eur. J. Pharmacol. 2016, 792, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Morin, C.; Rodr, E.; Blier, P.U.; Fortin, S. Potential Application of Eicosapentaenoic acid monoacylglyceride in the management of colorectal cancer. Mar. Drugs 2017, 15, 283. [Google Scholar] [CrossRef] [PubMed]
- Alsufyani, T. Metabolite Profiling of the Chemosphere of the Macroalga Ulva (Ulvales, Chlorophyta) and its Associated Bacteria. Ph.D. Thesis, Thuringia University and State Library, Jena, Germany, June 2014. [Google Scholar]
- Yi, Z. Biotechnological Approaches to Enhance Fucoxanthin Production in a Model Diatom Phaeodactylum tricornutum. Ph.D. Thesis, Faculty of Life and Environmental Sciences, University of Iceland, Reykjavík, Iceland, November 2018. [Google Scholar]
- Guschina, I.A.; Harwood, J.L. Lipids and lipid metabolism in eukaryotic algae. Prog. Lipid Res. 2006, 45, 160–186. [Google Scholar] [CrossRef] [PubMed]
- Sarpal, S.; Sharma, K.; Scott, J.; Kumar, R.; Sugmaran, V.; Chopra, A.; Bansal, V.; Rajagopalan, N.K. Compositional analyses of oil extracts of microalgae biomasses by NMR and chromatographic techniques. J. Anal. Bioanal. Sep. Tech. 2016, 1, 17–41. [Google Scholar]
- Miralto, A.; Barone, G.; Romano, G.; Poulet, S.A.; Ianora, A.; Russo, G.L.; Buttino, I.; Mazzarella, G.; Laabir, M.; Cabrini, M.; et al. The insidious effect of diatoms on copepod reproduction. Nature 1999, 402, 173–176. [Google Scholar] [CrossRef]
- Andrianasolo, E.H.; Haramaty, L.; Vardi, A.; White, E.; Lutz, R.; Falkowski, P. Apoptosis—Inducing galactolipids from a cultured marine diatom Phaeodactylum tricornutum. J. Nat. Prod. 2008, 71, 1197–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neumann, U.; Derwenskus, F.; Flister, V.F.; Schmid-staiger, U.; Hirth, T.; Bischo, S.C. Fucoxanthin, A carotenoid derived from Phaeodactylum tricornutum exerts antiproliferative and antioxidant activities in vitro. Antioxidants 2019, 8, 183. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.S.; Li, X.F.; Kang, K.H.; Ryu, B.; Kim, S.K. Stigmasterol isolated from marine microalgae Navicula incerta induces apoptosis in human hepatoma HepG2 cells. BMB Rep. 2014, 47, 433–438. [Google Scholar] [CrossRef] [Green Version]
- Rowland, S.J.; Belt, S.T.; Wraige, E.J.; Masse, G.; Roussakis, C.; Robert, J. Effects of temperature on polyunsaturation in cytostatic lipids of Haslea ostrearia. Phytochemistry 2001, 56, 597–602. [Google Scholar] [CrossRef]
- Nappo, M.; Berkov, S.; Massucco, C.; Di Maria, V.; Bastida, J.; Codina, C.; Avila, C.; Messina, P.; Zupo, V.; Zupo, S. Apoptotic activity of the marine diatom Cocconeis scutellum and eicosapentaenoic acid in BT20 cells. Pharm. Biol. 2012, 50, 529–535. [Google Scholar] [CrossRef] [PubMed]
- Nigjeh, S.E.; Yusoff, F.; Alitheen, N.B.M.; Rasoli, M.; Keong, Y.S.; Omar, A.R.B. Cytotoxic effect of ethanol extract of microalga, Chaetoceros calcitrans, and its mechanisms in inducing apoptosis in human breast cancer cell line. Biomed Res. Int. 2013, 2013, 8. [Google Scholar]
- Goh, S.H.; Alitheen, N.B.M.; Yusoff, F.M.; Yap, S.K.; Loh, S.P. Crude ethyl acetate extract of marine microalga, Chaetoceros calcitrans, induces apoptosis in MDA-MB-231 breast cancer cells. Pharmacogn. Mag. 2014, 10, 1–8. [Google Scholar]
- Dembitsky, V.M.; Maoka, T. Allenic and cumulenic lipids. Prog. Lipid Res. 2007, 46, 328–375. [Google Scholar] [CrossRef]
- Peng, J.; Yuan, J.; Wu, C.; Wang, J. Fucoxanthin, a marine carotenoid present in brown seaweeds and diatoms: Metabolism and bioactivities relevant to human health. Mar. Drugs 2011, 9, 1806–1828. [Google Scholar] [CrossRef]
- Xia, S.; Wang, K.; Wan, L.; Li, A.; Hu, Q.; Zhang, C. Production, characterization, and antioxidant activity of fucoxanthin from the marine diatom Odontella aurita. Mar. Drugs 2013, 11, 2667–2681. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.R.; Hosokawa, M.; Miyashita, K. Fucoxanthin: A marine carotenoid exerting anti-cancer effects by affecting multiple mechanisms. Mar. Drugs 2013, 11, 5130–5147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Foo, S.C.; Yusoff, F.M.; Ismail, M.; Basri, M.; Chan, K.W.; Khong, N.M.H.; Yau, S.K. Production of fucoxanthin-rich fraction (FxRF) from a diatom, Chaetoceros calcitrans (Paulsen) Takano 1968. Algal Res. 2015, 12, 26–32. [Google Scholar] [CrossRef]
- Moreau, D.; Tomasoni, C.; Jacquot, C.; Kaas, R.; Le, R.; Cadoret, J.; Muller-feuga, A.; Kontiza, I.; Vagias, C.; Roussis, V.; et al. Cultivated microalgae and the carotenoid fucoxanthin from Odontella aurita as potent anti-proliferative agents in bronchopulmonary and epithelial cell lines. Environ. Toxicol. Pharmacol. 2006, 22, 97–103. [Google Scholar] [CrossRef]
- Rengarajan, T.; Rajendran, P.; Nandakumar, N.; Balasubramanian, M.P.; Nishigaki, I. Cancer preventive efficacy of marine carotenoid fucoxanthin: Cell cycle arrest and apoptosis. Nutrients 2013, 5, 4978–4989. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Wang, J.; Wang, W.; Zhang, H.; Zhang, X.; Han, C. The chemical constituents and pharmacological actions of Cordyceps sinensis. Evidence-based complement. Altern. Med. 2015, 2015, 1–15. [Google Scholar]
- Prestegard, S.K.; Oftedal, L.; Coyne, R.T.; Nygaard, G.; Skjærven, K.H.; Knutsen, G.; Døskeland, S.O.; Herfindal, L. Marine benthic diatoms contain compounds able to induce leukemia cell death and modulate blood platelet activity. Mar. Drugs 2009, 7, 605–623. [Google Scholar] [CrossRef]
- Bag, B.G.; Barai, A.C. Self-assembly of naturally occurring stigmasterol in liquids yielding a fi brillar network and gel. RSC Adv. 2020, 10, 4755–4762. [Google Scholar] [CrossRef]
- Gastineau, R.; Turcotte, F.; Pouvreau, J.B.; Morançais, M.; Fleurence, J.; Windarto, E.; Prasetiya, F.S.; Arsad, S.; Jaouen, P.; Babin, M.; et al. Marennine, promising blue pigments from a widespread Haslea diatom species complex. Mar. Drugs 2014, 12, 3161–3189. [Google Scholar] [CrossRef] [Green Version]
- Carbonnelle, D.; Pondaven, P.; Morançais, M.; Massé, G.; Bosch, S.; Jacquot, C.; Briand, G.; Robert, J.M.; Roussakis, C. Antitumor and antiproliferative effects of an aqueous extract from the marine diatom Haslea ostrearia (Simonsen) against solid tumors: Lung carcinoma (NSCLC-N6), kidney carcinoma (E39) and melanoma (M96) cell lines. Anticancer Res. 1999, 19, 621–624. [Google Scholar]
- Stonik, I.V.; Stonik, I. Low-molecular-weight metabolites from diatoms: Structures, biological roles and biosynthesis. Mar. Biotechnol. 2015, 13, 3672–3709. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, K.; Wang, X.; Nie, S.; Chen, Z.G.; Shin, D.M. Therapeutic nanoparticles for drug delivery in cancer. Clin. Cancer Res. 2008, 14, 1310–1317. [Google Scholar] [CrossRef] [Green Version]
- De Jong, W.H.; Borm, P.J. Drug delivery and nanoparticles: Applications and hazards. Int. J. Nanomed. 2008, 3, 133–149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aw, M.S.; Simovic, S.; Yu, Y.; Addai-mensah, J.; Losic, D. Porous silica microshells from diatoms as biocarrier for drug delivery applications. Powder Technol. 2012, 223, 52–58. [Google Scholar] [CrossRef]
- Zhang, H.; Shahbazi, M.A.; Mäkilä, E.M.; da Silva, T.H.; Reis, R.L.; Salonen, J.J.; Hirvonen, J.T.; Santos, H.A. Biomaterials diatom silica microparticles for sustained release and permeation enhancement following oral delivery of prednisone and mesalamine. Biomaterials 2013, 34, 9210–9219. [Google Scholar] [CrossRef] [PubMed]
- Rea, I.; Martucci, N.M.; De Stefano, L.; Ruggiero, I.; Terracciano, M.; Dardano, P.; Migliaccio, N.; Arcari, P.; Taté, R.; Rendina, I.; et al. Diatomite biosilica nanocarriers for siRNA transport inside cancer cells. Biochim. Biophys. Acta 2014, 1840, 3393–3403. [Google Scholar] [CrossRef] [PubMed]
- Delalat, B.; Sheppard, V.C.; Rasi Ghaemi, S.; Rao, S.; Prestidge, C.A.; McPhee, G.; Rogers, M.L.; Donoghue, J.F.; Pillay, V.; Johns, T.G.; et al. Targeted drug delivery using genetically engineered diatom biosilica. Nat. Commun. 2015, 6, 8791. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delasoie, J.; Zobi, F. Natural diatom biosilica as microshuttles in drug delivery systems. Pharmaceutics 2019, 11, 537. [Google Scholar] [CrossRef] [Green Version]
- Terracciano, M.; Shahbazi, M.A.; Correia, A.; Rea, I.; Lamberti, A.; Stefano, L.D.; Santos, H.A. Surface bioengineering of diatomite based nanovectors for efficient intracellular uptake and drug delivery. Nanoscale 2015, 7, 20063–20074. [Google Scholar] [CrossRef] [Green Version]
- He, Q.; Shi, J.; Chen, F.; Zhu, M.; Zhang, L. Biomaterials an anticancer drug delivery system based on surfactant-templated mesoporous silica nanoparticles. Biomaterials 2010, 31, 3335–3346. [Google Scholar] [CrossRef]
- Cheng, Y.; Qin, S.; Ma, Y.; Chen, X.; Zhang, A.; Zhang, X. Super-pH-sensitive mesoporous silica nanoparticle-based drug delivery system for effective combination cancer therapy. ACS Biomater. Sci. Eng. 2019, 5, 1878–1886. [Google Scholar] [CrossRef]
- Maher, S.; Kumeria, T.; Wang, Y.; Kaur, G.; Fathalla, D.; Fetih, G.; Santos, A.; Habib, F.; Evdokiou, A.; Losic, D. From the mine to cancer therapy: Natural and biodegradable theranostic silicon nanocarriers from diatoms for sustained delivery of chemotherapeutics. Adv. Healthc. Mater. 2016, 5, 2667–2678. [Google Scholar] [CrossRef] [PubMed]
- Maher, S.; Santos, A.; Kumeria, T.; Kaur, G.; Lambert, M.; Forward, P.; Evdokiou, A.; Losic, D. Multifunctional microspherical magnetic and pH responsive carriers for combination anticancer therapy engineered by droplet-based microfluidics. J. Mater. Chem. B 2017, 5, 4097–4109. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, J.G.; Losic, D.; Rosengarten, G.; Vinayak, V.; Rorrer, G.; De Stefano, L.; Kroger, N.; Voelcker, N.; Zhang, Y.X.; Santos, H.A. Diatom Nanotechnology Progress and Emerging Applications; Royal Society of Chemistry: London, UK, 2017. [Google Scholar]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diatoms Species | Anticancer Compounds | Target Cells | IC50 | Time | References |
|---|---|---|---|---|---|
| Skeletonema marinoi | Monoacylglycerides (MAGs) | Haematological cancer cell line (U-937) | 5 µg/mL | 24 h | [28] |
| Colon cancer cell line (HCT-116) | 5 µg/mL | ||||
| MePR-2B normal cells | - | ||||
| Polyunsaturated aldehydes (PUAs 2-trans,4-trans-decadienal (DD)) | A549 | Not clarified | 48 h | [11] | |
| Colon adenocarcinoma metastatic ascites-deriving (COLO 205) | Not clarified | ||||
| Normal lung/brunch epithelial (BEAS-2B) | - | ||||
| Thalassiosira rotula Skeletonema costatum Pseudo-nitzschia delicatissima | Unsaturated aldehydes | Caco-2 cells | 11 ± 17 µg/mL | 48 h | [35] |
| Synedra acus | Chrysolaminaran | Human colon cancer cells (HT-29) | 54.5 µg/mL | 72 h | [18] |
| Colon cell line (DLD-1) | 47.7 µg/mL | ||||
| Phaeodactylum tricornutum | Nonyl 8-acetoxy-6-methyloctanoate (NAMO, fatty alcohol ester) | Human promyelocytic leukemia (HL-60) | 22.3 µg/mL | 48 h | [3] |
| Human lung carcinoma (A549) | 50 µg/mL | ||||
| Mouse melanoma (B16F10) | - | ||||
| Monogalactosyl diacylglycerols | Wild-type W2 Wild-type D3 | 52 µM 64 µM | 48 h | [36] | |
| Fucoxanthin | Caco-2 (derived from a human colon adenocarcinoma), Hep G2, and HeLa (derived from cervical cancer cells) | Not clarified | 48 h | [37] | |
| Navicula incerta | Stigmasterol (phytosterol) | Liver hepatocellular carcinoma (HepG2) | 8.25 µg/mL | 24 h | [38] |
| Haslea ostreria | hasla-6(17),9,13,23- tetraene | Human lung cancer (NSCLC-N6) | 3.8 µg/mL | 72 h | [39] |
| Marennine | Skin cancer (M96), lung cancer (NSCLC-N6), and kidney cancer (E39) | 30.2, 34.2, and 57.8 µg/mL | |||
| Cocconeis scutellum | Fraction 3 (eicosapentaenoic acid (EPA), diethyl ether extract) | Breast carcinoma (BT20) Human normal lymphocytes | Not clarified - | 24 h | [40] |
| Chaetoceros calcitrans | EtOH extract | MCF-7 | 3 µg/mL | 24 h | [41] |
| AcOEt extract | Breast adenocarcinoma (MDA-MB-231) | 60 µg/mL | 72 h | [42] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussein, H.A.; Abdullah, M.A. Anticancer Compounds Derived from Marine Diatoms. Mar. Drugs 2020, 18, 356. https://doi.org/10.3390/md18070356
Hussein HA, Abdullah MA. Anticancer Compounds Derived from Marine Diatoms. Marine Drugs. 2020; 18(7):356. https://doi.org/10.3390/md18070356
Chicago/Turabian StyleHussein, Hanaa Ali, and Mohd Azmuddin Abdullah. 2020. "Anticancer Compounds Derived from Marine Diatoms" Marine Drugs 18, no. 7: 356. https://doi.org/10.3390/md18070356
APA StyleHussein, H. A., & Abdullah, M. A. (2020). Anticancer Compounds Derived from Marine Diatoms. Marine Drugs, 18(7), 356. https://doi.org/10.3390/md18070356