Molecular and Functional Diversity of Crustin-Like Genes in the Shrimp Litopenaeus vannamei


Abstract
:1. Introduction
2. Results
2.1. Identification and Sequence Characterization of Crustin-Like Genes in L. vannamei
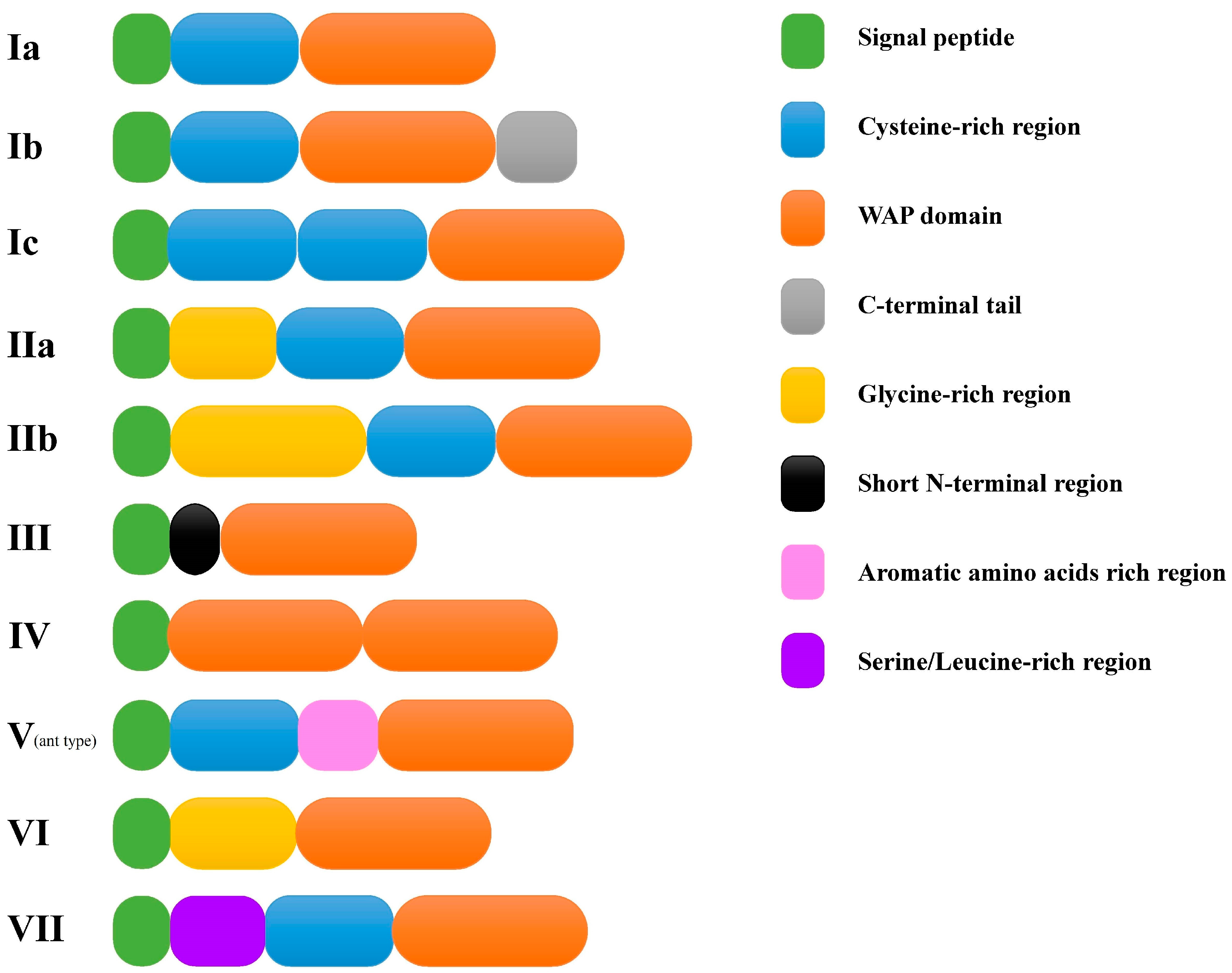
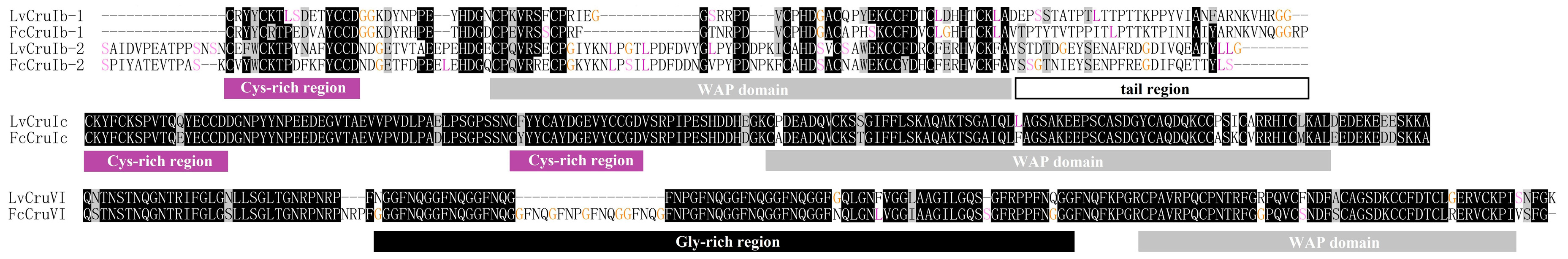
2.2. Identification of New Types and Sequence Characterization of Putative Crustins in L. vannamei
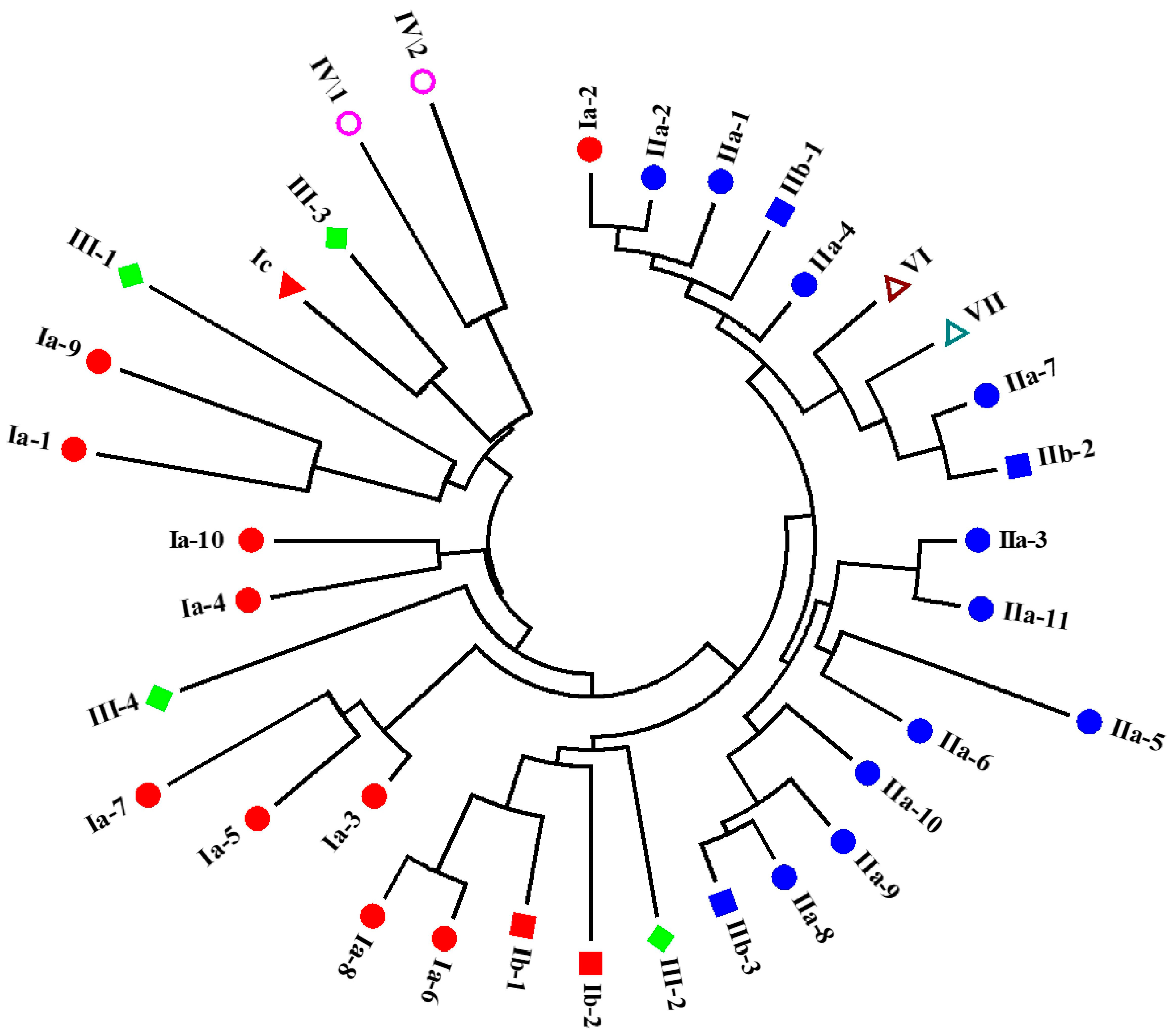
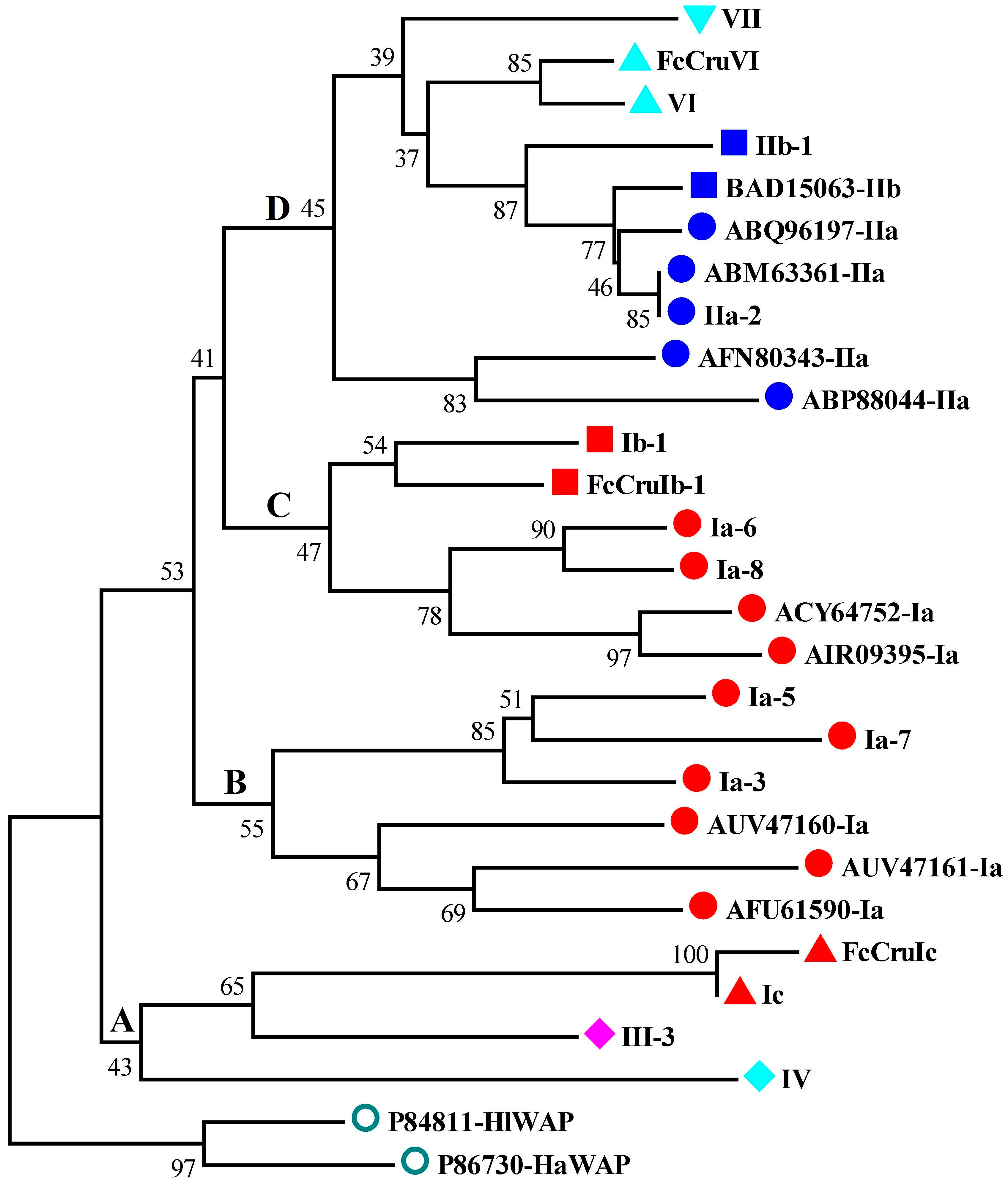
2.3. Phylogenetic Analysis of Different Types of Putative Crustins in Shrimp
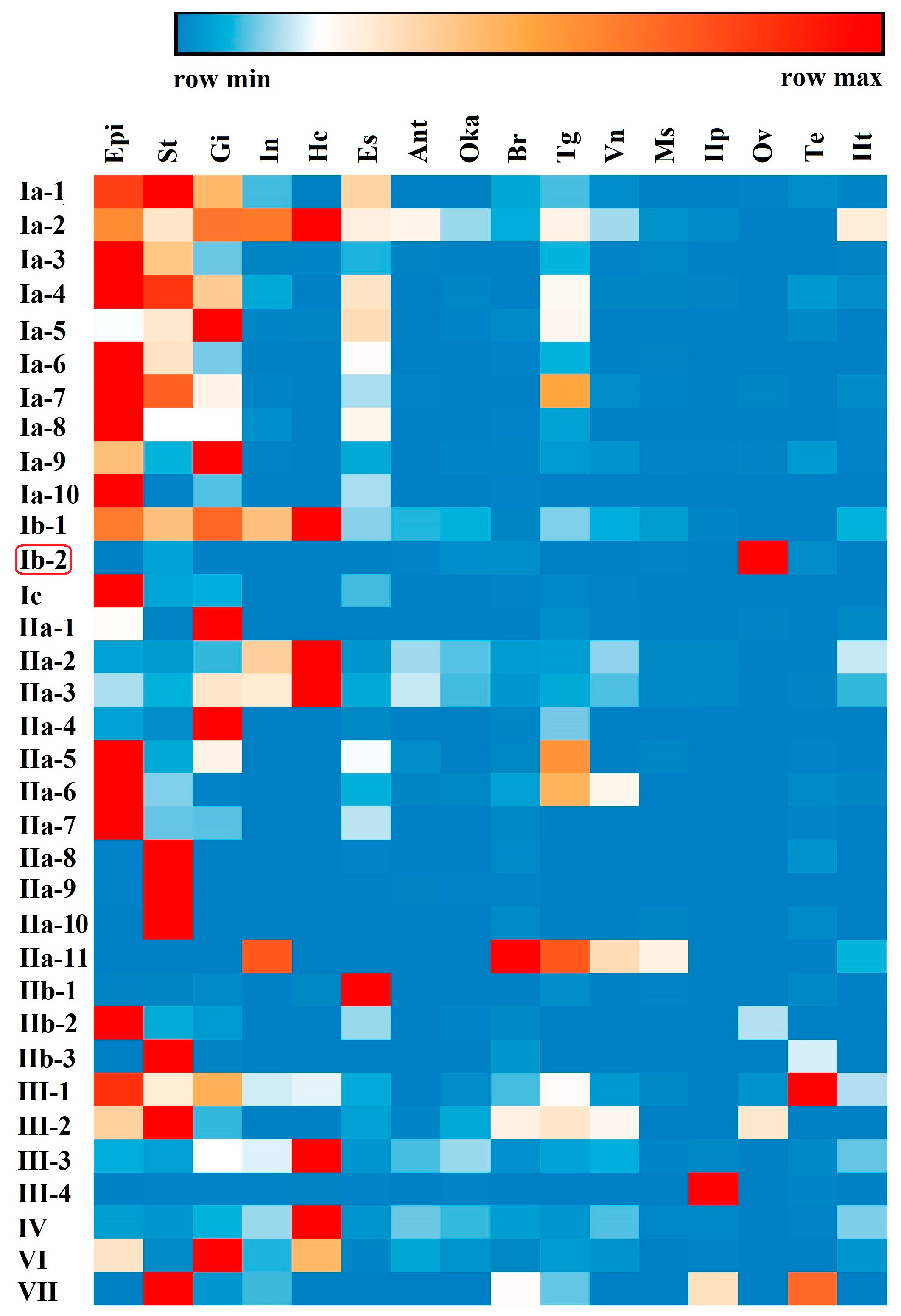
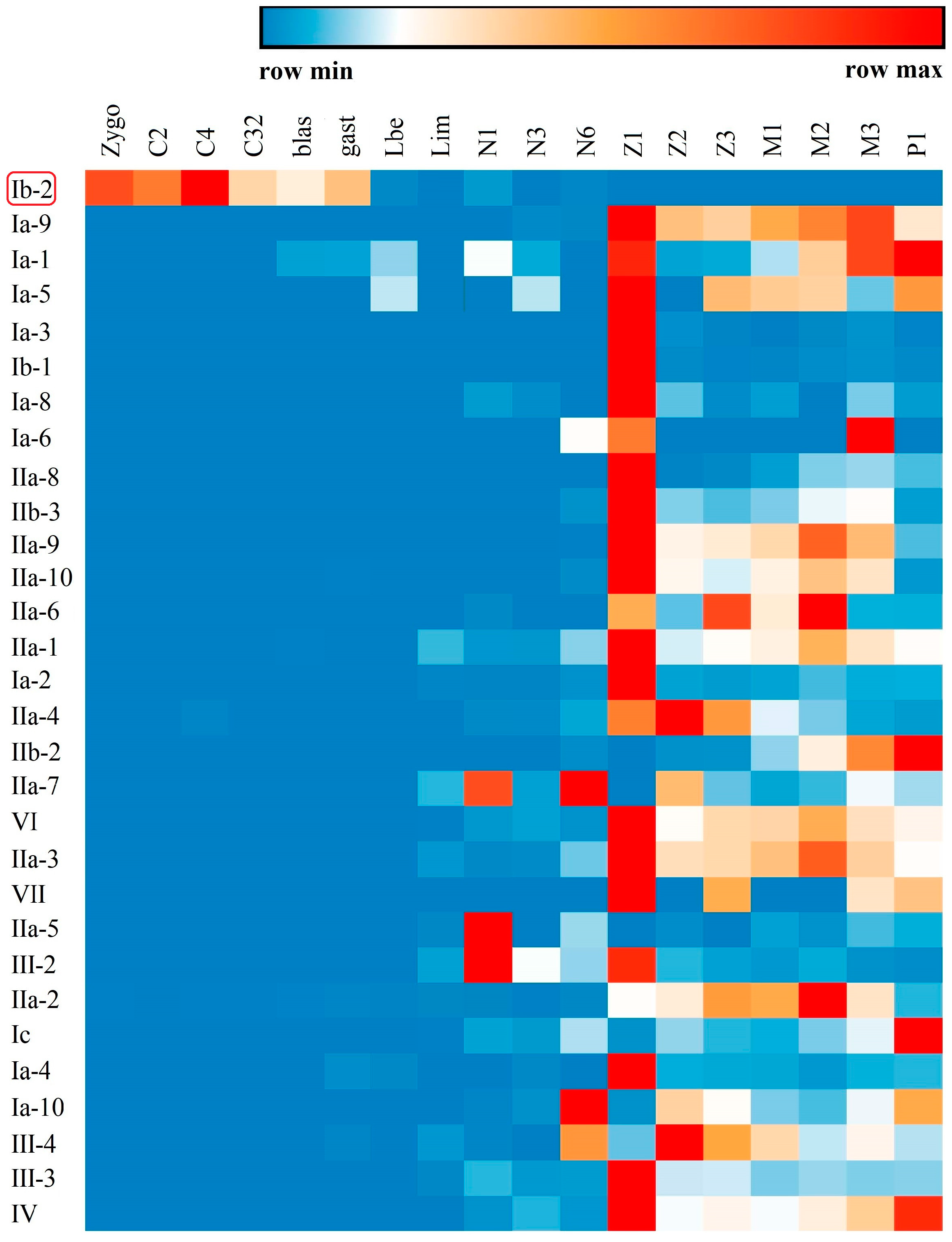
2.4. Spatial and Temporal Distribution of Crustin-Like Transcripts
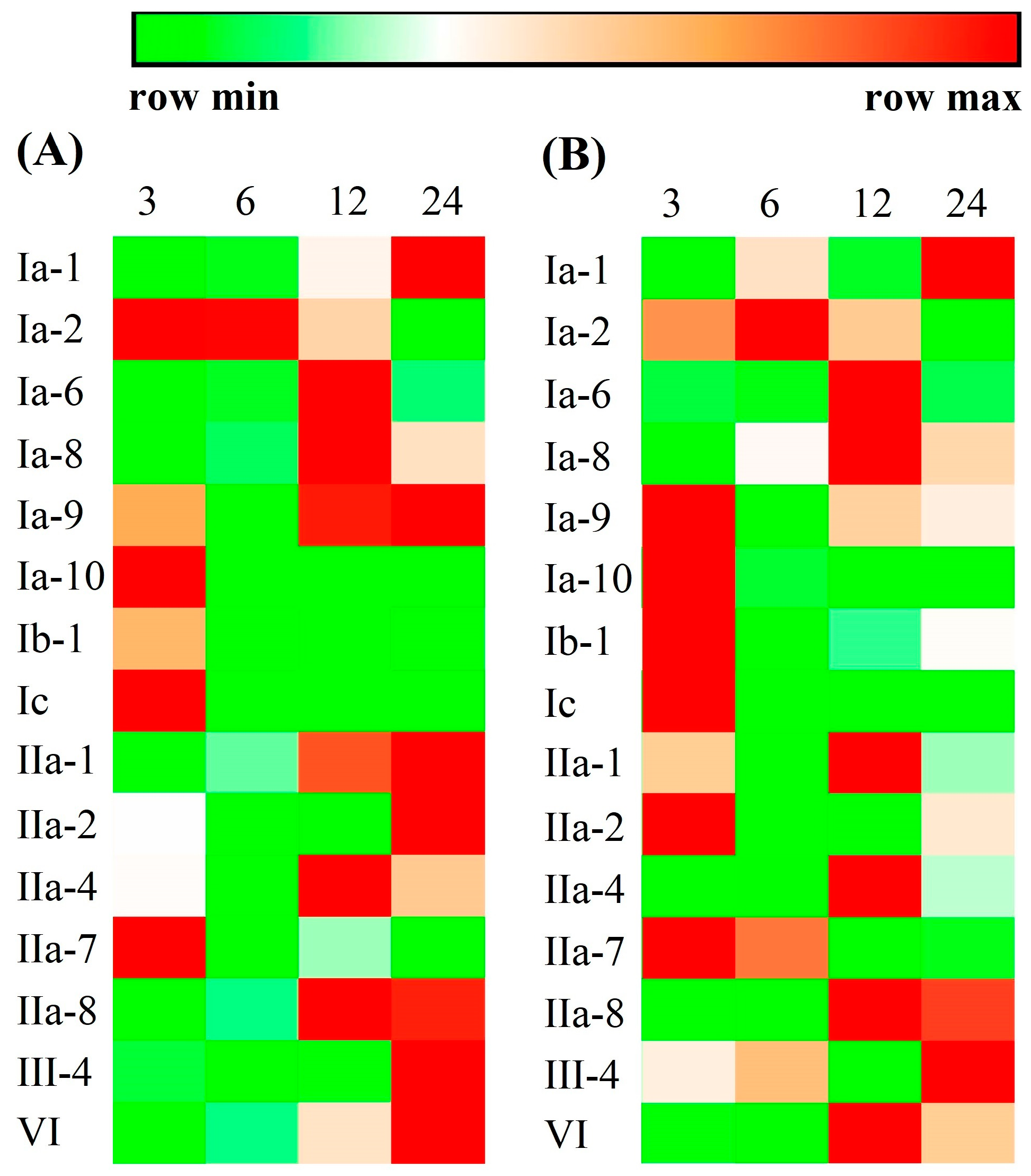
2.5. Immune Responses of Crustin-Like Transcripts on Vibrio parahaemolyticus and WSSV Infection
3. Discussion
4. Materials and Methods
4.1. Database
4.2. Sequence Identification
4.3. Sequence and Expression Analysis
4.4. Animals, Infection, and Tissue Collection
4.5. RNA Extraction and cDNA Synthesis
4.6. Quantitative PCR and Data Analysis
4.7. Ethical Statement
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef]
- Peschel, A.; Sahl, H.-G. The co-evolution of host cationic antimicrobial peptides and microbial resistance. Nat. Rev. Genet. 2006, 4, 529–536. [Google Scholar] [CrossRef] [PubMed]
- Destoumieux-Garzón, D.; Rosa, R.D.; Schmitt, P.; Barreto, C.; Vidal-Dupiol, J.; Mitta, G.; Gueguen, Y.; Bachère, E. Antimicrobial peptides in marine invertebrate health and disease. Philos. Trans. R. Soc. B Biol. Sci. 2016, 371, 20150300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosa, R.D.; Barracco, M.A. Antimicrobial peptides in crustaceans. Invertebr. Surviv. J. 2010, 7, 262–284. [Google Scholar]
- Tassanakajon, A.; Amparyup, P.; Somboonwiwat, K.; Supungul, P. Cationic Antimicrobial Peptides in Penaeid Shrimp. Mar. Biotechnol. 2010, 12, 487–505. [Google Scholar] [CrossRef]
- Tassanakajon, A.; Somboonwiwat, K.; Amparyup, P. Sequence diversity and evolution of antimicrobial peptides in invertebrates. Dev. Comp. Immunol. 2015, 48, 324–341. [Google Scholar] [CrossRef]
- Ranganathan, S.; Simpson, K.J.; Shaw, D.C.; Nicholas, K.R. The whey acidic protein family: A new signature motif and three-dimensional structure by comparative modeling. J. Mol. Graph. Model. 1999, 17, 106–113. [Google Scholar] [CrossRef]
- Hiemstra, P.S.; Maassen, R.J.; Stolk, J.; Heinzel-Wieland, R.; Steffens, G.J.; Dijkman, J.H. Antibacterial activity of antileukoprotease. Infect. Immun. 1996, 64, 4520–4524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schalkwijk, J.; Wiedow, O.; Hirose, S. The trappin gene family: Proteins defined by an N-terminal transglutaminase substrate domain and a C-terminal four-disulphide core. Biochem. J. 1999, 340 Pt 3, 569–577. [Google Scholar] [CrossRef]
- Smith, V.J.; Fernandes, J.M.O.; Kemp, G.D.; Hauton, C. Crustins: Enigmatic WAP domain-containing antibacterial proteins from crustaceans. Dev. Comp. Immunol. 2008, 32, 758–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Relf, J.M.; Chisholm, J.R.S.; Kemp, G.D.; Smith, V.J. Purification and characterization of a cysteine-rich 11.5-kDa antibacterial protein from the granular haemocytes of the shore crab, Carcinus maenas. Eur. J. Biochem. 1999, 264, 350–357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, V.J.; Chisholm, J.R.S. Antimicrobial Proteins in Crustaceans. In Phylogenetic Perspectives on the Vertebrate Immune System; Springer: Boston, MA, USA, 2001; Volume 484, pp. 95–112. [Google Scholar]
- Arockiaraj, J.; Gnanam, A.J.; Muthukrishnan, D.; Gudimella, R.; Milton, J.; Singh, A.; Muthupandian, S.; Kasi, M.; Bhassu, S. Crustin, a WAP domain containing antimicrobial peptide from freshwater prawn Macrobrachium rosenbergii: Immune characterization. Fish Shellfish. Immunol. 2013, 34, 109–118. [Google Scholar] [CrossRef] [PubMed]
- Battison, A.; Summerfield, R.; Patrzykat, A. Isolation and characterisation of two antimicrobial peptides from haemocytes of the American lobster Homarus americanus. Fish Shellfish. Immunol. 2008, 25, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Cui, Z.; Song, C.; Liu, Y.; Wang, S.; Li, Q.; Li, X. Crustins from eyestalk cDNA library of swimming crab Portunus trituberculatus: Molecular characterization, genomic organization and expression analysis. Fish Shellfish. Immunol. 2012, 33, 937–945. [Google Scholar] [CrossRef]
- Hauton, C.; Brockton, V.; Smith, V.J. Cloning of a crustin-like, single whey-acidic-domain, antibacterial peptide from the haemocytes of the European lobster, Homarus gammarus, and its response to infection with bacteria. Mol. Immunol. 2006, 43, 1490–1496. [Google Scholar] [CrossRef]
- Jiravanichpaisal, P.; Lee, S.Y.; Kim, Y.-A.; Andrén, T.; Söderhäll, I. Antibacterial peptides in hemocytes and hematopoietic tissue from freshwater crayfish Pacifastacus leniusculus: Characterization and expression pattern. Dev. Comp. Immunol. 2007, 31, 441–455. [Google Scholar] [CrossRef]
- Pisuttharachai, D.; Fagutao, F.F.; Yasuike, M.; Aono, H.; Yano, Y.; Murakami, K.; Kondo, H.; Aoki, T.; Hirono, I. Characterization of crustin antimicrobial proteins from Japanese spiny lobster Panulirus japonicus. Dev. Comp. Immunol. 2009, 33, 1049–1054. [Google Scholar] [CrossRef]
- Soonthornchai, W.; Rungrassamee, W.; Karoonuthaisiri, N.; Jarayabhand, P.; Klinbunga, S.; Söderhäll, K.; Jiravanichpaisal, P. Expression of immune-related genes in the digestive organ of shrimp, Penaeus monodon, after an oral infection by Vibrio harveyi. Dev. Comp. Immunol. 2010, 34, 19–28. [Google Scholar] [CrossRef]
- Sperstad, S.V.; Haug, T.; Paulsen, V.; Rode, T.M.; Strandskog, G.; Solem, S.T.; Styrvold, O.B.; Stensvåg, K. Characterization of crustins from the hemocytes of the spider crab, Hyas araneus, and the red king crab, Paralithodes camtschaticus. Dev. Comp. Immunol. 2009, 33, 583–591. [Google Scholar] [CrossRef]
- Amparyup, P.; Kondo, H.; Hirono, I.; Aoki, T.; Tassanakajon, A. Molecular cloning, genomic organization and recombinant expression of a crustin-like antimicrobial peptide from black tiger shrimp Penaeus monodon. Mol. Immunol. 2008, 45, 1085–1093. [Google Scholar] [CrossRef]
- Chen, D.; He, N.; Xu, X. Mj-DWD, a double WAP domain-containing protein with antiviral relevance in Marsupenaeus japonicus. Fish Shellfish. Immunol. 2008, 25, 775–781. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhu, S. Comparative genomics analysis of five families of antimicrobial peptide-like genes in seven ant species. Dev. Comp. Immunol. 2012, 38, 262–274. [Google Scholar] [CrossRef] [PubMed]
- Jia, Y.-P.; Sun, Y.-D.; Wang, Z.-H.; Wang, Q.; Wang, X.-W.; Zhao, X.-F.; Wang, J.-X. A single whey acidic protein domain (SWD)-containing peptide from fleshy prawn with antimicrobial and proteinase inhibitory activities. Aquaculture 2008, 284, 246–259. [Google Scholar] [CrossRef]
- Vatanavicharn, T.; Supungul, P.; Puanglarp, N.; Yingvilasprasert, W.; Tassanakajon, A. Genomic structure, expression pattern and functional characterization of crustinPm5, a unique isoform of crustin from Penaeus monodon. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2009, 153, 244–252. [Google Scholar] [CrossRef] [PubMed]
- Chang, Y.-T.; Lin, C.-Y.; Tsai, C.-Y.; Siva, V.; Chu, C.-Y.; Tsai, H.-J.; Song, Y.-L. The New Face of the Old Molecules: Crustin Pm4 and Transglutaminase Type I Serving as RNPs Down-Regulate Astakine-Mediated Hematopoiesis. PLoS ONE 2013, 8, e72793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suleiman, S.; Smith, V.J.; Dyrynda, E.A. Unusual tissue distribution of carcinin, an antibacterial crustin, in the crab, Carcinus maenas, reveals its multi-functionality. Dev. Comp. Immunol. 2017, 76, 274–284. [Google Scholar] [CrossRef] [Green Version]
- Rekha, R.; Vaseeharan, B.; Ishwarya, R.; Anjugam, M.; Alharbi, N.S.; Kadaikunnan, S.; Khaled, J.M.; Al-Anbr, M.N.; Govindarajan, M. Searching for crab-borne antimicrobial peptides: Crustin from Portunus pelagicus triggers biofilm inhibition and immune responses of Artemia salina against GFP tagged Vibrio parahaemolyticus Dahv2. Mol. Immunol. 2018, 101, 396–408. [Google Scholar] [CrossRef]
- Chen, Y.-H.; Lian, Y.-Y.; He, H.-H.; Yuan, K.; Zhang, C.-Z.; Yue, G.H.; He, J.-G. Functional characterization of an ER-stress responding Crustin gene in Litopenaeus vannamei. Fish Shellfish. Immunol. 2019, 84, 541–550. [Google Scholar] [CrossRef]
- Lai, A.G.; Aboobaker, A. Comparative genomic analysis of innate immunity reveals novel and conserved components in crustacean food crop species. BMC Genom. 2017, 18, 389. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Yuan, J.; Sun, Y.; Li, S.; Gao, Y.; Yu, Y.; Liu, C.; Wang, Q.; Lv, X.; Zhang, X.; et al. Penaeid shrimp genome provides insights into benthic adaptation and frequent molting. Nat. Commun. 2019, 10, 356. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Zhang, X.; Sun, Z.; Li, F.; Xiang, J. Transcriptome Analysis on Chinese Shrimp Fenneropenaeus chinensis during WSSV Acute Infection. PLoS ONE 2013, 8, e58627. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, C.; Luo, D.; Cui, Z.; Liu, Y.; Li, X.; Shi, G.; Wang, C. Polymorphism of crustins in the swimming crab (Portunus trituberculatus) and its association withVibrio alginolyticus. Aquac. Res. 2013, 46, 1261–1268. [Google Scholar] [CrossRef]
- Hanson, M.A.; Lemaitre, B. New insights on Drosophila antimicrobial peptide function in host defense and beyond. Curr. Opin. Immunol. 2020, 62, 22–30. [Google Scholar] [CrossRef] [PubMed]
- Zanchi, C.; Johnston, P.R.; Rolff, J. Evolution of defence cocktails: Antimicrobial peptide combinations reduce mortality and persistent infection. Mol. Ecol. 2017, 26, 5334–5343. [Google Scholar] [CrossRef]
- Padhi, A. Adaptive evolution of crustin antimicrobial peptides in decapods. Genetica 2012, 140, 197–203. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.-X.; Wang, J.-X.; Wang, X.-W. First identification and characterization of a triple WAP domain containing protein in Procambarus clarkii provides new insights into the classification and evolution of WAP proteins in crustacean. Fish Shellfish. Immunol. 2019, 94, 592–598. [Google Scholar] [CrossRef]
- Smith, V.J. Phylogeny of whey acidic protein (WAP) four-disulfide core proteins and their role in lower vertebrates and invertebrates. Biochem. Soc. Trans. 2011, 39, 1403–1408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres, A.M.; Wong, H.Y.; Desai, M.; Moochhala, S.; Kuchel, P.; Kini, R.M. Identification of a Novel Family of Proteins in Snake Venoms—Purification and structural characterization of nawaprin from Naja nigricollis snake venom. J. Biol. Chem. 2003, 278, 40097–40104. [Google Scholar] [CrossRef] [Green Version]
- Jiravanichpaisal, P.; Puanglarp, N.; Petkon, S.; Donnuea, S.; Söderhäll, I.; Söderhäll, K. Expression of immune-related genes in larval stages of the giant tiger shrimp, Penaeus monodon. Fish Shellfish. Immunol. 2007, 23, 815–824. [Google Scholar] [CrossRef] [PubMed]
- Treccani, L.; Mann, K.; Heinemann, F.; Fritz, M. Perlwapin, an Abalone Nacre Protein with Three Four-Disulfide Core (Whey Acidic Protein) Domains, Inhibits the Growth of Calcium Carbonate Crystals. Biophys. J. 2006, 91, 2601–2608. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transcriptome_ID | Genome_ID | Gene Symbol | NCBI Accession Number | ORF Length (bp) | Sequence Features |
|---|---|---|---|---|---|
| Unigene0010238 | XP_027238377 | Ia-1 | MT375557 | 318 | SP+Cys+WAP |
| Unigene0016512 | N/A | Ia-2 | MT375558 | 345 | SP+Cys+WAP |
| Unigene0015168 | N/A | Ia-3 | MT375559 | 306 | SP+Cys+WAP |
| Unigene0058358 | XP_027212735 | Ia-4 | MT375560 | 291 | SP+Cys+WAP |
| Unigene0088991 | N/A | Ia-5 | MT375561 | 309 | SP+Cys+WAP |
| Unigene0053725 | ROT80205 | Ia-6 | MT375562 | 327 | SP+Cys+WAP |
| Unigene0088992 | N/A | Ia-7 | MT375563 | 309 | SP+Cys+WAP |
| Unigene0053724 | XP_027219667 | Ia-8 | MT375564 | 363 | SP+Cys+WAP |
| Unigene0062421 | XP_027221901 | Ia-9 | MT375565 | 342 | SP+Cys+WAP |
| Unigene0047792 | XP_027209751 | Ia-10 | MT375566 | 339 | SP+Cys+WAP |
| Unigene0045871 | XP_027212782 | Ib-1 | MT375567/ATU82299 | 381 | SP+Cys+WAP+tail |
| Unigene0068591 | XP_027219394 | Ib-2 | MT375568 | 474 | SP+Cys+WAP+tail |
| Unigene0070992 | N/A | Ic | MT375569 | 546 | SP+Cys+Cys+WAP |
| Unigene0074242 | N/A | IIa-1 | MT375570 | 522 | SP+sGly+Cys+WAP |
| Unigene0014103 | XP_027208055 | IIa-2 | MT375571/AAL36890 | 492 | SP+sGly+Cys+WAP |
| Unigene0047624 | N/A | IIa-3 | MT375572/AFV77524 | 444 | SP+sGly+Cys+WAP |
| Unigene0075484 | N/A | IIa-4 | MT375573 | 492 | SP+sGly+Cys+WAP |
| Unigene0059807 | N/A | IIa-5 | MT375574 | 540 | SP+sGly+Cys+WAP |
| Unigene0070231 | XP_027213162 | IIa-6 | MT375575 | 456 | SP+sGly+Cys+WAP |
| Unigene0046286 | XP_027227947 | IIa-7 | MT375576 | 531 | SP+sGly+Cys+WAP |
| Unigene0003589 | N/A | IIa-8 | MT375577 | 471 | SP+sGly+Cys+WAP |
| Unigene0051595 | N/A | IIa-9 | MT375578 | 384 | SP+sGly+Cys+WAP |
| Unigene0060777 | N/A | IIa-10 | MT375579 | 465 | SP+sGly+Cys+WAP |
| Unigene0106537 | N/A | IIa-11 | MT375580 | 405 | SP+sGly+Cys+WAP |
| Unigene0076000 | N/A | IIb-1 | MT375581 | 732 | SP+lGly+Cys+WAP |
| Unigene0021981 | N/A | IIb-2 | MT375582 | 870 | SP+lGly+Cys+WAP |
| Unigene0035165 | N/A | IIb-3 | MT375583 | 720 | SP+lGly+Cys+WAP |
| Unigene0091111 | N/A | III-1 | MT375584 | 492 | SP+sN+WAP |
| Unigene0069142 | XP_027213161 | III-2 | MT375585 | 252 | SP+sN+WAP |
| Unigene0068231 | XP_027228308 | III-3 | MT375586/AY465833 | 282 | SP+sN+WAP |
| Unigene0047523 | N/A | III-4 | MT375587 | 282 | SP+sN+WAP |
| Unigene0075840 | N/A | IV | MT375588/ABR19819 | 276 | SP+WAP+WAP |
| Unigene0073524 | XP_027229424 | VI | MT375589 | 351 | SP+Gly+WAP |
| Unigene0106295 | XP_027236662 | VII | MT375590 | 516 | SP+Ser/Leu+Cys+WAP |
| Gene Symbol | Primer Name | Nucleotide Sequence (5’~3’) | Annealing Temperature (℃) |
|---|---|---|---|
| Ia-1 | Ia-1F | CTCTGGTGGACATTGACTGC | 55 |
| Ia-1R | GTTGAGGGCGTTGTGGTT | ||
| Ia-10 | Ia-10F | GGGTTGGCGTTACTTGGA | 52 |
| Ia-10R | GGGCAGGTTGGAAACTCATT | ||
| Ia-2 | Ia-2F | CCCACAAGTCCGTCCCACCT | 58 |
| Ia-2R | CTCCCAGACACCTGTCGAAGCAG | ||
| Ia-6 | Ia-6F | CGCCGATGGTTGCAGGTATT | 55 |
| Ia-6R | CATGATGGTCCAAGCAGGTGTC | ||
| Ia-8 | Ia-8F | CGCCGATGGTTGCAGGTAT | 55 |
| Ia-8R | CGTGATGGTCCAAGCAGGTGT | ||
| Ia-9 | Ia-9F | AATGGCGACCCTGATGTTG | 55 |
| Ia-9R | GGTGTCTAAGACGTTCTGCTCC | ||
| Ib-1 | Ib-1F | TGCGATGGCGGGAAAGAT | 55 |
| Ib-1R | GCAGCACTTCTCGTAGGGTTGG | ||
| Ic | Ic-F | GCAGTGCCTCAATCTTATCCTC | 52 |
| Ic-R | TCGGCTGTTACACCTTCATCTT | ||
| IIa-1 | IIa-1F | ACCGATCTTGAACCCGAGGGC | 58 |
| IIa-1R | TGCAGGCACCTGTCGAAGCAG | ||
| IIa-2 | IIa-2F | GGTGTTGGTGGTGGTTTCCC | 55 |
| IIa-2R | CAGTCGCTTGTGCCAGTTCC | ||
| IIa-4 | IIa-4F | TTGTGCTGTCTCCTGGTGGCTGTT | 58 |
| IIa-4R | CGCCAAAGAATCTTGCATTCCCTC | ||
| IIa-7 | IIa-7F | GCAGGTCACCCGAGAATCAGT | 55 |
| IIa-7R | CGAGGCATCTGTCGAAGCAG | ||
| IIa-8 | IIa-8F | GGCCTCATAGCGGGCATCG | 58 |
| IIa-8R | CCTGGTTCTGGGGCGTCTTG | ||
| III-4 | III-4F | GGAAATCCAGTCCGGCACTCT | 56 |
| III-4R | AAGCAGCGGTCCCAGCAA | ||
| VI | VI-F | CCCAGAACACCAACAGCACC | 55 |
| VI-R | GTCCACCGACGAAGTTACCG | ||
| 18S rRNA | 18S-F | TATACGCTAGTGGAGCTGGAA | 55 |
| 18S-R | GGGGAGGTAGTGACGAAAAAT |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Lv, X.; Yu, Y.; Zhang, X.; Li, F. Molecular and Functional Diversity of Crustin-Like Genes in the Shrimp Litopenaeus vannamei. Mar. Drugs 2020, 18, 361. https://doi.org/10.3390/md18070361
Li S, Lv X, Yu Y, Zhang X, Li F. Molecular and Functional Diversity of Crustin-Like Genes in the Shrimp Litopenaeus vannamei. Marine Drugs. 2020; 18(7):361. https://doi.org/10.3390/md18070361
Chicago/Turabian StyleLi, Shihao, Xinjia Lv, Yang Yu, Xiaojun Zhang, and Fuhua Li. 2020. "Molecular and Functional Diversity of Crustin-Like Genes in the Shrimp Litopenaeus vannamei" Marine Drugs 18, no. 7: 361. https://doi.org/10.3390/md18070361
APA StyleLi, S., Lv, X., Yu, Y., Zhang, X., & Li, F. (2020). Molecular and Functional Diversity of Crustin-Like Genes in the Shrimp Litopenaeus vannamei. Marine Drugs, 18(7), 361. https://doi.org/10.3390/md18070361