Immunomodulatory Effects of N-Acetyl Chitooligosaccharides on RAW264.7 Macrophages

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Effects of NACOS on the Non-Specific Immune Response of RAW264.7 Cells
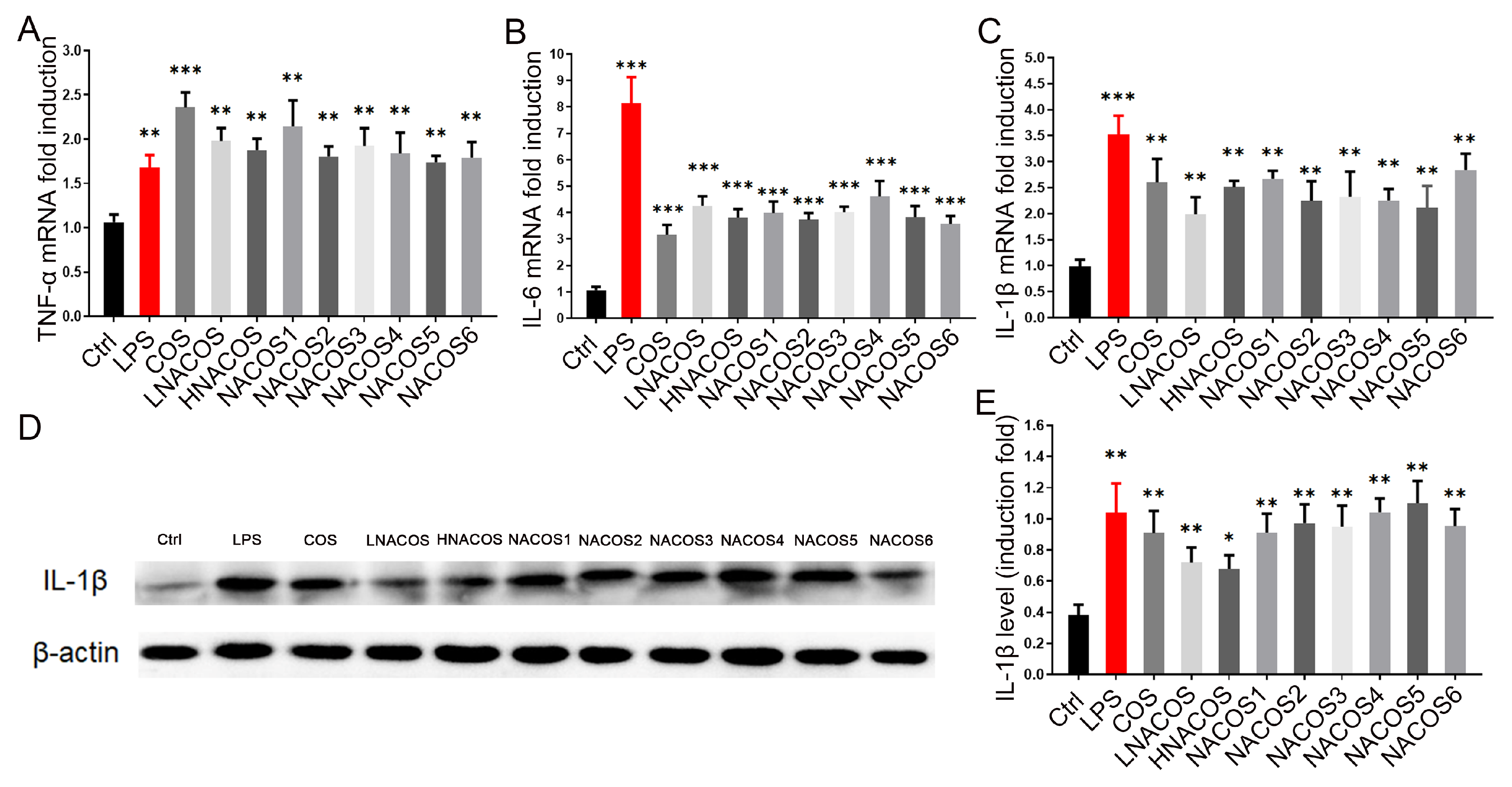
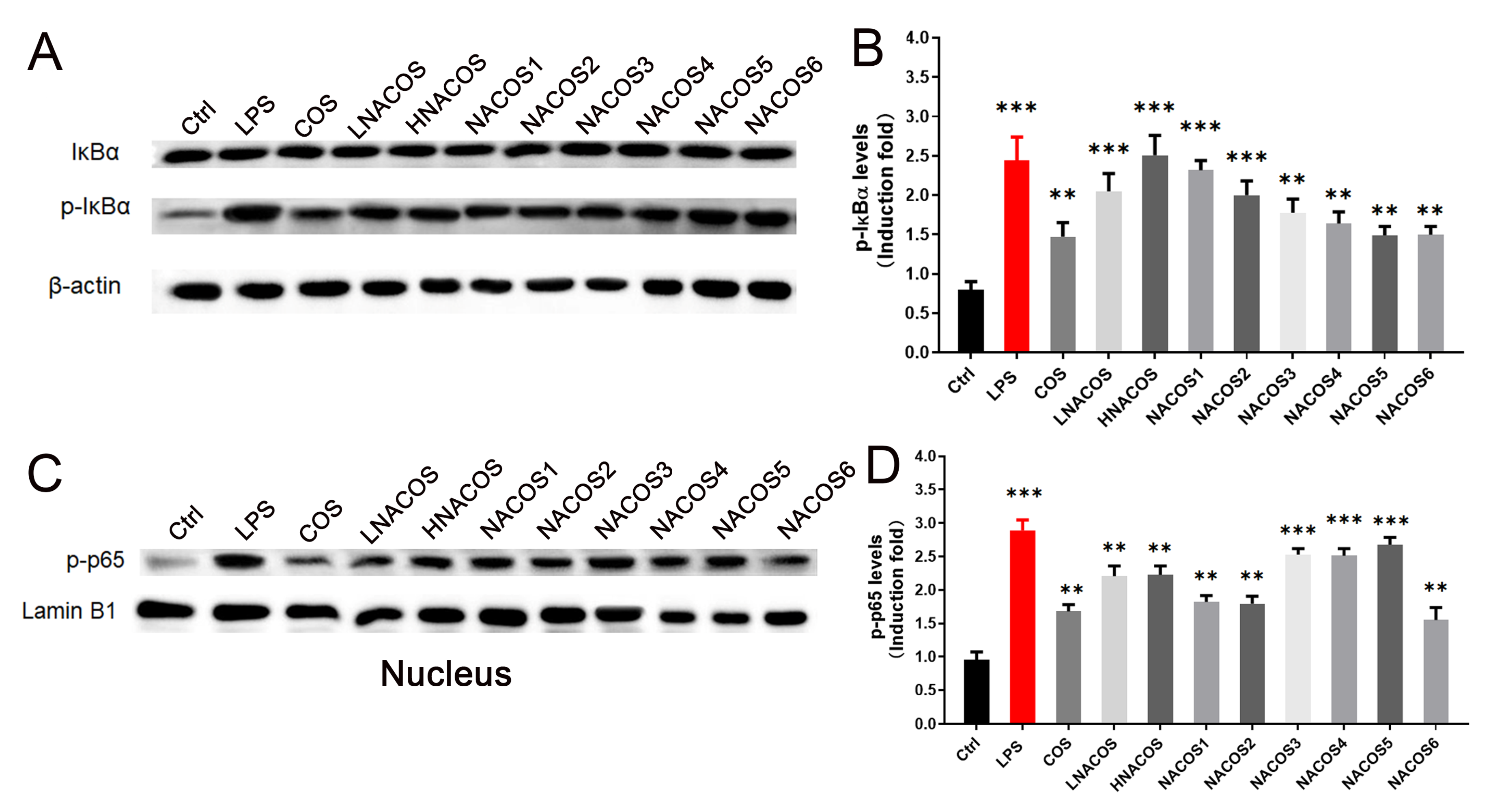
2.2. NACOS Promoted the Production of Pro-Inflammatory Cytokines via the NF-κB Pathway
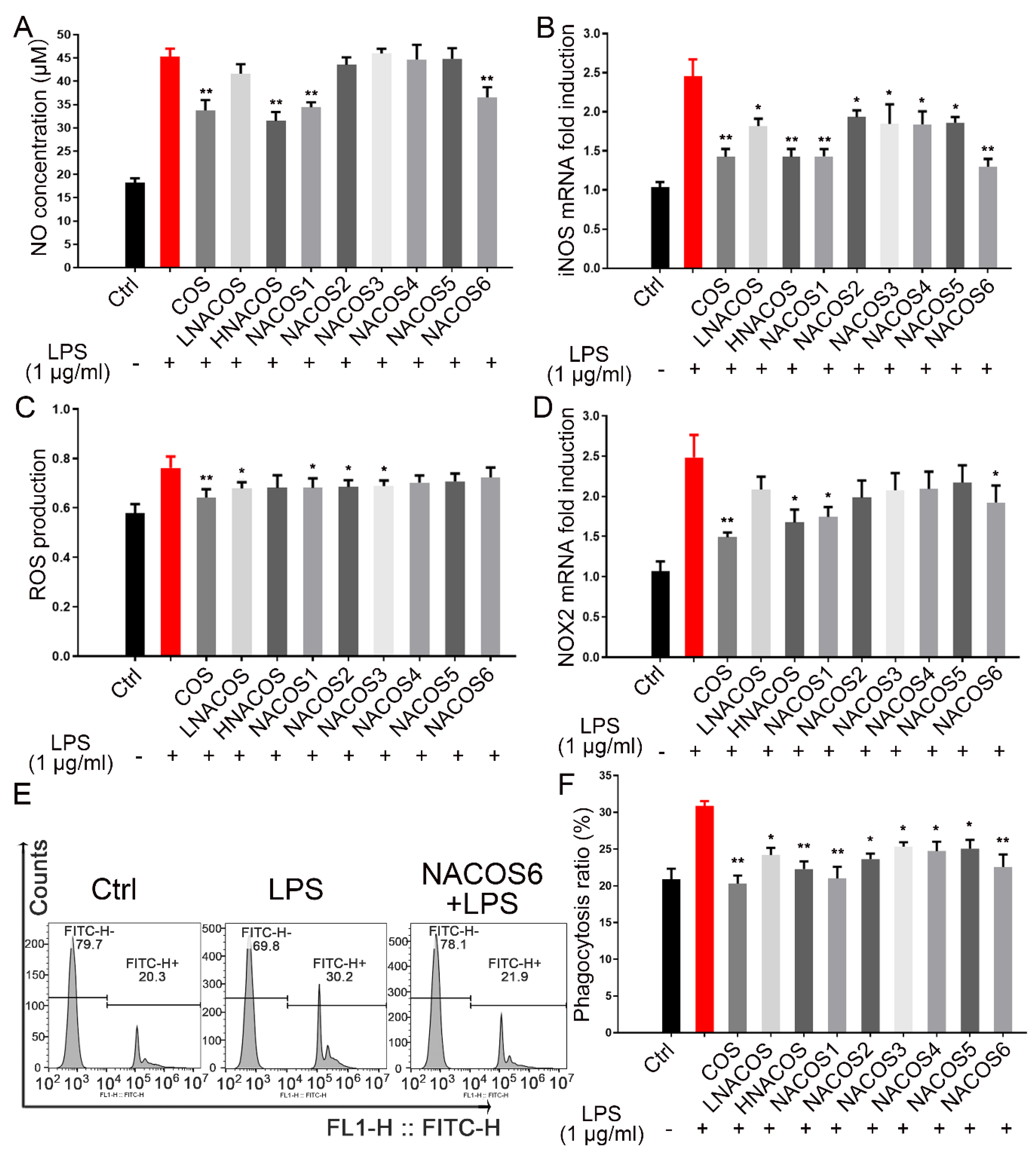
2.3. NACOS Inhibited the LPS-Induced RAW264.7 Inflammatory Response
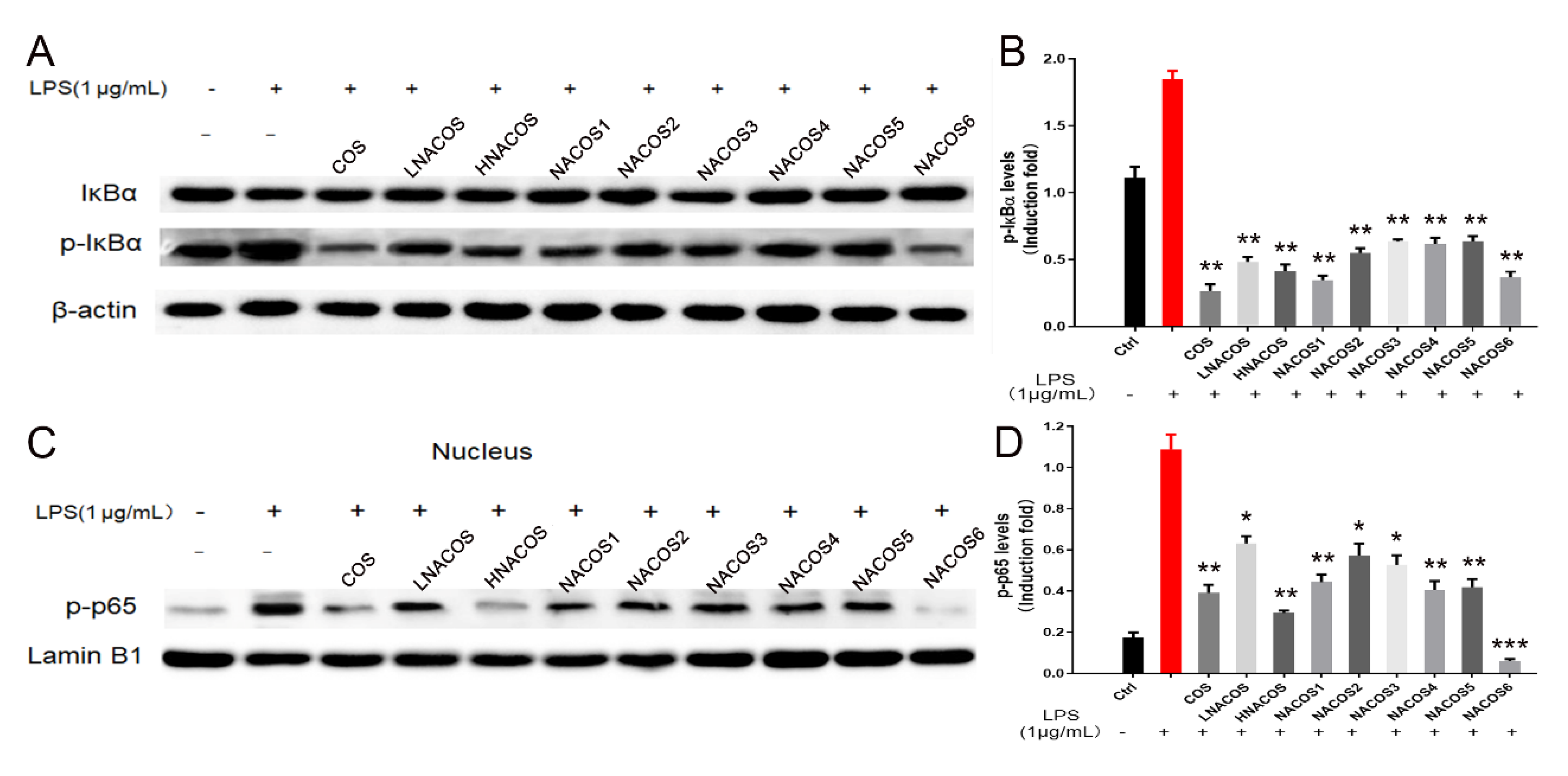
2.4. NACOS Inhibited LPS-Induced Inflammation through the NF-κB Pathway
3. Discussion
4. Materials and Methods
4.1. Materials and Reagents
4.2. Cell Culture and Treatment
4.3. Cell Viability Assay
4.4. Measurement of NO Production
4.5. Measurement of ROS Production
4.6. Phagocytosis Assay
4.7. Quantitative Real-Time PCR (qPCR)
4.8. Western Blotting Analysis
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Domard, A. A perspective on 30 years research on chitin and chitosan. Carbohydr. Polym. 2011, 84, 696–703. [Google Scholar] [CrossRef]
- Rinaudo, M. Chitin and chitosan: Properties and applications. Prog. Polym. Sci. 2006, 31, 603–632. [Google Scholar] [CrossRef]
- Liang, S.; Sun, Y.; Dai, X. A Review of the Preparation, Analysis and Biological Functions of Chitooligosaccharide. Int. J. Mol. Sci. 2018, 19, 2197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naveed, M.; Phil, L.; Sohail, M.; Hasnat, M.; Baig, M.; Ihsan, A.U.; Shumzaid, M.; Kakar, M.U.; Khan, T.M.; Akabar, M.D.; et al. Chitosan oligosaccharide (COS): An overview. Int. J. Biol. Macromol. 2019, 129, 827–843. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.; Liu, P.; Liu, J. Advance in chitosan hydrolysis by non-specific cellulases. Bioresour. Technol. 2008, 99, 6751–6762. [Google Scholar] [CrossRef] [PubMed]
- Viens, P.; Lacombe-Harvey, M.E.; Brzezinski, R. Chitosanases from Family 46 of Glycoside Hydrolases: From Proteins to Phenotypes. Mar. Drugs 2015, 13, 6566–6587. [Google Scholar] [CrossRef]
- Wei, X.; Wang, Y.; Zhu, Q.; Xiao, J.; Xia, W. Effects of chitosan pentamer and chitosan hexamerin vivoandin vitroon gene expression and secretion of cytokines. Food Agric. Immunol. 2009, 20, 269–280. [Google Scholar] [CrossRef]
- Kim, K.W.; Thomas, R.L. Antioxidative activity of chitosans with varying molecular weights. Food Chem. 2007, 101, 308–313. [Google Scholar] [CrossRef]
- Li, X.; Wang, J.; Chen, X.; Tian, J.; Li, L.; Zhao, M.; Jiao, Y.; Zhou, C. Effect of chitooligosaccharides on cyclin D1, bcl-xl and bcl-2 mRNA expression in A549 cells using quantitative PCR. Chin. Sci. Bull. 2011, 56, 1629–1632. [Google Scholar] [CrossRef] [Green Version]
- Tsai, G.J.; Su, W.H.; Chen, H.C.; Pan, C.L. Antimicrobial activity of shrimp chitin and chitosan from different treatments and applications of fish preservation. Fish. Sci. 2002, 68, 170–177. [Google Scholar] [CrossRef] [Green Version]
- Huang, R.; Mendis, E.; Kim, S.K. Improvement of ACE inhibitory activity of chitooligosaccharides (COS) by carboxyl modification. Bioorg. Med. Chem. 2005, 13, 3649–3655. [Google Scholar] [CrossRef] [PubMed]
- Kondo, Y.; Nakatani, A.; Hayashi, M.; Ito, M. Low molecular weight chitosan prevents the progression of low dose streptozotocin-induced slowly progressive diabetes mellitus in mice. Biol. Pharm. Bull. 2000, 23, 1458–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Zhou, Y.; Zhao, X.; Wang, H.; Wang, L.; Yuan, G.; Asim, M.; Wang, W.; Zeng, L.; Liu, X.; et al. Oligochitosan stimulated phagocytic activity of macrophages from blunt snout bream (Megalobrama amblycephala) associated with respiratory burst coupled with nitric oxide production. Dev. Comp. Immunol. 2014, 47, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Xing, R.; Liu, S.; Qin, Y.; Li, K.; Yu, H.; Li, P. Immunostimulatory Effects of Chitooligosaccharides on RAW 264.7 Mouse Macrophages via Regulation of the MAPK and PI3K/Akt Signaling Pathways. Mar. Drugs 2019, 17, 36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, B.; Wen, Z.S.; Huang, Y.J.; Xia, M.S.; Xiang, X.W.; Qu, Y.L. Molecular Weight-Dependent Immunostimulative Activity of Low Molecular Weight Chitosan via Regulating NF-kappaB and AP-1 Signaling Pathways in RAW264.7 Macrophages. Mar. Drugs 2016, 14, 169. [Google Scholar] [CrossRef] [Green Version]
- Qiao, Y.; Ruan, Y.; Xiong, C.; Xu, Q.; Wei, P.; Ma, P.; Bai, X.; Du, Y. Chitosan oligosaccharides suppressant LPS binding to TLR4/MD-2 receptor complex. Carbohydr. Polym. 2010, 82, 405–411. [Google Scholar] [CrossRef]
- Ma, P.; Liu, H.-T.; Wei, P.; Xu, Q.-S.; Bai, X.-F.; Du, Y.-G.; Yu, C. Chitosan oligosaccharides inhibit LPS-induced over-expression of IL-6 and TNF-α in RAW264.7 macrophage cells through blockade of mitogen-activated protein kinase (MAPK) and PI3K/Akt signaling pathways. Carbohydr. Polym. 2011, 84, 1391–1398. [Google Scholar] [CrossRef]
- Li, Y.; Liu, H.; Xu, Q.S.; Du, Y.G.; Xu, J. Chitosan oligosaccharides block LPS-induced O-GlcNAcylation of NF-kappaB and endothelial inflammatory response. Carbohydr. Polym. 2014, 99, 568–578. [Google Scholar] [CrossRef] [Green Version]
- Tokoro, A.; Tatewaki, N.; Suzuki, K.; Mikami, T.; Suzuki, S.; Suzuki, M. Growth-inhibitory effect of hexa-N-acetylchitohexaose and chitohexaose against Meth-A solid tumor. Chem. Pharm. Bull. 1988, 36, 784–790. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, M.; Watanabe, T.; Suzuki, S.; Suzuki, M. Effect of N-acetylchitohexaose against Candida albicans infection of tumor-bearing mice. Microbiol. Immunol. 1990, 34, 413–426. [Google Scholar] [CrossRef]
- Xu, Q.; Liu, M.; Liu, Q.; Wang, W.; Du, Y.; Yin, H. The inhibition of LPS-induced inflammation in RAW264.7 macrophages via the PI3K/Akt pathway by highly N-acetylated chitooligosaccharide. Carbohydr. Polym. 2017, 174, 1138–1143. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.W.; Liu, G.J.; Yang, Y.; Shao, K.; Wang, Y.T.; Liu, W.S.; Han, B.Q. N-Acetyl chitooligosaccharides attenuate amyloid beta-induced damage in animal and cell models of Alzheimer‘s disease. Process Biochem. 2019, 84, 161–171. [Google Scholar] [CrossRef]
- Ngo, D.-N.; Lee, S.-H.; Kim, M.-M.; Kim, S.-K. Production of chitin oligosaccharides with different molecular weights and their antioxidant effect in RAW 264.7 cells. J. Funct. Foods 2009, 1, 188–198. [Google Scholar] [CrossRef]
- Jung, W.-J.; Souleimanov, A.; Park, R.-D.; Smith, D.L. Enzymatic production of N-acetyl chitooligosaccharides by crude enzyme derived from Paenibacillus illioisensis KJA-424. Carbohydr. Polym. 2007, 67, 256–259. [Google Scholar] [CrossRef]
- Suzuki, K.; Tokoro, A.; Okawa, Y.; Suzuki, S.; Suzuki, M. Effect of N-acetyl chitooligosaccharides on activation of phagocytes. Microbiol. Immunol. 1986, 30, 777–787. [Google Scholar] [CrossRef]
- Xing, R.; Liu, Y.; Li, K.; Yu, H.; Liu, S.; Yang, Y.; Chen, X.; Li, P. Monomer composition of chitooligosaccharides obtained by different degradation methods and their effects on immunomodulatory activities. Carbohydr. Polym. 2017, 157, 1288–1297. [Google Scholar] [CrossRef]
- Naberezhnykh, G.A.; Gorbach, V.I.; Kalmykova, E.N.; Solov‘eva, T.F. Determination of the parameters of binding between lipopolysaccharide and chitosan and its N-acetylated derivative using a gravimetric piezoquartz biosensor. Biophys. Chem. 2015, 198, 9–13. [Google Scholar] [CrossRef]
- Sanchez, A.; Mengibar, M.; Fernandez, M.; Alemany, S.; Heras, A.; Acosta, N. Influence of Preparation Methods of Chitooligosaccharides on Their Physicochemical Properties and Their Anti-Inflammatory Effects in Mice and in RAW264.7 Macrophages. Mar. Drugs 2018, 16, 430. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, T.; Nakano, T.; Takamizawa, D.; Desaki, Y.; Ishii-Minami, N.; Nishizawa, Y.; Minami, E.; Okada, K.; Yamane, H.; Kaku, H.; et al. Two LysM receptor molecules, CEBiP and OsCERK1, cooperatively regulate chitin elicitor signaling in rice. Plant J. 2010, 64, 204–214. [Google Scholar] [CrossRef] [Green Version]
- Hayafune, M.; Berisio, R.; Marchetti, R.; Silipo, A.; Kayama, M.; Desaki, Y.; Arima, S.; Squeglia, F.; Ruggiero, A.; Tokuyasu, K.; et al. Chitin-induced activation of immune signaling by the rice receptor CEBiP relies on a unique sandwich-type dimerization. Proc. Natl. Acad. Sci. USA 2014, 111, E404–E413. [Google Scholar] [CrossRef] [Green Version]
- Azuma, K.; Osaki, T.; Wakuda, T.; Tsuka, T.; Imagawa, T.; Okamoto, Y.; Minami, S. Suppressive effects of N-acetyl-D-glucosamine on rheumatoid arthritis mouse models. Inflammation 2012, 35, 1462–1465. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.J.; Shi, D.; Mao, H.H.; Li, Z.W.; Liang, S.; Ke, Y.; Luo, X.C. Heterologous expression and characterization of an antifungal chitinase (Chit46) from Trichoderma harzianum GIM 3.442 and its application in colloidal chitin conversion. Int. J. Biol. Macromol. 2019, 134, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Deng, J.-J.; Mao, H.-H.; Fang, W.; Li, Z.-Q.; Shi, D.; Li, Z.-W.; Zhou, T.; Luo, X.-C. Enzymatic conversion and recovery of protein, chitin, and astaxanthin from shrimp shell waste. J. Clean. Prod. 2020, 271, 122655. [Google Scholar] [CrossRef]
- Bogdan, C. Nitric oxide and the immune response. Nat. Immunol. 2001, 2, 907–916. [Google Scholar] [CrossRef]
- Flannagan, R.S.; Cosio, G.; Grinstein, S. Antimicrobial mechanisms of phagocytes and bacterial evasion strategies. Nat. Rev. Microbiol. 2009, 7, 355–366. [Google Scholar] [CrossRef]
- Zhang, Y.; Choksi, S.; Chen, K.; Pobezinskaya, Y.; Linnoila, I.; Liu, Z.G. ROS play a critical role in the differentiation of alternatively activated macrophages and the occurrence of tumor-associated macrophages. Cell Res. 2013, 23, 898–914. [Google Scholar] [CrossRef] [Green Version]
- Murakami, A.; Ohigashi, H. Targeting NOX, INOS and COX-2 in inflammatory cells: Chemoprevention using food phytochemicals. Int. J. Cancer 2007, 121, 2357–2363. [Google Scholar] [CrossRef]
- Santos-Moriano, P.; Kidibule, P.; Miguez, N.; Fernandez-Arrojo, L.; Ballesteros, A.O.; Fernandez-Lobato, M.; Plou, F.J. Tailored Enzymatic Synthesis of Chitooligosaccharides with Different Deacetylation Degrees and Their Anti-Inflammatory Activity. Catalysts 2019, 9, 405. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.H.; Senevirathne, M.; Ahn, C.B.; Kim, S.K.; Je, J.Y. Factors affecting anti-inflammatory effect of chitooligosaccharides in lipopolysaccharides-induced RAW264.7 macrophage cells. Bioorg. Med. Chem. Lett. 2009, 19, 6655–6658. [Google Scholar] [CrossRef]
- Li, K.; Xing, R.; Liu, S.; Li, P. Advances in preparation, analysis and biological activities of single chitooligosaccharides. Carbohydr. Polym. 2016, 139, 178–190. [Google Scholar] [CrossRef]
- Komi, D.E.A.; Sharma, L.; Cruz, C.S.D. Chitin and Its Effects on Inflammatory and Immune Responses. Clin. Rev. Allergy Immunol. 2018, 54, 213–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müllebner, A.; Dorighello, G.G.; Kozlov, A.V.; Duvigneau, J.C. Interaction between Mitochondrial Reactive Oxygen Species, Heme Oxygenase, and Nitric Oxide Synthase Stimulates Phagocytosis in Macrophages. Front. Med. 2018, 4, 252. [Google Scholar] [CrossRef] [Green Version]
- Kleinert, H.; Schwarz, P.M.; Förstermann, U. Regulation of the Expression of Inducible Nitric Oxide Synthase. Biol. Chem. 2003, 384, 1343–1364. [Google Scholar] [CrossRef] [PubMed]
- Davydova, V.N.; Bratskaya, S.Y.; Gorbach, V.I.; Solov‘eva, T.F.; Kaca, W.; Yermak, I.M. Comparative study of electrokinetic potentials and binding affinity of lipopolysaccharides-chitosan complexes. Biophys. Chem. 2008, 136, 1–6. [Google Scholar] [CrossRef]
- Naberezhnykh, G.A.; Gorbach, V.I.; Likhatskaya, G.N.; Davidova, V.N.; Solov‘eva, T.F. Interaction of chitosans and their N-acylated derivatives with lipopolysaccharide of gram-negative bacteria. Biochemistry 2008, 73, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Golks, A.; Guerini, D. The O-linked N-acetylglucosamine modification in cellular signalling and the immune system. EMBO Rep. 2008, 9, 748–753. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.; Xie, M.; Men, L.; Du, J. O-GlcNAcylation in immunity and inflammation: An intricate system (Review). Int. J. Mol. Med. 2019, 44, 363–374. [Google Scholar] [CrossRef] [Green Version]
- Baudoin, L.; Issad, T. O-GlcNAcylation and Inflammation: A Vast Territory to Explore. Front. Endocrinol. 2014, 5, 235. [Google Scholar] [CrossRef] [Green Version]
- Pizzolatti, A.L.A.; Gaudig, F.; Seitz, D.; Roesler, C.R.M.; Salmoria, G.V. Glucosamine Hydrochloride and N-Acetylglucosamine Influence the Response of Bovine Chondrocytes to TGF-beta3 and IGF in Monolayer and Three-Dimensional Tissue Culture. Tissue Eng. Regen. Med. 2018, 15, 781–791. [Google Scholar] [CrossRef]
- Shikhman, A.R.; Brinson, D.C.; Valbracht, J.; Lotz, M.K. Differential metabolic effects of glucosamine and N-acetylglucosamine in human articular chondrocytes. Osteoarthr. Cartil. 2009, 17, 1022–1028. [Google Scholar] [CrossRef] [Green Version]
- Zou, L.; Yang, S.; Champattanachai, V.; Hu, S.; Chaudry, I.H.; Marchase, R.B.; Chatham, J.C. Glucosamine improves cardiac function following trauma-hemorrhage by increased protein O-GlcNAcylation and attenuation of NF-{kappa}B signaling. Am. J. Physiol. Heart Circ. Physiol. 2009, 296, H515–H523. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeuchi, T.; Nagasaka, M.; Shimizu, M.; Tamura, M.; Arata, Y. N-acetylglucosamine suppresses osteoclastogenesis in part through the promotion of O-GlcNAcylation. Bone Rep. 2016, 5, 15–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryczko, M.C.; Pawling, J.; Chen, R.; Rahman, A.M.A.; Yau, K.; Copeland, J.K.; Zhang, C.; Surendra, A.; Guttman, D.S.; Figeys, D.; et al. Metabolic Reprogramming by Hexosamine Biosynthetic and Golgi N-Glycan Branching Pathways. Sci. Rep. 2016, 6, 23043. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Barreda, D.R.; Zhang, Y.A.; Boshra, H.; Gelman, A.E.; Lapatra, S.; Tort, L.; Sunyer, J.O. B lymphocytes from early vertebrates have potent phagocytic and microbicidal abilities. Nat. Immunol. 2006, 7, 1116–1124. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Samples | Composition (%) | DA (%) | ||||||
|---|---|---|---|---|---|---|---|---|
| DP1 | DP2 | DP3 | DP4 | DP5 | DP6 | DP7 | ||
| COS | 7.5 | 11.0 | 21.9 | 25.9 | 18.1 | 8.7 | 3.1 | >95 |
| LNACOS | 1.5 | 94.8 | 2.8 | 0.9 | - | - | - | 8.7 |
| HNACOS | - | 37.4 | 10.4 | 32.3 | 6.1 | 13.8 | - | 8.6 |
| Genes | Forward Primer (5′ 3′) | Reverse Primer (5′ 3′) |
|---|---|---|
| iNOS | GGTAGTAGTAGAATGGAGATAGG | CTACCTAAGATAGCAGTTGATG |
| NOX2 | ACCAGACAGACTTGAGAATG | GCTGTGCTATGTTGCTCTAG |
| IL-1β | ATCTCGCAGCAGCACATC | CCAGCAGGTTATCATCATCATC |
| TNF-α | CACGCTCTTCTGTCTACTG | ACTTGGTGGTTTGCTAC |
| IL-6 | AATTAAGCCTCCGACTTGTG | CACGCTCTTCTGTCTACTG |
| β-actin | GCTGTGCTATGTTGCTCTAG | TCGTTGCCAATAGTGATGAC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deng, J.-J.; Li, Z.-Q.; Mo, Z.-Q.; Xu, S.; Mao, H.-H.; Shi, D.; Li, Z.-W.; Dan, X.-M.; Luo, X.-C. Immunomodulatory Effects of N-Acetyl Chitooligosaccharides on RAW264.7 Macrophages. Mar. Drugs 2020, 18, 421. https://doi.org/10.3390/md18080421
Deng J-J, Li Z-Q, Mo Z-Q, Xu S, Mao H-H, Shi D, Li Z-W, Dan X-M, Luo X-C. Immunomodulatory Effects of N-Acetyl Chitooligosaccharides on RAW264.7 Macrophages. Marine Drugs. 2020; 18(8):421. https://doi.org/10.3390/md18080421
Chicago/Turabian StyleDeng, Jun-Jin, Zong-Qiu Li, Ze-Quan Mo, Shun Xu, He-Hua Mao, Dan Shi, Zhi-Wei Li, Xue-Ming Dan, and Xiao-Chun Luo. 2020. "Immunomodulatory Effects of N-Acetyl Chitooligosaccharides on RAW264.7 Macrophages" Marine Drugs 18, no. 8: 421. https://doi.org/10.3390/md18080421
APA StyleDeng, J. -J., Li, Z. -Q., Mo, Z. -Q., Xu, S., Mao, H. -H., Shi, D., Li, Z. -W., Dan, X. -M., & Luo, X. -C. (2020). Immunomodulatory Effects of N-Acetyl Chitooligosaccharides on RAW264.7 Macrophages. Marine Drugs, 18(8), 421. https://doi.org/10.3390/md18080421