Studies of Conorfamide-Sr3 on Human Voltage-Gated Kv1 Potassium Channel Subtypes


 and
and 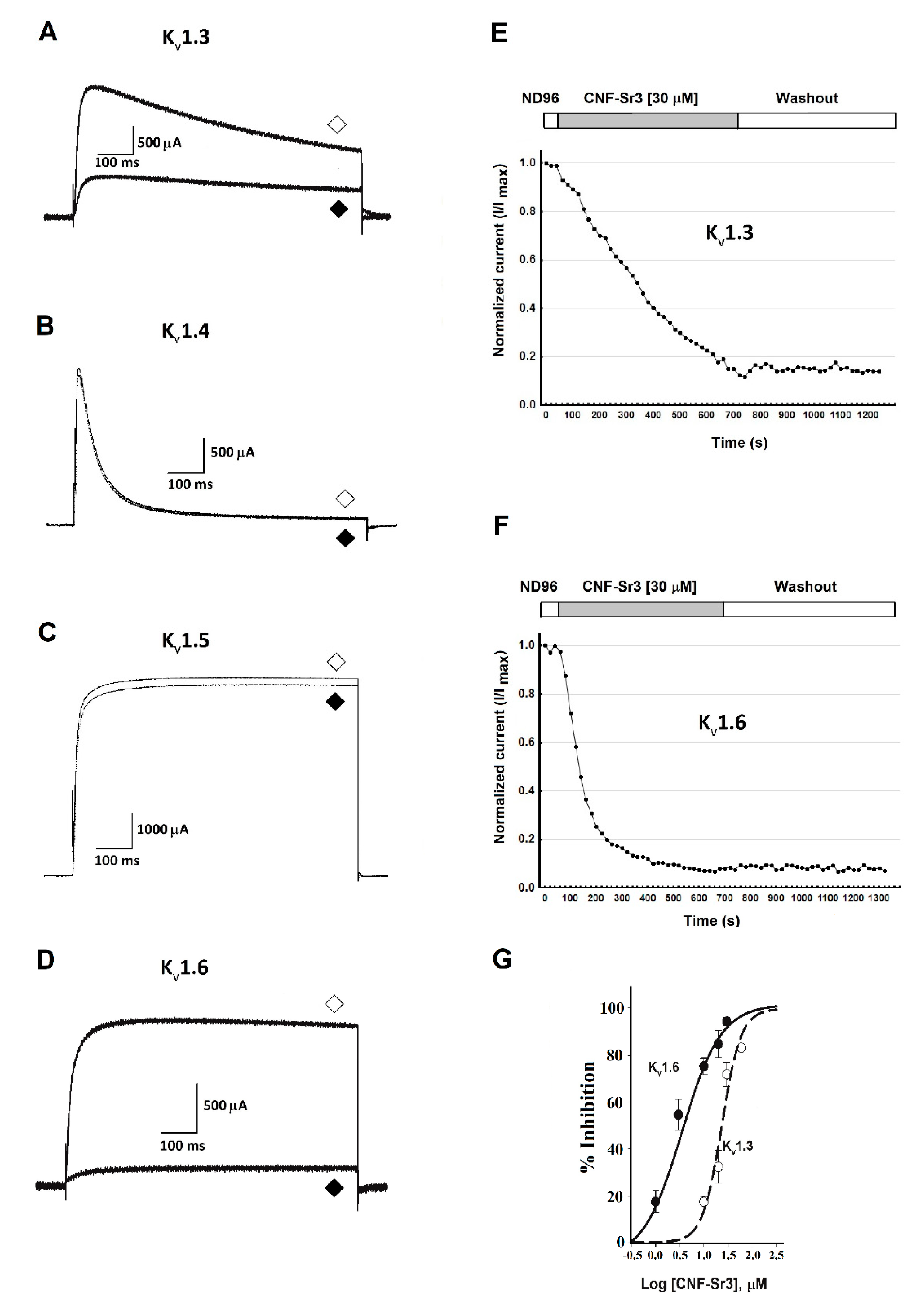{kind=link}
Abstract
:1. Introduction
2. Results
Evaluation of CNF-Sr3 on Kv1 channels Expressed in Oocytes
3. Discussion
4. Materials and Methods
4.1. Synthetic CNF-Sr3
4.2. Electrophysiology Assay in Oocytes
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Campos-Lira, E.; Carrillo, E.; Aguilar, M.B.; Gajewiak, J.; Gómez-Lagunas, F.; López-Vera, E. Conorfamide-Sr3, a Structurally Novel Specific Inhibitor of the Shaker K+ Channel. Toxicon 2017, 138, 53–58. [Google Scholar] [CrossRef]
- Puillandre, N.; Duda, T.F.; Meyer, C.; Olivera, B.M.; Bouchet, P. One, four or 100 genera? A New Classification of the Cone Snails. J. Molluscan Stud. 2015, 81, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Terlau, H.; Olivera, B.M. Conus Venoms: A Rich Source of Novel Ion Channel-Targeted Peptides. Physiol. Rev. 2004, 84, 41–68. [Google Scholar] [CrossRef] [Green Version]
- Akondi, K.B.; Muttenthaler, M.; Dutertre, S.; Kaas, Q.; Craik, D.J.; Lewis, R.J.; Alewood, P.F. Discovery, Synthesis, and Structure–Activity Relationships of Conotoxins. Chem. Rev. 2014, 114, 5815–5847. [Google Scholar] [CrossRef]
- Lebbe, E.K.M.; Tytgat, J. In the picture: Disulfide-poor conopeptides, a class of pharmacologically interesting compounds. J. Venom. Anim. Toxins Incl. Trop. Dis. 2016, 22, 1–15. [Google Scholar] [CrossRef] [Green Version]
- López-Vera, E.; Aguilar, M.B.; Heimer de la Cotera, E.P. FMRFamide and Related Peptides in the Phylum Mollusca. Peptides 2008, 29, 310–317. [Google Scholar] [CrossRef]
- Maillo, M.; Aguilar, M.B.; Lopéz-Vera, E.; Craig, A.G.; Bulaj, G.; Olivera, B.M.; Heimer De La Cotera, E.P. Conorfamide, a Conus Venom Peptide Belonging to the RFamide Family of Neuropeptides. Toxicon 2002, 40, 401–407. [Google Scholar] [CrossRef]
- Aguilar, M.B.; Luna-Ramírez, K.S.; Echeverría, D.; Falcón, A.; Olivera, B.M.; Heimer de la Cotera, E.P.; Maillo, M. Conorfamide-Sr2, a gamma-carboxyglutamate-containing FMRFamide-related peptide from the Venom of Conus spurius with activity in mice and mollusks. Peptides 2008, 29, 186–195. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, W.D.; Trout, W.E. The Behavior of Four Neurological Mutants of Drosophila. Genetics 1969, 61, 399–409. [Google Scholar]
- Kamb, A.; Iverson, L.E.; Tanouye, M.A. Molecular characterization of Shaker, a Drosophila gene that encodes a potassium channel. Cell 1987, 50, 405–413. [Google Scholar] [CrossRef]
- Swanson, R.; Marshall, J.; Smith, J.S.; Williams, J.B.; Boyle, M.B.; Folander, K.; Luneau, C.J.; Antanavage, J.; Oliva, C.; Buhrow, S.A.; et al. Cloning and expression of cDNA and genomic clones encoding three delayed rectifier potassium channels in rat brain. Neuron 1990, 4, 929–939. [Google Scholar] [CrossRef]
- Papazian, D.M.; Schwarz, T.L.; Tempel, B.L.; Jan, Y.N.; Jan, L.Y. Cloning of genomic and complementary DNA from Shaker, a putative potassium channel gene from Drosophila. Science 1987, 237, 749–753. [Google Scholar] [CrossRef]
- Roberds, S.L.; Tamkun, M.M. Cloning and tissue-specific expression of five voltage-gated potassium channel cDNAs expressed in rat heart. Proc. Natl. Acad. Sci. USA 2006, 88, 1798–1802. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Jen, J.C.; Lee, H.; Baloh, R.W.; Nelson, S.F.; Sabatti, C. A novel mutation in KCNA1 causes episodic ataxia without myokymia. Hum. Mutat. 2004, 24, 536. [Google Scholar]
- Frolov, R.V.; Bagati, A.; Casino, B.; Singh, S. Potassium Channels in Drosophila: Historical Breakthroughs, Significance, and Perspectives. J. Neurogenet. 2012, 26, 275–290. [Google Scholar] [CrossRef] [PubMed]
- Martel, P.; Leo, D.; Fulton, S.; Bérard, M.; Trudeau, L.E. Role of Kv1 Potassium Channels in Regulating Dopamine Release and Presynaptic D2 Receptor Function. PLoS ONE 2011, 6, e20402. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, S.; Baba, A.; Kaida, K.; Utsugisawa, K.; Kita, Y.; Tsugawa, J.; Ogawa, G.; Nagane, Y.; Kuwana, M.; Suzuki, N. Cardiac involvements in myasthenia gravis associated with anti-Kv1.4 antibodies. Eur. J. Neurol. 2014, 21, 223–230. [Google Scholar] [CrossRef]
- Ding, W.G.; Xie, Y.; Toyoda, F.; Matsuura, H. Improved Functional Expression of Human Cardiac Kv1.5 Channels and Trafficking-Defective Mutants by Low Temperature Treatment. PLoS ONE 2014, 9, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Nattel, S.; Yue, L.; Wang, Z. Cardiac Ultrarapid Delayed Rectifiers. Cell. Physiol. Biochem. 1999, 9, 217–226. [Google Scholar] [CrossRef]
- Comes, N.; Serrano-Albarrás, A.; Capera, J.; Serrano-Novillo, C.; Condom, E.; y Cajal, S.R.; Ferreres, J.C.; Felipe, A. Involvement of potassium channels in the progression of cancer to a more malignant phenotype. Biochim. Biophys. Acta 2015, 1848, 2477–2492. [Google Scholar] [CrossRef] [Green Version]
- Shon, K.J.; Stocker, M.; Terlau, H.; Stühmer, W.; Jacobsen, R.; Walker, C.; Grilley, M.; Watkins, M.; Hillyard, D.R.; Gray, W.R.; et al. κ-Conotoxin PVIIA is a peptide inhibiting the shaker K+ Channel. J. Biol. Chem. 1998, 273, 33–38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahdavi, S.; Kuyucak, S. Why the Drosophila Shaker K+ Channel Is Not a Good Model for Ligand Binding to Voltage-Gated Kv1 Channels. Biochemistry 2013, 52, 1631–1640. [Google Scholar] [CrossRef]
- Imperial, J.S.; Bansal, P.S.; Alewood, P.F.; Daly, N.L.; Craik, D.J.; Sporning, A.; Terlau, H.; López-Vera, E.; Bandyopadhyay, P.K.; Olivera, B.M. A Novel Conotoxin Inhibitor of Kv1.6 Channel and nAChR Subtypes Defines a New Superfamily of Conotoxins. Biochemistry 2006, 45. [Google Scholar] [CrossRef] [PubMed]
- Lewis, R.J.; Dutertre, S.; Vetter, I.; Christie, M.J. Conus Venom Peptide Pharmacology. Pharmacol. Rev. 2012, 64, 259–298. [Google Scholar] [CrossRef]
- Sudarslal, S.; Singaravadivelan, G.; Ramasamy, P.; Ananda, K.; Sarma, S.P.; Sikdar, S.K.; Krishnan, K.S.; Balaram, P. A novel 13 residue acyclic peptide from the marine snail, Conus monile, targets potassium channels. Biochem. Biophys. Res. Commun. 2004, 317, 682–688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imperial, J.S.; Chen, P.; Sporning, A.; Terlau, H.; Daly, N.L.; Craik, D.J.; Alewood, P.F.; Olivera, B.M. Tyrosine-rich Conopeptides Affect Voltage-gated K+ Channels. J. Biol. Chem. 2008, 283, 23026–23032. [Google Scholar] [CrossRef] [Green Version]
- Terlau, H.; Shon, K.J.; Grilley, M.; Stocker, M.; Stühmer, W.; Olivera, B.M. Strategy for rapid immobilization of prey by a fish-hunting marine snail. Nature 1996, 381, 148–151. [Google Scholar] [CrossRef]
- Craig, A.G.; Zafaralla, G.; Cruz, L.J.; Santos, A.D.; Hillyard, D.R.; Dykert, J.; Rivier, J.E.; Gray, W.R.; Imperial, J.; DelaCruz, R.G.; et al. An O-Glycosylated Neuroexcitatory Conus Peptide. Biochemistry 1998, 37, 16019–16025. [Google Scholar] [CrossRef]
- Ferber, M.; Sporning, A.; Jeserich, G.; DeLaCruz, R.; Watkins, M.; Olivera, B.M.; Terlau, H. A Novel Conus Peptide Ligand for K+ Channels. J. Biol. Chem. 2003, 278, 2177–2183. [Google Scholar] [CrossRef] [Green Version]
- Kauferstein, S.; Huys, I.; Lamthanh, H.; Stöcklin, R.; Sotto, F.; Menez, A.; Tytgat, J.; Mebs, D. A novel conotoxin inhibiting vertebrate voltage-sensitive potassium channels. Toxicon 2003, 42, 43–52. [Google Scholar] [CrossRef]
- Aguilar, M.B.; Pérez-Reyes, L.I.; López, Z.; de la Cotera, E.P.H.; Falcón, A.; Ayala, C.; Galván, M.; Salvador, C.; Escobar, L.I. Peptide Sr11a from Conus spurius is a novel peptide blocker for Kv1 potassium channels. Peptides 2010, 31, 1287–1291. [Google Scholar] [CrossRef]
- Al-Sabi, A.; Lennartz, D.; Ferber, M.; Gulyas, J.; Rivier, J.E.F.; Olivera, B.M.; Carlomagno, T.; Terlau, H. kM-Conotoxin RIIIK, Structural and Functional Novelty in a K+ Channel Antagonist. Biochemistry 2004, 43, 8625–8635. [Google Scholar] [CrossRef]

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Vera, E.; Martínez-Hernández, L.; Aguilar, M.B.; Carrillo, E.; Gajewiak, J. Studies of Conorfamide-Sr3 on Human Voltage-Gated Kv1 Potassium Channel Subtypes. Mar. Drugs 2020, 18, 425. https://doi.org/10.3390/md18080425
López-Vera E, Martínez-Hernández L, Aguilar MB, Carrillo E, Gajewiak J. Studies of Conorfamide-Sr3 on Human Voltage-Gated Kv1 Potassium Channel Subtypes. Marine Drugs. 2020; 18(8):425. https://doi.org/10.3390/md18080425
Chicago/Turabian StyleLópez-Vera, Estuardo, Luis Martínez-Hernández, Manuel B. Aguilar, Elisa Carrillo, and Joanna Gajewiak. 2020. "Studies of Conorfamide-Sr3 on Human Voltage-Gated Kv1 Potassium Channel Subtypes" Marine Drugs 18, no. 8: 425. https://doi.org/10.3390/md18080425
APA StyleLópez-Vera, E., Martínez-Hernández, L., Aguilar, M. B., Carrillo, E., & Gajewiak, J. (2020). Studies of Conorfamide-Sr3 on Human Voltage-Gated Kv1 Potassium Channel Subtypes. Marine Drugs, 18(8), 425. https://doi.org/10.3390/md18080425