
Exogenous Antioxidants Improve the Accumulation of Saturated and Polyunsaturated Fatty Acids in Schizochytrium sp. PKU#Mn4


 ,
, 
 and
and 
Abstract
:1. Introduction
2. Results
2.1. Screening Potential Antioxidants
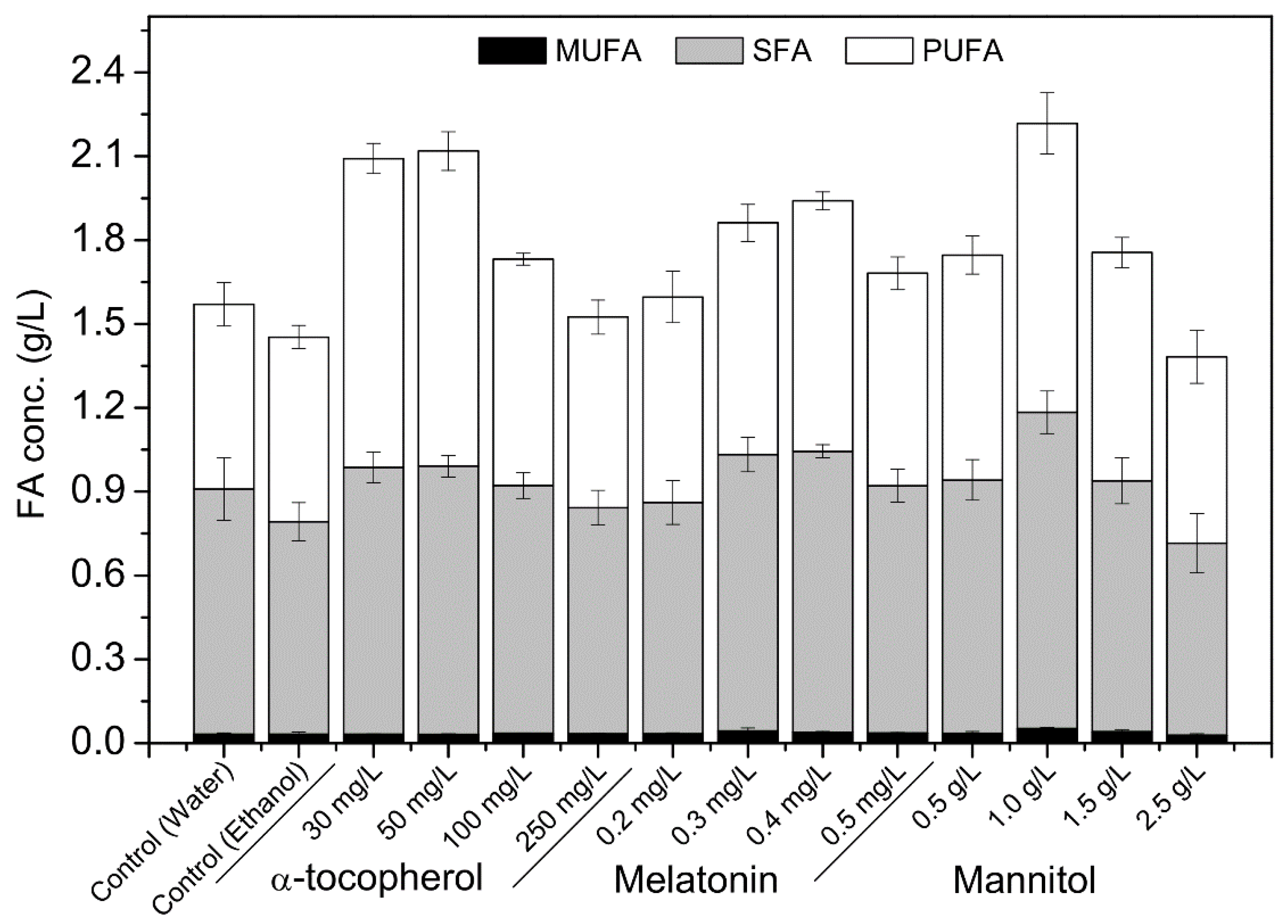
2.2. Effects of Different Concentrations of Single Antioxidants
2.3. Interaction Effects of Ascorbic Acid and Mannitol in Batch Culture
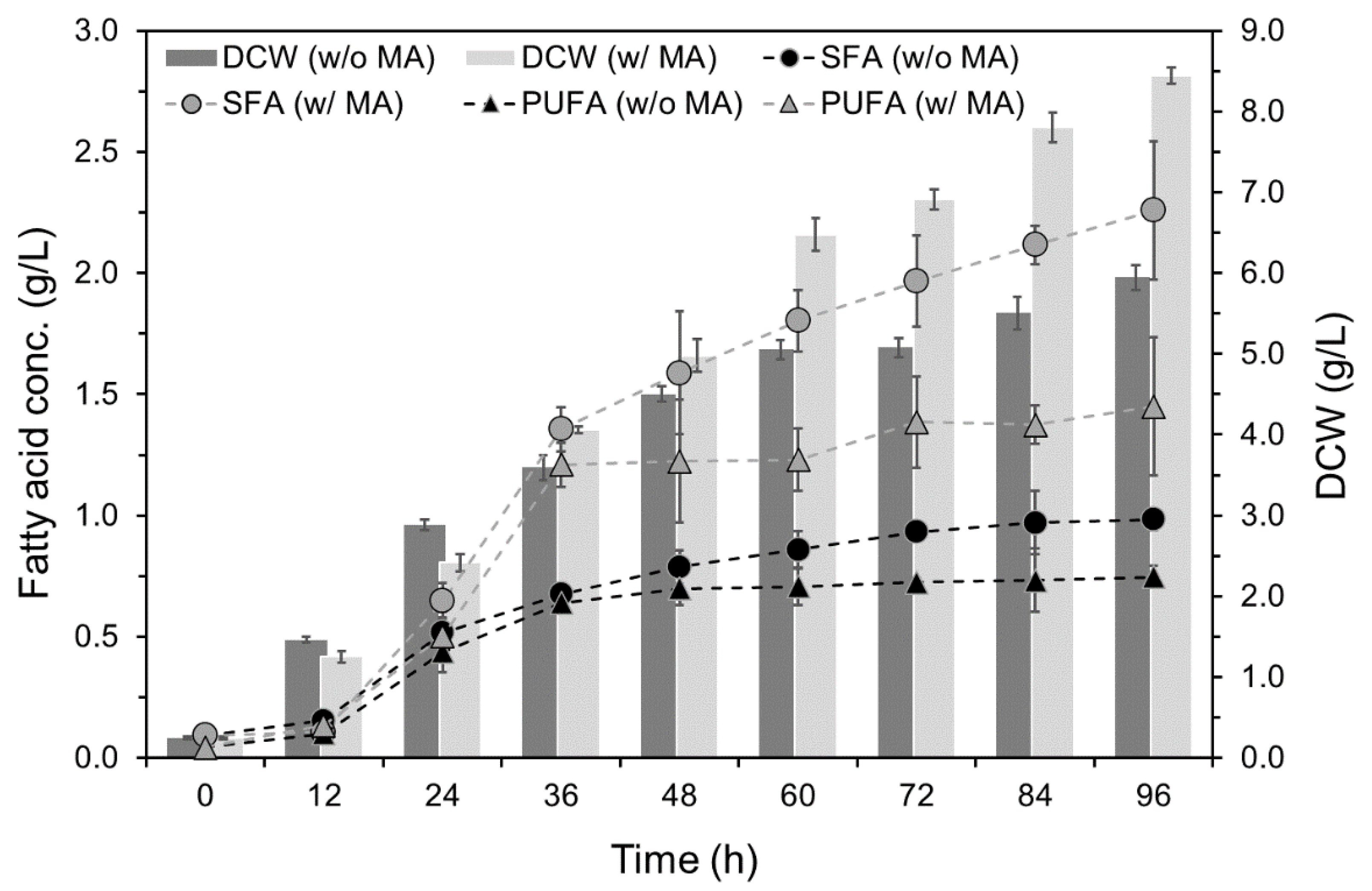
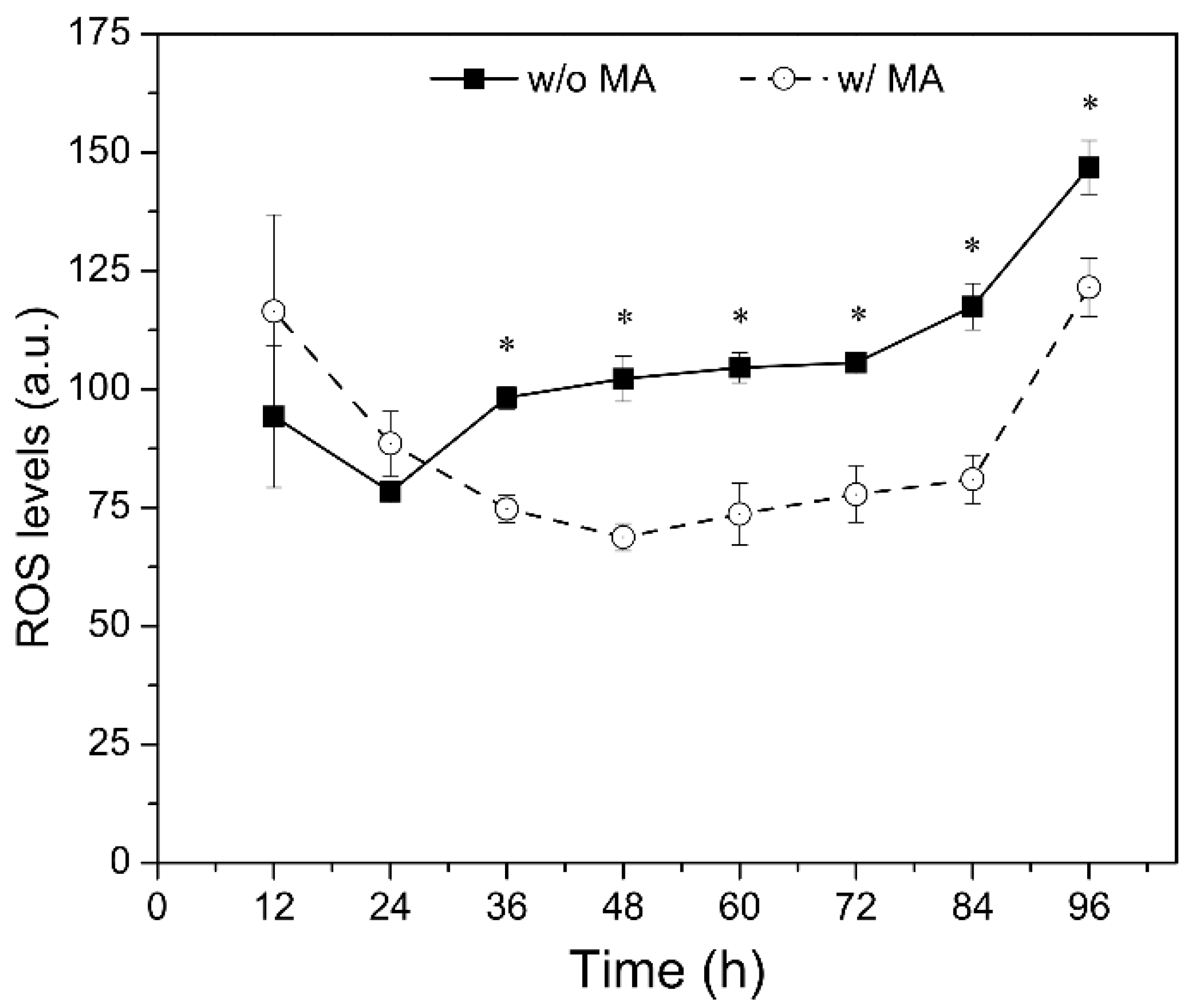
2.4. Effects of Mannitol and Ascorbic Acid Supplemention on Fed-Batch Culture
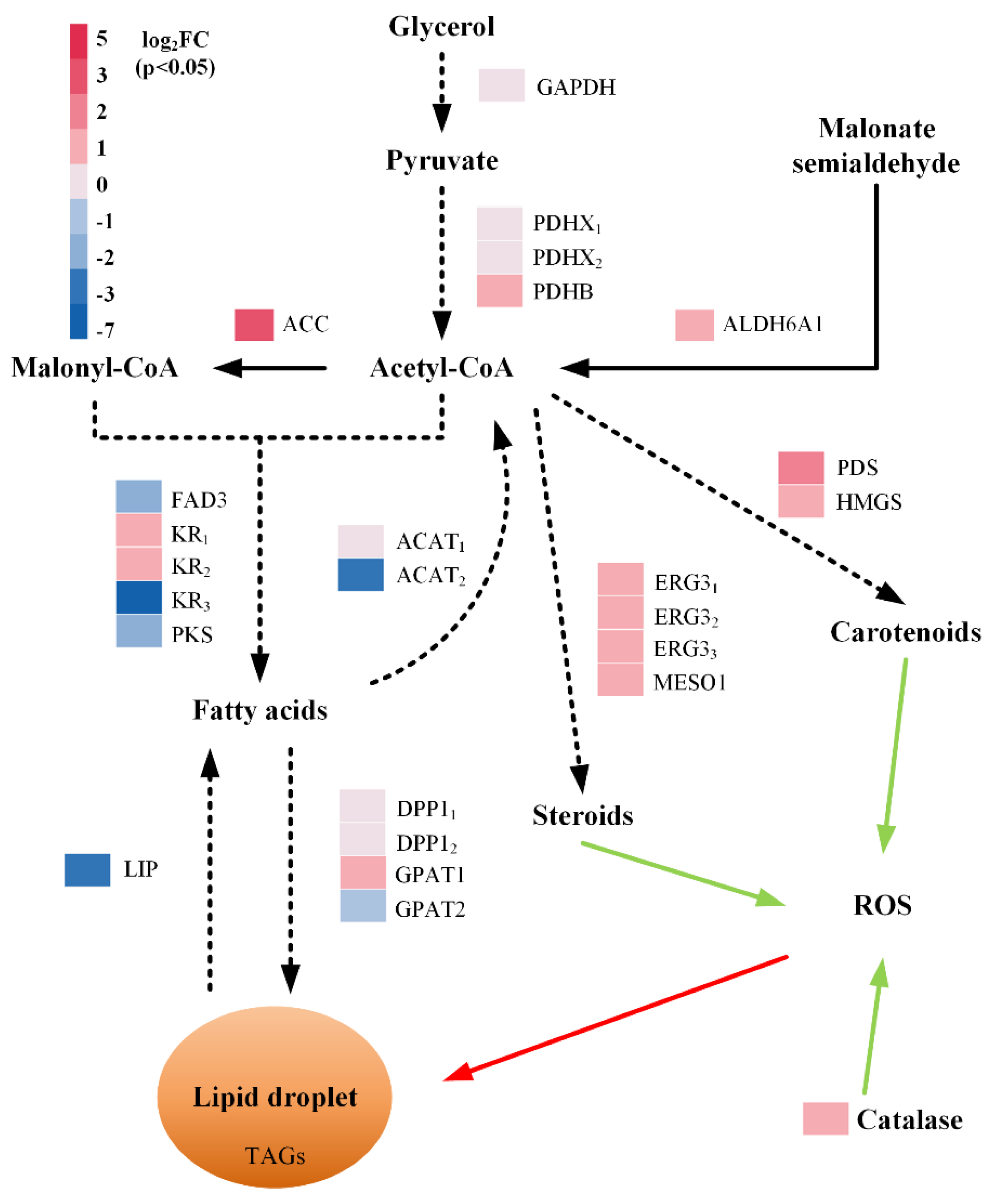
2.5. Effects of Mannitol and Ascorbic Acid Supplementation at the Transcriptome Level
3. Discussion
3.1. Antioxidants Affect Growth and Fatty Acids Accumulation
3.2. Exogenous Antioxidants Trigger Differential Expression of Lipid Biosynthetic Genes
4. Materials and Methods
4.1. Microorganism and Batch Fermentation
4.2. Experimental Design and Statistical Analyses
4.3. Quantification of Dry Cell Weight and Fatty Acids
4.4. Determination of Intracellular ROS
4.5. Fed-Batch Fermentation
4.6. Transcriptome Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gupta, A.; Barrow, C.J.; Puri, M. Omega-3 biotechnology: Thraustochytrids as a novel source of omega-3 oils. Biotechnol. Adv. 2012, 30, 1733–1745. [Google Scholar] [CrossRef] [PubMed]
- Du, F.; Wang, Y.Z.; Xu, Y.S.; Shi, T.Q.; Liu, W.Z.; Sun, X.M.; Huang, H. Biotechnological production of lipid and terpenoid from thraustochytrids. Biotechnol. Adv. 2021, 48, 107725. [Google Scholar] [CrossRef] [PubMed]
- Chandrasekaran, K.; Roy, R.K.; Chadha, A. Docosahexaenoic acid production by a novel high yielding strain of Thraustochytrium sp. of Indian origin: Isolation and bioprocess optimization studies. Algal Res. 2018, 32, 93–100. [Google Scholar] [CrossRef]
- Leyton, A.; Flores, L.; Shene, C.; Chisti, Y.; Larama, G.; Asenjo, J.A.; Armenta, R.E. Antarctic Thraustochytrids as Sources of Carotenoids and High-Value Fatty Acids. Mar. Drugs 2021, 19, 386. [Google Scholar] [CrossRef]
- Zhang, A.; Xie, Y.; He, Y.; Wang, W.; Sen, B.; Wang, G. Bio-based squalene production by Aurantiochytrium sp. through optimization of culture conditions, and elucidation of the putative biosynthetic pathway genes. Bioresour. Technol. 2019, 287, 121415. [Google Scholar] [CrossRef]
- Aasen, I.M.; Ertesvåg, H.; Heggeset, T.M.B.; Liu, B.; Brautaset, T.; Vadstein, O.; Ellingsen, T.E. Thraustochytrids as production organisms for docosahexaenoic acid (DHA), squalene, and carotenoids. Appl. Microbiol. Biotechnol. 2016, 100, 4309–4321. [Google Scholar] [CrossRef]
- Guo, D.-S.; Ji, X.-J.; Ren, L.-J.; Li, G.-L.; Sun, X.-M.; Chen, K.-Q.; Gao, S.; Huang, H. Development of a scale-up strategy for fermentative production of docosahexaenoic acid by Schizochytrium sp. Chem. Eng. Sci. 2018, 176, 600–608. [Google Scholar] [CrossRef]
- Huang, T.Y.; Lu, W.C.; Chu, I.M. A fermentation strategy for producing docosahexaenoic acid in Aurantiochytrium limacinum SR21 and increasing C22:6 proportions in total fatty acid. Bioresour. Technol. 2012, 123, 8–14. [Google Scholar] [CrossRef]
- Li, J.; Liu, R.; Chang, G.; Li, X.; Chang, M.; Liu, Y.; Jin, Q.; Wang, X. A strategy for the highly efficient production of docosahexaenoic acid by Aurantiochytrium limacinum SR21 using glucose and glycerol as the mixed carbon sources. Bioresour. Technol. 2015, 177, 51–57. [Google Scholar] [CrossRef]
- Ling, X.; Guo, J.; Liu, X.; Zhang, X.; Wang, N.; Lu, Y.; Ng, I.S. Impact of carbon and nitrogen feeding strategy on high production of biomass and docosahexaenoic acid (DHA) by Schizochytrium sp. LU310. Bioresour. Technol. 2015, 184, 139–147. [Google Scholar] [CrossRef]
- Qu, L.; Ji, X.J.; Ren, L.J.; Nie, Z.K.; Feng, Y.; Wu, W.J.; Ouyang, P.K.; Huang, H. Enhancement of docosahexaenoic acid production by Schizochytrium sp. using a two-stage oxygen supply control strategy based on oxygen transfer coefficient. Lett. Appl. Microbiol. 2011, 52, 22–27. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Ji, X.; Huang, H.; Qu, L.; Feng, Y.; Tong, Q.; Ouyang, P. Development of a Stepwise Aeration Control Strategy for Efficient Docosahexaenoic Acid Production by Schizochytrium sp. Appl. Microbiol. Biotechnol. 2010, 87, 1649–1656. [Google Scholar] [CrossRef] [PubMed]
- Heggeset, T.M.B.; Ertesvåg, H.; Liu, B.; Ellingsen, T.E.; Vadstein, O.; Aasen, I.M. Lipid and DHA-production in Aurantiochytrium sp.—Responses to nitrogen starvation and oxygen limitation revealed by analyses of production kinetics and global transcriptomes. Sci. Rep. 2019, 9, 19470. [Google Scholar] [CrossRef] [PubMed]
- Jakobsen, A.N.; Aasen, I.M.; Josefsen, K.D.; Strøm, A.R. Accumulation of Docosahexaenoic Acid-Rich Lipid in Thraustochytrid Aurantiochytrium sp. strain T66: Effects of N and P Starvation and O2 Limitation. Appl. Microbiol. Biotechnol. 2008, 80, 297–306. [Google Scholar] [CrossRef]
- Chodchoey, K.; Verduyn, C. Growth, fatty acid profile in major lipid classes and lipid fluidity of Aurantiochytrium mangrovei Sk-02 as a function of growth temperature. Braz. J. Microbiol. 2012, 43, 187–200. [Google Scholar] [CrossRef] [Green Version]
- Taoka, Y.; Nagano, N.; Okita, Y.; Izumida, H.; Sugimoto, S.; Hayashi, M. Influences of Culture Temperature on the Growth, Lipid Content and Fatty Acid Composition of Aurantiochytrium sp. Strain mh0186. Mar. Biotechnol. 2009, 11, 368–374. [Google Scholar] [CrossRef]
- Shi, K.; Gao, Z.; Shi, T.-Q.; Song, P.; Ren, L.-J.; Huang, H.; Ji, X.-J. Reactive Oxygen Species-Mediated Cellular Stress Response and Lipid Accumulation in Oleaginous Microorganisms: The State of the Art and Future Perspectives. Front. Microbiol. 2017, 8, 793. [Google Scholar] [CrossRef]
- Sajjadi, B.; Chen, W.-Y.; Raman, A.A.A.; Ibrahim, S. Microalgae lipid and biomass for biofuel production: A comprehensive review on lipid enhancement strategies and their effects on fatty acid composition. Renew. Sustain. Energy Rev. 2018, 97, 200–232. [Google Scholar] [CrossRef]
- Sun, X.M.; Ren, L.J.; Ji, X.J.; Huang, H. Enhancing biomass and lipid accumulation in the microalgae Schizochytrium sp. by addition of fulvic acid and EDTA. AMB Express 2018, 8, 150. [Google Scholar] [CrossRef] [Green Version]
- Zhang, S.; He, Y.; Sen, B.; Chen, X.; Xie, Y.; Keasling, J.D.; Wang, G. Alleviation of reactive oxygen species enhances PUFA accumulation in Schizochytrium sp. through regulating genes involved in lipid metabolism. Metab. Eng. Commun. 2018, 6, 39–48. [Google Scholar] [CrossRef]
- Kitchener, R.L.; Grunden, A.M. Methods for enhancing cyanobacterial stress tolerance to enable improved production of biofuels and industrially relevant chemicals. Appl. Microbiol. Biotechnol. 2018, 102, 1617–1628. [Google Scholar] [CrossRef]
- Zhang, S.; He, Y.; Sen, B.; Wang, G. Reactive oxygen species and their applications toward enhanced lipid accumulation in oleaginous microorganisms. Bioresour. Technol. 2020, 307, 123234. [Google Scholar] [CrossRef]
- Santos-Sánchez, N.F.; Salas-Coronado, R.; Villanueva-Cañongo, C.; Hernández-Carlos, B. Antioxidant Compounds and Their Antioxidant Mechanism. In Antioxidants; Shalaby, E., Ed.; IntechOpen: London, UK, 2019. [Google Scholar]
- Ahmad, P.; Jaleel, C.A.; Salem, M.A.; Nabi, G.; Sharma, S. Roles of enzymatic and nonenzymatic antioxidants in plants during abiotic stress. Crit. Rev. Biotechnol. 2010, 30, 161–175. [Google Scholar] [CrossRef]
- Zhao, Y.; Li, D.; Xu, J.-W.; Zhao, P.; Li, T.; Ma, H.; Yu, X. Melatonin enhances lipid production in Monoraphidium sp. QLY-1 under nitrogen deficiency conditions via a multi-level mechanism. Bioresour. Technol. 2018, 259, 46–53. [Google Scholar] [CrossRef]
- Liu, B.; Liu, J.; Sun, P.; Ma, X.; Jiang, Y.; Chen, F. Sesamol Enhances Cell Growth and the Biosynthesis and Accumulation of Docosahexaenoic Acid in the Microalga Crypthecodinium cohnii. J. Agric. Food. Chem. 2015, 63, 5640–5645. [Google Scholar] [CrossRef]
- Ren, L.; Sun, X.; Ji, X.; Chen, S.; Guo, D.; Huang, H. Enhancement of Docosahexaenoic Acid Synthesis by Manipulation of Antioxidant Capacity and Prevention of Oxidative Damage in Schizochytrium sp. Bioresour. Technol. 2017, 223, 141–148. [Google Scholar] [CrossRef] [Green Version]
- Gaffney, M.; O’Rourke, R.; Murphy, R. Manipulation of fatty acid and antioxidant profiles of the microalgae Schizochytrium sp. through flaxseed oil supplementation. Algal Res. 2014, 6, 195–200. [Google Scholar] [CrossRef]
- Singh, D.; Mathur, A.S.; Tuli, D.K.; Puri, M.; Barrow, C.J. Propyl gallate and butylated hydroxytoluene influence the accumulation of saturated fatty acids, omega-3 fatty acid and carotenoids in thraustochytrids. J. Funct. Foods 2015, 15, 186–192. [Google Scholar] [CrossRef]
- Zhao, Y.; Yue, C.; Ding, W.; Li, T.; Xu, J.-W.; Zhao, P.; Ma, H.; Yu, X. Butylated hydroxytoluene induces astaxanthin and lipid production in Haematococcus pluvialis under high-light and nitrogen-deficiency conditions. Bioresour. Technol. 2018, 266, 315–321. [Google Scholar] [CrossRef]
- Na, C.; Junmu, X.; Yongjie, F.; Yongteng, Z.; Xuya, Y.; Jun-Wei, X.; Tao, L.; Peng, Z. Antioxidants enhance lipid productivity in Heveochlorella sp. Yu. Algal Res. 2021, 55, 102235. [Google Scholar]
- Li, D.; Zhao, Y.; Ding, W.; Zhao, P.; Xu, J.W.; Li, T.; Ma, H.; Yu, X. A strategy for promoting lipid production in green microalgae Monoraphidium sp. QLY-1 by combined melatonin and photoinduction. Bioresour. Technol. 2017, 235, 104–112. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.; Zhao, P.; Peng, J.; Zhao, Y.; Xu, J.-W.; Li, T.; Reiter, R.J.; Ma, H.; Yu, X. Melatonin enhances astaxanthin accumulation in the green microalga Haematococcus pluvialis by mechanisms possibly related to abiotic stress tolerance. Algal Res. 2018, 33, 256–265. [Google Scholar] [CrossRef]
- Davis, M.S.; Solbiati, J.; Cronan, J.E., Jr. Overproduction of acetyl-CoA carboxylase activity increases the rate of fatty acid biosynthesis in Escherichia coli. J. Biol. Chem. 2000, 275, 28593–28598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.; Pei, G.; Diao, J.; Chen, Z.; Liu, L.; Chen, L.; Zhang, W. Screening and transcriptomic analysis of Crypthecodinium cohnii mutants with high growth and lipid content using the acetyl-CoA carboxylase inhibitor sethoxydim. Appl. Microbiol. Biotechnol. 2017, 101, 6179–6191. [Google Scholar] [CrossRef]
- Fathy, W.; Essawy, E.; Tawfik, E.; Khedr, M.; Abdelhameed, M.S.; Hammouda, O.; Elsayed, K. Recombinant overexpression of the Escherichia coli acetyl-CoA carboxylase gene in Synechocystis sp. boosts lipid production. J. Basic Microbiol. 2021, 61, 330–338. [Google Scholar] [CrossRef]
- Che, R.; Huang, L.; Xu, J.-W.; Zhao, P.; Li, T.; Ma, H.; Yu, X. Effect of fulvic acid induction on the physiology, metabolism, and lipid biosynthesis-related gene transcription of Monoraphidium sp. FXY-10. Bioresour. Technol. 2017, 227, 324–334. [Google Scholar] [CrossRef]
- Liu, Y.; Singh, P.; Sun, Y.; Luan, S.; Wang, G. Culturable Diversity and Biochemical Features of Thraustochytrids from Coastal Waters of Southern China. Appl. Microbiol. Biotechnol. 2014, 98, 3241–3255. [Google Scholar] [CrossRef]
- Wang, Q.; Ye, H.; Xie, Y.; He, Y.; Sen, B.; Wang, G. Culturable Diversity and Lipid Production Profile of Labyrinthulomycete Protists Isolated from Coastal Mangrove Habitats of China. Mar. Drugs 2019, 17, 268. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Ye, H.; Sen, B.; Xie, Y.; He, Y.; Park, S.; Wang, G. Improved production of docosahexaenoic acid in batch fermentation by newly-isolated strains of Schizochytrium sp. and Thraustochytriidae sp. through bioprocess optimization. Synth. Sys. Biotechnol. 2018, 3, 121–129. [Google Scholar] [CrossRef]
- Tjørve, K.M.C.; Tjørve, E. The use of Gompertz models in growth analyses, and new Gompertz-model approach: An addition to the Unified-Richards family. PLoS ONE 2017, 12, e0178691. [Google Scholar] [CrossRef]
- Kim, D.; Langmead, B.; Salzberg, S.L. HISAT: A fast spliced aligner with low memory requirements. Nat. Methods 2015, 12, 357–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pertea, M.; Pertea, G.M.; Antonescu, C.M.; Chang, T.C.; Mendell, J.T.; Salzberg, S.L. StringTie enables improved reconstruction of a transcriptome from RNA-seq reads. Nat. Biotechnol. 2015, 33, 290–295. [Google Scholar] [CrossRef] [Green Version]
- Mortazavi, A.; Williams, B.A.; McCue, K.; Schaeffer, L.; Wold, B. Mapping and quantifying mammalian transcriptomes by RNA-Seq. Nat. Methods 2008, 5, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factor | Antioxidant | Low Level (−1) | High Level (+1) |
|---|---|---|---|
| A | Ascorbic acid | 9.0 g/L | 13.5 g/L |
| B | α-tocopherol | 0.50 g/L | 0.75 g/L |
| C | Tea extract | 0.50 g/L | 0.75 g/L |
| D | Melatonin | 0.25 mg/L | 0.375 mg/L |
| E | Mannitol | 1.0 g/L | 1.5 g/L |
| F | Sesamol | 70 mg/L | 105 mg/L |
| G | BHT | 2 mg/L | 3 mg/L |
| Source | p-Value | Source | p-Value |
|---|---|---|---|
| Response 1: DCW (g/L) | Response 2: TFA (mg/g DCW) | ||
| Model | 0.0447 | Model | 0.0180 |
| A: ascorbic acid | 0.3917 | A: ascorbic acid | 0.6802 |
| B: α-tocopherol | 0.0406 | B: α-tocopherol | 0.0203 |
| C: tea extract | 0.1094 | D: melatonin | 0.0206 |
| D: melatonin | 0.1397 | E: mannitol | 0.1194 |
| E: mannitol | 0.0141 | F: sesamol | 0.1643 |
| F: sesamol | 0.4890 | G: BHT | 0.6473 |
| AE | 0.1069 | AE | 0.0487 |
| AF | 0.1061 | AF | 0.0166 |
| AG | 0.0047 | ||
| BD | 0.0697 | ||
| Treatment | DCW (g/L) | MUFA (g/L) | SFA (g/L) | PUFA (g/L) | SFA/PUFA |
|---|---|---|---|---|---|
| Control | 5.89 ± 0.23 | 0.03 ± 0.00 | 0.88 ± 0.11 | 0.66 ± 0.08 | 1.32 |
| Mannitol (1 g/L) | 5.51 ± 0.33 | 0.04 ± 0.01 | 1.11 ± 0.17 | 0.97 ± 0.13 | 1.14 |
| Mannitol (1 g/L) + Ascorbic acid (1 g/L) | 8.49 ± 0.18 | 0.08 ± 0.01 | 2.26 ± 0.27 | 1.45 ± 0.04 | 1.56 |
| Mannitol (1 g/L) + Ascorbic acid (2 g/L) | 7.96 ± 0.28 | 0.06 ± 0.01 | 2.26 ± 0.07 | 1.19 ± 0.09 | 1.89 |
| Mannitol (1 g/L) + Ascorbic acid (3 g/L) | 6.79 ± 0.12 | 0.04 ± 0.00 | 1.87 ± 0.16 | 1.01 ± 0.14 | 1.85 |
| Mannitol (1 g/L) + Ascorbic acid (6 g/L) | 4.67 ± 0.33 | 0.04 ± 0.00 | 1.31 ± 0.19 | 0.73 ± 0.07 | 1.80 |
| Mannitol (1 g/L) + Ascorbic acid (9 g/L) | 3.38 ± 0.00 | 0.02 ± 0.00 | 0.79 ± 0.06 | 0.59 ± 0.02 | 1.35 |
| Mannitol (1 g/L) + Ascorbic acid (12 g/L) | 1.84 ± 0.03 | 0.01 ± 0.00 | 0.45 ± 0.02 | 0.34 ± 0.03 | 1.32 |
| Mannitol (1 g/L) + Ascorbic acid (15 g/L) | 1.22 ± 0.02 | 0.01 ± 0.00 | 0.29 ± 0.02 | 0.28 ± 0.03 | 1.07 |
| Dependent Variable | a (g/L) | R (g/L·h−1) | λ (h) | Residual Standard Error | |
|---|---|---|---|---|---|
| DCW | w/o MA | 5.799 *** | 0.102 *** | −1.791 | 0.2223 |
| w/MA | 9.276 *** | 0.118 *** | 3.478 | 0.2045 | |
| SFA | w/o MA | 0.977 *** | 0.021 *** | 1.763 | 0.0434 |
| w/MA | 2.191 *** | 0.047 *** | 9.832 * | 0.1017 | |
| PUFA | w/o MA | 0.729 *** | 0.030 *** | 9.069 *** | 0.0194 |
| w/MA | 1.369 *** | 0.056 ** | 13.723 ** | 0.0948 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Chen, X.; Sen, B.; Bai, M.; He, Y.; Wang, G. Exogenous Antioxidants Improve the Accumulation of Saturated and Polyunsaturated Fatty Acids in Schizochytrium sp. PKU#Mn4. Mar. Drugs 2021, 19, 559. https://doi.org/10.3390/md19100559
Zhang S, Chen X, Sen B, Bai M, He Y, Wang G. Exogenous Antioxidants Improve the Accumulation of Saturated and Polyunsaturated Fatty Acids in Schizochytrium sp. PKU#Mn4. Marine Drugs. 2021; 19(10):559. https://doi.org/10.3390/md19100559
Chicago/Turabian StyleZhang, Sai, Xiaohong Chen, Biswarup Sen, Mohan Bai, Yaodong He, and Guangyi Wang. 2021. "Exogenous Antioxidants Improve the Accumulation of Saturated and Polyunsaturated Fatty Acids in Schizochytrium sp. PKU#Mn4" Marine Drugs 19, no. 10: 559. https://doi.org/10.3390/md19100559
APA StyleZhang, S., Chen, X., Sen, B., Bai, M., He, Y., & Wang, G. (2021). Exogenous Antioxidants Improve the Accumulation of Saturated and Polyunsaturated Fatty Acids in Schizochytrium sp. PKU#Mn4. Marine Drugs, 19(10), 559. https://doi.org/10.3390/md19100559