
Anti-Inflammatory Activity of Fucoidan Extracts In Vitro

 ,
,  , , , ,
, , , ,  , ,
, ,  and
and
Abstract
:1. Introduction
2. Results
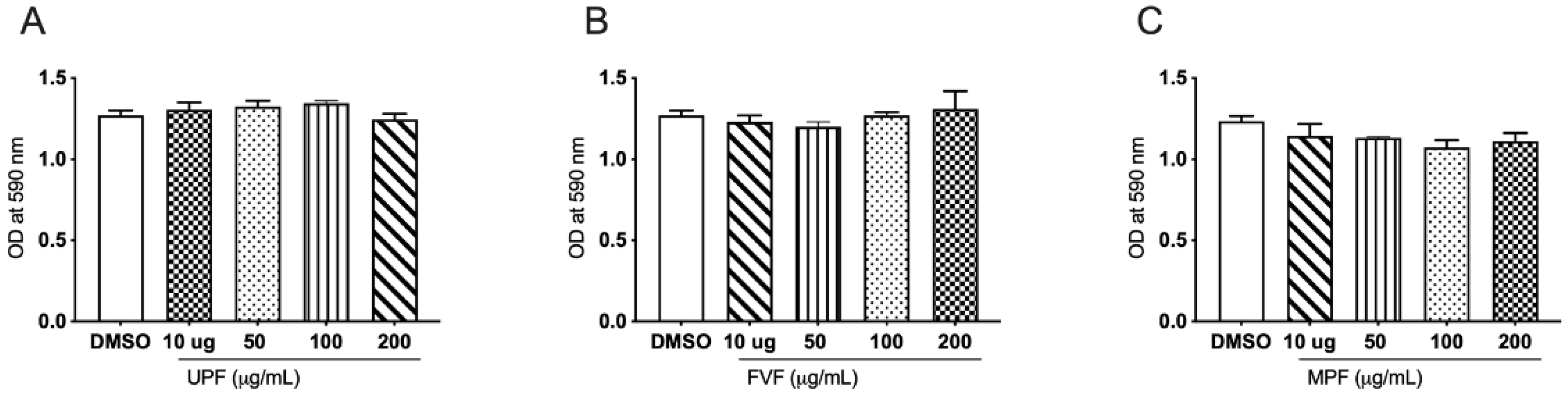
2.1. Effect of Fucoidan Extracts on Cell Viability
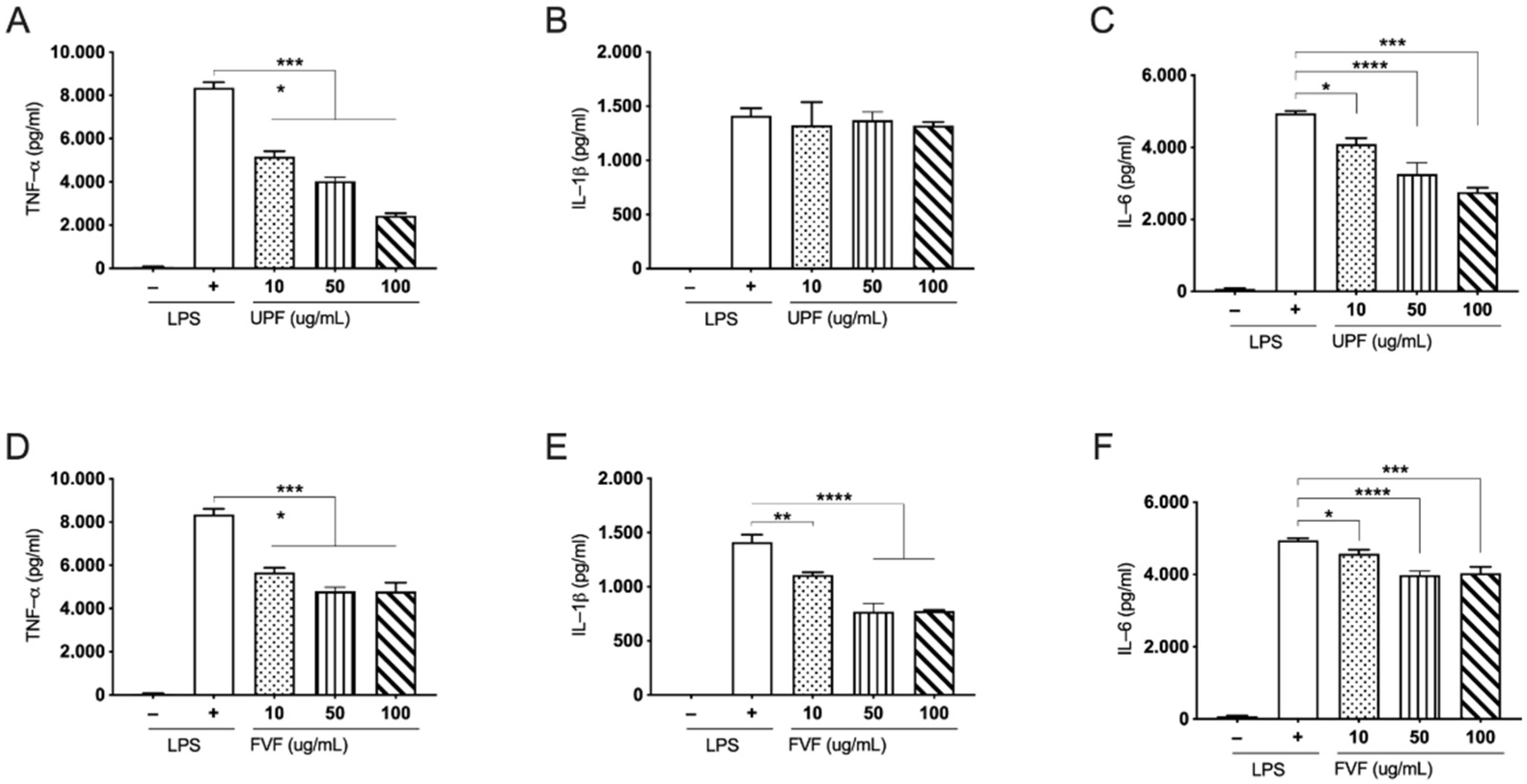
2.2. Effects of UPF and FVF in LPS Induced TNF-α, IL-1β, and IL-6 in Human PBMCs
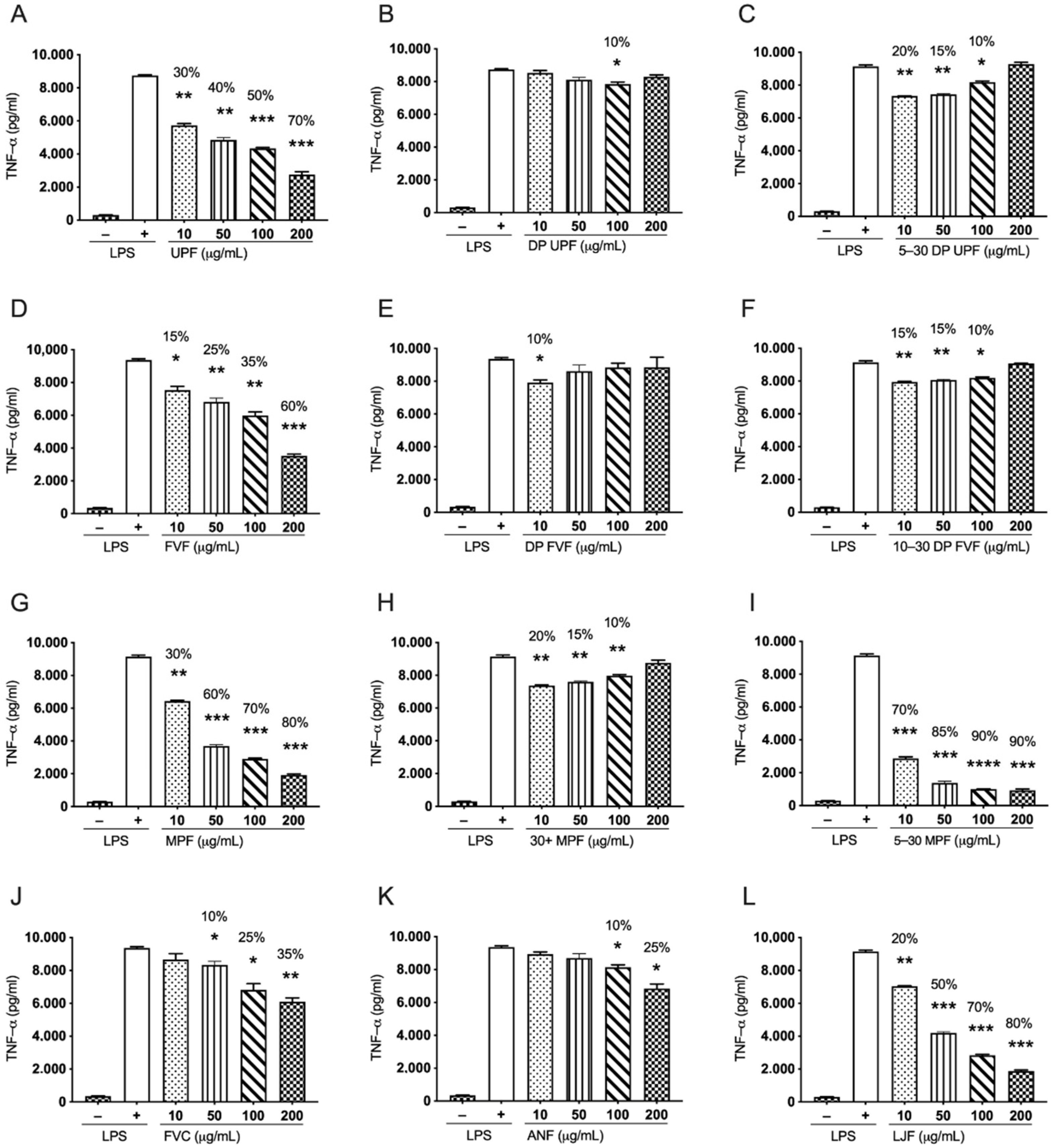
2.3. Effect of Fucoidan Extracts on LPS Induced TNF-α Pro-Inflammatory Cytokine in Human THP-1 Monocytes
2.3.1. Undaria pinnatifida Fucoidan (UPF) and Its Derived Extracts Suppress LPS Induced TNF-α in Human THP-1 Cells
2.3.2. Fucus vesiculosus Fucoidan (FVF) and Its Derived Extracts Suppress LPS Stimulated TNF-α in Human THP-1 Cells
2.3.3. Macrocystis pyrifera Fucoidan (MPF) and Its Derived Extracts Suppress LPS Stimulated TNF-α in Human THP-1 Cells
2.3.4. Nutritional Grade Fucoidan from Fucus vesiculosus with High Polyphenol Content (FVC), Ascophyllum nodosum (ANF), and Laminaria japonica (LJF) Inhibit LPS Induced TNF-α in Human THP-1 Cells in a Dose-Dependent Manner
2.4. Effect of Fucoidan Extracts on LPS Induced Pro-Inflammatory Cytokine IL-1β in Human THP-1 Monocytes
2.4.1. Undaria pinnatifida Fucoidan (UPF) and Its Derived Extracts Suppress LPS Induced IL-1β in Human THP-1 Cells
2.4.2. Fucus vesiculosus Fucoidan (FVF) and Its Derived Extracts Suppress LPS Stimulated IL-1β in Human THP-1 Cells
2.4.3. Macrocystis pyrifera Fucoidan (MPF) and Its Derived Extracts Suppress LPS Induced IL-1β in Human THP-1 Cells
2.4.4. Fucoidan Extracts from Fucus vesiculosus with Higher Polyphenol Content (FVC), Ascophyllum nodosum (ANF), and Laminaria japonica (LJF) Inhibit LPS Induced IL-1β in Human THP-1 Cells in a Dose-Dependent Manner
3. Discussion
4. Materials and Methods
4.1. Cell Viability Assay
4.2. PBMC Cell Culture, Fucoidan Treatment, and Stimulation
4.3. THP-1 Cell Culture, Fucoidan Treatment, and LPS Stimulation
4.4. Enzyme-Linked Immunosorbent Assay ELISA
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Senthilkumar, K.; Kim, S.-K. Chapter 11—Anticancer Effects of Fucoidan. In Advances in Food and Nutrition Research; Kim, S.-K., Ed.; Academic Press: Cambridge, MA, USA, 2014; pp. 195–213. [Google Scholar]
- Carvalho, A.C.; Sousa, R.B.; Franco, X.; Costa, J.V.G.; Neves, L.M.; Ribeiro, R.A.; Sutton, R.; Criddle, D.N.; Soares, P.M.; de Souza, M.H. Protective Effects of Fucoidan, a P- and L-Selectin Inhibitor, in Murine Acute Pancreatitis. Pancreas 2014, 43, 82–87. [Google Scholar] [CrossRef]
- Fitton, J.H. Therapies from Fucoidan; Multifunctional Marine Polymers. Mar. Drugs 2011, 9, 1731–1760. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xing, M.; Cao, Q.; Ji, A.; Liang, H.; Song, S. Biological Activities of Fucoidan and the Factors Mediating Its Therapeutic Effects: A Review of Recent Studies. Mar. Drugs 2019, 17, 183. [Google Scholar] [CrossRef] [Green Version]
- Luthuli, S.; Wu, S.; Cheng, Y.; Zheng, X.; Wu, M.; Tong, H. Therapeutic Effects of Fucoidan: A Review on Recent Studies. Mar. Drugs 2019, 17, 487. [Google Scholar] [CrossRef] [Green Version]
- Rani, V.; Prabhu, A.; Venkatesan, J.; Kim, S.K. Seaweed Polysaccharides: Promising Molecules for Biotechnological Applications. In Comprehensive Glycoscience, 2nd ed.; Barchi, J.J., Ed.; Elsevier: Oxford, UK, 2021; pp. 131–141. [Google Scholar]
- Fitton, J.H.; Stringer, D.N.; Karpiniec, S.S. Therapies from Fucoidan: An Update. Mar. Drugs 2015, 13, 5920–5946. [Google Scholar] [CrossRef] [Green Version]
- Alekseyenko, T.V.; Zhanayeva, S.Y.; Venediktova, A.A.; Zvyagintseva, T.N.; Kuznetsova, T.A.; Besednova, N.N.; Korolenko, T. Antitumor and antimetastatic activity of fucoidan, a sulfated polysaccharide isolated from the Okhotsk sea Fucus evanescens brown alga. Bull. Exp. Biol. Med. 2007, 143, 730–732. [Google Scholar] [CrossRef]
- Ale, M.T.; Maruyama, H.; Tamauchi, H.; Mikkelsen, J.D.; Meyer, A.S. Fucoidan from Sargassum sp. and Fucus vesiculosus reduces cell viability of lung carcinoma and melanoma cells in vitro and activates natural killer cells in mice in vivo. Int. J. Biol. Macromol. 2011, 49, 331–336. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhang, Q.; Chen, L.; Ren, S.; Xu, P.; Tang, Y.; Luo, D. Higher specificity of the activity of low molecular weight fucoidan for thrombin-induced platelet aggregation. Thromb. Res. 2010, 125, 419–426. [Google Scholar] [CrossRef]
- Kim, K.T.; Rioux, L.E.; Turgeon, S.L. Alpha-amylase and alpha-glucosidase inhibition is differentially modulated by fucoidan obtained from Fucus vesiculosus and Ascophyllum nodosum. Phytochemistry 2014, 98, 27–33. [Google Scholar] [CrossRef]
- Wang, P.; Liu, Z.; Liu, X.; Teng, H.; Zhang, C.; Hou, L.; Zou, X. Anti-Metastasis Effect of Fucoidan from Undaria pinnatifida Sporophylls in Mouse Hepatocarcinoma Hca-F Cells. PLoS ONE 2014, 9, e106071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.W.; Liu, Q.; Thorlacius, H. Inhibition of Selectin Function and Leukocyte Rolling Protects Against Dextran Sodium Sulfate-Induced Murine Colitis. Scand. J. Gastroenterol. 2001, 36, 270–275. [Google Scholar] [CrossRef]
- Lean, Q.Y.; Eri, R.D.; Fitton, J.H.; Patel, R.P.; Gueven, N. Fucoidan Extracts Ameliorate Acute Colitis. PLoS ONE 2015, 10, e0128453. [Google Scholar] [CrossRef] [Green Version]
- Semenov, A.V.; Mazurov, A.V.; Preobrazhenskaia, M.E.; Ushakova, N.A.; Mikhaĭlov, V.I.; Berman, A.E.; Usov, A.I.; Nifant’ev, N.E.; Bovin, N.V. Sulfated polysaccharides as inhibitors of receptor activity of P-selectin and P-selectin-dependent inflam-mation. Voprosy Meditsinskoi Khimii 1998, 44, 135–144. [Google Scholar]
- Ale, M.T.; Mikkelsen, J.D.; Meyer, A.S. Important determinants for fucoidan bioactivity: A critical review of structure-function relations and extraction methods for fucose-containing sulfated polysaccharides from brown seaweeds. Mar. Drugs 2011, 9, 2106–2130. [Google Scholar] [CrossRef] [Green Version]
- Sang, V.T.; Kim, S.-K. Fucoidans as a natural bioactive ingredient for functional foods. J. Funct. Foods 2013, 5, 16–27. [Google Scholar]
- Zhang, W.; An, E.-K.; Park, H.-B.; Hwang, J.; Dhananjay, Y.; Kim, S.-J.; Eom, H.-Y.; Oda, T.; Kwak, M.; Lee, P.C.-W.; et al. Ecklonia cava fucoidan has potential to stimulate natural killer cells in vivo. Int. J. Biol. Macromol. 2021, 185, 111–121. [Google Scholar] [CrossRef]
- Phull, A.R.; Kim, S.J. Fucoidan as bio-functional molecule: Insights into the anti-inflammatory potential and associated molecular mechanisms. J. Funct. Foods 2017, 38, 415–426. [Google Scholar] [CrossRef]
- Lin, Z.; Tan, X.; Zhang, Y.; Li, F.; Luo, P.; Liu, H. Molecular Targets and Related Biologic Activities of Fucoidan: A Review. Mar. Drugs 2020, 18, 376. [Google Scholar] [CrossRef]
- Wallace, K.L. Immunopathology of inflammatory bowel disease. World J. Gastroenterol. 2014, 20, 6–21. [Google Scholar] [CrossRef]
- Heinsbroek, S.E.M.; Gordon, S. The role of macrophages in inflammatory bowel diseases. Expert Rev. Mol. Med. 2009, 11, e14. [Google Scholar] [CrossRef]
- Zhu, W.; Yu, J.; Nie, Y.; Shi, X.; Liu, Y.; Li, F.; Zhang, X.-L. Disequilibrium of M1 and M2 Macrophages Correlates with the Development of Experimental Inflammatory Bowel Diseases. Immunol. Investig. 2014, 43, 638–652. [Google Scholar] [CrossRef] [PubMed]
- Boyum, A. Isoation of leukocytes from human blood. Further observations. Methylcellulose, dextran, and ficoll as erythro-cyteaggregating agents. Scand. J. Clin. Lab. Investig. Suppl. 1968, 9731–9750. [Google Scholar]
- Chanput, W.; Mes, J.J.; Wichers, H.J. THP-1 cell line: An in vitro cell model for immune modulation approach. Int. Immunopharmacol. 2014, 23, 37–45. [Google Scholar] [CrossRef]
- Berridge, M.V.; Herst, P.M.; Tan, A.S. Tetrazolium dyes as tools in cell biology: New insights into their cellular reduction. Biotechnol. Annu. Rev. 2005, 11, 127–152. [Google Scholar] [CrossRef]
- Stockert, J.C.; Horobin, R.W.; Colombo, L.L.; Blázquez-Castro, A. Tetrazolium salts and formazan products in Cell Biology: Viability assessment, fluorescence imaging, and labeling perspectives. Acta Histochem. 2018, 120, 159–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Furman, D.; Campisi, J.; Verdin, E.; Carrera-Bastos, P.; Targ, S.; Franceschi, C.; Ferrucci, L.; Gilroy, D.W.; Fasano, A.; Miller, G.W.; et al. Chronic inflammation in the etiology of disease across the life span. Nat. Med. 2019, 25, 1822–1832. [Google Scholar] [CrossRef] [PubMed]
- Cox, A.J.; Cripps, A.W.; Taylor, P.A.; Fitton, J.H.; West, N.P. Fucoidan Supplementation Restores Fecal Lysozyme Concentrations in High-Performance Athletes: A Pilot Study. Mar. Drugs 2020, 18, 412. [Google Scholar] [CrossRef]
- Lahrsen, E.; Schoenfeld, A.-K.; Alban, S. Size-dependent pharmacological activities of differently degraded fucoidan fractions from Fucus vesiculosus. Carbohydr. Polym. 2018, 189, 162–168. [Google Scholar] [CrossRef]
- Trincone, A. Enzymatic Technologies for Marine Polysaccharides; CRC Press: Boca Raton, FL, USA, 2019. [Google Scholar]
- Choi, J.-I.; Kim, H.-J. Preparation of low molecular weight fucoidan by gamma-irradiation and its anticancer activity. Carbohydr. Polym. 2013, 97, 358–362. [Google Scholar] [CrossRef]
- Lemarchand, C.; Gref, R.; Couvreur, P. Polysaccharide-decorated nanoparticles. Eur. J. Pharm. Biopharm. 2004, 58, 327–341. [Google Scholar] [CrossRef] [PubMed]
- Atashrazm, F.; Lowenthal, R.M.; Woods, G.M.; Holloway, A.F.; Dickinson, J.L. Fucoidan and Cancer: A Multifunctional Molecule with Anti-Tumor Potential. Mar. Drugs 2015, 13, 2327–2346. [Google Scholar] [CrossRef] [Green Version]
- Aisa, Y.; Miyakawa, Y.; Nakazato, T.; Shibata, H.; Saito, K.; Ikeda, Y.; Kizaki, M. Fucoidan induces apoptosis of human HS-Sultan cells accompanied by activation of caspase-3 and down-regulation of ERK Pathways. Am. J. Hematol. 2004, 78, 7–14. [Google Scholar] [CrossRef]
- Ishikawa, C.; Mori, N. In vitro and in vivo anti-primary effusion lymphoma activities of fucoidan extracted from Cladosiphon okamuranus Tokida. Oncol. Rep. 2017, 38, 3197–3204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, J.-O.; Song, M.-G.; Kim, Y.-N.; Park, J.-I.; Kwak, J.-Y. The mechanism of fucoidan-induced apoptosis in leukemic cells: Involvement of ERK1/2, JNK, glutathione, and nitric oxide. Mol. Carcinog. 2010, 49, 771–782. [Google Scholar] [CrossRef]
- Park, H.S.; Hwang, H.J.; Kim, G.-Y.; Cha, H.-J.; Kim, W.-J.; Kim, N.D.; Yoo, Y.H.; Choi, Y.H. Induction of Apoptosis by Fucoidan in Human Leukemia U937 Cells through Activation of p38 MAPK and Modulation of Bcl-2 Family. Mar. Drugs 2013, 11, 2347–2364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Wang, H.; Shao, Q.; Kong, B.; Qu, X. Fucoidan modulates cytokine production and migration of THP-1-derived macrophages via colony-stimulating factor. Mol. Med. Rep. 2017, 15, 2325–2332. [Google Scholar] [CrossRef] [Green Version]
- Bittkau, K.S.; Dörschmann, P.; Blümel, M.; Tasdemir, D.; Roider, J.; Klettner, A.; Alban, S. Comparison of the Effects of Fucoidans on the Cell Viability of Tumor and Non-Tumor Cell Lines. Mar. Drugs 2019, 17, 441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braegger, C.; Nicholls, S.; Murch, S.; MacDonald, T.; Stephens, S. Tumour necrosis factor alpha in stool as a marker of intestinal inflammation. Lancet 1992, 339, 89–91. [Google Scholar] [CrossRef]
- Breese, E.J.; Michie, C.A.; Nicholls, S.W.; Murch, S.H.; Williams, C.B.; Domizio, P.; Walker-Smith, J.A.; Macdonald, T.T. Tumor necrosis factor α-producing cells in the intestinal mucosa of children with inflammatory bowel disease. Gastroenterology 1994, 106, 1455–1466. [Google Scholar] [CrossRef]
- Farrugia, M.; Baron, B. The role of TNF-α in rheumatoid arthritis: A focus on regulatory T cells. J. Clin. Transl. Res. 2016, 2, 84–90. [Google Scholar] [CrossRef]
- Kumar, S.; Joos, G.; Boon, L.; Tournoy, K.; Provoost, S.; Maes, T. Role of tumor necrosis factor-alpha and its receptors in diesel exhaust particle-induced pulmonary inflammation. Sci. Rep. 2017, 7, 11508. [Google Scholar] [CrossRef] [Green Version]
- Maeda, M.; Watanabe, N.; Neda, H.; Yamauchi, N.; Okamoto, T.; Sasaki, H.; Tsuji, Y.; Akiyama, S.; Tsuji, N.; Niitsu, Y. Serum Tumor Necrosis Factor Activity in Inflammatory Bowel Disease. Immunopharmacol. Immunotoxicol. 1992, 14, 451–461. [Google Scholar] [CrossRef]
- Dosh, R.H.; Jordan-Mahy, N.; Sammon, C.; Le Maitre, C. Interleukin 1 is a key driver of inflammatory bowel disease-demonstration in a murine IL-1Ra knockout model. Oncotarget 2019, 10, 3559–3575. [Google Scholar] [CrossRef] [Green Version]
- Mudter, J.; Neurath, M.F. Il-6 signaling in inflammatory bowel disease: Pathophysiological role and clinical relevance. Inflamm. Bowel Dis. 2007, 13, 1016–1023. [Google Scholar] [CrossRef] [PubMed]
- Eapen, M.S.; Hansbro, P.M.; McAlinden, K.; Kim, R.Y.; Ward, C.; Hackett, T.-L.; Walters, E.H.; Sohal, S.S. Abnormal M1/M2 macrophage phenotype profiles in the small airway wall and lumen in smokers and chronic obstructive pulmonary disease (COPD). Sci. Rep. 2017, 7, 13392. [Google Scholar] [CrossRef]
- Eapen, M.; Sohal, S.S. Understanding novel mechanisms of microbial pathogenesis in chronic lung disease: Implications for new therapeutic targets. Clin. Sci. 2018, 132, 375–379. [Google Scholar] [CrossRef] [PubMed]
- Schildberger, A.; Rossmanith, E.; Eichhorn, T.; Strassl, K.; Weber, V. Monocytes, Peripheral Blood Mononuclear Cells, and THP-1 Cells Exhibit Different Cytokine Expression Patterns following Stimulation with Lipopolysaccharide. Mediat. Inflamm. 2013, 2013, 697972. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, L.S.; Tsunawaki, S. Expression of NADPH oxidases and enhanced H2O2-generating activity in human coronary artery endothelial cells upon induction with tumor necrosis factor-α. Int. Immunopharmacol. 2008, 8, 1377–1385. [Google Scholar] [CrossRef]
- Guzik, T.J.; Korbut, R.; Adamek-Guzik, T. Nitric oxide and superoxide in inflammation and immune regulation. J. Physiol. Pharmacol. 2003, 54, 469–487. [Google Scholar] [PubMed]
- Pugazhenthi, S.; Zhang, Y.; Bouchard, R.; Mahaffey, G. Induction of an Inflammatory Loop by Interleukin-1β and Tumor Necrosis Factor-α Involves NF-kB and STAT-1 in Differentiated Human Neuroprogenitor Cells. PLoS ONE 2013, 8, e69585. [Google Scholar]
- Moe, K.T.; Khairunnisa, K.; Yin, N.O.; Chin-Dusting, J.; Wong, P.; Cheong Wong, M. Tumor necrosis factor-α-induced nuclear factor-kappaB activation in human cardiomyocytes is mediated by NADPH oxidase. J. Physiol. Biochem. 2014, 70, 769–779. [Google Scholar] [CrossRef]
- Tanaka, T.; Narazaki, M.; Kishimoto, T. IL-6 in inflammation, immunity, and disease. Cold Spring Harb. Perspect. Biol. 2014, 6, a016295. [Google Scholar] [CrossRef]
- He, G.; Karin, M. NF-κB and STAT3-key players in liver inflammation and cancer. Cell Res. 2011, 21, 159–168. [Google Scholar] [CrossRef] [Green Version]
- Gabay, C. Interleukin-6 and chronic inflammation. Arthritis Res. 2006, 8, S3. [Google Scholar] [CrossRef] [Green Version]
- Hurst, S.M.; Wilkinson, T.; McLoughlin, R.; Jones, S.; Horiuchi, S.; Yamamoto, N.; Rose-John, S.; Fuller, G.M.; Topley, N.; Jones, S.A. IL-6 and Its Soluble Receptor Orchestrate a Temporal Switch in the Pattern of Leukocyte Recruitment Seen during Acute Inflammation. Immunity 2001, 14, 705–714. [Google Scholar] [CrossRef] [Green Version]
- Jeong, J.-W.; Hwang, S.J.; Han, M.H.; Lee, D.-S.; Yoo, J.S.; Choi, I.-W.; Cha, H.-J.; Kim, S.; Kim, H.-S.; Kim, G.-Y.; et al. Fucoidan inhibits lipopolysaccharide-induced inflammatory responses in RAW 264.7 macrophages and zebrafish larvae. Mol. Cell. Toxicol. 2017, 13, 405–417. [Google Scholar] [CrossRef]
- Ni, L.; Wang, L.; Fu, X.; Duan, D.; Jeon, Y.-J.; Xu, J.; Gao, X. In vitro and in vivo anti-inflammatory activities of a fucose-rich fucoidan isolated from Saccharina japonica. Int. J. Biol. Macromol. 2020, 156, 717–729. [Google Scholar] [CrossRef] [PubMed]
- Hsu, H.-Y.; Chiu, S.-L.; Wen, M.-H.; Chen, K.-Y.; Hua, K.-F. Ligands of Macrophage Scavenger Receptor Induce Cytokine Expression via Differential Modulation of Protein Kinase Signaling Pathways. J. Biol. Chem. 2001, 276, 28719–28730. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Oda, T.; Yu, Q.; Jin, J.-O. Fucoidan from Macrocystis pyrifera Has Powerful Immune-Modulatory Effects Compared to Three Other Fucoidans. Mar. Drugs 2015, 13, 1084–1104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Park, H.-B.; Yadav, D.; Hwang, J.; An, E.-K.; Eom, H.-Y.; Kim, S.-J.; Kwak, M.; Lee, P.C.-W.; Jin, J.-O. Comparison of human peripheral blood dendritic cell activation by four fucoidans. Int. J. Biol. Macromol. 2021, 174, 477–484. [Google Scholar] [CrossRef] [PubMed]
- Widdrington, J.D.; Gomez-Duran, A.; Pyle, A.; Ruchaud-Sparagano, M.-H.; Scott, J.; Baudouin, S.V.; Rostron, A.J.; Lovat, P.; Chinnery, P.F.; Simpson, A.J. Exposure of Monocytic Cells to Lipopolysaccharide Induces Coordinated Endotoxin Tolerance, Mitochondrial Biogenesis, Mitophagy, and Antioxidant Defenses. Front. Immunol. 2018, 9, 2217. [Google Scholar] [CrossRef]
- Sudhakaran, V.A.; Panwar, H.; Chauhan, R.; Duary, R.K.; Rathore, R.K.; Batish, V.K.; Grover, S. Modulation of anti-inflammatory response in lipopolysaccharide stimulated human THP-1 cell line and mouse model at gene expression level with indigenous putative probiotic lactobacilli. Genes Nutr. 2013, 8, 637–648. [Google Scholar] [CrossRef] [Green Version]
- Chometon, T.Q.; Siqueira, M.D.S.; Sant´anna, J.C.; Almeida, M.R.; Gandini, M.; Nogueira, A.C.M.D.A.; Antas, P.R.Z. A protocol for rapid monocyte isolation and generation of singular human monocyte-derived dendritic cells. PLoS ONE 2020, 15, e0231132. [Google Scholar] [CrossRef] [Green Version]
- Citkowska, A.; Szekalska, M.; Winnicka, K. Possibilities of Fucoidan Utilization in the Development of Pharmaceutical Dosage Forms. Mar. Drugs 2019, 17, 458. [Google Scholar] [CrossRef] [Green Version]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Filisetti-Cozzi, T.M.; Carpita, N.C. Measurement of uronic acids without interference from neutral sugars. Anal. Biochem. 1991, 197, 157–162. [Google Scholar] [CrossRef]
- Dodgson, K.S. Determination of inorganic sulphate in studies on the enzymic and non-enzymic hydrolysis of carbohydrate and other sulphate esters. Biochem. J. 1961, 78, 312–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Autissier, P.; Soulas, C.; Burdo, T.H.; Williams, C.K. Evaluation of a 12-color flow cytometry panel to study lymphocyte, monocyte, and dendritic cell subsets in humans. Cytom. Part A J. Int. Soc. Adv. Cytom. 2010, 77, 410–419. [Google Scholar] [CrossRef] [PubMed]
- Davila, J.A.A.; Rios, A.H.D.L. An Overview of Peripheral Blood Mononuclear Cells as a Model for Immunological Research of Toxoplasma gondii and Other Apicomplexan Parasites. Front. Cell. Infect. Microbiol. 2019, 9, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Species | Origin | Description |
|---|---|---|---|
| UPF | Undaria pinnatifida | South American | ≥85% purity |
| DP UPF | Undaria pinnatifida | South American | ≥90% purity and depyrogenated |
| 5–30 DP UPF | Undaria pinnatifida | South American | ≥90% purity, depyrogenated and 5–30 kDa |
| FVF | Fucus vesiculosus | Canadian | ≥90% purity |
| DP FVF | Fucus vesiculosus | Canadian | ≥90% purity and depyrogenated |
| 10–30 DP FVF | Fucus vesiculosus | Canadian | >90% purity, depyrogenated and 10–30 kDa |
| FVC | Fucus vesiculosus | Canadian | ≥85% purity |
| MPF | Macrocystis pyrifera | South American | ≥85% purity |
| 30+ DP MPF | Macrocystis pyrifera | South American | ≥85% purity, ≥30 kDa and depyrogenated |
| 5–30 DP MPF | Macrocystis pyrifera | South American | ≥85% purity, 5–30 kDa and depyrogenated |
| ANF | Ascophyllum nodosum | Canadian | ≥85% purity |
| LJF | Laminaria japonica | Japan | ≥85% purity |
| Co | Total Carbohydrates (%) | Uronic Acid (%) | Polyphenols (%) | SO4 (%) | Cations (%) | Peak MW (kDa) |
|---|---|---|---|---|---|---|
| UPF | 54.2 | 4.1 | <2.5 | 24.9 | 8.0 | 40.4 |
| DP UPF | 64.0 | 0.9 | <2.5 | 31.0 | 6.8 | 134.2 |
| 5–30 DP UPF | 61.3 | 0.7 | <2.5 | 27.6 | 6.0 | 8.2 |
| FVF | 66.5 | 3.6 | 2.5 | 25.9 | 6.2 | 35.8 |
| DP FVF | 67.5 | 2.0 | <2.5 | 26.0 | 11.2 | 54.0 |
| 10–30 DP FVF | 71.5 | 2.2 | <2.5 | 26.9 | 8.8 | 21.3 |
| FVC | 59.6 | 6.2 | 15.5 | 14.0 | 5.7 | 48.8 |
| MPF | 51.1 | 6.1 | <2.5 | 25.7 | 7.9 | 66.0 |
| 30+ DP MPF | 56.9 | 5.7 | <2.5 | 23.0 | 10.8 | 32.5 |
| 5–30 DP MPF | 54.6 | 7.1 | <2.5 | 19.7 | 10.8 | 17.4 |
| ANF | 66.6 | 4.1 | 17.5 | 24.2 | 5.0 | 115.5 |
| LJF | 55.2 | 23.8 | <2.5 | 18.6 | 5.0 | 597.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ahmad, T.; Eapen, M.S.; Ishaq, M.; Park, A.Y.; Karpiniec, S.S.; Stringer, D.N.; Sohal, S.S.; Fitton, J.H.; Guven, N.; Caruso, V.; et al. Anti-Inflammatory Activity of Fucoidan Extracts In Vitro. Mar. Drugs 2021, 19, 702. https://doi.org/10.3390/md19120702
Ahmad T, Eapen MS, Ishaq M, Park AY, Karpiniec SS, Stringer DN, Sohal SS, Fitton JH, Guven N, Caruso V, et al. Anti-Inflammatory Activity of Fucoidan Extracts In Vitro. Marine Drugs. 2021; 19(12):702. https://doi.org/10.3390/md19120702
Chicago/Turabian StyleAhmad, Tauseef, Mathew Suji Eapen, Muhammad Ishaq, Ah Young Park, Samuel S. Karpiniec, Damien N. Stringer, Sukhwinder Singh Sohal, J. Helen Fitton, Nuri Guven, Vanni Caruso, and et al. 2021. "Anti-Inflammatory Activity of Fucoidan Extracts In Vitro" Marine Drugs 19, no. 12: 702. https://doi.org/10.3390/md19120702
APA StyleAhmad, T., Eapen, M. S., Ishaq, M., Park, A. Y., Karpiniec, S. S., Stringer, D. N., Sohal, S. S., Fitton, J. H., Guven, N., Caruso, V., & Eri, R. (2021). Anti-Inflammatory Activity of Fucoidan Extracts In Vitro. Marine Drugs, 19(12), 702. https://doi.org/10.3390/md19120702