
Haloferax mediterranei Cells as C50 Carotenoid Factories

 , ,
, ,  ,
, 

Abstract
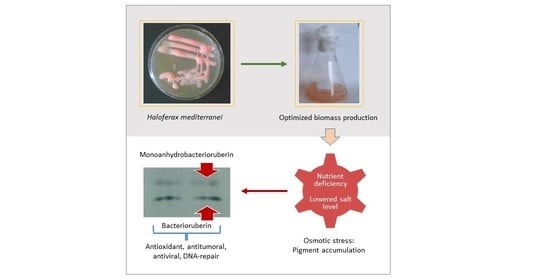
:
1. Introduction
2. Results
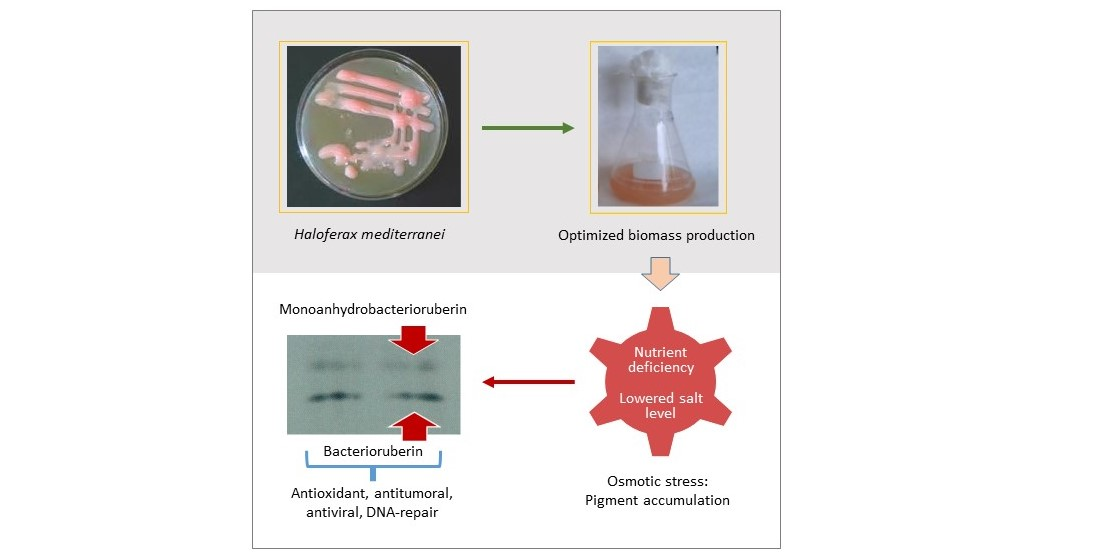
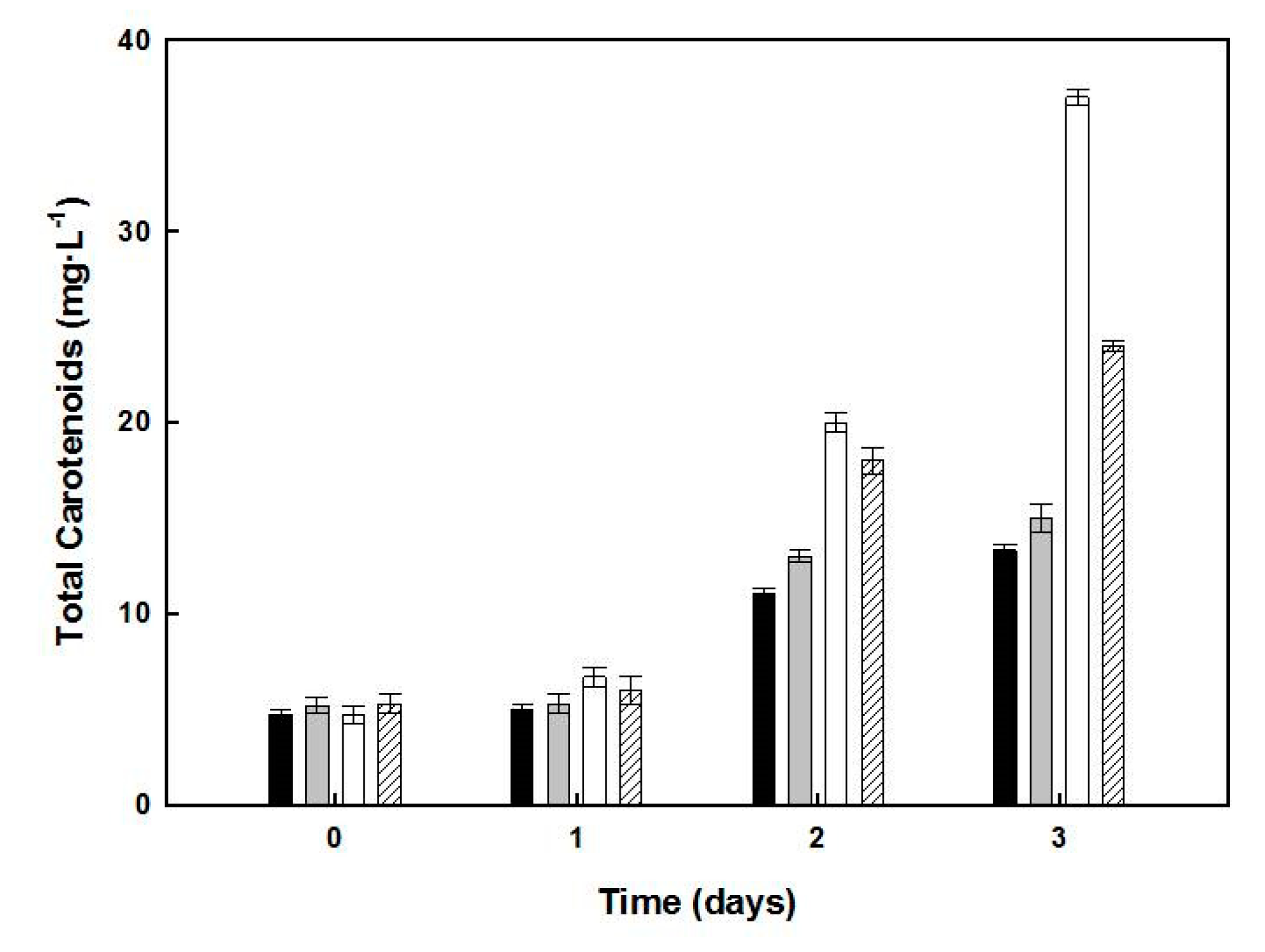
2.1. Carotenoids Content of Hfx. mediterranei All along the Growth Curve
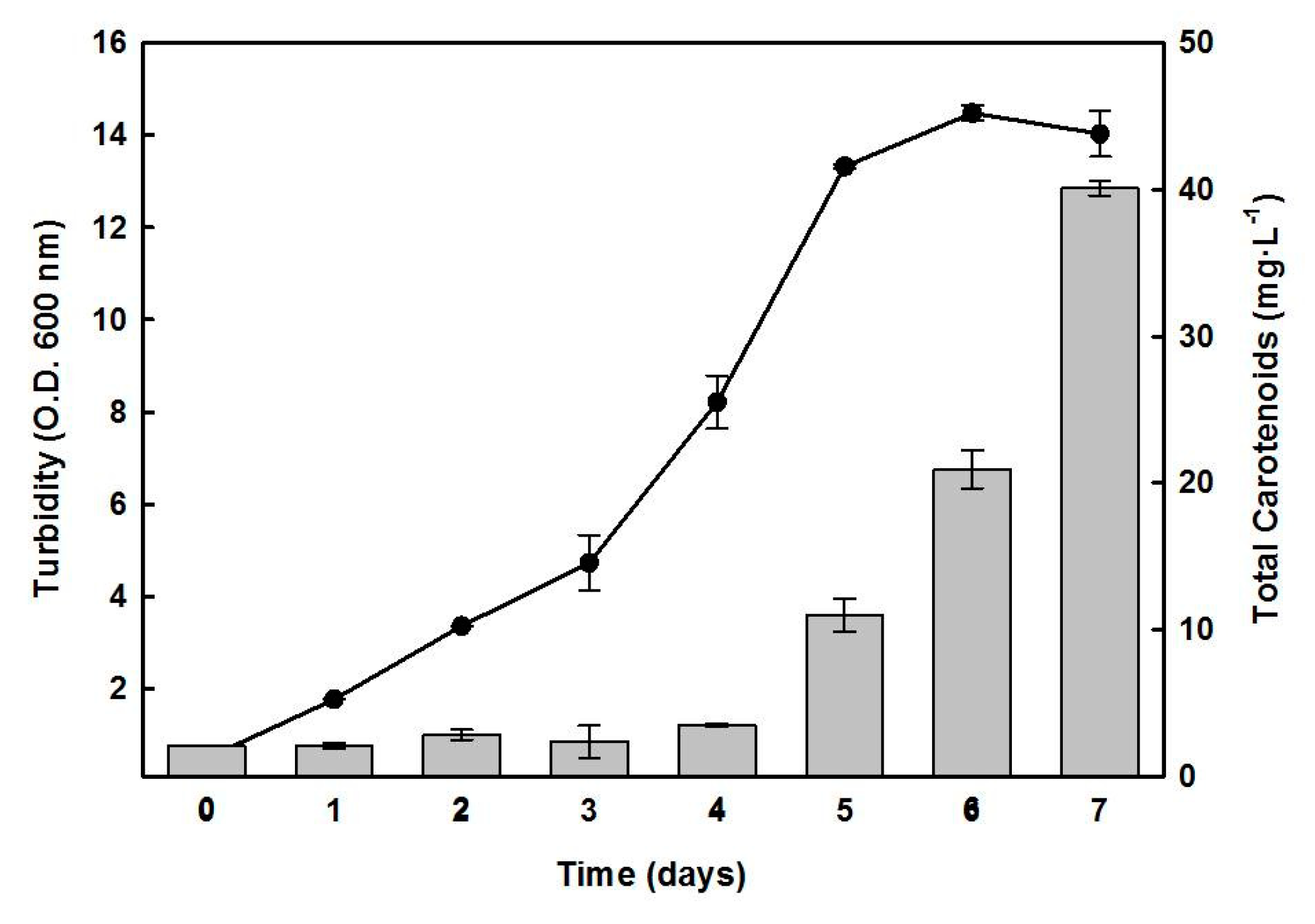
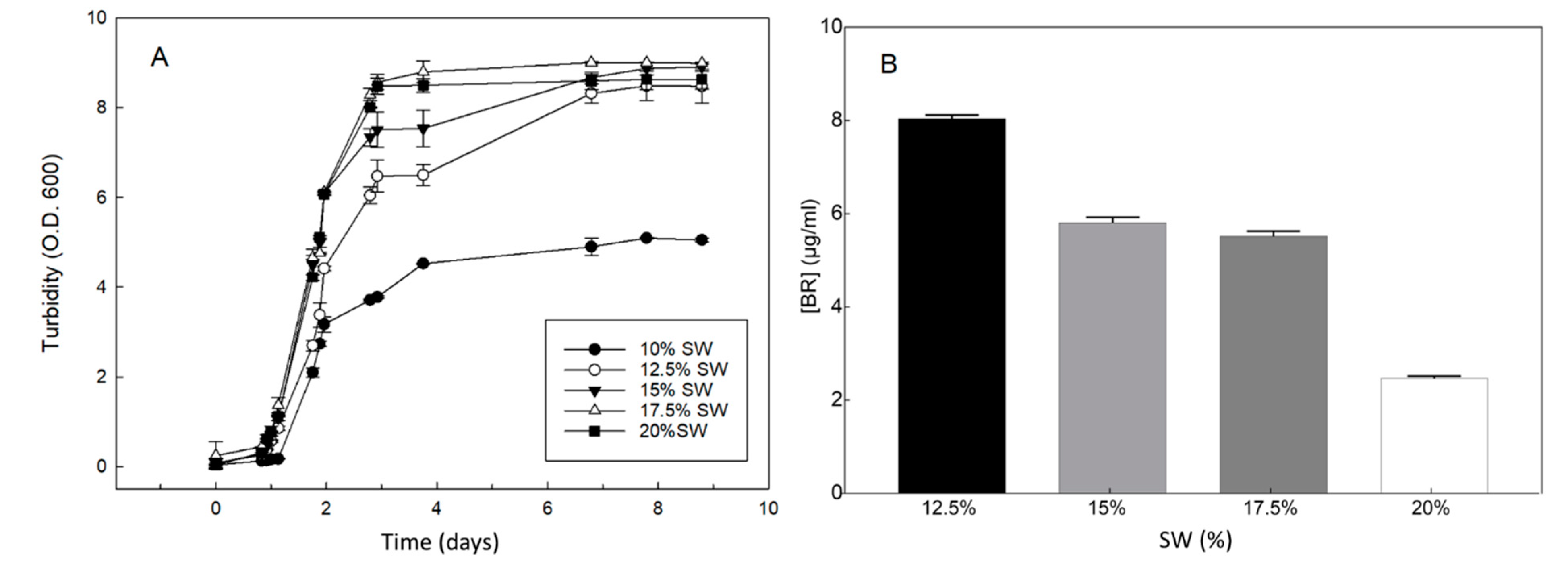
2.2. Effects of Nutrient Limitation on the Growth of Hfx. mediterranei and Synthesis of C50 Carotenoids
2.3. Effect of Inorganic Salts on the Growth and BR Production of Hfx. mediterranei
2.4. Effect of Carbon/Nitrogen (C/N) Ratio in the Medium on BR Production by Hfx. mediterranei
2.5. Effects of Magnesium or Sulfate on the Growth and Synthesis of Pigments by Hfx. mediterranei
3. Discussion
4. Materials and Methods
4.1. Microorganism and Standard Growth Conditions
4.2. Cultivation in the Absence of Glucose and/or Yeast Extracts
4.3. Growth Determination
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Oren, A. Thermodinamic limits to microbial life at high salt concentrations. Environ. Microbiol. 2011. [Google Scholar] [CrossRef]
- Amoozegar, M.A.; Siroosi, M.; Atashgahi, S.; Smidt, H.; Ventosa, A. Systematics of haloarchaea and biotechnological potential of their hydrolytic enzymes. Microbiology 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koller, M.; Maršálek, L.; de Sousa, M.M.; Braunegg, G. Producing microbial polyhydroxyalkanoate (PHA) biopolyesters in a sustainable manner. New Biotechnol. 2017, 25, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Wu, L.P.; Liu, X.-B.; Hou, J.; Zhao, L.L.; Chen, J.Y.; Zhao, D.-H.; Xiang, H. Biodegradation and biocompatibility of haloarchaea-produced poly(3-hydroxybutyrate-co-3-hydroxyvalerate) copolymers. Biomaterials 2017, 139, 172–186. [Google Scholar] [CrossRef] [PubMed]
- Rodrigo-Baños, M.; Garbayo, I.; Vílchez, C.; Bonete, M.J.; Martínez-Espinosa, R.M. Carotenoids from Haloarchaea and their potential in biotechnology. Mar. Drugs 2015, 13, 5508–5532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giani, M.; Garbayo, I.; Vílchez, C.; Martínez-Espinosa, R.M. Haloarchaeal Carotenoids: Healthy Novel Compounds from Extreme Environments. Mar. Drugs 2019, 17, 524. [Google Scholar] [CrossRef] [Green Version]
- Montero-Lobato, Z.; Ramos-Merchante, A.; Fuentes, J.L.; Sayago, A.; Fernández-Recamales, A.; Martínez-Espinosa, R.M.; Vega, J.M.; Vílchez, C.; Garbayo, I. Optimization of growth and carotenoid production by Haloferax mediterranei using response surface methodology. Mar. Drugs 2018, 16, 372. [Google Scholar] [CrossRef] [Green Version]
- Jones, D.L.; Baxter, B.K. DNA repair and photoprotection: Mechanisms of overcoming environmental ultraviolet radiation exposure in halophilic archaea. Front. Microbiol. 2017, 8, 01882. [Google Scholar] [CrossRef]
- Saito, T.; Miyabe, Y.; Ide, H.; Yamamoto, O. Hydroxyl radical scavenging ability of bacterioruberin. Rad. Phys. Chem. 1997, 50, 267–269. [Google Scholar] [CrossRef]
- Zalazar, L.; Pagola, P.; Miro, M.V.; Churio, M.S.; Cerletti, M.; Martínez, C.; Iniesta-Cuerda, M.; Soler, A.J.; Cesari, A.; De Castro, R. Bacterioruberin extracts from a genetically modified hyperpigmented Haloferax volcanii strain: Antioxidant activity and bioactive properties on sperm cells. J. Appl. Microbiol. 2019, 126, 796–810. [Google Scholar] [CrossRef]
- Shahmohammadi, H.R.; Asgarani, E.; Terato, H.; Saito, T.; Ohyama, Y.; Gekko, K.; Yamamoto, O.; Ide, H. Protective roles of bacterioruberin and intracellular KCl in the resistance of Halobacterium salinarium against DNA-damaging Agents. J. Radiat. Res. 2008, 39, 251–262. [Google Scholar] [CrossRef] [Green Version]
- DasSarma, S.; DasSarma, P.; Laye, V.J.; Schwieterman, E.W. Extremophilic models for astrobiology: Haloarchaeal survival strategies and pigments for remote sensing. Extremophiles 2020, 24, 31–41. [Google Scholar] [CrossRef]
- Mussagy, C.U.; Winterburn, J.; Santos-Ebinuma, V.C.; Pereira, J.F.B. Production and extraction of carotenoids produced by microorganisms. Appl. Microbiol. Biotechnol. 2019, 103, 1095–1114. [Google Scholar] [CrossRef]
- Sui, L.; Liu, L.; Deng, Y. Characterization of halophilic C50 carotenoid-producing archaea isolated from solar saltworks in Bohai Bay, China. Chin. J. Oceanol. Limnol. 2014, 32, 1280–1287. [Google Scholar] [CrossRef]
- Naziri, D.; Hamidi, M.; Hassanzadeh, S.; Tarhriz, V.; Zanjani, B.M.; Nazemyieh, H.; Hejazi, M.A.; Hejazi, M.S. Analysis of carotenoid production by Halorubrum sp. TBZ126; an extremely halophilic archeon from Urmia lake. Adv. Pharm. Bull 2014, 4, 61–67. [Google Scholar] [PubMed] [Green Version]
- Hechler, T.; Frech, M.; Pfeifer, F. Glucose inhibits the formation of gas vesicles in Haloferax volcanii transformants. Environ. Microbiol. 2008, 10, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Calegari-Santos, R.; Diogo, R.A.; Fontana, J.D.; Bonfim, T.M.B. Carotenoid Production by Halophilic Archaea under Different Culture Conditions. Curr. Microbiol. 2016, 72, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Soliman, G.S.H.; Trueper, H.G. Halobacterium pharaonis sp. nov., a new, extremely haloalkaliphilic archaebacterium with low magnesium requirement. Zentral Bakteriol. Angew. Okol. Microbiol 1982, 3, 318–329. [Google Scholar] [CrossRef]
- Squillaci, G.; Parrella, R.; Carbone, V.; Minasi, P.; La Cara, F.; Morana, A. Carotenoids from the extreme halophilic archaeon Haloterrigena turkmenica: Identification and antioxidant activity. Extremophiles 2017, 21, 933–945. [Google Scholar] [CrossRef]
- Chen, C.W.; Hsu, S.H.; Lin, M.T.; Hsu, Y.H. Mass production of C50 carotenoids by Haloferax mediterranei in using extruded rice bran and starch under optimal conductivity of brined medium. Bioprocess Biosyst. Eng. 2015, 38, 2361–2367. [Google Scholar] [CrossRef]
- Fang, C.J.; Ku, K.L.; Lee, M.H.; Su, N.W. Influence of nutritive factors on C50 carotenoids production by Haloferax mediterranei ATCC 33500 with two-stage cultivation. Bioresour. Technol. 2010, 101, 6487–6493. [Google Scholar] [CrossRef]
- Hamidi, M. Optimization of total carotenoid production by Halorubrum sp. TBZ126 using Response surface methodology. J. Microb. Biochem. Technol. 2014, 6, 286–294. [Google Scholar] [CrossRef]
- Yatsunami, R.; Ando, A.; Yang, Y.; Takaichi, S.; Kohno, M.; Matsumura, Y.; Ikeda, H.; Fukui, T.; Nakasone, K.; Fujita, N.; et al. Identification of carotenoids from the extremely halophilic archaeon Haloarcula japonica. Front. Microbiol. 2014, 5, 100. [Google Scholar] [CrossRef]
- Zuo, Z.Q.; Xue, Q.; Zhou, J.; Zhao, D.H.; Han, J.; Xiang, H. Engineering Haloferax mediterranei as an efficient platform for high level production of lycopene. Front. Microbiol. 2018, 9, 2893. [Google Scholar] [CrossRef] [PubMed]
- Lamers, P.P.; Janssen, M.; De Vos, R.C.H.; Bino, R.J.; Wijffels, R.H. Carotenoid and fatty acid metabolism in nitrogen-starved Dunaliella salina, a unicellular green microalga. J. Biotechnol. 2012, 162, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Simionato, D.; Block, M.A.; La Rocca, N.; Jouhet, J.; Maréchal, E.; Finazzi, G.; Morosinotto, T. The response of Nannochloropsis gaditana to nitrogen starvation includes de novo biosynthesis of triacylglycerols, a decrease of chloroplast galactolipids, and reorganization of the photosynthetic apparatus. Eukaryot. Cell 2013, 12, 665–676. [Google Scholar] [CrossRef] [Green Version]
- Orosa, M.; Valero, J.F.; Herrero, C.; Abalde, J. Comparison of the accumulation of astaxanthin in Haematococcus pluvialis and other green microalgae under N-starvation and high light conditions. Biotechnol. Lett. 2001, 23, 1079–1085. [Google Scholar] [CrossRef] [Green Version]
- Sahli, K.; Gomri, M.A.; Esclapez, J.; Gómez-Villegas, P.; Ghennai, O.; Bonete, M.J.; León, R.; Kharroub, K. Bioprospecting and characterization of pigmented halophilic archaeal strains from Algerian hypersaline environments with analysis of carotenoids produced by Halorubrum sp. BS2. J. Basic. Microbiol. 2020, 60, 624–638. [Google Scholar] [CrossRef]
- Giani, M.; Martínez-Espinosa, R.M. Carotenoids as a Protection Mechanism against Oxidative Stress in Haloferax Mediterr. Antioxidants 2020, 9, 1060. [Google Scholar] [CrossRef]
- Mullakhanbhai, M.F.; Larsen, H. Halobacterium volcanii spec. nov., a Dead Sea halobacterium with a moderate salt requirement. Arch. Microbiol 1975, 104, 207–214. [Google Scholar] [CrossRef]
- Oren, A. Halobacterium sodomense sp. nov., a Dead Sea halobacterium with an extremely high magnesium requirement. Int. J. Syst. Bacteriol 1983, 33, 381–386. [Google Scholar] [CrossRef]
- Strand, A.; Shivaji, S.; Liaaen-Jensen, S. Bacterial carotenoids 55. C50 carotenoids 25. Revised structures of carotenoids associated with membranes in psychrotrophic Micrococcus roseus. Biochem. Syst. Ecol. 1997, 25, 547–552. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Culture 1 | C/N Ratio | Turbidity (OD 600 nm) | Pigmentation Level 1 | |
|---|---|---|---|---|
| SW (%) | Nitrate (mM) | |||
| 25 | 5 | 5.6 | 2.1 | - |
| 25 | 25 | 1.1 | 2.3 | ++ |
| 25 | 100 | 0.28 | 2.4 | +++ |
| 15 | 5 | 5.6 | 1.8 | + |
| 15 | 25 | 1.1 | 1.9 | +++ |
| 15 | 100 | 0.28 | 2.2 | ++++ |
| 15 | 100 (S) | 0.28 | 2.0 | +++++ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Giani, M.; Montero-Lobato, Z.; Garbayo, I.; Vílchez, C.; Vega, J.M.; Martínez-Espinosa, R.M. Haloferax mediterranei Cells as C50 Carotenoid Factories. Mar. Drugs 2021, 19, 100. https://doi.org/10.3390/md19020100
Giani M, Montero-Lobato Z, Garbayo I, Vílchez C, Vega JM, Martínez-Espinosa RM. Haloferax mediterranei Cells as C50 Carotenoid Factories. Marine Drugs. 2021; 19(2):100. https://doi.org/10.3390/md19020100
Chicago/Turabian StyleGiani, Micaela, Zaida Montero-Lobato, Inés Garbayo, Carlos Vílchez, José M. Vega, and Rosa María Martínez-Espinosa. 2021. "Haloferax mediterranei Cells as C50 Carotenoid Factories" Marine Drugs 19, no. 2: 100. https://doi.org/10.3390/md19020100
APA StyleGiani, M., Montero-Lobato, Z., Garbayo, I., Vílchez, C., Vega, J. M., & Martínez-Espinosa, R. M. (2021). Haloferax mediterranei Cells as C50 Carotenoid Factories. Marine Drugs, 19(2), 100. https://doi.org/10.3390/md19020100