
A New Micromonospora Strain with Antibiotic Activity Isolated from the Microbiome of a Mid-Atlantic Deep-Sea Sponge

 ,
,  ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Results
2.1. Sponge Sample Collection and Processing
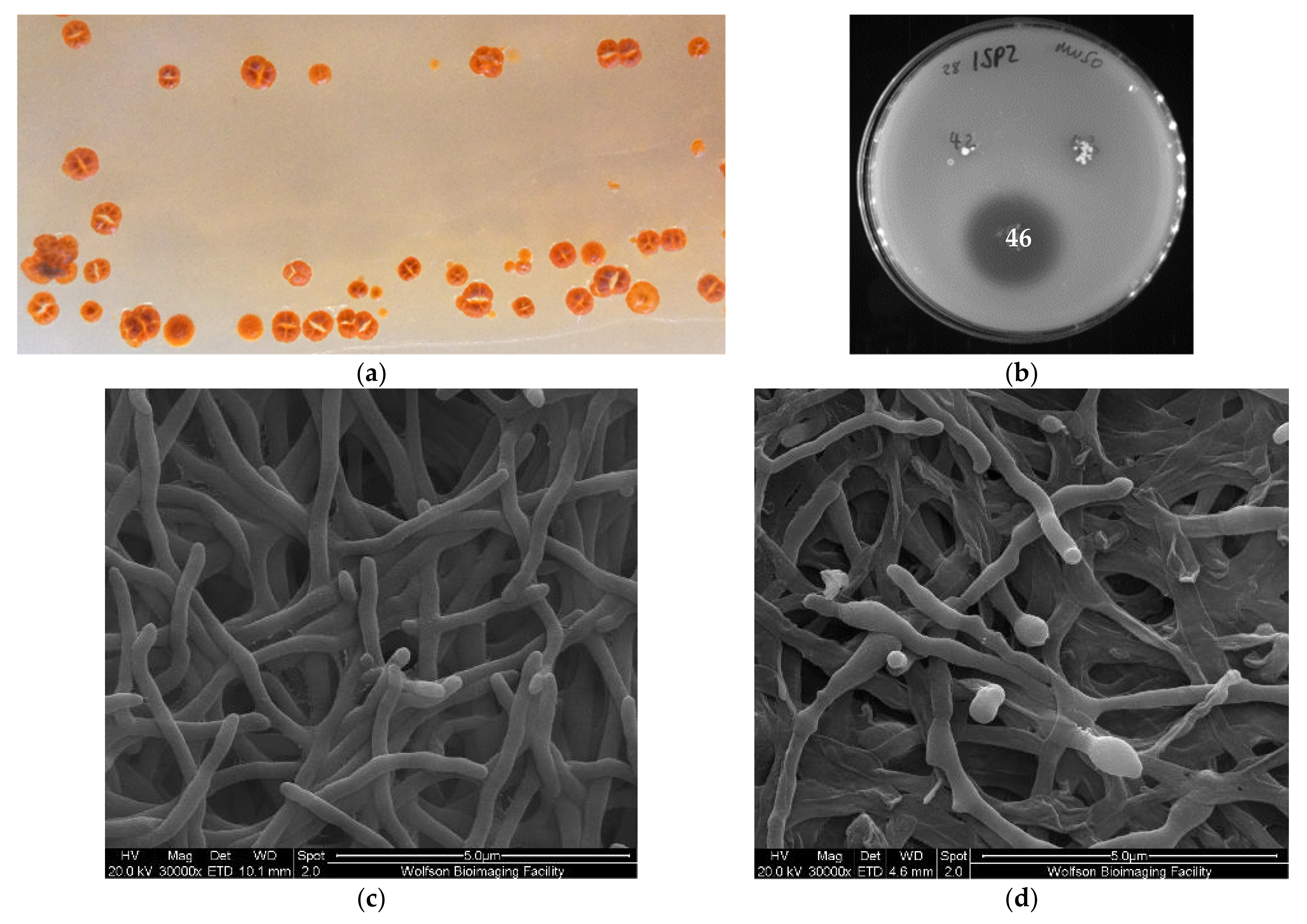
2.2. Isolation of a Micromonospora Strain with Antibiotic Activity
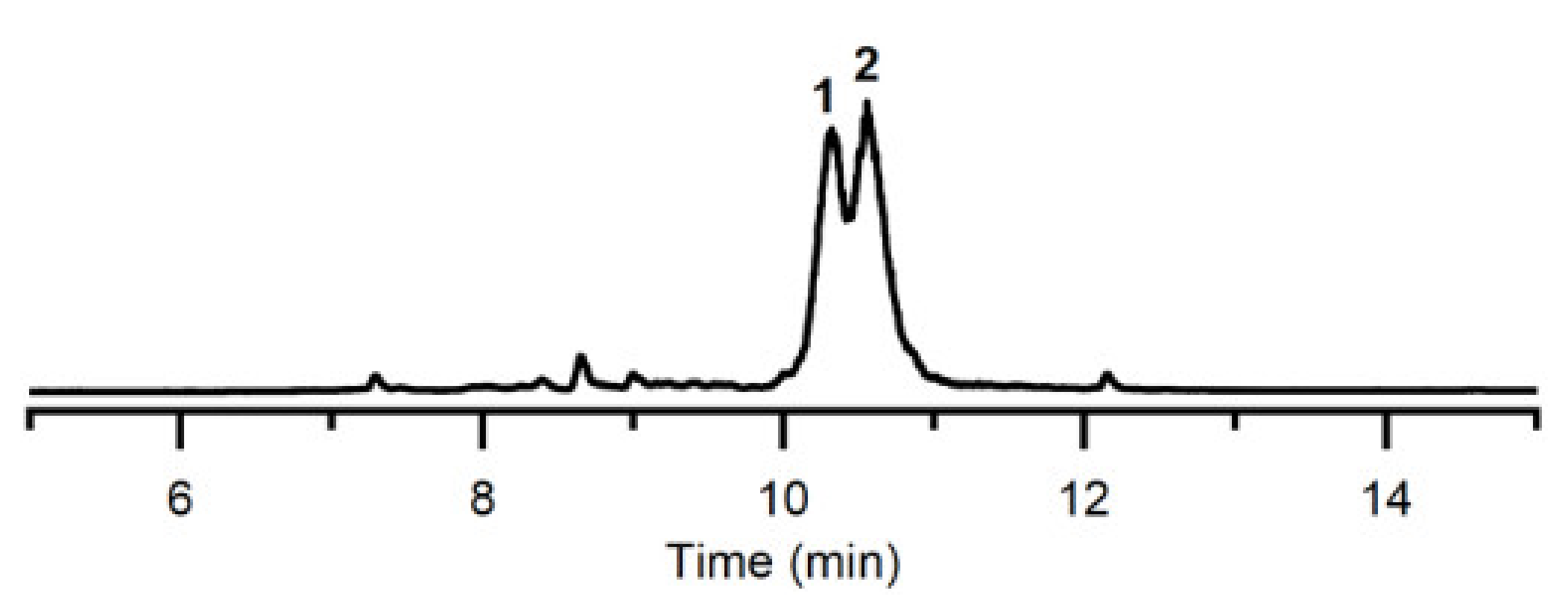
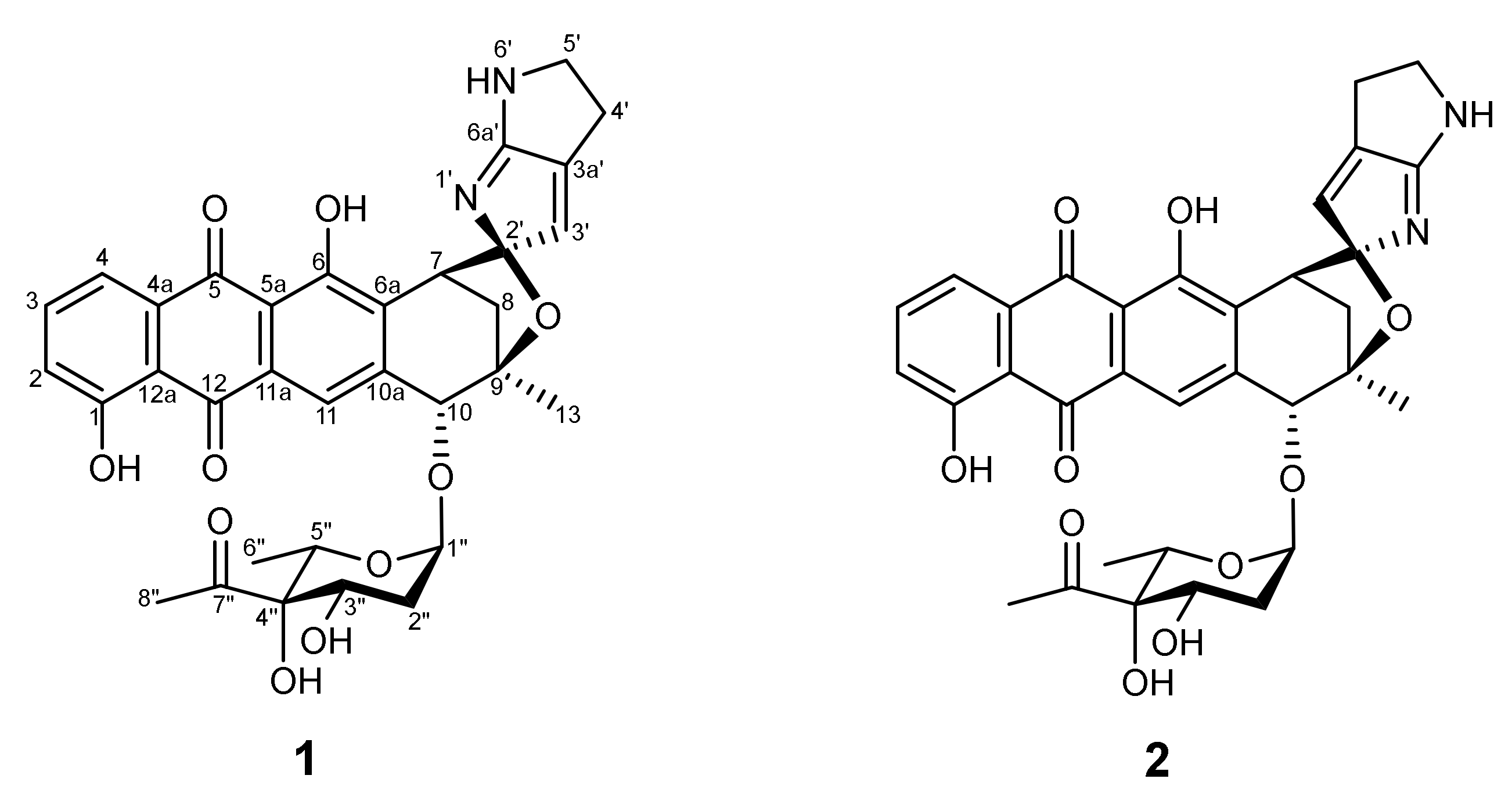
2.3. Identification of Quinocycline Antibiotics
2.4. Biological Activities
2.5. Genome Sequencing and Assembly
2.6. Quinocycline-Producing Gene Cluster
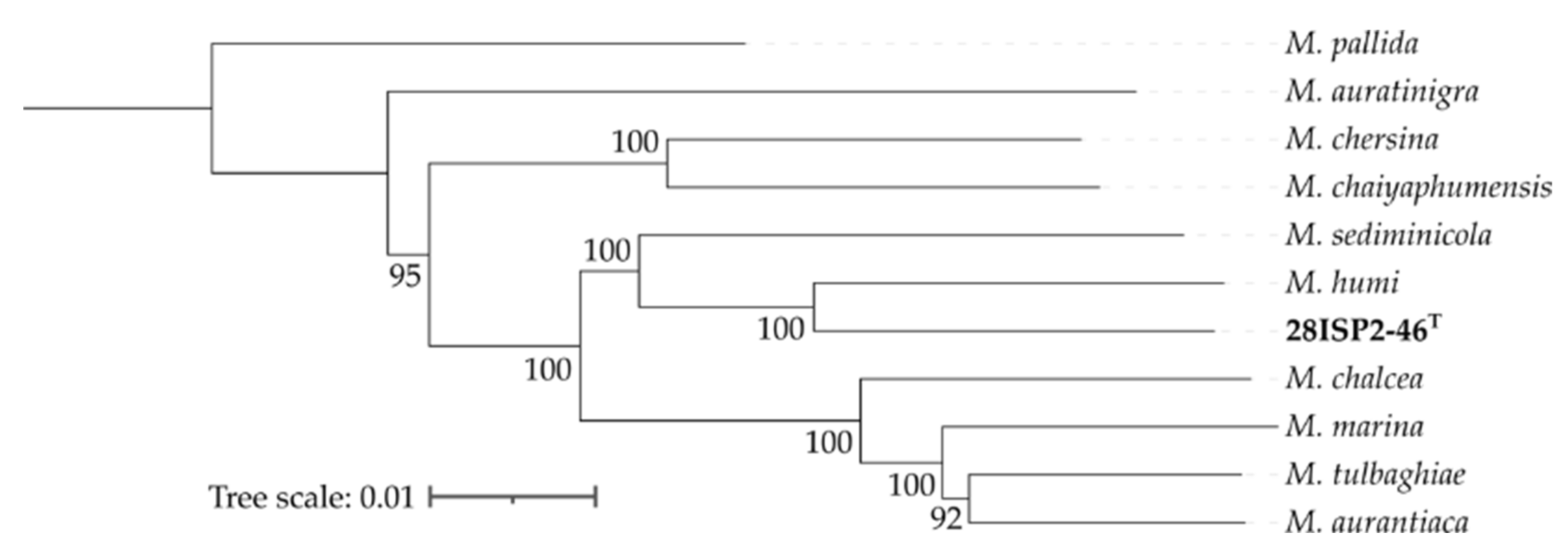
2.7. Genome Comparison of Strain 28ISP2-46T with Other Micromonospora Species
3. Discussion
Description of ‘Micromonospora robiginosa’ sp. nov.
4. Materials and Methods
4.1. Collection of Sponge Samples
4.2. Sponge Processing and Isolation of Bacterial Strains
4.3. 16S rRNA Gene Sequencing of Strain 28ISP2-46T
4.4. Cultivation Conditions of 28ISP2-46T
4.5. Scanning Electron Microscopy of 28ISP2-46T
4.6. Soft Agar Overlay Antibiotic Activity Assay
4.7. Small-Volume Liquid Culture Antibiotic Activity Assay of 28ISP2-46
4.8. Analysis and Identification of the Active Compound
4.9. Antimicrobial Susceptibility Testing of the Active Compound
4.10. Cytotoxicity Assay
4.11. Genome Sequencing
4.12. Genome Assembly and Mining
4.13. Genome BGC Comparison
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Resistance, T.R.o.A. Tackling Drug-Resistant Infections Globally: Final Report and Recommendations. Available online: https://amr-review.org/sites/default/files/160525_Final%20paper_with%20cover.pdf (accessed on 17 July 2019).
- Coates, A.R.; Halls, G.; Hu, Y. Novel classes of antibiotics or more of the same? Br. J. Pharmacol. 2011, 163, 184–194. [Google Scholar] [CrossRef]
- CDC. Antibiotic Resistance Threats in the United States, 2019; U.S. Department of Health and Human Services: Atlanta, GA, USA, 2019.
- Medema, M.H.; Blin, K.; Cimermancic, P.; de Jager, V.; Zakrzewski, P.; Fischbach, M.A.; Weber, T.; Takano, E.; Breitling, R. antiSMASH: Rapid identification, annotation and analysis of secondary metabolite biosynthesis gene clusters in bacterial and fungal genome sequences. Nucleic Acids Res. 2011, 39, 339–346. [Google Scholar] [CrossRef]
- Tracanna, V.; de Jong, A.; Medema, M.H.; Kuipers, O.P. Mining prokaryotes for antimicrobial compounds: From diversity to function. FEMS Microbiol. Rev. 2017, 41, 417–429. [Google Scholar] [CrossRef] [PubMed]
- Taylor, M.W.; Radax, R.; Steger, D.; Wagner, M. Sponge-associated microorganisms: Evolution, ecology, and biotechnological potential. Microbiol. Mol. Biol. Rev. 2007, 71, 295–347. [Google Scholar] [CrossRef] [PubMed]
- Webster, N.S.; Taylor, M.W. Marine sponges and their microbial symbionts: Love and other relationships. Environ. Microbiol. 2012, 14, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Mehbub, M.F.; Lei, J.; Franco, C.; Zhang, W. Marine sponge derived natural products between 2001 and 2010: Trends and opportunities for discovery of bioactives. Mar. Drugs 2014, 12, 4539–4577. [Google Scholar] [CrossRef]
- Gerwick, W.H.; Fenner, A.M. Drug discovery from marine microbes. Microb. Ecol. 2013, 65, 800–806. [Google Scholar] [CrossRef]
- Williams, S.E.; Stennett, H.L.; Back, C.R.; Tiwari, K.; Ojeda Gomez, J.; Challand, M.R.; Hendry, K.R.; Spencer, J.; Essex-Lopresti, A.E.; Willis, C.L.; et al. The Bristol Sponge Microbiome Collection: A unique repository of deep-sea microorganisms and associated natural products. Antibiotics 2020, 9, 509. [Google Scholar] [CrossRef]
- Van der Meij, A.; Worsley, S.F.; Hutchings, M.I.; van Wezel, G.P. Chemical ecology of antibiotic production by actinomycetes. FEMS Microbiol. Rev. 2017, 41, 392–416. [Google Scholar] [CrossRef]
- Igarashi, Y.; Higuchi, H.; Oki, T.; Furumai, T. NMR analysis of quinocycline antibiotics: Structure determination of kosinostatin, an antitumor substance from Micromonospora sp. TP-A0468. J. Antibiot. 2002, 55, 134–140. [Google Scholar] [CrossRef]
- Celmer, W.D.; Murai, K.; Rao, K.V.; Tanner, F.W., Jr.; Marsh, W.S. The quinocycline complex. I. Isolation and characterization. Antibiot. Annu. 1957, 5, 484–492. [Google Scholar] [PubMed]
- Furumai, T.; Igarashi, Y.; Higuchi, H.; Saito, N.; Oki, T. Kosinostatin, a quinocycline antibiotic with antitumor activity from Micromonospora sp. TP-A0468. J. Antibiot. 2002, 55, 128–133. [Google Scholar] [CrossRef]
- Tulinsky, A. Structure of isoquinocycline A. X-Ray crystallographic determination. J. Am. Chem. Soc. 1964, 86, 5368. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Auch, A.F.; Klenk, H.-P.; Göker, M. Genome sequence-based species delimitation with confidence intervals and improved distance functions. BMC Bioinform. 2013, 14, 60. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Göker, M. TYGS is an automated high-throughput platform for state-of-the-art genome-based taxonomy. Nat. Commun. 2019, 10, 2182. [Google Scholar] [CrossRef] [PubMed]
- Lefort, V.; Desper, R.; Gascuel, O. FastME 2.0: A comprehensive, accurate, and fast distance-based phylogeny inference program. Mol. Biol. Evol. 2015, 32, 2798–2800. [Google Scholar] [CrossRef] [PubMed]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. antiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, 81–87. [Google Scholar] [CrossRef]
- Ma, H.M.; Zhou, Q.; Tang, Y.M.; Zhang, Z.; Chen, Y.S.; He, H.Y.; Pan, H.X.; Tang, M.C.; Gao, J.F.; Zhao, S.Y.; et al. Unconventional origin and hybrid system for construction of pyrrolopyrrole moiety in kosinostatin biosynthesis. Chem. Biol. 2013, 20, 796–805. [Google Scholar] [CrossRef]
- Carro, L.; Nouioui, I.; Sangal, V.; Meier-Kolthoff, J.P.; Trujillo, M.E.; Montero-Calasanz, M.d.C.; Sahin, N.; Smith, D.L.; Kim, K.E.; Peluso, P.; et al. Genome-based classification of micromonosporae with a focus on their biotechnological and ecological potential. Sci. Rep. 2018, 8, 525. [Google Scholar] [CrossRef] [PubMed]
- El-Naggar, M.Y. Kosinostatin, a major secondary metabolite isolated from the culture filtrate of Streptomyces violaceusniger strain HAL64. J. Microbiol. 2007, 45, 262–267. [Google Scholar] [PubMed]
- König, C.M.; Harms, K.; Koert, U. Stereoselective synthesis of methyl 7-dihydro-trioxacarcinoside B. Org. Lett. 2007, 9, 4777–4779. [Google Scholar] [CrossRef] [PubMed]
- Cordes, J.; Harms, K.; Koert, U. Synthesis of the isoquinocycline-pyrrolopyrrole substructure. Org. Lett. 2010, 12, 3808–3811. [Google Scholar] [CrossRef]
- Breuning, M.A.; Harms, K.; Koert, U. The imidato-alkenyllithium route for the synthesis of the isoquinocycline-pyrrolopyrrole substructure. Org. Lett. 2011, 13, 1402–1405. [Google Scholar] [CrossRef]
- Kitamura, M.; Kubo, K.; Yoshinaga, S.; Matsuzaki, H.; Ezaki, K.; Matsuura, T.; Matsuura, D.; Fukuzumi, N.; Araki, K.; Narasaki, M. Synthetic study of kosinostatin aglycone: Synthesis of BCDE rings using alkoxycarbonylmethylation of diazonaphthoquinone. Tetrahedron Lett. 2014, 55, 1653–1656. [Google Scholar] [CrossRef]
- Igarashi, Y. Antitumor Compounds from Actinomycetes in Deep-sea Water of Toyama Bay. In Handbook of Anticancer Drugs from Marine Origin; Kim, S.-K., Ed.; Springer: Cham, Switzerland, 2015; pp. 367–382. [Google Scholar]
- Quigley, G.J.; Wang, A.H.; Ughetto, G.; van der Marel, G.; van Boom, J.H.; Rich, A. Molecular structure of an anticancer drug-DNA complex: Daunomycin plus d(CpGpTpApCpG). Proc. Natl. Acad. Sci. USA 1980, 77, 7204–7208. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Completing bacterial genome assemblies with multiplex MinION sequencing. Microb. Genom. 2017, 3. [Google Scholar] [CrossRef]
- Zhou, Q.; Luo, G.C.; Zhang, H.Z.; Tang, G.L. Discovery of 16-demethylrifamycins by removing the predominant polyketide biosynthesis pathway in Micromonospora sp. strain TP-A0468. Appl. Environ. Microb. 2019, 85. [Google Scholar] [CrossRef]
- Wang, Z.; Fu, P.; Liu, P.; Wang, P.; Hou, J.; Li, W.; Zhu, W. New pyran-2-ones from alkalophilic actinomycete, Nocardiopsis alkaliphila sp. nov. YIM-80379. Chem. Biodivers. 2013, 10, 281–287. [Google Scholar] [CrossRef]
- Viegelmann, C.; Margassery, L.M.; Kennedy, J.; Zhang, T.; Brien, C.; Gara, F.; Morrissey, J.P.; Dobson, A.D.W. Metabolomic profiling and genomic study of a marine sponge-associated Streptomyces sp. Mar. Drugs 2014, 12, 3323–3351. [Google Scholar] [CrossRef]
- Intra, B.; Greule, A.; Bechthold, A.; Euanorasetr, J.; Paululat, T.; Panbangred, W. Thailandins A and B, new polyene macrolactone compounds isolated from Actinokineospora bangkokensis strain 44EHWT, possessing antifungal activity against anthracnose fungi and pathogenic yeasts. J. Agric. Food Chem. 2016, 64, 5171–5179. [Google Scholar] [CrossRef]
- Xu, G.; Kong, L.; Gong, R.; Xu, L.; Gao, Y.; Jiang, M.; Cai, Y.-S.; Hong, K.; Hu, Y.; Liu, P.; et al. Coordinated biosynthesis of the purine nucleoside antibiotics aristeromycin and coformycin in actinomycetes. Appl. Environ. Microb. 2018, 84, e01860-18. [Google Scholar] [CrossRef] [PubMed]
- Vértesy, L.; Aretz, W.; Knauf, M.; Markus, A.; Vogel, M.; Wink, J. Feglymycin, a novel inhibitor of the replication of the human immunodeficiency virus. Fermentation, isolation and structure elucidation. J. Antibiot. 1999, 52, 374–382. [Google Scholar] [CrossRef]
- Rausch, S.; Hänchen, A.; Denisiuk, A.; Löhken, M.; Schneider, T.; Süssmuth, R.D. Feglymycin is an inhibitor of the enzymes MurA and MurC of the peptidoglycan biosynthesis pathway. ChemBioChem 2011, 12, 1171–1173. [Google Scholar] [CrossRef] [PubMed]
- Olson, J.B.; Lord, C.C.; McCarthy, P.J. Improved recoverability of microbial colonies from marine sponge samples. Microb. Ecol. 2000, 40, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Sipkema, D.; Schippers, K.; Maalcke, W.J.; Yang, Y.; Salim, S.; Blanch, H.W. Multiple approaches to enhance the cultivability of bacteria associated with the marine sponge Haliclona (gellius) sp. Appl. Environ. Microb. 2011, 77, 2130–2140. [Google Scholar] [CrossRef] [PubMed]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Koren, S.; Walenz, B.P.; Berlin, K.; Miller, J.R.; Bergman, N.H.; Philippy, A.M. Canu: Scalable and accurate long-read assembly via adaptive k-mer weighting and repeat separation. Genome Res. 2017, 27, 722–736. [Google Scholar] [CrossRef]
- Peng, Y.; Leung, H.C.M.; Yiu, S.M.; Chin, F.Y.L. IDBA—A practical iterative de Bruijn graph de novo assembler. In Research in Computational Molecular Biology, Proceedings of the 14th Annual International Conference, RECOMB 2010, Lisbon, Portugal, 25–28 April 2010; Lecture Notes in Computer Science; Berger, B., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; Volume 6044. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef] [PubMed]
- Letunic, I.; Bork, P. Interactive Tree Of Life (iTOL) v4: Recent updates and new developments. Nucleic Acids Res. 2019, 47, W256–W259. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | MIC90 (µM) | |
|---|---|---|
| 2 | Sitafloxacin | |
| A. baumannii ATCC 17978 | 25 | 0.0781 |
| A. baumannii efflux KO a | 1.563 | <0.0195 |
| P. aeruginosa ATCC 33359 | >100.0 | 0.469 (±0.156) |
| P. aeruginosa efflux KO b | 12.5 | 0.156 |
| E. coli BW25113 | >100.0 | <0.0195 |
| E. coli efflux KO c | 9.375 (±3.125) | <0.0195 |
| E. coli IMP d | <0.1953 | <0.0195 |
| K. pneumoniae ATCC 10031 | 12.5 | <0.0195 |
| S. aureus SH1000 | 1.562 | 0.117 (±0.039) |
| Compound ID | IC50 (µM) | Cell Viability at 100 µM (%) |
|---|---|---|
| 2 | 13.3 | 0.0 |
| CONTROL | 22.5 | 0.0 |
| Illumina Only | Nanopore Only | Illumina/Nanopore Assembly | |
|---|---|---|---|
| Number of contigs | 637 | 196 | 1 |
| Total length (Mb) | 6.62 | 8.06 | 6.64 |
| Shortest contig | 123 nt | 1.03 kb | 6.64 Mb |
| Largest contig | 1.36 kb | 6.70 Mb | 6.64 Mb |
| Average coverage a | 36.3 | 313 | 290 |
| Coverage std dev | 19.9 | 137 | 113 |
| N50 (kb) | 22.3 | 6700 | 6640 |
| L50 | 92 | 1 | 1 |
| Mapped reads | 98.5% | 99.6% | 98.5% |
| Error rate | 1.27% | 10.8% | 0.44% |
| GC content | 72.4% | 72.3% | 72.5% |
| Region | Type | Cluster Size (kb) | Most Similar Known Cluster | Strain of Most Similar Known Cluster | Similarity a |
|---|---|---|---|---|---|
| 1 | T3PKS | 40.6 | Alkyl-O-dihydrogeranyl-methoxyhydroquinones | Actinoplanes missouriensis 431 | 71% |
| 2 | Terpene | 19.8 | Isorenieratene | Streptomyces argillaceus | 25% |
| 3 | Terpene | 16.8 | Phosphonoglycans | Glycomyces sp. NRRL B-16210 | 3% |
| 4 | NAGGN | 14.7 | No similar cluster | ||
| 5 | NRPS, T1PKS | 61.7 | Bleomycin | Streptomyces verticillus | 6% |
| 6 | Lanthipeptide | 21.5 | No similar cluster | ||
| 7 | NRPS, T1PKS | 66.3 | Nostopeptolide | Nostoc sp. GSV224 | 25% |
| 8 | Lanthipeptide | 21.6 | SapB | Streptomyces coelicolor A3(2) | 75% |
| 9 | T1PKS, NRPS-like | 214 | Rifamycin | Salinispora arenicola CNS-205 | 38% |
| 10 | NRPS, arylpolyene | 84.8 | Kedarcidin | Streptoalloteichus sp. ATCC 53650 | 13% |
| 11 | Terpene | 19.3 | Nocathiacin | Nocardia sp. ATCC 202099 | 4% |
| 12 | T2PKS | 71.2 | Formicamycins A-M | Streptomyces sp. KY5 | 13% |
| 13 | NRPS | 49.1 | Azicemicin | Kibdelosporangium sp. MJ126-NF4 | 13% |
| 14 | Siderophore | 11.8 | Desferrioxamine | Streptomyces sp. ID38640 | 100% |
| 15 | Oligosaccharide, terpene, lanthipeptide | 51.0 | Lobosamide | Micromonospora sp. RL09-050-HVF-A | 13% |
| 16 | NRPS, T3PKS | 114 | Feglymycin | Streptomyces sp. DSM 11171 | 63% |
| 17 | T2PKS, ectoine, NRPS, T1PKS, other | 158 | Kosinostatin | Micromonospora sp. TP-A0468 | 77% |
| 18 | Bacteriocin | 10.8 | Lymphostin | Salinispora tropica CNB-440 | 33% |
| 19 | Terpene | 21.0 | No similar cluster |
| Gene Group | Predicted Function | Colour in Figure 6 |
|---|---|---|
| kstA | Synthesise the anthracycline and modify its rings. | Red |
| kstB | Load nicotinic acid and convert it to a pyrrole. | Yellow |
| kstC | Convert the pyrrole and phosphoribosyl pyrophosphate to the pyrrolopyrrole. | Green |
| kstD | Synthesise the branched deoxy-octose and attach it to the anthracycline. | Blue |
| kstRg | Transcription factors that control the expression of the BGC. | Black |
| kstRs | Provide self-resistance to kosinostatin. | White |
| kstU | Not involved in kosinostatin biosynthesis. | Grey |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Back, C.R.; Stennett, H.L.; Williams, S.E.; Wang, L.; Ojeda Gomez, J.; Abdulle, O.M.; Duffy, T.; Neal, C.; Mantell, J.; Jepson, M.A.; et al. A New Micromonospora Strain with Antibiotic Activity Isolated from the Microbiome of a Mid-Atlantic Deep-Sea Sponge. Mar. Drugs 2021, 19, 105. https://doi.org/10.3390/md19020105
Back CR, Stennett HL, Williams SE, Wang L, Ojeda Gomez J, Abdulle OM, Duffy T, Neal C, Mantell J, Jepson MA, et al. A New Micromonospora Strain with Antibiotic Activity Isolated from the Microbiome of a Mid-Atlantic Deep-Sea Sponge. Marine Drugs. 2021; 19(2):105. https://doi.org/10.3390/md19020105
Chicago/Turabian StyleBack, Catherine R., Henry L. Stennett, Sam E. Williams, Luoyi Wang, Jorge Ojeda Gomez, Omar M. Abdulle, Thomas Duffy, Christopher Neal, Judith Mantell, Mark A. Jepson, and et al. 2021. "A New Micromonospora Strain with Antibiotic Activity Isolated from the Microbiome of a Mid-Atlantic Deep-Sea Sponge" Marine Drugs 19, no. 2: 105. https://doi.org/10.3390/md19020105
APA StyleBack, C. R., Stennett, H. L., Williams, S. E., Wang, L., Ojeda Gomez, J., Abdulle, O. M., Duffy, T., Neal, C., Mantell, J., Jepson, M. A., Hendry, K. R., Powell, D., Stach, J. E. M., Essex-Lopresti, A. E., Willis, C. L., Curnow, P., & Race, P. R. (2021). A New Micromonospora Strain with Antibiotic Activity Isolated from the Microbiome of a Mid-Atlantic Deep-Sea Sponge. Marine Drugs, 19(2), 105. https://doi.org/10.3390/md19020105