
Distribution of Tetrodotoxin in Pacific Oysters (Crassostrea gigas)


 , ,
, ,
Abstract
:1. Introduction
2. Results
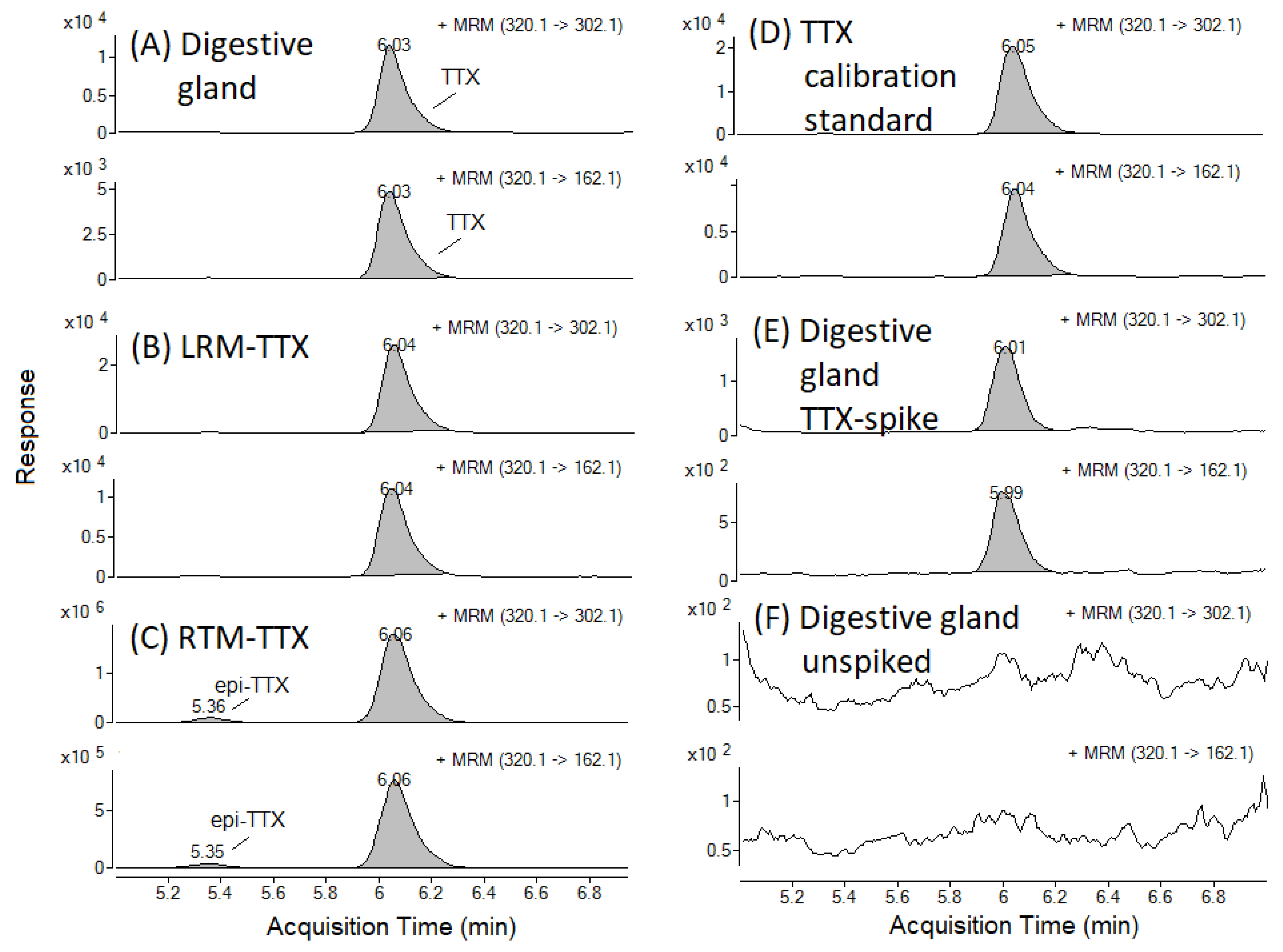
2.1. TTX Recovery in C. gigas Matrices and Matrix Effect on TTX Signal
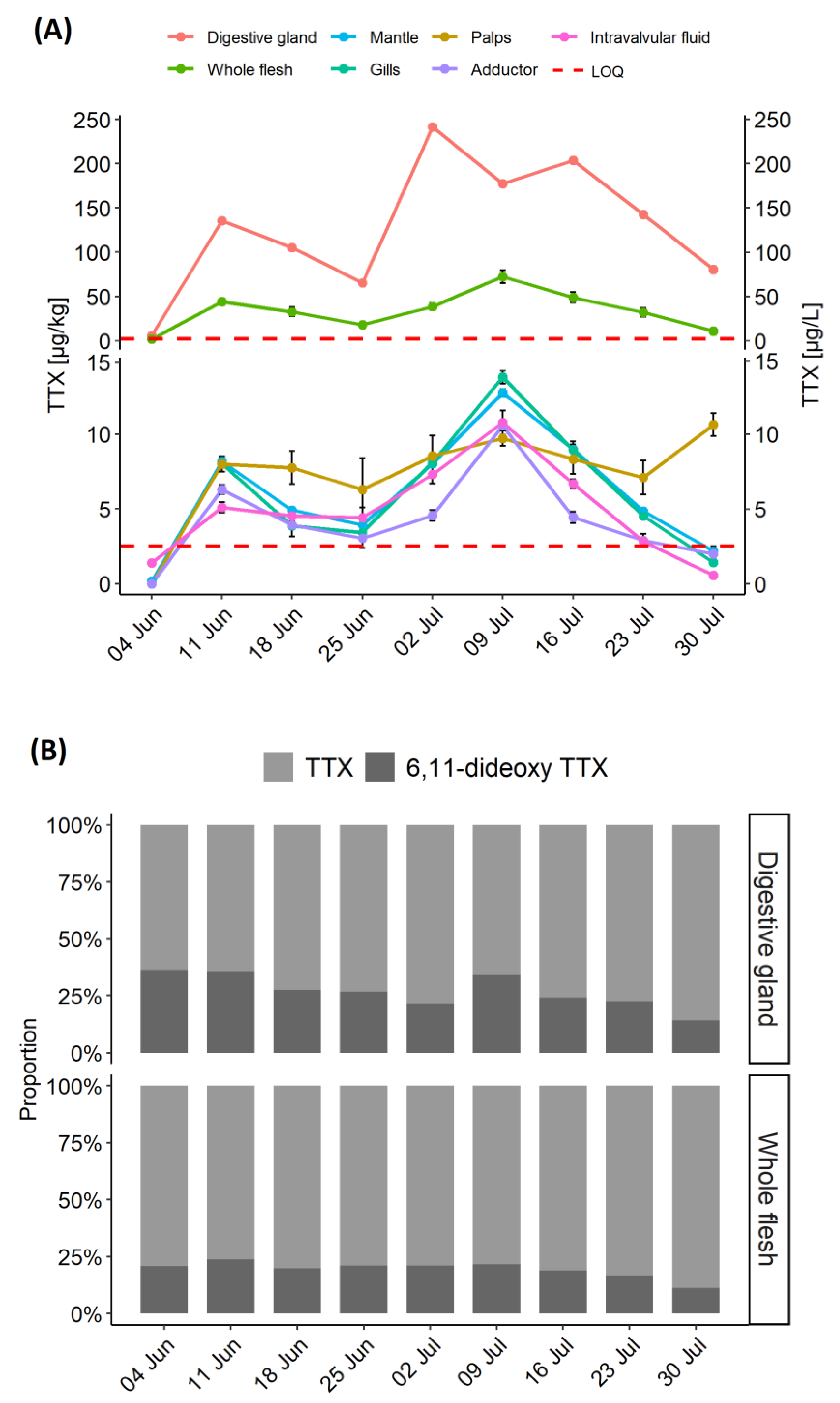
2.2. Time Series Study: TTX Concentrations in C. gigas Tissues
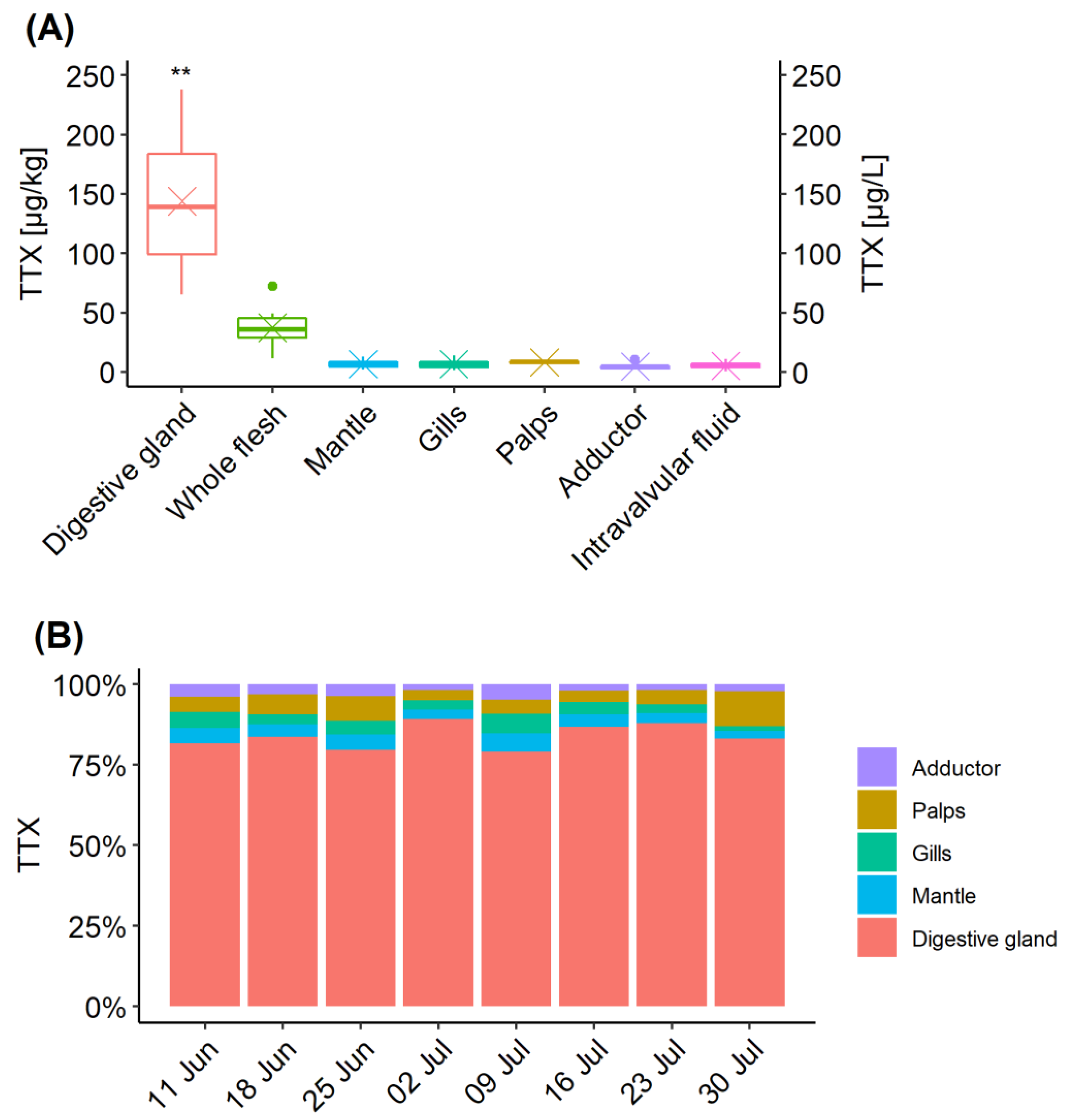
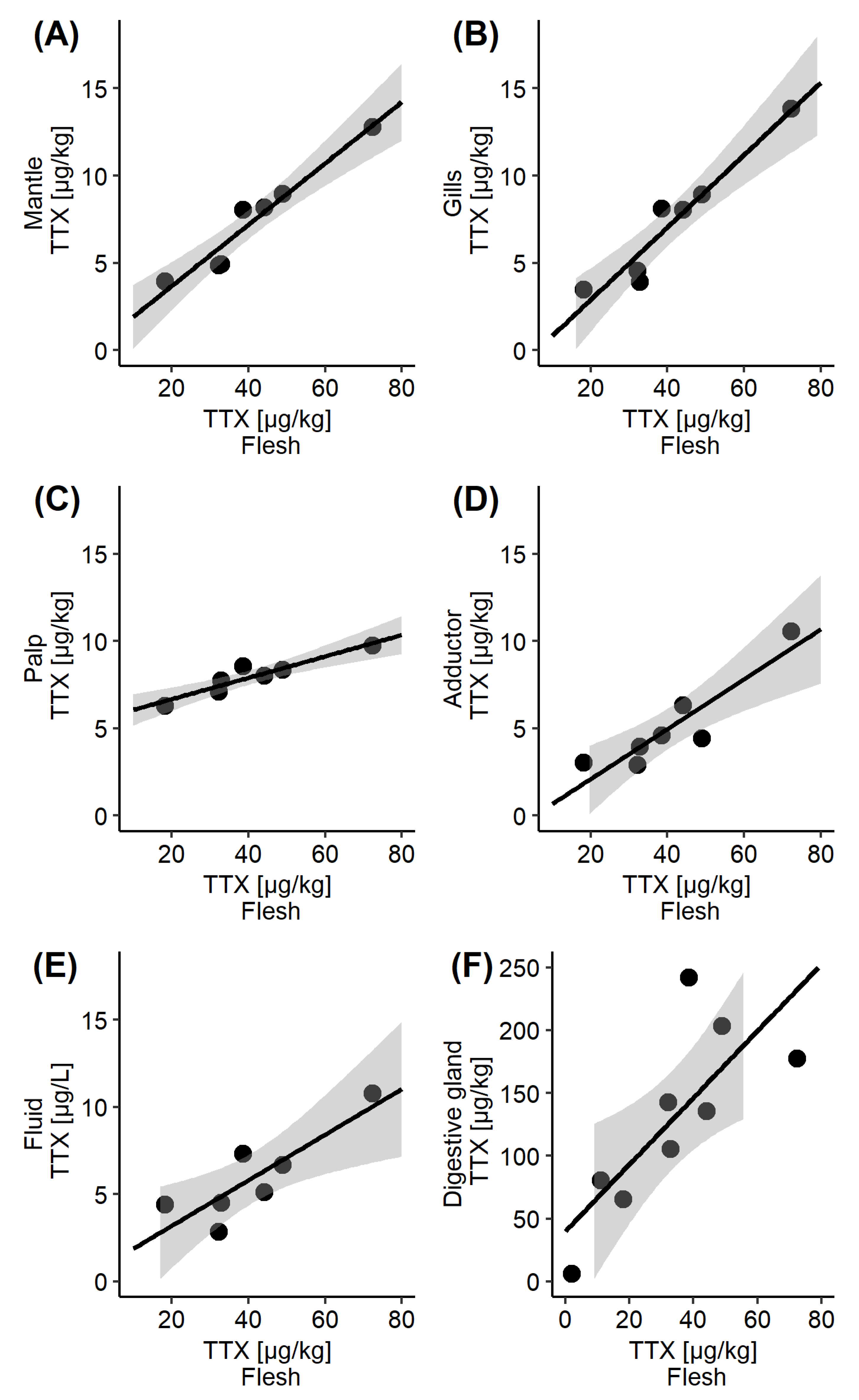
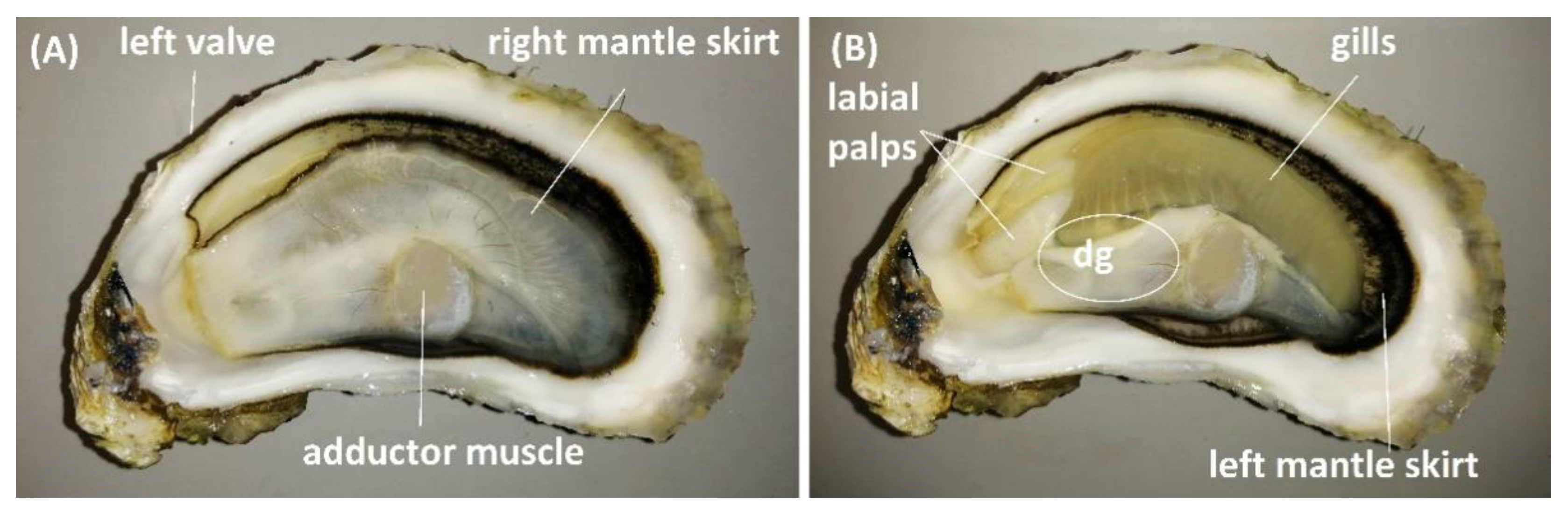
2.3. TTX Distribution in C. gigas Tissues
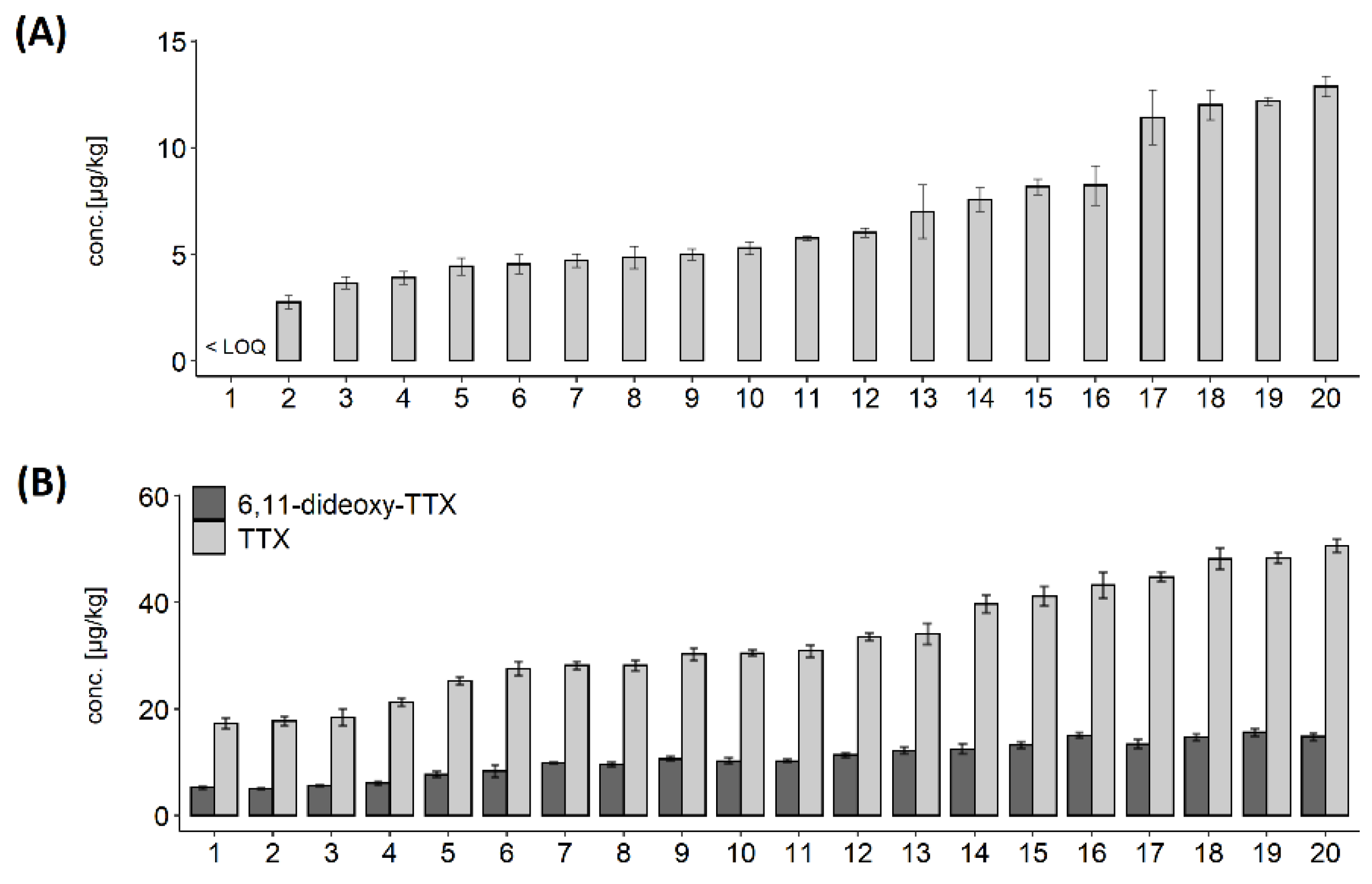
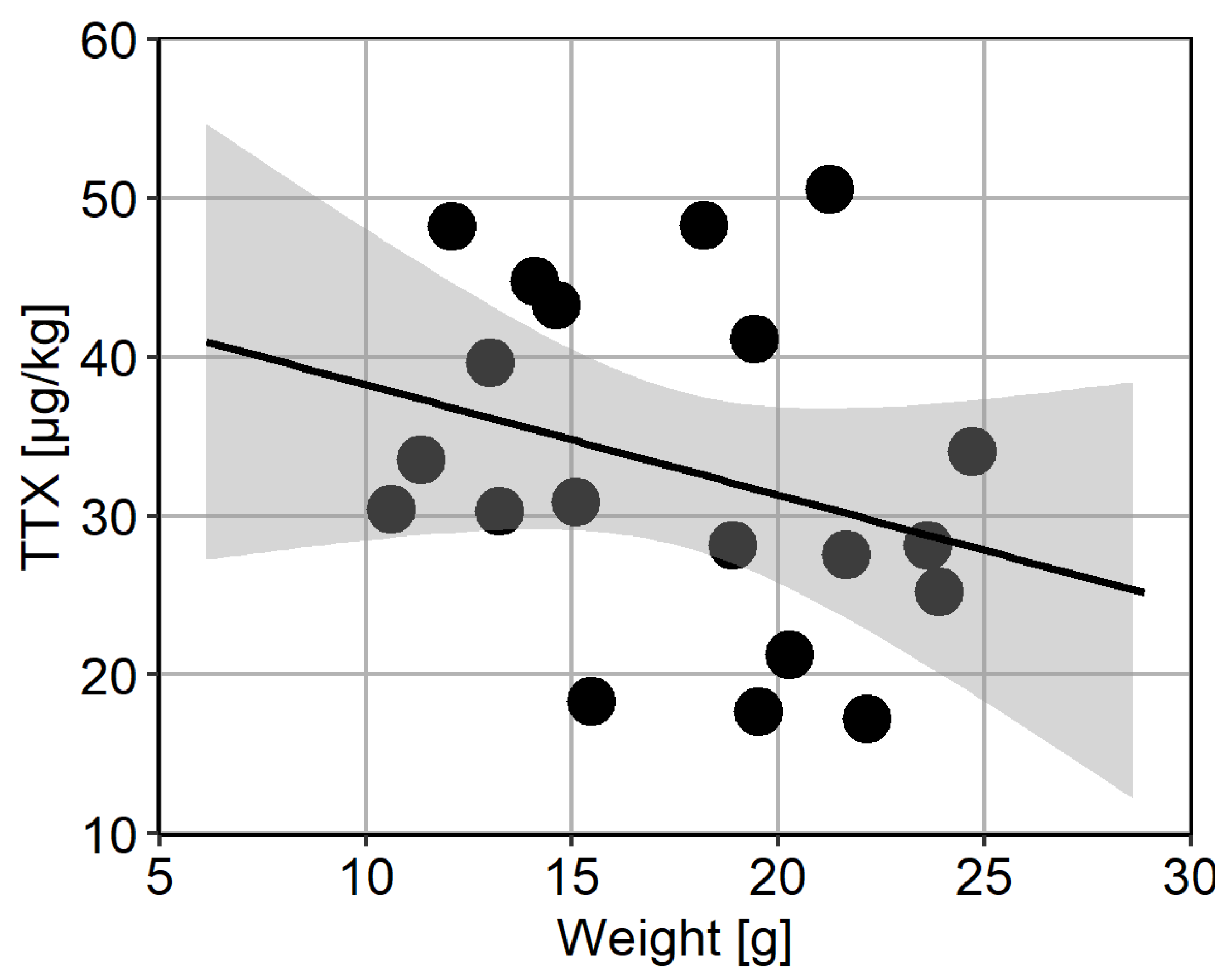
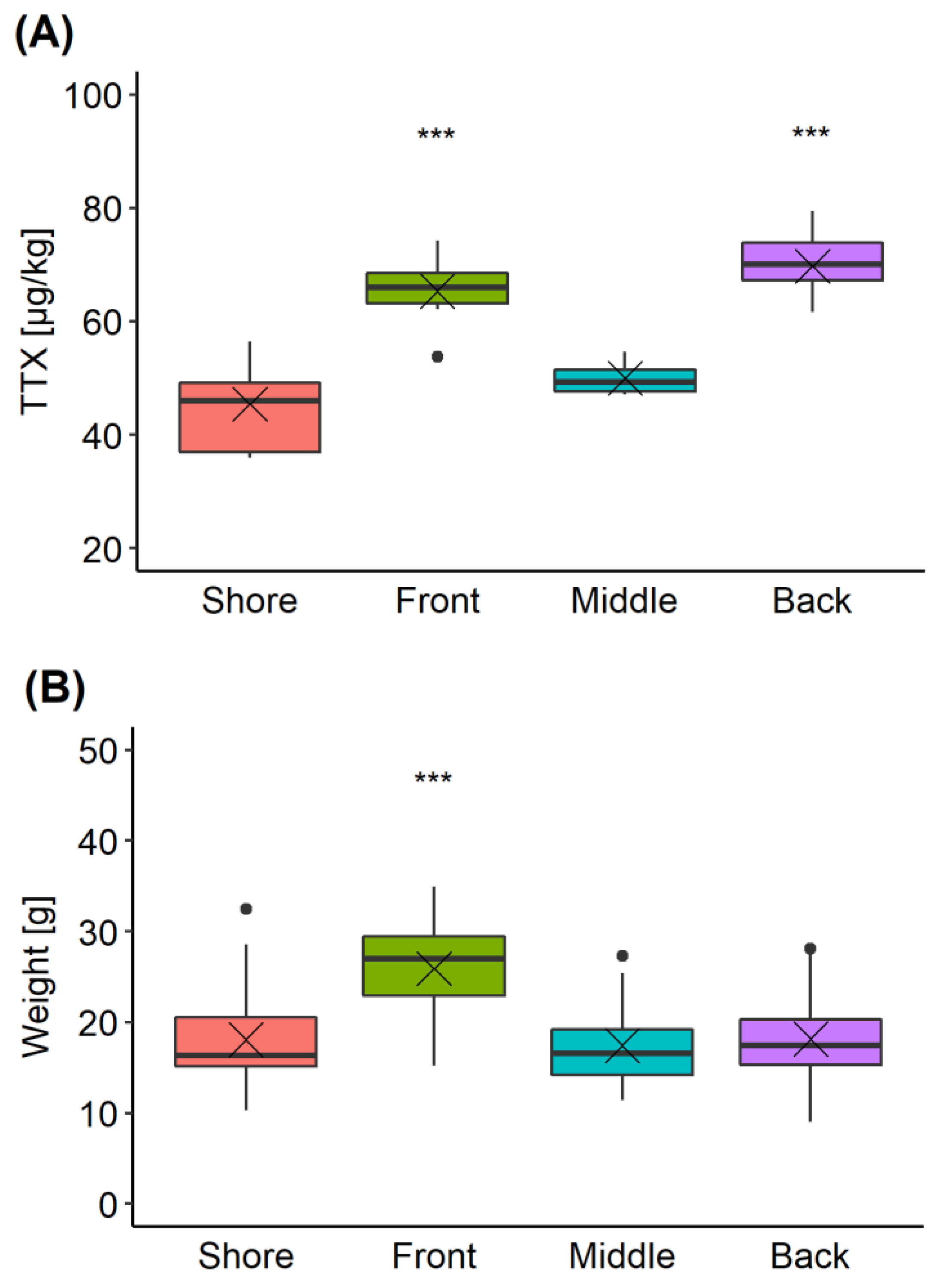
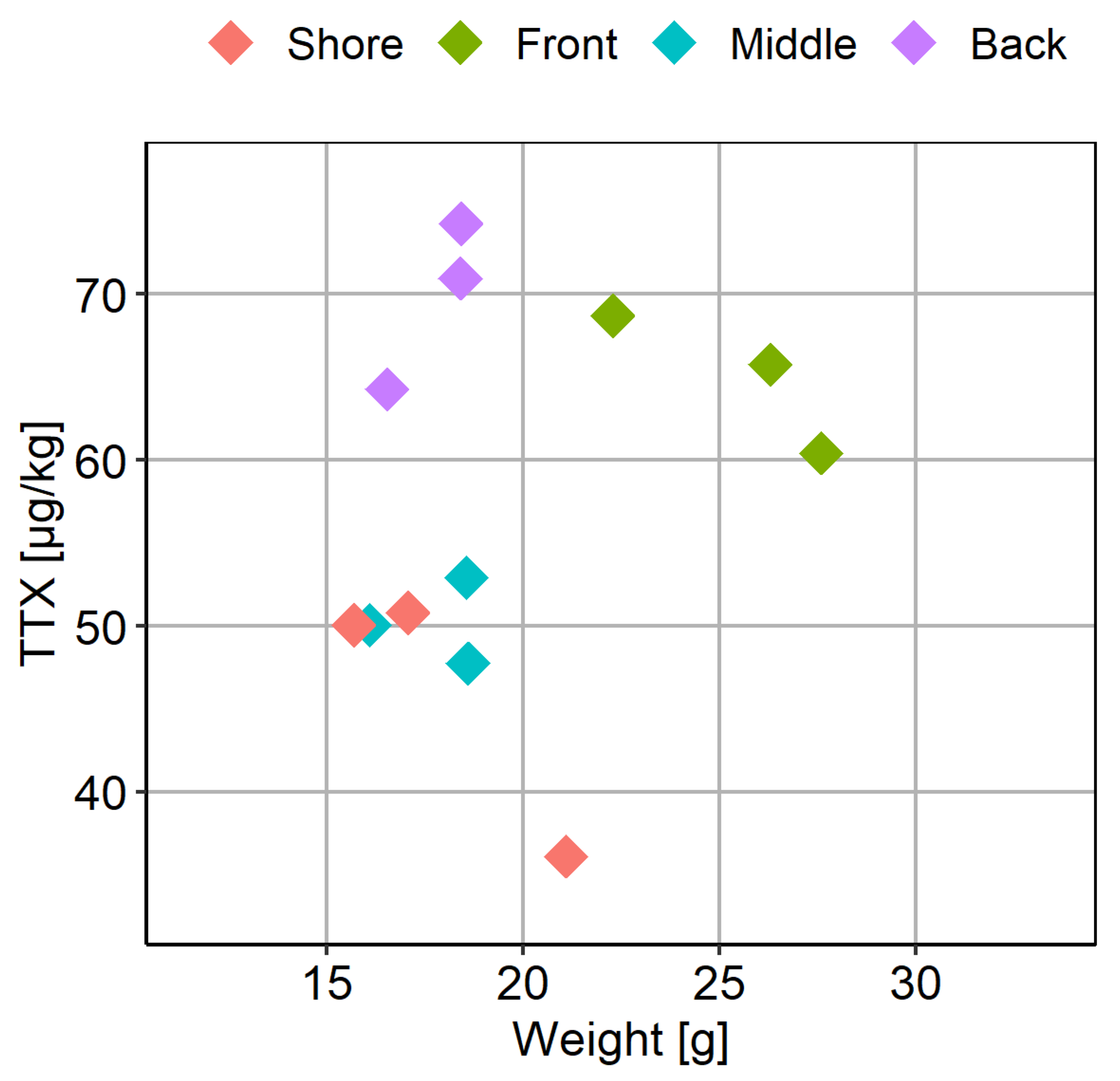
2.4. Intra-Population TTX Variability
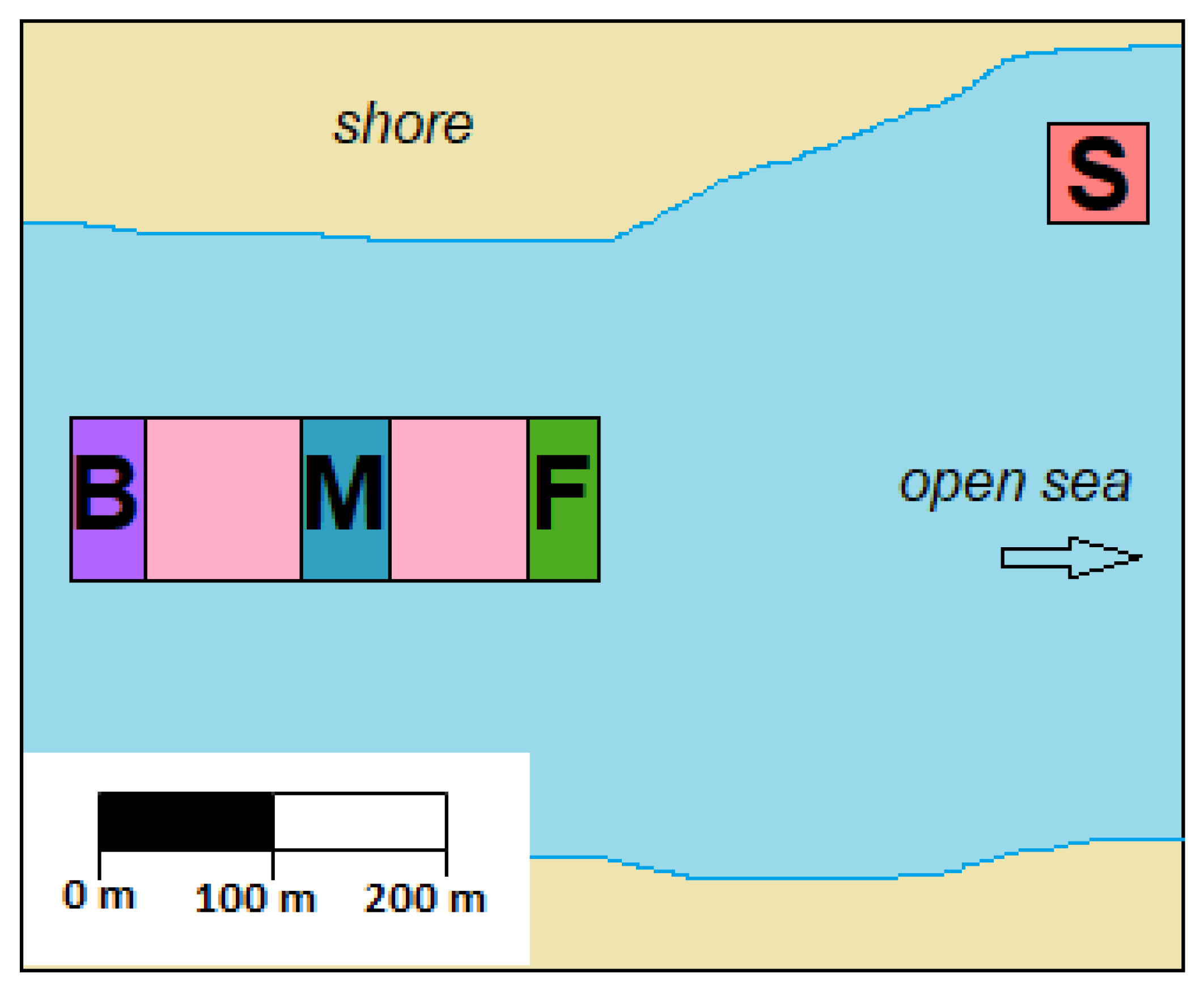
2.5. Inter-Population TTX Variability
3. Discussion
4. Materials and Methods
4.1. Reagents and Chemicals
- Laboratory Reference Material (LRM-TTX-PO), containing TTX (181 µg/kg) and a range of TTX analogues, was prepared by Cefas as described by Turner et al. (2017) [28];
- Non-certified mussel reference material (RM-TTX-Mus), containing TTX (2275 µg/kg of wet tissue homogenate) and a range of TTX analogues [29], was obtained from National Research Council Canada (NRCC, Halifax, Canada). After 0.35 ± 0.05 g of the freeze-dried powder was reconstituted in 1.65 mL of deionised water, vortex mixed for 30 s and sonicated for 1 min in ultrasonic bath, it was subjected to TTX extraction procedure. This material is not currently commercially available, and therefore it was extracted only once, and the cleaned extract was diluted weekly to check the performance of the instrumental detection method;
- Retention time marker (RTM-TTX), used for qualitative identification of retention times of TTX and multiple TTX analogues, was prepared from freeze-dried tissue of the sea slug Pleurobranchaea maculata (Cawthron Natural Compounds, Nelson, New Zealand), as described by Turner et al. (2017) [28].
4.2. Toxin Analysis
4.2.1. Extraction, Clean-Up and Dilution
4.2.2. Liquid Chromatography—Tandem Mass Spectrometry
4.3. C. gigas Collection and Preparation
4.3.1. Time Series of Tetrodotoxins in C. gigas
4.3.2. TTX Recovery
4.3.3. Intra-Population Variability
4.3.4. Inter-Population Variability
4.4. Data Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Noguchi, T.; Onuki, K.; Arakawa, O. Tetrodotoxin Poisoning Due to Pufferfish and Gastropods, and Their Intoxication Mechanism. Int. Sch. Res. Netw. ISRN Toxicol. 2011, 2011, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Bane, V.; Lehane, M.; Dikshit, M.; O’Riordan, A.; Furey, A. Tetrodotoxin: Chemistry, toxicity, source, distribution and detection. Toxins (Basel) 2014, 6, 693–755. [Google Scholar] [CrossRef] [Green Version]
- Guardone, L.; Maneschi, A.; Meucci, V.; Gasperetti, L.; Nucera, D.; Armani, A. A Global Retrospective Study on Human Cases of Tetrodotoxin (TTX) Poisoning after Seafood Consumption. Food Rev. Int. 2019, 645–667. [Google Scholar] [CrossRef]
- Chau, R.; Kalaitzis, J.A.; Neilan, B.A. On the origins and biosynthesis of tetrodotoxin. Aquat. Toxicol. 2011, 104, 61–72. [Google Scholar] [CrossRef]
- Kodama, M.; Sato, S.; Ogata, T. Alexandrium tamarense as a source of Tetrodotoxin in the scallop Patinopecten yessoensis. In Proceedings of the Toxic Phytoplankton Blooms in the Sea, Proceeding of the 5th International Conference on Toxic Marine Phytoplankton, Newport, RI, USA, 28 October–1 November 1991; Elsevier: New York, NY, USA, 1993; pp. 401–406. [Google Scholar]
- Ogilvie, S.; Taylor, D.; McNabb, P.; Hamon, D.; Nathan, P.; Anderson, A. Tetrodotoxin in Kaimoana: Science and Mātauranga Mitigating Health Risks from a Lethal Neurotoxin; Rep. No. 2219; Cawthron Institute: Nelson, New Zealand, 2012; pp. 1–46. [Google Scholar]
- McNabb, P.S.; Taylor, D.I.; Ogilvie, S.C.; Wilkinson, L.; Anderson, A.; Hamon, D.; Wood, S.A.; Peake, B.M. First detection of tetrodotoxin in the bivalve Paphies australis by liquid chromatography coupled to triple quadrupole mass spectrometry with and without precolumn reaction. J. AOAC Int. 2014, 97, 325–333. [Google Scholar] [CrossRef]
- Biessy, L.; Smith, K.F.; Harwood, D.T.; Boundy, M.J.; Hawes, I.; Wood, S.A. Spatial variability and depuration of tetrodotoxin in the bivalve Paphies australis from New Zealand. Toxicon X 2019, 2, 100008. [Google Scholar] [CrossRef]
- Biessy, L.; Smith, K.F.; Boundy, M.J.; Webb, S.C.; Hawes, I.; Wood, S.A. Distribution of tetrodotoxin in the New Zealand Clam, Paphies Australis, established using immunohistochemistry and liquid chromatography-tandem quadrupole mass spectrometry. Toxins (Basel) 2018, 10, 282. [Google Scholar] [CrossRef] [Green Version]
- Numano, S.; Kudo, Y.; Cho, Y.; Konoki, K.; Yotsu-Yamashita, M. Temporal Variation of the Profile and Concentrations of Paralytic Shellfish Toxins and Tetrodotoxin in the Scallop, Patinopectedn yessoensis, Cultured in a Bay of East Japan. Mar. Drugs 2019, 17, 653. [Google Scholar] [CrossRef] [Green Version]
- Boundy, M.J.; Biessy, L.; Roughan, B.; Nicolas, J.; Harwood, D.T. Survey of Tetrodotoxin in New Zealand Bivalve Molluscan Shellfish over a 16-Month Period. Toxins (Basel) 2020, 12, 512. [Google Scholar] [CrossRef]
- Han, C.; Zhang, X.; Li, L.; Chen, S.; Yan, Z.; Gao, X.; Chang, J. Analysis and Evaluation of Tetrodotoxin in Coastal Aquatic Products of Zhejiang Province. J. Coast. Res. 2018, 83, 380. [Google Scholar] [CrossRef]
- Fernández-Ortega, J.F.; Morales’de los Santos, J.; Herrera-Gutiérrez, M.E.; Fernández-Sánchez, V.; Loureo, P.R.; Rancaño, A.A.; Téllez-Andrade, A. Seafood intoxication by tetrodotoxin: First case in europe. J. Emerg. Med. 2010, 39, 612–617. [Google Scholar] [CrossRef]
- Rodriguez, P.; Alfonso, A.; Vale, C.; Alfonso, C.; Vale, P.; Tellez, A.; Botana, L.M. First toxicity report of tetrodotoxin and 5,6,11-trideoxyTTX in the trumpet shell Charonia lampas lampas in Europe. Anal. Chem. 2008, 80, 5622–5629. [Google Scholar] [CrossRef]
- EFSA; Knutsen, H.K.; Alexander, J.; Barregård, L.; Bignami, M.; Brüschweiler, B.; Ceccatelli, S.; Cottrill, B.; Dinovi, M.; Edler, L.; et al. Risks for public health related to the presence of tetrodotoxin (TTX) and TTX analogues in marine bivalves and gastropods. EFSA J. 2017, 15, 4752. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.; Azevedo, J.; Rodriguez, P.; Alfonso, A.; Botana, L.M.; Vasconcelos, V. New gastropod vectors and tetrodotoxin potential expansion in temperate waters of the Atlantic Ocean. Mar. Drugs 2012, 10, 712–726. [Google Scholar] [CrossRef] [Green Version]
- Turner, A.D.; Powell, A.; Schofield, A.; Lees, D.N.; Baker-Austin, C. Detection of the pufferfish toxin tetrodotoxin in European bivalves, England, 2013 to 2014. Eurosurveillance 2015, 20, 21009. [Google Scholar] [CrossRef]
- Vlamis, A.; Katikou, P.; Rodriguez, I.; Rey, V.; Alfonso, A.; Papazachariou, A.; Zacharaki, T.; Botana, A.M.; Botana, L.M. First detection of tetrodotoxin in greek shellfish by UPLC-MS/MS potentially linked to the presence of the dinoflagellate Prorocentrum minimum. Toxins (Basel) 2015, 7, 1779–1807. [Google Scholar] [CrossRef] [Green Version]
- Turner, A.D.; Dhanji-Rapkova, M.; Coates, L.; Bickerstaff, L.; Milligan, S.; O’Neill, A.; Faulkner, D.; McEneny, H.; Baker-Austin, C.; Lees, D.N.; et al. Detection of Tetrodotoxin Shellfish Poisoning (TSP) toxins and causative factors in bivalve molluscs from the UK. Mar. Drugs 2017, 15, 277. [Google Scholar] [CrossRef] [Green Version]
- Gerssen, A.; Bovee, T.H.F.; Klijnstra, M.D.; Poelman, M.; Portier, L.; Hoogenboom, R.L.A.P. First Report on the Occurrence of Tetrodotoxins in Bivalve Mollusks in The Netherlands. Toxins (Basel) 2018, 10, 450. [Google Scholar] [CrossRef] [Green Version]
- Bordin, P.; Dall’Ara, S.; Tartaglione, L.; Antonelli, P.; Calfapietra, A.; Varriale, F.; Guiatti, D.; Milandri, A.; Dell’Aversano, C.; Arcangeli, G.; et al. First occurrence of tetrodotoxins in bivalve mollusks from Northern Adriatic Sea (Italy). Food Control 2021, 120, 107510. [Google Scholar] [CrossRef]
- Dell’Aversano, C.; Tartaglione, L.; Polito, G.; Dean, K.; Giacobbe, M.; Casabianca, S.; Capellacci, S.; Penna, A.; Turner, A.D. First detection of tetrodotoxin and high levels of paralytic shellfish poisoning toxins in shellfish from Sicily (Italy) by three different analytical methods. Chemosphere 2019, 215, 881–892. [Google Scholar] [CrossRef]
- Katikou, P. Public Health Risks Associated with Tetrodotoxin and Its Analogues in European Waters: Recent Advances after The EFSA Scientific Opinion. Toxins (Basel) 2019, 11, 240. [Google Scholar] [CrossRef] [Green Version]
- Biessy, L.; Boundy, M.J.; Smith, K.F.; Harwood, D.T.; Hawes, I.; Wood, S.A. Tetrodotoxin in marine bivalves and edible gastropods: A mini-review. Chemosphere 2019, 236, 124404. [Google Scholar] [CrossRef]
- Allison, S.; Hardy, M.; Hayward, K.; Cameron, T.C.; Underwood, G.J.C. Strongholds of Ostrea edulis populations in estuaries in Essex, SE England and their association with traditional oyster aquaculture: Evidence to support a MPA designation. J. Mar. Biol. Assoc. UK 2020, 100, 27–36. [Google Scholar] [CrossRef] [Green Version]
- Eurostat. Available online: https://ec.europa.eu/eurostat/data/database (accessed on 12 January 2021).
- STECF Scientific, Technical and Economic Committee for Fisheries (STECF). Economic Report of the EU Aquaculture Sector (STECF-18-19); Publications Office of the European Union: Luxemburg, 2018; ISBN 9789279235405. [Google Scholar]
- Turner, A.D.; Boundy, M.J.; Dhanji-Rapkova, M. Development and single-laboratory validation of a liquid chromatography tandem mass spectrometry method for quantitation of tetrodotoxin in mussels and oysters. J. AOAC Int. 2017, 100, 1469–1482. [Google Scholar] [CrossRef]
- Turner, A.D.; Dhanji-Rapkova, M.; Fong, S.Y.T.; Hungerford, J.; McNabb, P.S.; Boundy, M.J.; Harwood, D.T. Ultrahigh-Performance Hydrophilic Interaction Liquid Chromatography with Tandem Mass Spectrometry Method for the Determination of Paralytic Shellfish Toxins and Tetrodotoxin in Mussels, Oysters, Clams, Cockles, and Scallops: Collaborative Study. J. AOAC Int. 2020, 103, 533–562. [Google Scholar] [CrossRef]
- Ward, J.E.; Shumway, S.E. Separating the grain from the chaff: Particle selection in suspension- and deposit-feeding bivalves. J. Exp. Mar. Bio. Ecol. 2004, 300, 83–130. [Google Scholar] [CrossRef]
- Yotsu-Yamashita, M.; Sugimoto, A.; Terakawa, T.; Shoji, Y.; Miyazawa, T.; Yasumoto, T. Purification, characterization, and cDNA cloning of a novel soluble saxitoxin and tetrodotoxin binding protein from plasma of the puffer fish, Fugu pardalis. Eur. J. Biochem. 2001, 268, 5937–5946. [Google Scholar] [CrossRef]
- Nagashima, Y.; Yamamoto, K.; Shimakura, K.; Shiomi, K. A tetrodotoxin-binding protein in the hemolymph of shore crab Hemigrapsus sanguineus: Purification and properties. Toxicon 2002, 40, 753–760. [Google Scholar] [CrossRef]
- De Goeij, P.; Luttikhuizen, P.C.; Van Der Meer, J.; Piersma, T. Facilitation on an intertidal mudflat: The effect of siphon nipping by flatfish on burying depth of the bivalve Macoma balthica. Oecologia 2001, 126, 500–506. [Google Scholar] [CrossRef] [Green Version]
- Blanco, J.; Acosta, C.P.; Bermúdez de la Puente, M.; Salgado, C. Depuration and anatomical distribution of the amnesic shellfish poisoning (ASP) toxin domoic acid in the king scallop Pecten maximus. Aquat. Toxicol. 2002, 60, 111–121. [Google Scholar] [CrossRef]
- Blanco, J.; Acosta, C.P.; Mariño, C.; Muñiz, S.; Martín, H.; Moroño, Á.; Correa, J.; Arévalo, F.; Salgado, C. Depuration of domoic acid from different body compartments of the king scallop Pecten maximus grown in raft culture and natural bed. Aquat. Living Resour. 2006, 19, 257–265. [Google Scholar] [CrossRef]
- Bresnan, E.; Fryer, R.J.; Fraser, S.; Smith, N.; Stobo, L.; Brown, N.; Turrell, E. The relationship between Pseudo-nitzschia (Peragallo) and domoic acid in Scottish shellfish. Harmful Algae 2017, 63, 193–202. [Google Scholar] [CrossRef]
- Mafra, L.L.; Bricelj, V.M.; Fennel, K. Domoic acid uptake and elimination kinetics in oysters and mussels in relation to body size and anatomical distribution of toxin. Aquat. Toxicol. 2010, 100, 17–29. [Google Scholar] [CrossRef] [Green Version]
- Blogoslawski, W.J.; Stewart, M.E. Joint Sanitation Seminar on North Pacific Clams, Juneau, Alaska, Public Health Service. Mar. Biol. 1978, 45, 261–264. [Google Scholar] [CrossRef]
- Quayle, D. Animal detoxification. In Proceedings of the Joint Sanitation Seminar on North Pacific Clams, Juneau, AK, USA, 24–25 September 1965; Public Health Service: Juneau, AK, USA, 1965; pp. 24–25. [Google Scholar]
- Shumway, S. A review of the effects of algal bloooms on shellfish and aquaculture. J. World Aquac. Soc. 1990, 21, 65–104. [Google Scholar] [CrossRef]
- Møhlenberg, F.; Riisgård, H.U. Efficiency of particle retention in 13 species of suspension feeding bivalves. Ophelia 1978, 17, 239–246. [Google Scholar] [CrossRef]
- Kodama, M.; Sato, S.; Sakamoto, S.; Ogata, T. Occurrence of tetrodotoxin in Alexandrium tamarense, a causative dinoflagellate of paralytic shellfish poisoning. Toxicon 1996, 34, 1101–1105. [Google Scholar] [CrossRef]
- Rodríguez, I.; Alfonso, A.; Alonso, E.; Rubiolo, J.A.; Roel, M.; Vlamis, A.; Katikou, P.; Jackson, S.A.; Menon, M.L.; Dobson, A.; et al. The association of bacterial C 9 -based TTX-like compounds with Prorocentrum minimum opens new uncertainties about shellfish seafood safety. Sci. Rep. 2017, 7, 1–12. [Google Scholar] [CrossRef]
- Cognie, B.; Barillé, L.; Rincé, Y. Selective feeding of the oyster Crassostrea gigas fed on a natural microphytobenthos assemblage. Estuaries 2001, 24, 126–134. [Google Scholar] [CrossRef]
- White, A.W.; Shumway, S.E.; Nassif, J.; Whittaker, D.K. Variation in levels of paralytic shellfish toxins among individual shellfish. J. Shellfish Res. 1992, 11, 209. [Google Scholar]
- Álvarez, G.; Uribe, E.; Regueiro, J.; Martin, H.; Gajardo, T.; Jara, L.; Blanco, J. Depuration and anatomical distribution of domoic acid in the surf clam Mesodesma donacium. Toxicon 2015, 102, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Quayle, D.B. Paralytic shellfish poisoning in British Columbia. Fish. Res. Board Can. Bull. 1969, 168, 68. [Google Scholar]
- Aalvik, B.; Framstad, K. Assay and detoxification experiments with mytilotoxin in mussels (Mytilus edulis L.) from Nordasstraumen, western Norway, 1979 and 1980. Sarsia 1981, 66, 143–146. [Google Scholar] [CrossRef]
- Bogan, Y.M.; Harkin, A.L.; Gillespie, J.; Kennedy, D.J.; Hess, P.; Slater, J.W. The influence of size on domoic acid concentration in king scallop, Pecten maximus (L.). Harmful Algae 2007, 6, 15–28. [Google Scholar] [CrossRef]
- Bricelj, V.M.; Shumway, S.E. Paralytic shellfish toxins in bivalve molluscs: Occurrence, transfer kinetics, and biotransformation. Rev. Fish. Sci. 1998, 6, 315–383. [Google Scholar] [CrossRef]
- Moroño, A.; Franco, J.; Miranda, M.; Reyero, M.I.; Blanco, J. The effect of mussel size, temperature, seston volume, food quality and volume-specific toxin concentration on the uptake rate of PSP toxins by mussels (Mytilus galloprovincialis Lmk). J. Exp. Mar. Bio. Ecol. 2001, 257, 117–132. [Google Scholar] [CrossRef]
- Cefas Classification of Bivalve Mollusc Production Areas in England and Wales. Sanitary Survey Report. Available online: https://www.cefas.co.uk/data-and-publications/sanitary-surveys/england-and-wales/reports/ (accessed on 2 September 2020).
- Noguchi, T.; Arakawa, O.; Takatani, T. TTX accumulation in pufferfish. Comp. Biochem. Physiol.-Part D Genomics Proteomics 2006, 1, 145–152. [Google Scholar] [CrossRef]
- Magarlamov, T.Y.; Melnikova, D.I.; Chernyshev, A.V. Tetrodotoxin-producing bacteria: Detection, distribution and migration of the toxin in aquatic systems. Toxins (Basel) 2017, 9, 166. [Google Scholar] [CrossRef]
- Matsumura, K. Reexamination of tetrodotoxin production by bacteria. Appl. Environ. Microbiol. 1995, 61, 3468–3470. [Google Scholar] [CrossRef] [Green Version]
- Matsumura, K. No Ability To Produce Tetrodotoxin in Bacteria Letter to the Editor No Ability To Produce Tetrodotoxin in Bacteria. Appl. Environ. Microbiol. 2001, 67, 2393–2395. [Google Scholar] [CrossRef] [Green Version]
- Strand, M.; Hedström, M.; Seth, H.; McEvoy, E.G.; Jacobsson, E.; Göransson, U.; Andersson, H.S.; Sundberg, P. The bacterial (Vibrio alginolyticus) production of tetrodotoxin in the ribbon worm Lineus longissimus-Just a false positive? Mar. Drugs 2016, 14, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, A.D.; Fenwick, D.; Powell, A.; Dhanji-Rapkova, M.; Ford, C.; Hatfield, R.G.; Santos, A.; Martinez-Urtaza, J.; Bean, T.P.; Baker-Austin, C.; et al. New invasive Nemertean species (Cephalothrix Simula) in England with high levels of tetrodotoxin and a microbiome linked to toxin metabolism. Mar. Drugs 2018, 16, 452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, S.; Harada, R.M.; Defelice, S.V.; Bienfang, P.K.; Li, Q.X. Bacterial production of tetrodotoxin in the pufferfish Arothron hispidus. Nat. Prod. Res. 2009, 23, 1630–1640. [Google Scholar] [CrossRef] [PubMed]
- Melnikova, D.I.; Vlasenko, A.E.; Magarlamov, T.Y. Stable Tetrodotoxin Production by Bacillus sp. Strain 1839. Mar. Drugs 2019, 17, 704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaelli, P.M.; Theis, K.R.; Williams, J.E.; O’Connell, L.A.; Foster, J.A.; Eisthen, H.L. The skin microbiome facilitates adaptive tetrodotoxin production in poisonous newts. Elife 2020, 9, e53898. [Google Scholar] [CrossRef]
- Palmer, R.E.; Williams, L.G. Effect of particle concentration on filtration efficiency of the bay scallop Argopecten irradians and the oyster Crassostrea virginica. Ophelia 1980, 19, 163–174. [Google Scholar] [CrossRef]
- Cabello, A.E.; Espejo, R.T.; Romero, J. Tracing Vibrio parahaemolyticus in oysters (Tiostrea chilensis) using a Green Fluorescent Protein tag. J. Exp. Mar. Bio. Ecol. 2005, 327, 157–166. [Google Scholar] [CrossRef]
- Matsuyama, Y.; Shumway, S. Impacts of harmful algal blooms on shellfisheries aquaculture. In New Technologies in Shellfisheries Aquaculture; Woodhead Publishing: Sawston, UK, 2009; pp. 580–609. [Google Scholar]
- Ramanan, R.; Kim, B.H.; Cho, D.H.; Oh, H.M.; Kim, H.S. Algae-bacteria interactions: Evolution, ecology and emerging applications. Biotechnol. Adv. 2016, 34, 14–29. [Google Scholar] [CrossRef] [Green Version]
- Lima-Mendez, G.; Faust, K.; Henry, N.; Decelle, J.; Colin, S.; Carcillo, F.; Chaffron, S.; Ignacio-Espinosa, J.C.; Roux, S.; Vincent, F.; et al. Determinants of community structure in the global plankton interactome. Ocean Plankt. 2015, 348, 1262073. [Google Scholar] [CrossRef] [Green Version]
- Hattenrath-Lehmann, T.K.; Gobler, C.J. Identification of unique microbiomes associated with harmful algal blooms caused by Alexandrium fundyense and Dinophysis acuminata. Harmful Algae 2017, 68, 17–30. [Google Scholar] [CrossRef]
- R Core Team R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. Available online: http://www.R-project.org/ (accessed on 15 May 2020).











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Acidification | pH after Acidification | Heating (5 Min Boil) | TTX Recovery Mean ± s.d. and (RSD) 1 |
|---|---|---|---|---|
| A | no | 7.38 | yes | 57 ± 3 (6%) |
| B | no | no | 91 ± 6 (7%) | |
| C | Glacial acetic acid | 3.75 | yes | 87 ± 3 (4%) |
| D | Glacial acetic acid | no | 84 ± 5 (5%) | |
| E | 1% acetic acid | 3.22 | yes | 66 ± 3 (5%) |
| F | 1% acetic acid | no | 68 ± 6 (8%) |
| Matrix | r2 | MM/Solvent Slope (%) | TTX Recovery Mean ± s.d. and (RSD) 1 |
|---|---|---|---|
| Intravalvular fluid | 0.997 | 104 | 91 ± 6 (7%) |
| Mantle | 0.999 | 100 | 86 ± 8 (9%) |
| Gills | 0.996 | 123 | 81 ± 6 (7%) |
| Whole flesh 2 | 0.995 | 117 | 76 ± 6 (8%) |
| Whole flesh 3 | 0.998 | 74 | 66 ± 2 (4%) |
| Adductor | 0.999 | 83 | 68 ± 3 (5%) |
| Digestive gland | 1.000 | 55 | 60 ± 5 (8%) |
| Palp | 0.997 | 55 | 37 ± (12%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dhanji-Rapkova, M.; Turner, A.D.; Baker-Austin, C.; Huggett, J.F.; Ritchie, J.M. Distribution of Tetrodotoxin in Pacific Oysters (Crassostrea gigas). Mar. Drugs 2021, 19, 84. https://doi.org/10.3390/md19020084
Dhanji-Rapkova M, Turner AD, Baker-Austin C, Huggett JF, Ritchie JM. Distribution of Tetrodotoxin in Pacific Oysters (Crassostrea gigas). Marine Drugs. 2021; 19(2):84. https://doi.org/10.3390/md19020084
Chicago/Turabian StyleDhanji-Rapkova, Monika, Andrew D. Turner, Craig Baker-Austin, Jim F. Huggett, and Jennifer M. Ritchie. 2021. "Distribution of Tetrodotoxin in Pacific Oysters (Crassostrea gigas)" Marine Drugs 19, no. 2: 84. https://doi.org/10.3390/md19020084
APA StyleDhanji-Rapkova, M., Turner, A. D., Baker-Austin, C., Huggett, J. F., & Ritchie, J. M. (2021). Distribution of Tetrodotoxin in Pacific Oysters (Crassostrea gigas). Marine Drugs, 19(2), 84. https://doi.org/10.3390/md19020084