
Stereochemical Determination of Fistularins Isolated from the Marine Sponge Ecionemia acervus and Their Regulatory Effect on Intestinal Inflammation

 and
and 
Abstract
:1. Introduction
2. Results and Discussion
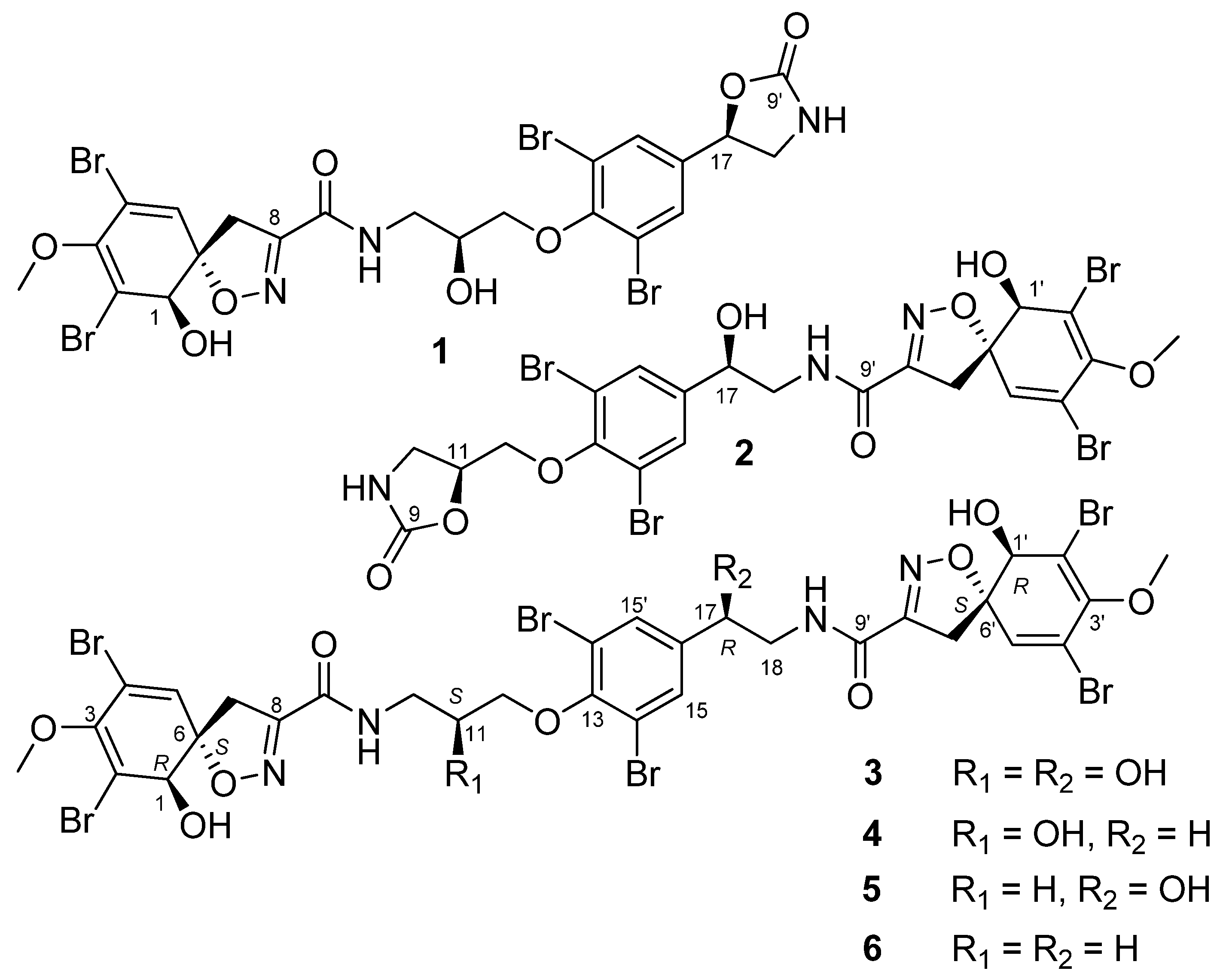
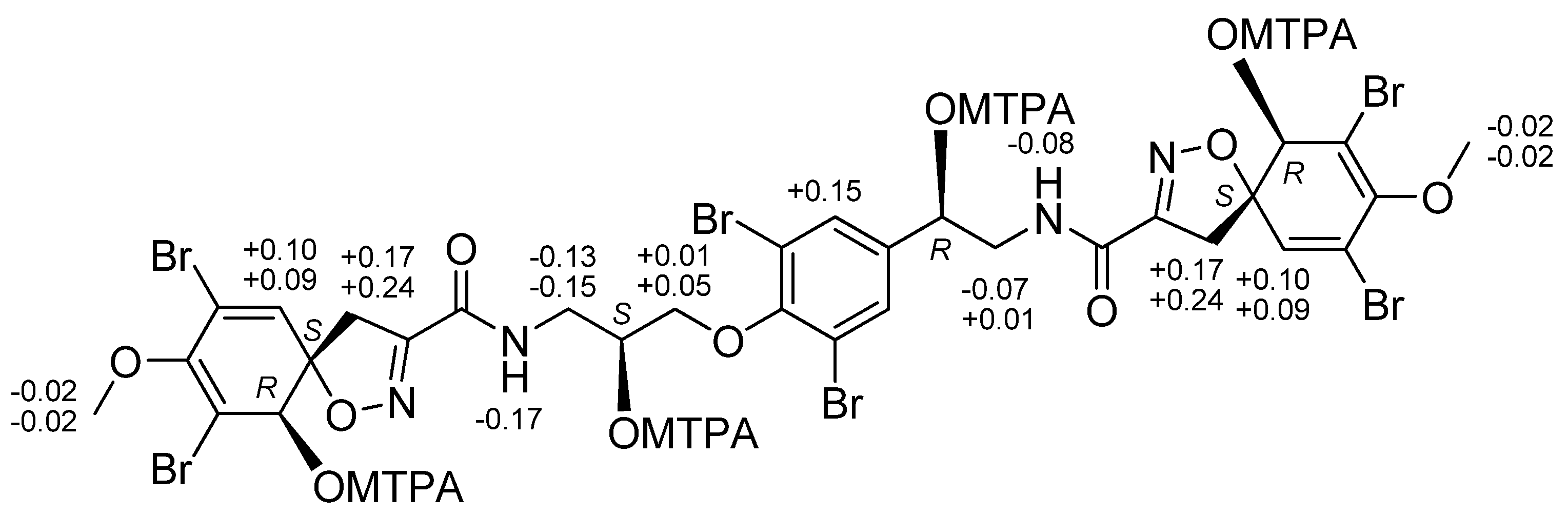
2.1. Structures of Compounds 1–6 Isolated from Ecionemia acervus
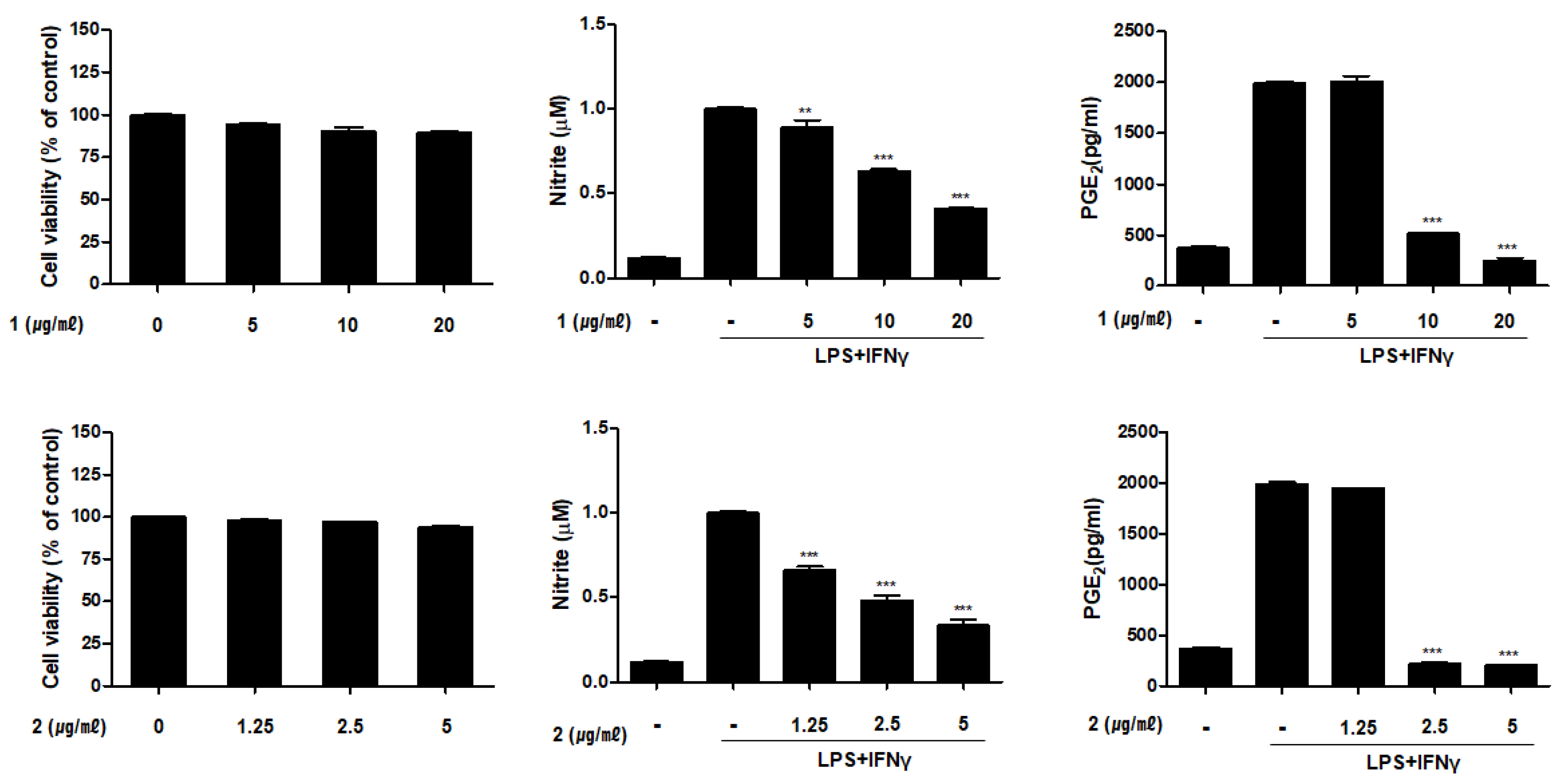
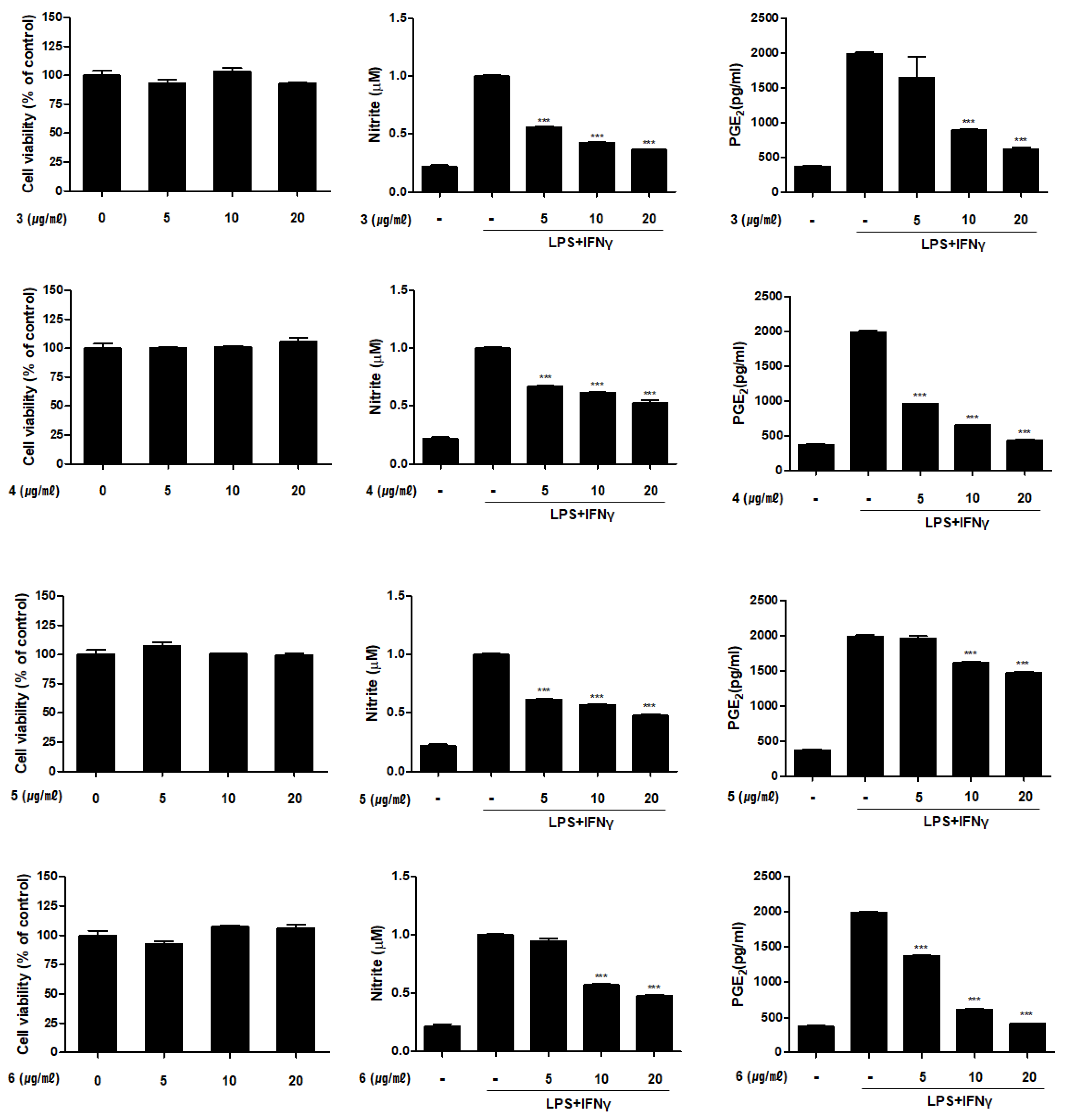
2.2. Inhibitory Activities of Compounds 1–6 on the Productioin of NO and PGE2 in THP-1 Macrophages Activated with LPS and IFNγ
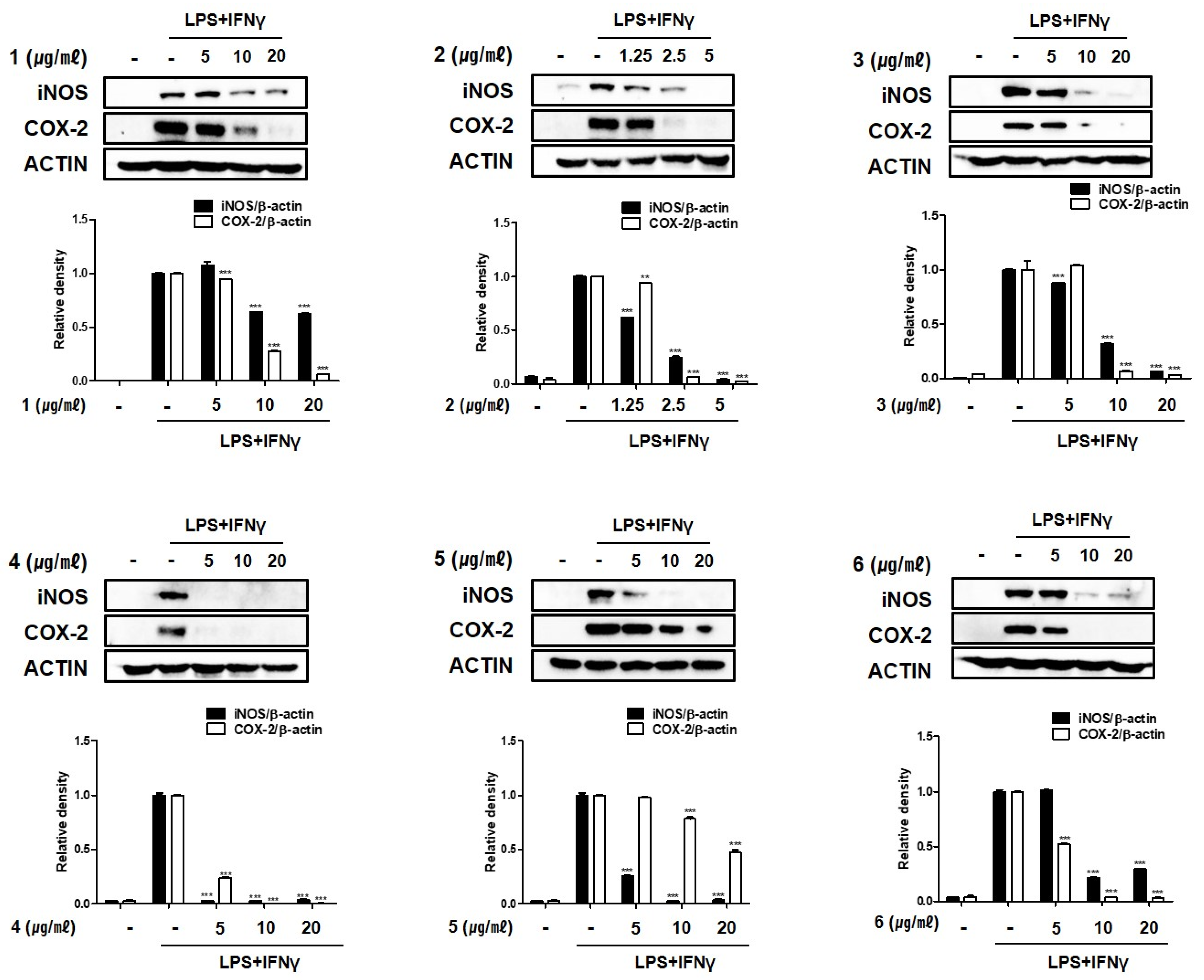
2.3. Compounds 1–6 Inhibit the Expression of Pro-Inflammatory Proteins iNOS and COX-2 in THP-1 Macrophages Co-Cultured with Caco-2 Cells
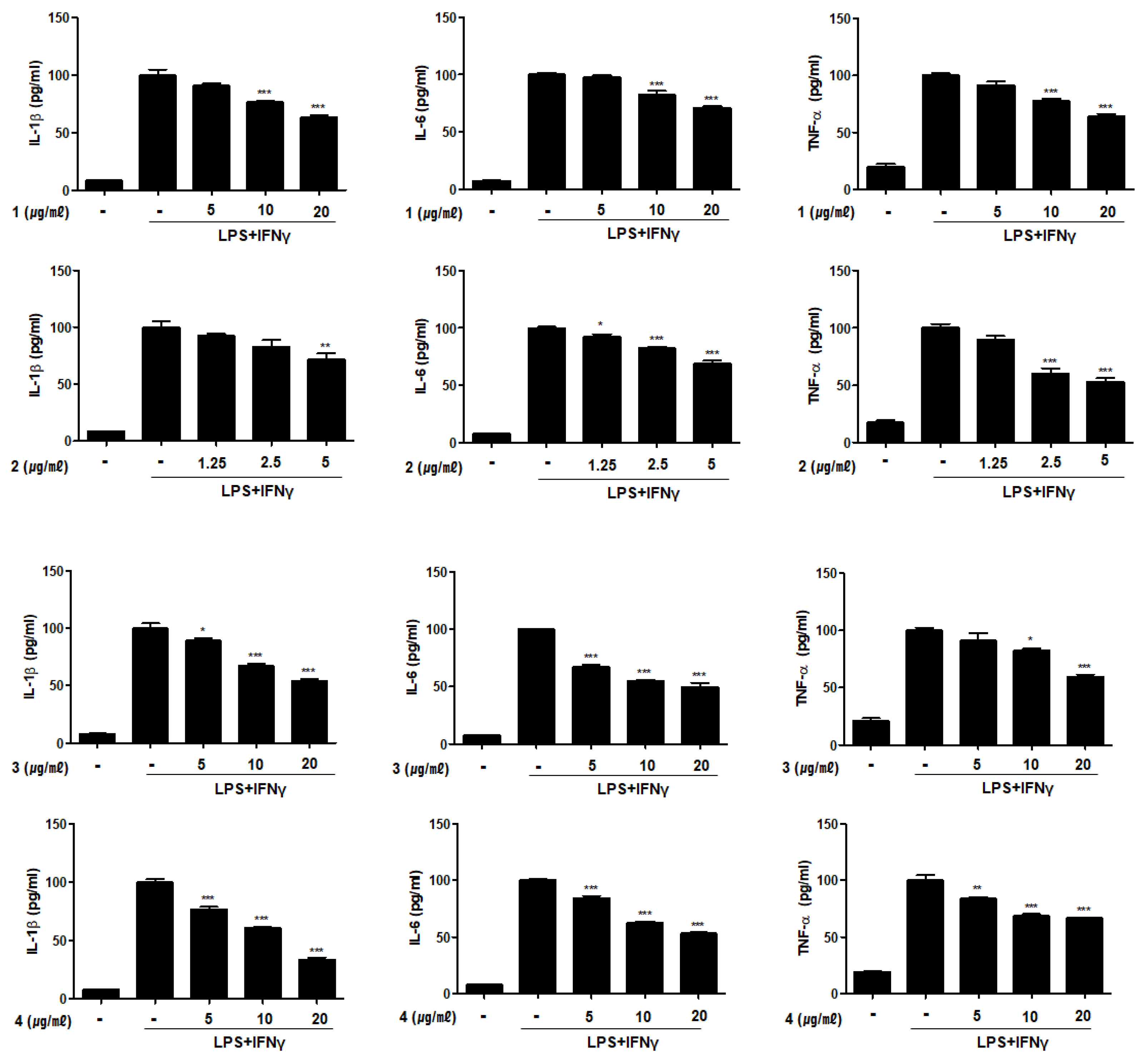
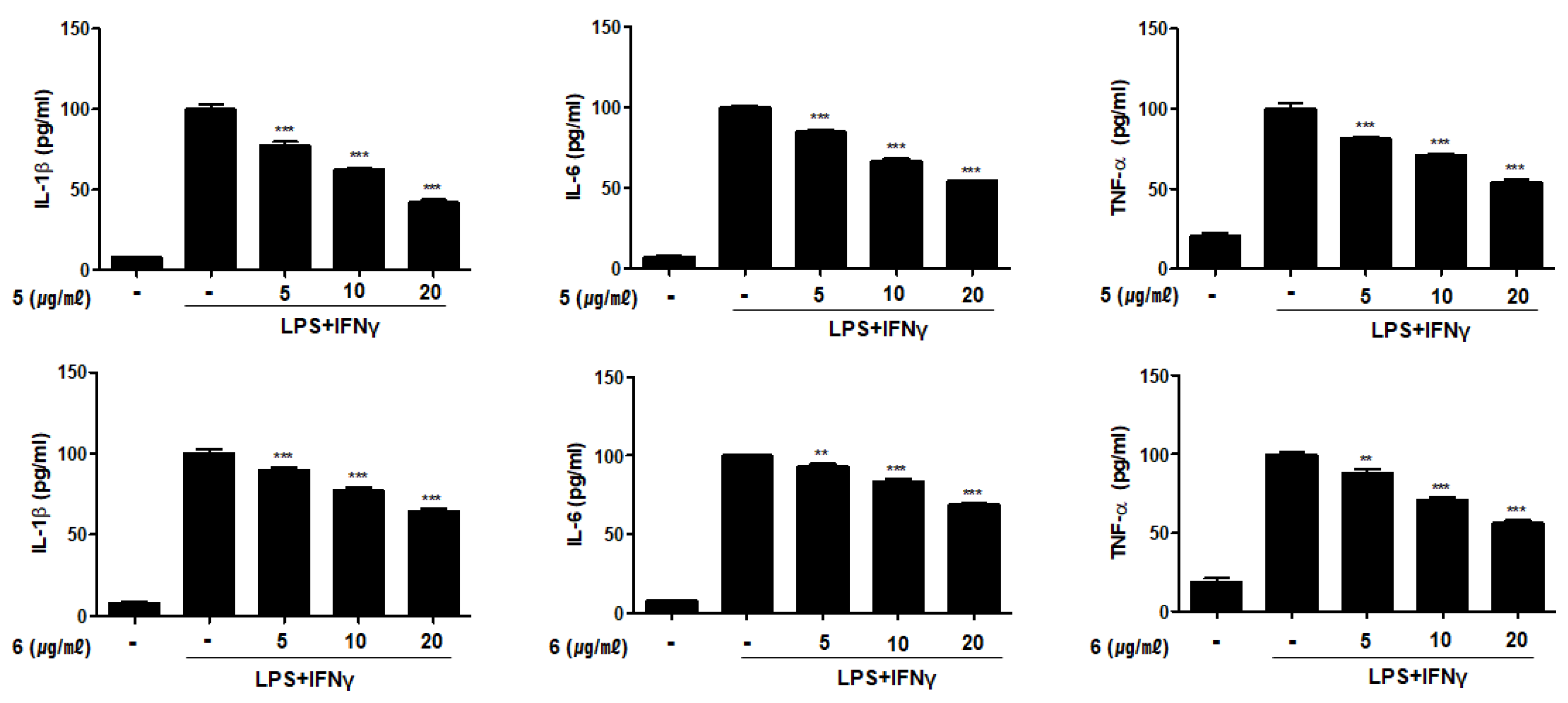
2.4. Compounds 1–6 Inhibit the Production of Pro-Inflammatory Cytokines in THP-1 Macrophages Co-Cultured with Caco-2 Cells
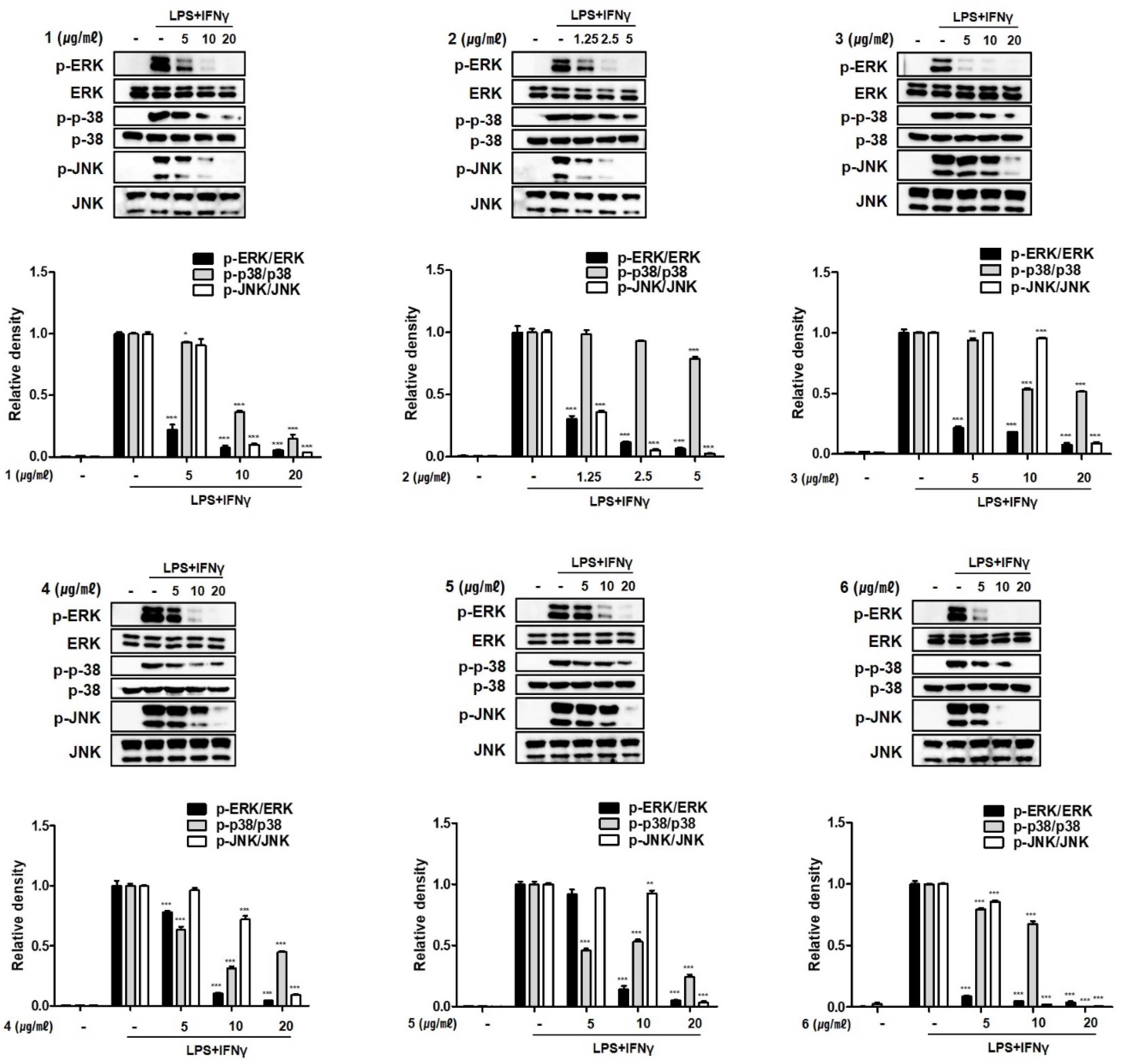
2.5. Compounds 1–6 Inhibit MAPK Phosphorylation in THP-1 Macrophages Co-Cultured with Caco-2 Cells
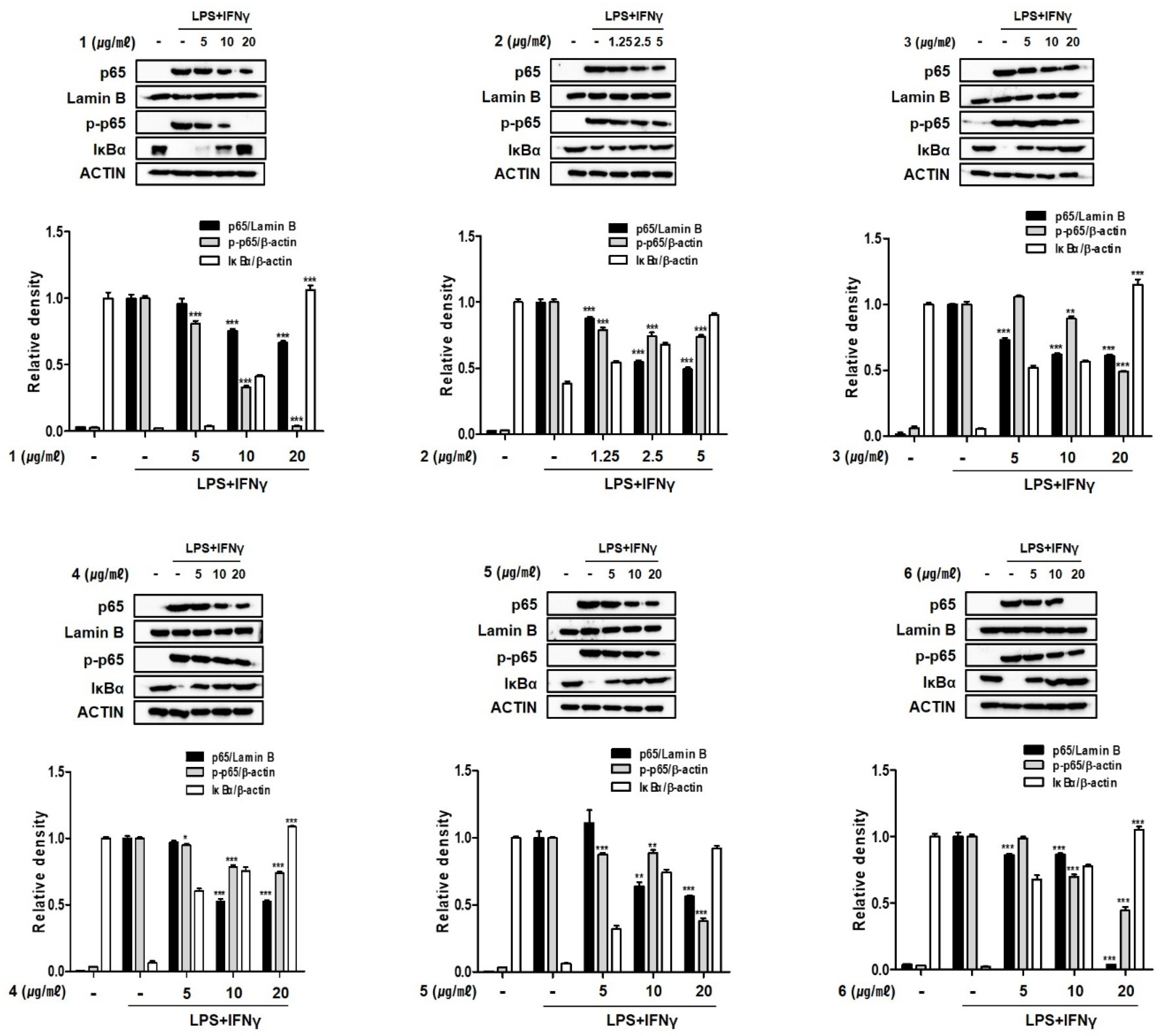
2.6. Compounds 1–6 Inhibit the Nuclear Translocation of Nuclear Factor Kappa B (NF-κB) in THP-1 Macrophages Co-Cultured with Caco-2 Cells
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Chemicals and Antibodies
3.3. Collection of Marine Sponge, E. acervus
3.4. Isolation of Fistularins from E. acervus
3.5. Mosher’s Reaction of FS-3
3.6. DP4+ Probability of Compound 1
3.7. Cell Cultures
3.8. Differentiation of THP-1 to Macrophages
3.9. In Vitro Co-Culture Model of Intestine
3.10. Measurment of TEER Values
3.11. Cytotoxicity Assay
3.12. Measurement of NO and Inflammatory Cytokines
3.13. Western Blotting
3.14. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2020, 37, 175–223. [Google Scholar] [CrossRef]
- Kaur, K.; Kumar, V.; Sharma, A.K.; Gupta, G.K. Isoxazoline containing natural products as anticancer agents: A review. Eur. J. Med. Chem. 2014, 77, 121. [Google Scholar] [CrossRef] [PubMed]
- Galeano, E.; Thomas, O.P.; Robledo, S.; Munoz, D.; Martinez, A. Antiparasitic Bromotyrosine Derivatives from the Marine Sponge Verongula rigida. Mar. Drugs 2011, 9, 1902–1913. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Li, J.; Hamann, M.T. The Marine Bromotyrosine Derivatives. In The Alkaloids: Chemistry and Biology; Cordell, G.A., Ed.; Elsevier: Amsterdam, The Netherlands, 2005; Volume 61, p. 59. [Google Scholar]
- Gopichand, Y.; Schmitz, F.J. Marine natural products: Fistularin-1, -2 and -3 from the sponge Aplysina fistularis forma fulva. Tetrahedron Lett. 1979, 41, 3921. [Google Scholar] [CrossRef]
- Rogers, E.W.; de Oliveira, M.F.; Berlinck, R.G.S.; König, G.M.; Molinski, T.F. Stereochemical Heterogeneity in Verongid Sponge Metabolites. Absolute Stereochemistry of (+)-Fistularin-3 and (+)-11-epi-Fistularin-3 by Microscale LCMS-Marfey’s Analysis. J. Nat. Prod. 2005, 68, 891. [Google Scholar] [CrossRef]
- Floren, C.; Kim, K.R.; Schnekenburger, M.; Kim, H.-J.; Moriou, C.; Debitus, C.; Dicato, M.; Al-Mourabit, A.; Han, B.W.; Diederich, M. Synergistic AML cell death induction by marine cytotoxin (+)-1(R), 6(S), 1′ (R), 6′(S), 11(R), 17(S)-fistularin-3 and Bcl-2 inhibitor venetoclax. Mar. Drugs 2018, 16, 518. [Google Scholar] [CrossRef] [Green Version]
- Ciminiello, P.; Fattorusso, E.; Forino, M.; Magno, S. Chemistry of Verongida sponges VIII. Bromocompounds from the Mediterranean sponges Aplysina aerophoba and Aplysina cavernicola. Tetrahedron 1997, 53, 6565. [Google Scholar] [CrossRef]
- Mancini, I.; Guella, G.; Laboute, P.; Debitus, C.; Pietra, F. Hemifistularin 3: A degraded peptide or biogenetic precursors? Isolation from a sponge of the order Verogida from the coral sea or generation from base treatment of 11-oxofistularin 3. J. Chem. Soc. Perkin Trans. 1 1993, 24, 3121–3125. [Google Scholar] [CrossRef]
- König, G.M.; Wright, A.D. Agelorins A and B, and 11-epi-Fistularin-3, three new antibacterial fistularin-3 derivatives from the tropical marine sponge Agelas oroides. Heterocycles 1993, 36, 1351. [Google Scholar]
- Compagnone, R.S.; Avila, R.; Suárez, A.I.; Abrams, O.V.; Rangel, H.R.; Arvelo, F.; Piña, I.C.; Merentes, E. 11-deoxyfistularin-3, a new cytotoxic metabolite fro, the Caribbean sponge Aplysina fistularis insularis. J. Nat. Prod. 1999, 62, 1443. [Google Scholar] [CrossRef]
- Nicacio, K.J.; Ióca, L.P.; Fróes, A.M.; Leomil, L.; Appolinario, L.R.; Thompson, C.C.; Thompson, F.L.; Ferreira, A.G.; Williams, D.E.; Anderson, R.J.; et al. Cultures of the marine bacterium Pseuvibrio denitrificans Ab134 produce bromotyrosine-derived alkaloids previously only isolated from marine sponges. J. Nat. Prod. 2017, 80, 235. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.M.; Kim, N.-H.; Lee, S.; Kim, Y.N.; Heo, J.-D.; Jeong, E.J.; Rho, J.-R. Deacetylphylloketal, a new phylloketal derivative from a marine sponge, genus Phyllospongia, with potent anti-inflammatory activity in vitro co-culture model of intestine. Mar. Drugs 2019, 17, 634. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.M.; Kim, N.-H.; Lee, S.; Kim, Y.N.; Heo, J.-D.; Rho, J.-R.; Jeong, E.J. (10Z)-Debromohymenialdisine from marine sponge Stylissa sp. regulates intestinal inflammatory responses in co-culture model of epithelial Caco-2 cells and THP-1 macrophages cells. Molecules 2019, 24, 3394. [Google Scholar] [CrossRef] [Green Version]
- Kim, N.-H.; Lee, S.M.; Kim, Y.N.; Jeon, Y.-J.; Heo, J.-D.; Jeong, E.J.; Rho, J.-R. Standardized fraction of Turbinaria ornata alleviates dextran sulfate sodium-induced chronic colitis in C57BL/6 mice via upregulating of FOXP3 + regulatory T cells. Biomolecules 2020, 10, 1463. [Google Scholar] [CrossRef]
- Martín, M.J.; Rodríguez-Acebes, R.; García-Ramos, Y.; Martínez, V.; Murcia, C.; Digón, I.; Marco, I.; Pelay-Gimeno, M.; Fernández, R.; Reyes, F.; et al. Stellatolides, a new cyclodepsipeptide family from the sponge Ecionemia acervus: Isolation, solid-phase total synthesis, and full structure assignment of stellatolide A. J. Am. Chem. Soc. 2014, 136, 6754–6762. [Google Scholar] [CrossRef]
- Das, P.; Hamme II, A.T. Divergent and concise syntheses of spiroisoxazolines: First total synthesis of 11-deoxyfistularin-3. Eur. J. Org. Chem. 2015, 23, 5159–5166. [Google Scholar] [CrossRef]
- Kernan, M.R.; Cambie, R.C.; Bergquist, P.R. Chemistry of sponges, VII. 11,19-dideoxyfistularin 3 and 11-hydroxyaerlthionin, bromotyrosine derivatives from Pseudoceratina durissima. J. Nat. Prod. 1990, 53, 615. [Google Scholar] [CrossRef]
- Grimblat, N.; Zanardi, M.; Sarotti, A.M. Beyond DP4: An improved probability for the stereochemical assignment of isomeric compounds using quantum chemical calculations of NMR shifts. J. Org. Chem. 2015, 80, 12526–12534. [Google Scholar] [CrossRef] [PubMed]
- Müller, E.; Christopoulos, P.F.; Halder, S.; Lunde, A.; Beraki, K.; Speth, M.; Øynebråten, I.; Corthay, A. Toll-Like Receptor Ligands and Interferon-gamma Synergize for Induction of Antitumor M1 Macrophages. Front Immunol. 2017, 8, 1383. [Google Scholar] [CrossRef]
- Sharon, P.; Stenson, W.F. Enhanced synthesis of leukotriene B4 by colonic mucosa in inflammatory bowel disease. Gastroenterology 1984, 86, 453. [Google Scholar] [CrossRef]
- Lauritsen, K.; Laursen, L.S.; Bukhave, K.; Rask-Madsen, J. Effects of topical 5-aminosalicylic acid and prednisolone on prostaglandin E2, and leukotriene B4 levels determined by equilibrium in vivo dialysis of rectum in relapsing ulcerative colitis. Gastroenterology 1986, 91, 837. [Google Scholar] [CrossRef]
- Singer, I.I.; Kawka, D.W.; Schloemann, S.; Tessner, T.; Riehl, T.; Stenson, W.F. Cyclooxygenase 2 is induced in colonic epithelial cells in inflammatory bowel disease. Gastroenterology 1998, 115, 297. [Google Scholar] [CrossRef]
- Economou, J.S.; Rhoades, R.; Essner, W.H.; McBride, J.C.; Morton, D.L. Genetic analysis of the human tumor necrosis factor a/cachectin promoter region in a macrophage cell line. J. Exp. Med. 1989, 170, 321. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Takada, Y.; Shishodia, S.; Gutierrez, A.M.; Oommen, O.V.; Ichikawa, H. Nuclear transcription factor NF-kappa B: Role in biology and medicine. Indian J. Exp. Biol. 2004, 42, 341–353. [Google Scholar]
- Mitsuyama, K.; Sata, M.; Tanikawa, K. Significance of interleukin-6 in patients with inflammatory bowel disease. Gastroenterol. Jpn. 1991, 26, 20. [Google Scholar] [CrossRef] [PubMed]
- Rosen, M.J.; Dhawan, A.; Saeed, S.A. Inflammatory Bowel Disease in Children and Adolescents. Jama Pediatr. 2015, 169, 1053. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waetzig, G.H.; Seegert, D.; Rosenstiel, P.; Nikolaus, S.; Schreiber, S. p38 Mitogen-activated protein kinase is activated and linked to TNFalpha signaling in inflammatory bowel disease. J. Immunol. 2002, 168, 5342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mitsuyama, K.; Suzuki, A.; Tomiyasu, N.; Tsuruta, O.; Kitazaki, S.; Takeda, T.; Satoh, Y.; Bennett, B.L.; Toyonaga, A.; Sata, M. Pro-inflammatory signaling by Jun-N-terminal kinase in inflammatory bowel disease. Int. J. Mol. Med. 2006, 17, 449. [Google Scholar] [CrossRef] [Green Version]
- Miampamba, M. 743 Rdea119, a potent and highly selective MEK inhibitor ameliorates murine colitis. Gastroenterology 2008, 134 (Suppl. 1), A-107. [Google Scholar] [CrossRef]
- Sandle, G.I.; Hayslett, J.P.; Binder, H.J. Effect of glucocorticoids on rectal transport in normal subjects and patients with ulcerative colitis. Gut 1986, 27, 309. [Google Scholar] [CrossRef] [Green Version]
- Reinecker, H.C.; Steffen, M.; Withoeft, T.; Pflueger, I.; Schreiber, S.; MacDermott, R.P.; Raedler, A. Enhanced secretion of tumour necrosis factor-alpha, IL-6, and IL-1 beta by isolated lamina propria mononuclear cells from patients with ulcerative colitis and Crohn’s disease. Clin. Exp. Immunol. 1993, 94, 174. [Google Scholar] [CrossRef]
- Lappas, M.; Permezel, M.; Georgiou, H.M.; Rice, G.E. Nuclear factor kappa B regulation of proinflammatory cytokines in human gestational tissues in vitro. Biol. Reprod. 2002, 67, 668. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Appleby, S.B.; Ristimäki, A.; Neilson, K.; Narko, K.; Hla, T. Structure of the human cyclo-oxygenase-2 gene. Biochem. J. 1994, 302, 723. [Google Scholar] [CrossRef]
- Park, H.J.; Kim, I.T.; Won, J.H. Anti-inflammatory activities of ent16alphaH,17-hydroxy-kauran-19-oic acid isolated from the roots of Siegesbeckia pubescens are due to the inhibition of iNOS and COX-2 expression in RAW 264.7 macrophages via NF-kappaB inactivation. Eur. J. Pharmacol. 2007, 558, 185. [Google Scholar] [CrossRef]
- Oeckinghaus, A.; Ghosh, S. The NF-kappaB family of transcription factors and its regulation. Cold Spring Harb. Perspect. Biol. 2009, 1, a000034. [Google Scholar] [CrossRef]
- Kleiveland, C.R. Chapter 18: Co-culture Caco-2/immune cells. Impact Food Bioact. Health 2015. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.M.; Kim, N.H.; Ji, Y.K.; Kim, Y.N.; Jeon, Y.-J.; Heo, J.D.; Jeong, E.J.; Rho, J.-R. Sulfoquinovosylmonoacylglycerols regulating intestinal inflammation in co-culture system from the brown alga Turbinaria ornata. Algae 2020, 35, 201. [Google Scholar] [CrossRef]
- Puyana, M.; Pawlik, J.; Blum, J.; Fenical, W. Metabolite variability in Caribbean sponges of the genus Aplysina. Rev. Bras. Farmacon. 2015, 25, 592–599. [Google Scholar] [CrossRef] [Green Version]
- Rogers, E.W.; Molinski, T.F. Highly Polar spoiroisoxazolines from the sponge Aplysina fulva. J. Nat. Prod. 2007, 70, 1191–1194. [Google Scholar] [CrossRef] [Green Version]
- Thompson, C.C.; Kruger, R.H.; Thompson, F.L. Unlocking marine biotechnology in developing world. Trens Biotechnol. 2017, 35, 1119–1121. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | FS-3 (11S, 17R) | epi-FS-3 (11R, 17S) * | ||
|---|---|---|---|---|
| 13C | 1H | 13C | 1H | |
| 1, 1′ | 75.1, 75.1 | 4.17, s/4.19, s | 75.2, 75.3 | 4.18, s/4.19, s |
| 2, 2′ | 122.1, 122.2 | 122.1, 122.1 | ||
| 3, 3′ | 148.7, 148.7 | 148.8, 148.8 | ||
| 4, 4′ | 113.8, 113.8 | 113.8, 113.9 | ||
| 5, 5′ | 132.2, 132.3 | 6.53, s/6.55, s | 132.3, 132.4 | 6.52, s/6.53, s |
| 6, 6′ | 91.3, 91.3 | 91.8, 91.8 | ||
| 7, 7′ | 39.5, 39.5 | 3.21/3.86, d (18.1) | 40.0, 40.0 | 3.19 /3.85, d (18.0) |
| 3.18/3.83, d (18.1) | 3.16 /3.82, d (18.0) | |||
| 8, 8′ | 155.1, 155.1 | 155.1, 155.2 | ||
| 9, 9′ | 160.3, 160.4 | 160.5, 160.5 | ||
| 10 | 43.4 | 3.52, dd (13.9, 7.3) | 43.6 | 3.54, m |
| 3.78, dd (13.7, 4.7) | 3.80, m | |||
| 11 | 69.7 | 4.25, m | 69.9 | 4.25, m |
| 12 | 75.8 | 4.02, dd (9.1, 5.9) | 75.9 | 4.02, dd (9.1, 5.5) |
| 4.07, dd (9.1, 5.6) | 4.06, dd (9.1, 5.5) | |||
| 13 | 152.6 | 152.7 | ||
| 14, 14′ | 118.3, 118.3 | 118.4, 118.4 | ||
| 15, 15′ | 131.4, 131.4 | 7.67, s | 131.5, 131.5 | 7.66, s |
| 16 | 143.3 | 143.3 | ||
| 17 | 71.3 | 4.90, dd (7.3, 4.7) | 71.3 | 4.90, dd (7.7, 4.3) |
| 18 | 47.4 | 3.48, dd (13.7, 7.3) | 47.7 | 3.49, m |
| 3.60, dd (13.7, 4.7) | 3.63, m | |||
| OCH3 | 60.2 | 3.73, s | 60.2 | 3.73, s |
| NH | 7.69, br t (5.4) | 7.62, br t (6.0) | ||
| NH’ | 7.75, br t (6.1) | 7.66, br t (6.0) | ||
| Compounds (20 μM) | NO | PGE2 |
|---|---|---|
| Relative % | ||
| Non-treated control | 0.00 | |
| LPS + IFNγ only | 100.00 | |
| 1 | 32.97 ± 0.85 | −7.50 ± 3.03 |
| 2 * | 24.46 ± 7.42 | −10.40 ± 0.39 |
| 3 | 19.39 ± 0.66 | 15.54 ± 1.95 |
| 4 | 39.82 ± 4.94 | 3.70 ± 1.61 |
| 5 | 33.24 ± 3.09 | 67.98 ± 1.60 |
| 6 | 33.22 ± 2.65 | 2.63 ± 0.51 |
| Compounds (20 μM) | IL-1β | IL-6 | TNF-α |
|---|---|---|---|
| Relative % | |||
| Non-treated control | 0.00 | ||
| LPS+ IFNγ only | 100.00 | ||
| 1 | 58.62 ± 3.00 | 67.32 ± 2.13 | 58.97 ± 4.75 |
| 2 * | 67.66 ± 11.00 | 64.89 ± 5.23 | 46.84 ± 6.21 |
| 3 | 48.17 ± 3.00 | 42.94 ± 6.77 | 54.21 ± 3.46 |
| 4 | 25.05 ± 3.41 | 46.96 ± 1.14 | 62.03 ± 1.06 |
| 5 | 34.37 ± 2.96 | 48.25 ± 0.62 | 48.19 ± 3.47 |
| 6 | 60.07 ± 2.95 | 64.82 ± 1.60 | 50.83 ± 2.47 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ji, Y.K.; Lee, S.M.; Kim, N.-H.; Tu, N.V.; Kim, Y.N.; Heo, J.D.; Jeong, E.J.; Rho, J.-R. Stereochemical Determination of Fistularins Isolated from the Marine Sponge Ecionemia acervus and Their Regulatory Effect on Intestinal Inflammation. Mar. Drugs 2021, 19, 170. https://doi.org/10.3390/md19030170
Ji YK, Lee SM, Kim N-H, Tu NV, Kim YN, Heo JD, Jeong EJ, Rho J-R. Stereochemical Determination of Fistularins Isolated from the Marine Sponge Ecionemia acervus and Their Regulatory Effect on Intestinal Inflammation. Marine Drugs. 2021; 19(3):170. https://doi.org/10.3390/md19030170
Chicago/Turabian StyleJi, Yeong Kwang, Seon Min Lee, Na-Hyun Kim, Nguyen Van Tu, Yun Na Kim, Jeong Doo Heo, Eun Ju Jeong, and Jung-Rae Rho. 2021. "Stereochemical Determination of Fistularins Isolated from the Marine Sponge Ecionemia acervus and Their Regulatory Effect on Intestinal Inflammation" Marine Drugs 19, no. 3: 170. https://doi.org/10.3390/md19030170
APA StyleJi, Y. K., Lee, S. M., Kim, N. -H., Tu, N. V., Kim, Y. N., Heo, J. D., Jeong, E. J., & Rho, J. -R. (2021). Stereochemical Determination of Fistularins Isolated from the Marine Sponge Ecionemia acervus and Their Regulatory Effect on Intestinal Inflammation. Marine Drugs, 19(3), 170. https://doi.org/10.3390/md19030170