
Enhancement of Xanthophyll Synthesis in Porphyra/Pyropia Species (Rhodophyta, Bangiales) by Controlled Abiotic Factors: A Systematic Review and Meta-Analysis



Abstract
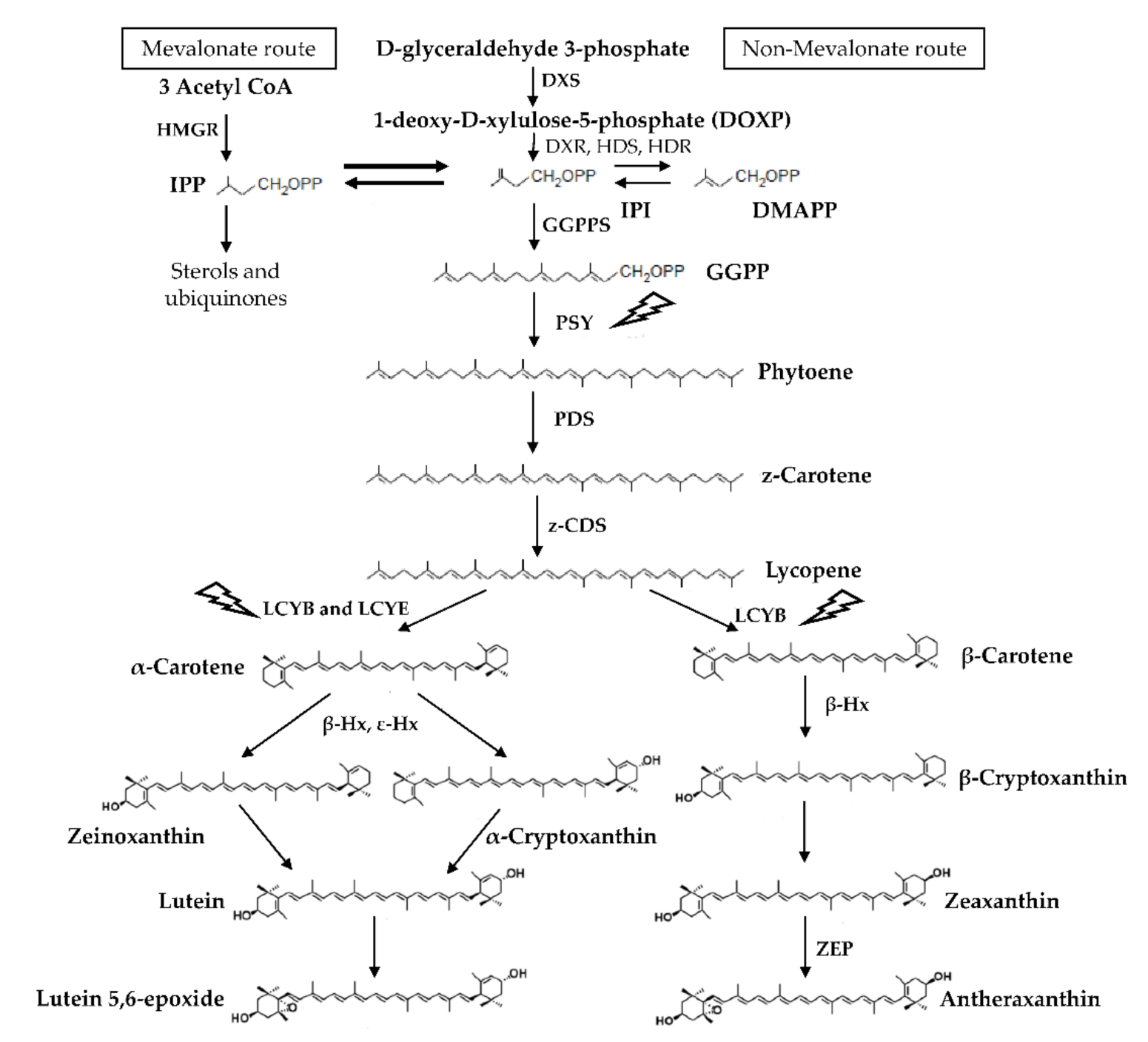
:1. Introduction
2. Results
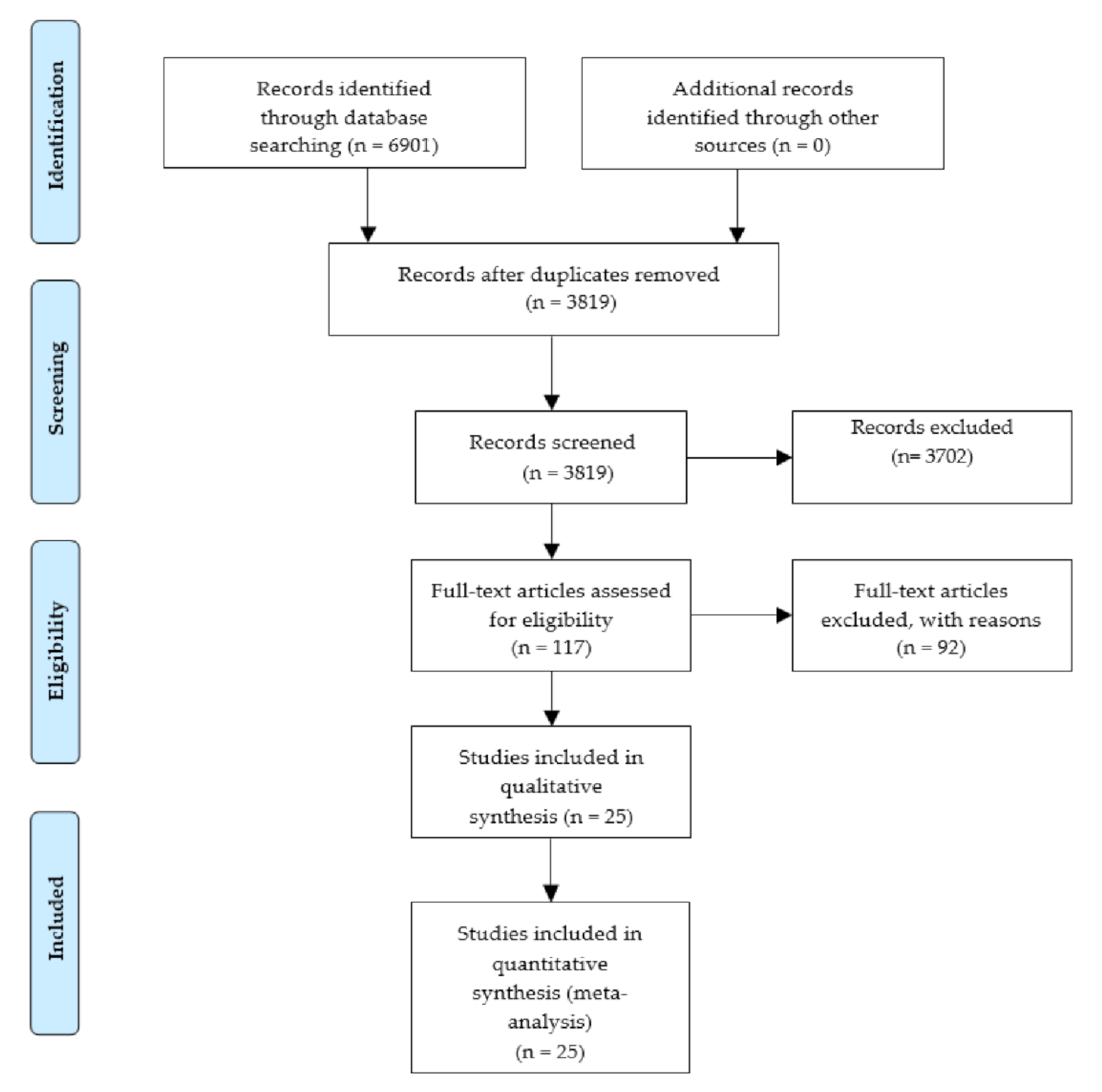
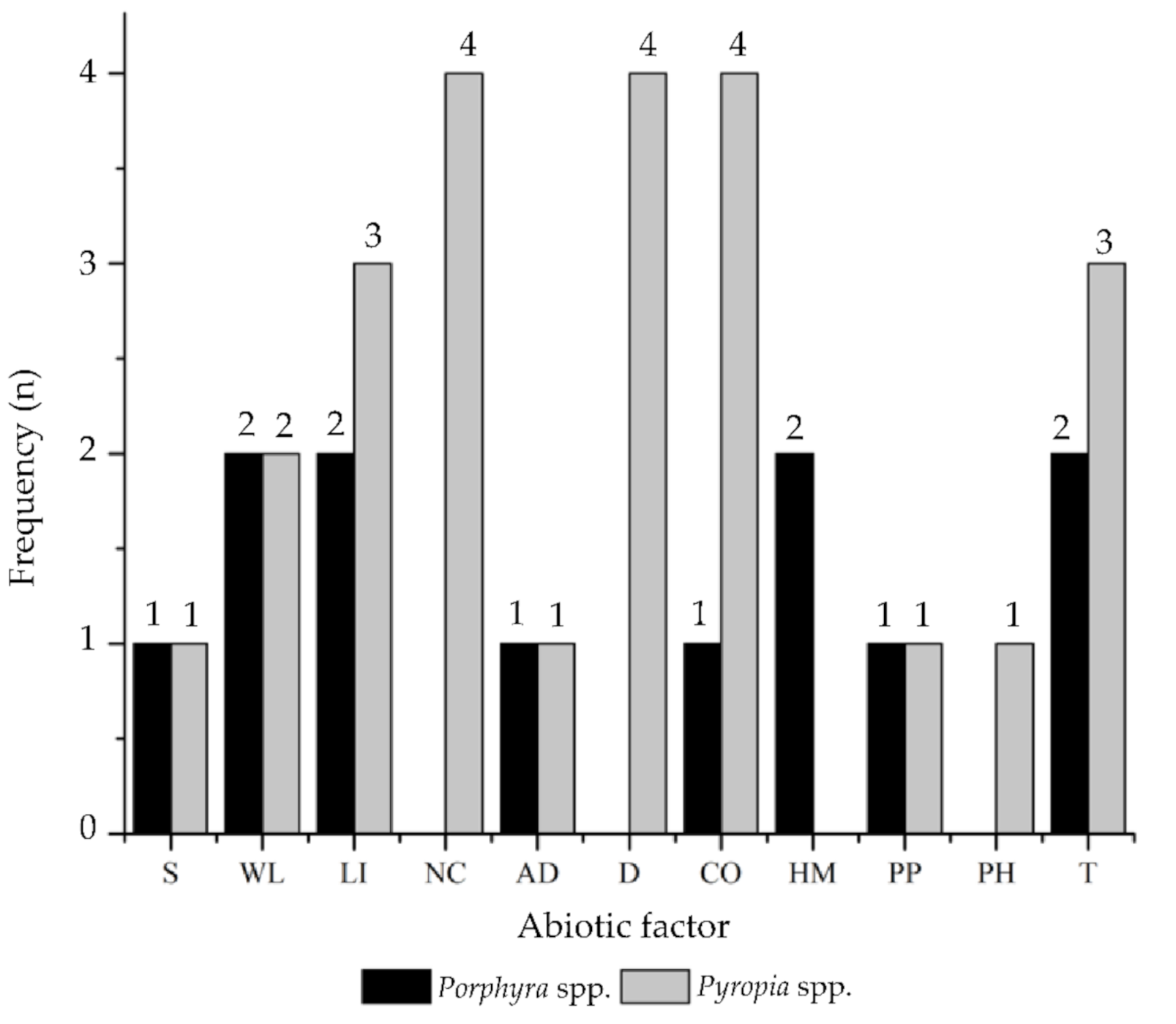
2.1. Systematic Review and Data Extraction
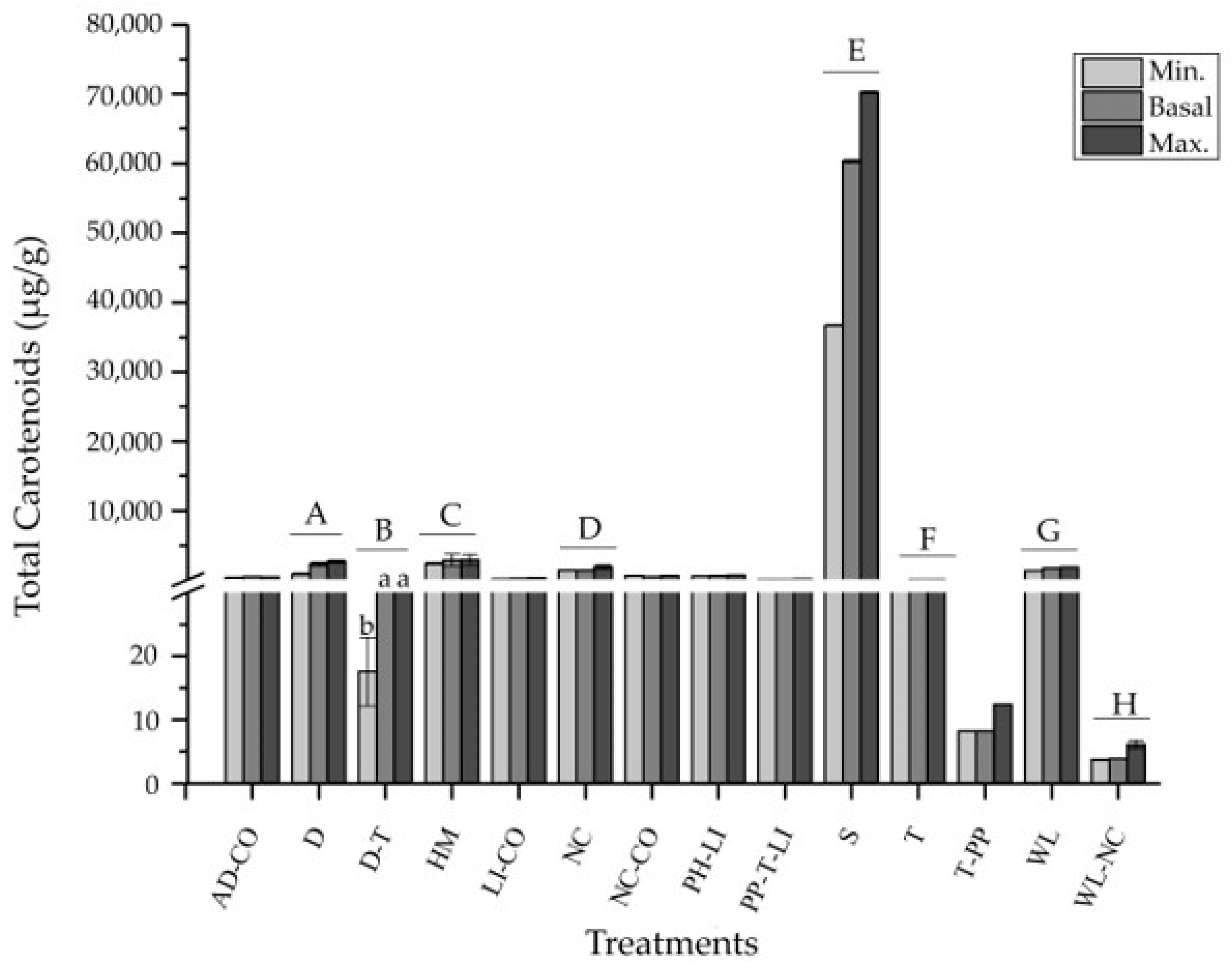
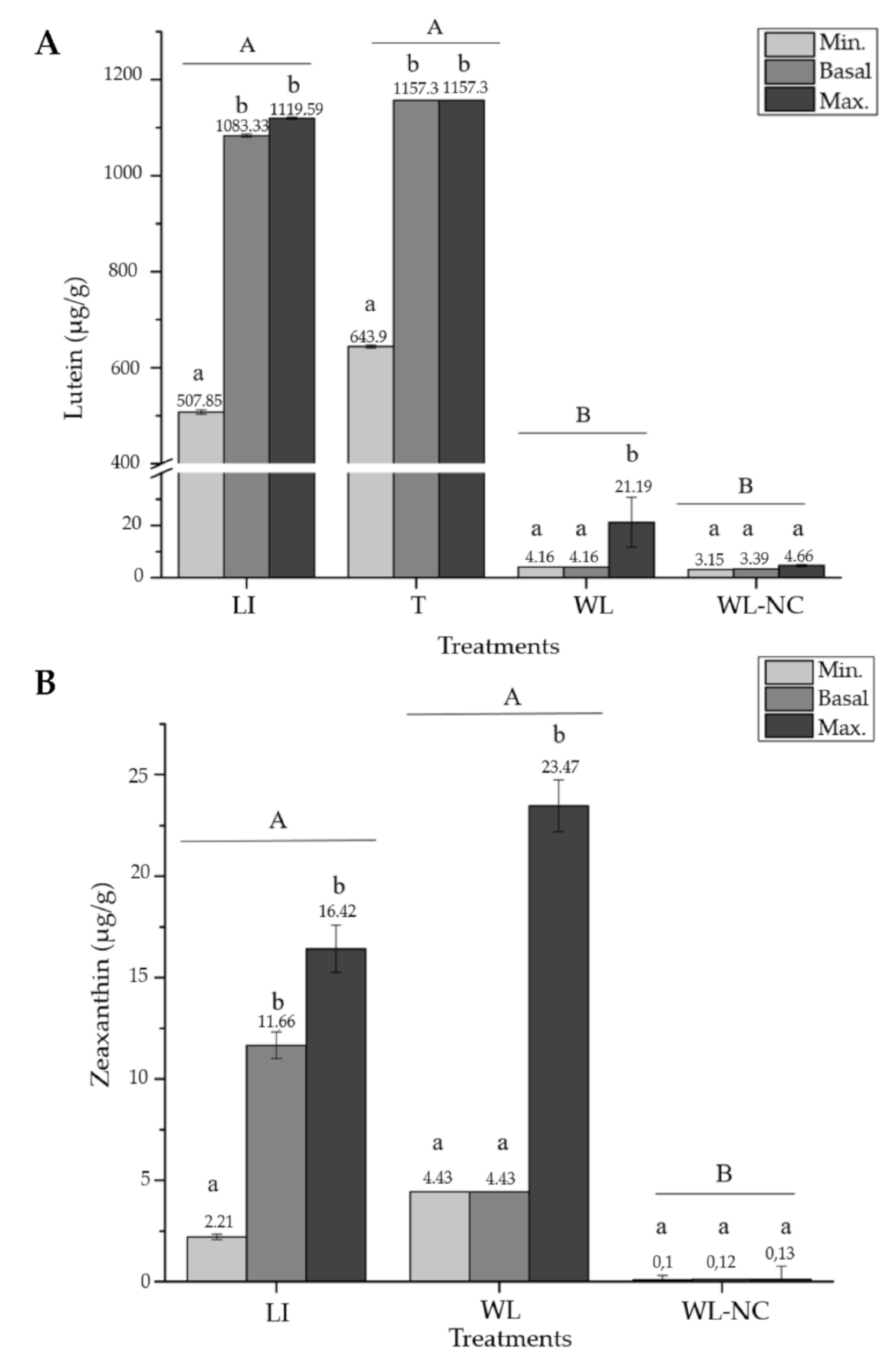
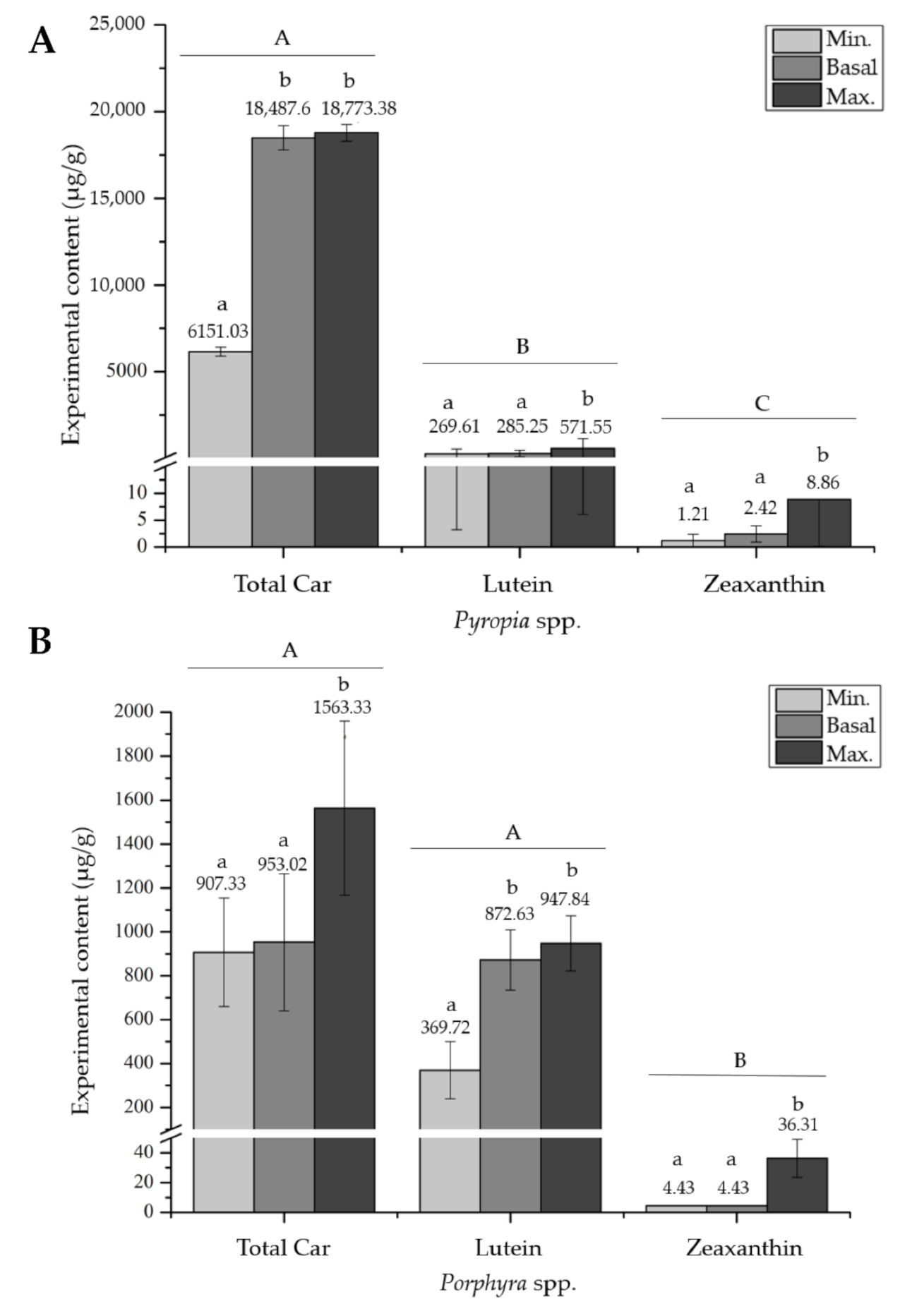
2.2. Meta-Analysis
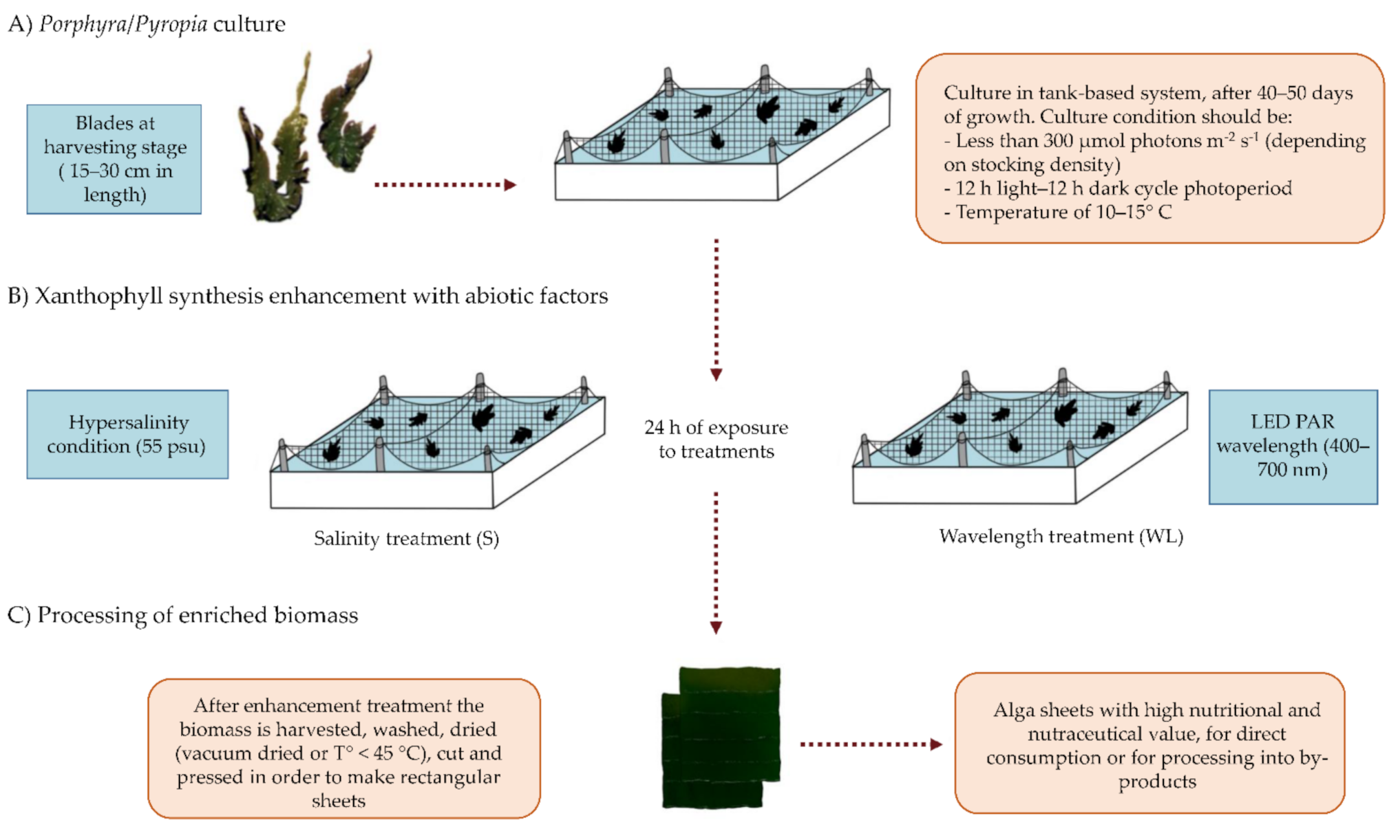
2.3. Suggested Culture Method
3. Discussion
4. Materials and Methods
4.1. Systematic Review
4.2. Data Extraction
4.3. Meta-Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rodriguez-Concepcion, M.; Avalos, J.; Bonet, M.L.; Boronat, A.; Gomez-Gomez, L.; Hornero-Mendez, D.; Limon, M.C.; Meléndez-Martínez, A.J.; Olmedilla-Alonso, B.; Palou, A.; et al. A global perspective on carotenoids: Metabolism, biotechnology, and benefits for nutrition and health. Prog. Lipid Res. 2018, 70, 62–93. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaczor, A.; Baranska, M. Carotenoids: Nutrition, Analysis and Technology, 1st ed.; Wiley Blackwell: Chichester, UK; Hoboken, NJ, USA, 2016; ISBN 978-1-118-62226-1. [Google Scholar]
- Widomska, J.; Subczynski, W.K. Why has Nature Chosen Lutein and Zeaxanthin to Protect the Retina? J. Clin. Exp. Ophthalmol. 2014, 5, 326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krinsky, N.I.; Johnson, E.J. Carotenoid actions and their relation to health and disease. Mol. Asp. Med. 2005, 26, 459–516. [Google Scholar] [CrossRef] [PubMed]
- Juturu, V. Lutein, Brain, and Neurological Functions. In Bioactive Nutraceuticals and Dietary Supplements in Neurological and Brain Disease; Elsevier: Amsterdam, The Netherlands, 2015; pp. 41–47. ISBN 978-0-12-411462-3. [Google Scholar]
- Abdel-Aal, E.S.; Akhtar, H.; Zaheer, K.; Ali, R. Dietary Sources of Lutein and Zeaxanthin Carotenoids and Their Role in Eye Health. Nutrients 2013, 5, 1169–1185. [Google Scholar] [CrossRef] [Green Version]
- Vachali, P.P.; Besch, B.M.; Bernstein, P.S. Carotenoids and Age-Related Macular Degeneration. In Handbook of Nutrition, Diet and the Eye; Elsevier: Amsterdam, The Netherlands, 2014; pp. 77–84. ISBN 978-0-12-401717-7. [Google Scholar]
- Buscemi, S.; Corleo, D.; Di Pace, F.; Petroni, M.L.; Satriano, A.; Marchesini, G. The Effect of Lutein on Eye and Extra-Eye Health. Nutrients 2018, 10, 1321. [Google Scholar] [CrossRef] [Green Version]
- Terasaki, M.; Mutoh, M.; Fujii, G.; Takahashi, M.; Ishigamori, R.; Masuda, S. Potential ability of xanthophylls to prevent obesity associated cancer. World J. Pharmacol. 2014, 3, 140–152. [Google Scholar] [CrossRef]
- Sumantran, V.N.; Zhang, R.; Lee, D.S.; Wicha, M.S. Differential Regulation of Apoptosis in Normal versus Transformed Mammary Epithelium by Lutein and Retinoic Acid. Cancer Epidemiol. Biomark. Prev. 2000, 9, 257–263. Available online: https://cebp.aacrjournals.org/content/9/3/257.long (accessed on 3 November 2020).
- Rengasamy, K.R.R.; Mahomoodally, M.F.; Aumeeruddy, M.Z.; Zengin, G.; Xiao, J.; Kim, D.H. Bioactive compounds in seaweeds: An overview of their biological properties and safety. Food Chem. Toxicol. 2020, 135, 111013. [Google Scholar] [CrossRef]
- FAO. Fishery and Aquaculture Statistics. Global Aquaculture Production 1950–2019. Available online: http://www.fao.org/fishery/statistics/global-aquaculture-production/query/en (accessed on 10 April 2021).
- Cao, J.; Wang, J.; Wang, S.; Xu, X. Porphyra Species: A Mini-Review of Its Pharmacological and Nutritional Properties. J. Med. Food 2016, 19, 111–119. [Google Scholar] [CrossRef]
- Cho, T.J.; Rhee, M.S. Health Functionality and Quality Control of Laver (Porphyra, Pyropia): Current Issues and Future Perspectives as an Edible Seaweed. Mar. Drugs 2019, 18, 14. [Google Scholar] [CrossRef] [Green Version]
- Cazzonelli, C.I. Carotenoids in nature: Insights from plants and beyond. Funct. Plant. Biol. 2011, 38, 833. [Google Scholar] [CrossRef] [Green Version]
- Formaggio, E.; Cinque, G.; Bassi, R. Functional Architecture of the Major Light-harvesting Complex from Higher Plants. J. Mol. Biol. 2001, 314, 1157–1166. [Google Scholar] [CrossRef]
- Bailey, S.; Grossman, A. Photoprotection in Cyanobacteria: Regulation of Light Harvesting. Photochem. Photobiol. 2008, 84, 1410–1420. [Google Scholar] [CrossRef]
- Alboresi, A.; Dall’Osto, L.; Aprile, A.; Carillo, P.; Roncaglia, E.; Cattivelli, L.; Bassi, R. Reactive oxygen species and transcript analysis upon excess light treatment in wild-type Arabidopsis thaliana vs a photosensitive mutant lacking zeaxanthin and lutein. BMC Plant. Biol. 2011, 11, 62. [Google Scholar] [CrossRef] [Green Version]
- Cunningham, F.X.; Gantt, E., Jr. Genes and enzymes of carotenoid biosynthesis in plants. Annu. Rev. Plant. Physiol. Plant. Mol. Biol. 1998, 49, 557–583. [Google Scholar] [CrossRef]
- Pizarro, L.; Stange, C. Light-dependent regulation of carotenoid biosynthesis in plants. Cienc. Investig. Agrar. 2009, 36, 143–162. [Google Scholar] [CrossRef]
- Koizumi, J.; Takatani, N.; Kobayashi, N.; Mikami, K.; Miyashita, K.; Yamano, Y.; Wada, A.; Maoka, T.; Hosokawa, M. Carotenoid Profiling of a Red Seaweed Pyropia yezoensis: Insights into Biosynthetic Pathways in the Order Bangiales. Mar. Drugs 2018, 16, 426. [Google Scholar] [CrossRef] [Green Version]
- Schubert, N.; García-Mendoza, E.; Pacheco-Ruiz, I. Carotenoid composition of marine red algae. J. Phycol. 2006, 42, 1208–1216. [Google Scholar] [CrossRef]
- Schubert, N.; García-Mendoza, E. Photoinhibition in red algal species with different carotenoid profiles. J. Phycol. 2008, 44, 1437–1446. [Google Scholar] [CrossRef]
- Flores-Molina, M.R.; Thomas, D.; Lovazzano, C.; Núñez, A.; Zapata, J.; Kumar, M.; Correa, J.A.; Contreras-Porcia, L. Desiccation stress in intertidal seaweeds: Effects on morphology, antioxidant responses and photosynthetic performance. Aquat. Bot. 2014, 113, 90–99. [Google Scholar] [CrossRef]
- Takaichi, S.; Yokoyama, A.; Mochimaru, M.; Uchida, H.; Murakami, A. Carotenogenesis diversification in phylogenetic lineages of Rhodophyta. J. Phycol. 2016, 52, 329–338. [Google Scholar] [CrossRef]
- Römer, S.; Fraser, P.D. Recent advances in carotenoid biosynthesis, regulation and manipulation. Planta 2005, 221, 305–308. [Google Scholar] [CrossRef]
- Howitt, C.A.; Pogson, B.J. Carotenoid accumulation and function in seeds and non-green tissues. Plant. Cell Environ. 2006, 29, 435–445. [Google Scholar] [CrossRef]
- Stange, C.; Flores, C. Carotenoids and Photosynthesis—Regulation of Carotenoid Biosynthesis by Photoreceptors. In Advances in Photosynthesis—Fundamental Aspects; Najafpour, M., Ed.; InTech: Rijeka, Croatia, 2012; ISBN 978-953-307-928-8. [Google Scholar]
- Llorente, B. Regulation of Carotenoid Biosynthesis in Photosynthetic Organs. In Carotenoids in Nature; Stange, C., Ed.; Subcellular Biochemistry; Springer International Publishing: Cham, Switzerland, 2016; Volume 79, pp. 141–160. ISBN 978-3-319-39124-3. [Google Scholar]
- Oslan, S.N.H.; Shoparwe, N.F.; Yusoff, A.H.; Rahim, A.A.; Chang, C.S.; Tan, J.S.; Oslan, S.N.; Arumugam, K.; Ariff, A.B.; Sulaiman, A.Z.; et al. A Review on Haematococcus pluvialis Bioprocess Optimization of Green and Red Stage Culture Conditions for the Production of Natural Astaxanthin. Biomolecules 2021, 11, 256. [Google Scholar] [CrossRef]
- Bohne, F.; Linden, H. Regulation of carotenoid biosynthesis genes in response to light in Chlamydomonas reinhardtii. Biochim. Biophys. Acta BBA—Gene Struct. Expr. 2002, 1579, 26–34. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, Z.-W.; Zhong, C.-H.; Ding, L.; Zheng, Y.-Y.; Zhang, G. Effects of Concentrated Seawater on Photosynthetic Pigment Contents in Porphyra haitanensis. Pol. J. Environ. Stud. 2020, 29, 2475–2480. [Google Scholar] [CrossRef]
- Zhu, X.; Zou, D.; Huang, Y.; Cao, J.; Sun, Y.; Chen, B.; Chen, X. Physiological responses of Porphyra haitanensis (Rhodophyta) to copper and cadmium exposure. Bot. Mar. 2017, 60, 27–37. [Google Scholar] [CrossRef]
- Li, G.; Mai, G.; Zhang, J.; Lin, Q.; Ni, G.; Tan, Y.; Huang, L.; Zou, D. Algal density alleviates the elevated CO2-caused reduction on growth of Porphyra haitanensis (Bangiales, Rhodophyta), a species farmed in China. Aquac. Res. 2020, 51, 3879–3887. [Google Scholar] [CrossRef]
- Li, Y.X.; Zhou, S.; Zhao, F.J.; Liu, Y.; Fan, P.P.; Wang, G.C. Physiological responses of Porphyra haitanesis to different copper and zinc concentrations. Braz. J. Oceanogr. 2010, 58, 261–267. [Google Scholar] [CrossRef]
- Oh, S.; Lee, E.; Choe, E. Light Effects on Lipid Oxidation, Antioxidants, and Pigments in Dried Laver (Porphyra) during Storage. Food Sci. Biotechnol. 2014, 23, 701–709. [Google Scholar] [CrossRef]
- Son, S.; Choe, E. Toasting Effects on the Lipid Oxidation, Antioxidants, and Pigments of Dried Laver (Porphyra spp.). Korean J. Food Sci. Technol. 2014, 46, 677–681. [Google Scholar] [CrossRef] [Green Version]
- Bouzon, Z.L.; Chow, F.; Zitta, C.S.; dos Santos, R.W.; Ouriques, L.C.; de L. Felix, M.R.; Osorio, L.K.P.; Gouveia, C.; de Paula Martins, R.; Latini, A.; et al. Effects of Natural Radiation, Photosynthetically Active Radiation and Artificial Ultraviolet Radiation-B on the Chloroplast Organization and Metabolism of Porphyra acanthophora var. brasiliensis (Rhodophyta, Bangiales). Microsc. Microanal. 2012, 18, 1467–1479. [Google Scholar] [CrossRef] [PubMed]
- Green, L.A.; Neefus, C.D. Effects of temperature, light level, and photoperiod on the physiology of Porphyra umbilicalis Kützing from the Northwest Atlantic, a candidate for aquaculture. J. Appl. Phycol. 2015, 28, 1815–1826. [Google Scholar] [CrossRef]
- Aguilera, J.; Jiménez, C.; Figueroa, F.L.; Lebert, M.; Häder, D.-P. Effect of ultraviolet radiation on thallus absorption and photosynthetic pigments in the red alga Porphyra umbilicalis. J. Photochem. Photobiol. B: Biol. 1999, 48, 75–82. [Google Scholar] [CrossRef]
- Uribe, E.; Vega-Gálvez, A.; Heredia, V.; Pastén, A.; Di Scala, K. An Edible Red seaweed (Pyropia orbicularis): Influence of vacuum drying on physicochemical composition, bioactive compounds, antioxidant capacity, and pigments. J. Appl. Phycol. 2018, 30, 673–683. [Google Scholar] [CrossRef]
- Uribe, E.; Vega-Gálvez, A.; García, V.; Pastén, A.; Rodríguez, K.; López, J.; Di Scala, K. Evaluation of physicochemical composition and bioactivity of a red seaweed (Pyropia orbicularis) as affected by different drying technologies. Dry. Technol. 2020, 38, 1218–1230. [Google Scholar] [CrossRef]
- Pereira, D.T.; Filipin, E.P.; Ramlov, F.; Maraschin, M.; Bouzon, Z.L.; Simioni, C. Pyropia acanthophora var. brasiliensis EC Oliveira and Coll (Rhodophyta: Bangiales) cultivated in seawater under laboratory conditions favors the production of economically important secondary metabolites. Braz. J. Biol. Sci. 2018, 5, 85–93. [Google Scholar] [CrossRef]
- Pereira, D.T.; Schmidt, É.C.; Filipin, E.P.; Pilatti, F.K.; Ramlov, F.; Maraschin, M.; Bouzon, Z.L.; Simioni, C. Effects of ultraviolet radiation on the morphophysiology of the macroalga Pyropia acanthophora var. brasiliensis (Rhodophyta, Bangiales) cultivated at high concentrations of nitrate. Acta Physiol. Plant. 2020, 42, 61. [Google Scholar] [CrossRef]
- Xie, X.; Lu, X.; Wang, L.; He, L.; Wang, G. High light intensity increases the concentrations of β-carotene and zeaxanthin in marine red macroalgae. Algal Res. 2020, 47, 101852. [Google Scholar] [CrossRef]
- Xin-shu, L.; Zi-hui, F.; Pei-min, H. Effects of solar radiation on photosynthesis and pigmentation in the red alga Pyropia yezoensis Ueda (Bangiales, Rhodophyta). Indian J. Geo-Mar. Sci. 2014, 43, 473–4780. [Google Scholar]
- Wang, W.J.; Li, X.L.; Sun, T.Q.; Liang, Z.R.; Liu, F.L.; Sun, X.T.; Wang, F.J. Effects of periodical drying and non-drying on nutrient content and desiccation tolerance of an intertidal Pyropia yezoensis strain subject to farming conditions. J. Appl. Phycol. 2018, 31, 1897–1906. [Google Scholar] [CrossRef]
- Samanta, P.; Shin, S.; Jang, S.; Kim, J.K. Comparative assessment of salinity tolerance based on physiological and biochemical performances in Ulva australis and Pyropia yezoensis. Algal Res. 2019, 42, 101590. [Google Scholar] [CrossRef]
- Kim, S.; Yoon, S.C.; Yoo, M.H.; Park, K.W.; Park, S.R.; Youn, S.-H. Physiological Responses of Cultured Seaweed Pyropia yezoensis to Phosphorous Limitation in the Nakdong River Estuary, Korea. Ocean. Sci. J. 2018, 54, 129–139. [Google Scholar] [CrossRef]
- Li, X.; Wang, W.-J.; Liu, F.-L.; Liang, Z.-R.; Sun, X.-T.; Yao, H.-Q.; Wang, F.-J. Periodical drying or no drying during aquaculture affects the desiccation tolerance of a sublittoral Pyropia yezoensis Strain. J. Appl. Phycol. 2017, 30, 697–705. [Google Scholar] [CrossRef]
- Bao, M.; Wang, J.; Xu, T.; Wu, H.; Li, X.; Xu, J. Rising CO2 levels alter the responses of the red macroalga Pyropia yezoensis under light stress. Aquaculture 2018, 501, 325–330. [Google Scholar] [CrossRef]
- Urrea-Victoria, V. Efeito do estresse térmico sobre respostas fisiológicas, composição química e potencial antioxidante de Sargassum stenophyllum (Fucales, Ochrophyta) e Pyropia spiralis (Bangiales, Rhodophyta). Ph.D. Thesis, Universidade de São Paulo, São Paulo, Brasil, 2019. [Google Scholar]
- Wu, H. Effect of different light qualities on growth, pigment content, chlorophyll fluorescence, and antioxidant enzyme activity in the red alga Pyropia haitanensis (Bangiales, Rhodophyta). BioMed Res. Int. 2016, 2016, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Jiang, H.; Zou, D.; Lou, W.; Deng, Y.; Zeng, X. Effects of seawater acidification and alkalization on the farmed seaweed, Pyropia haitanensis (Bangiales, Rhodophyta), grown under different irradiance conditions. Algal Res. 2018, 31, 413–420. [Google Scholar] [CrossRef]
- Chen, B.; Zou, D.; Yang, Y. Increased iron availability resulting from increased CO2 enhances carbon and nitrogen metabolism in the economical marine red macroalga Pyropia haitanensis (Rhodophyta). Chemosphere 2017, 173, 444–451. [Google Scholar] [CrossRef]
- Chen, B.; Zou, D.; Ma, J. Interactive effects of elevated CO2 and nitrogen–phosphorus supply on the physiological properties of Pyropia haitanensis (Bangiales, Rhodophyta). J. Appl. Phycol. 2016, 28, 1235–1243. [Google Scholar] [CrossRef]
- Latorre, N.; Castañeda, F.; Meynard, A.; Rivas, J.; Contreras, L. First approach of characterization of bioactive compound in Pyropia orbicularis during the daily tidal cycle. Lat. Am. J. Aquat. 2019, 47, 826–840. [Google Scholar] [CrossRef]
- Zapata, J.; Meynard, A.; Anguita, C.; Espinoza, C.; Alvear, P.; Kumar, M.; Contreras-Porcia, L. Non-Random distribution and ecophysiological differentiation of Pyropia species (Bangiales, Rhodophyta) through environmental gradients. J. Phycol. 2019, 55, 1140–1153. [Google Scholar] [CrossRef] [PubMed]
- Lalegerie, F.; Gager, L.; Stiger-Pouvreau, V.; Connan, S. The stressful life of red and brown seaweeds on the temperate intertidal zone: Effect of abiotic and biotic parameters on the physiology of macroalgae and content variability of particular metabolites. In Advances in Botanical Research; Elsevier: Amsterdam, The Netherlands, 2020; Volume 95, pp. 247–287. ISBN 978-0-08-102710-3. [Google Scholar]
- Pereira, D.T.; Simioni, C.; Filipin, E.P.; Bouvie, F.; Ramlov, F.; Maraschin, M.; Bouzon, Z.L.; Schmidt, É.C. Effects of salinity on the physiology of the red macroalga, Acanthophora spicifera (Rhodophyta, Ceramiales). Acta Bot. Bras. 2017, 31, 555–565. [Google Scholar] [CrossRef] [Green Version]
- Cazzaniga, S.; Bressan, M.; Carbonera, D.; Agostini, A.; Dall’Osto, L. Differential Roles of Carotenes and Xanthophylls in Photosystem I Photoprotection. Biochemistry 2016, 55, 3636–3649. [Google Scholar] [CrossRef] [PubMed]
- Havaux, M.; Dall’Osto, L.; Bassi, R. Zeaxanthin Has Enhanced Antioxidant Capacity with Respect to All Other Xanthophylls in Arabidopsis Leaves and Functions Independent of Binding to PSII Antennae. Plant. Physiology. 2007, 145, 1506–1520. [Google Scholar] [CrossRef] [Green Version]
- Korbee, N.; Lopez Figueroa, F.; Aguilera, J. Effect of light quality on the accumulation of photosynthetic pigments, proteins and mycosporine-like amino acids in the red alga Porphyra leucosticta (Bangiales, Rhodophyta). J. Photobiol. 2005, 80, 71–78. [Google Scholar] [CrossRef]
- Meynard, A.; Zapata, J.; Salas, N.; Betancourtt, C.; Pérez-Lara, G.; Castañeda, F.; Ramírez, M.E.; Bulboa Contador, C.; Guillemin, M.-L.; Contreras-Porcia, L. Genetic and morphological differentiation of Porphyra and Pyropia species (Bangiales, Rhodophyta) coexisting in a rocky intertidal in Central Chile. J. Phycol. 2019, 55, 297–313. [Google Scholar] [CrossRef]
- Schweikert, K.; Sutherland, J.E.; Burritt, D.J.; Hurd, C.L. Analysis of spatial and temporal diversity and distribution of Porphyra (Rhodophyta) in southeastern New Zealand supported by the use of molecular tools. J. Phycol. 2012, 48, 530–538. [Google Scholar] [CrossRef]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; PRISMA Group. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef] [Green Version]
- Gury, M.D.; Gury, G.M. Algaebase, World-Wide Electronic Publication. Available online: https://www.algaebase.org/ (accessed on 12 January 2021).
- RStudio Team. RStudio: Integrated Development for R; RStudio, PBC: Boston, MA, USA, 2020. [Google Scholar]
- Borenstein, M. (Ed.) Introduction to Meta-Analysis; John Wiley & Sons: Chichester, UK, 2009; ISBN 978-0-470-05724-7. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Exclusion Criteria | Articles Excluded |
|---|---|
| 1. Species different from Porphyra spp. or Pyropia spp. were used. | 11 |
| 2. Carotenoid content was not reported. | 50 |
| 3. The experimental factor evaluated was a biological factor. | 1 |
| 4. The carotenoid content was evaluated at baseline, without the alteration of an experimental factor. | 13 |
| 5. The methodology of the experimental factor or treatment was not descriptively explained. | 1 |
| 6. The control condition was not reported. | 3 |
| 7. The magnitude of the experimental factor was not specified. | 1 |
| 8. Experimental factor or treatment was evaluated on the conchocelis, spores or reproductive structures. | 6 |
| 9. Article was not available in electronic databases. | 6 |
| Genus | Specie | Total Carotenoids | Lutein | Zeaxanthin | Measure Method | Ref. |
|---|---|---|---|---|---|---|
| Porphyra | Po. haitanensis | 62.5–10,000 | - | - | SP 1 | [32,33,34,35] |
| Po. sp. | - | 430–1117 | - | LC-UV 2 | [36,37] | |
| Po. acanthophora var. brasiliensis | - | 4.16–30.71 | 4.43–36.31 | LC-UV | [38] | |
| Po. umbilicalis | + | - | - | SP | [39,40] | |
| Pyropia | Py. orbicularis | 2.2–170.84 | - | - | SP | [41,42] |
| Py. acanthophora var. brasiliensis | 3.63–12.4 | 3.22–6.1 | 0.07–0.15 | SP/LC-UV | [43,44] | |
| Py. yezoensis | 225–2.2 × 104 | 536–1137 | 2.36–17.58 | SP/LC-UV | [45,46,47,48,49,50,51] | |
| Py. spiralis | 500–1.5 × 104 | - | - | SP | [52] | |
| Py. haitanensis | 280–800 | - | - | SP | [53,54,55,56] |
| Variable | Classification of Variables | N° of Articles |
|---|---|---|
| Genus | Porphyra | 9 |
| Pyropia | 16 | |
| Measured compound | Total carotenoids | 21 |
| Lutein | 5 | |
| Zeaxanthin | 3 | |
| Measure method | LC-UV | 5 |
| Spectrophotometry | 20 | |
| Experimental factor | Salinity (S) | 2 |
| Wavelength (WL) | 4 | |
| Light Intensity (LI) | 5 | |
| Nutrients Composition (NC) | 4 | |
| Algal/Culture Density (AD) | 2 | |
| Desiccation (D) | 4 | |
| CO2 (CO) | 5 | |
| Heavy Metals (HM) | 2 | |
| Photoperiod (PP) | 2 | |
| pH (PH) | 1 | |
| Temperature (T) | 5 | |
| Treatments | S | 2 |
| WL | 3 | |
| WL-NC | 1 | |
| LI | 2 | |
| LI-CO | 1 | |
| NC | 1 | |
| NC-CO | 2 | |
| AD-CO | 2 | |
| D | 3 | |
| D-T | 1 | |
| HM | 2 | |
| PP-T-LI | 1 | |
| PH-LI | 1 | |
| T | 2 | |
| T-PP | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Piña, F.; Contreras-Porcia, L. Enhancement of Xanthophyll Synthesis in Porphyra/Pyropia Species (Rhodophyta, Bangiales) by Controlled Abiotic Factors: A Systematic Review and Meta-Analysis. Mar. Drugs 2021, 19, 221. https://doi.org/10.3390/md19040221
Piña F, Contreras-Porcia L. Enhancement of Xanthophyll Synthesis in Porphyra/Pyropia Species (Rhodophyta, Bangiales) by Controlled Abiotic Factors: A Systematic Review and Meta-Analysis. Marine Drugs. 2021; 19(4):221. https://doi.org/10.3390/md19040221
Chicago/Turabian StylePiña, Florentina, and Loretto Contreras-Porcia. 2021. "Enhancement of Xanthophyll Synthesis in Porphyra/Pyropia Species (Rhodophyta, Bangiales) by Controlled Abiotic Factors: A Systematic Review and Meta-Analysis" Marine Drugs 19, no. 4: 221. https://doi.org/10.3390/md19040221
APA StylePiña, F., & Contreras-Porcia, L. (2021). Enhancement of Xanthophyll Synthesis in Porphyra/Pyropia Species (Rhodophyta, Bangiales) by Controlled Abiotic Factors: A Systematic Review and Meta-Analysis. Marine Drugs, 19(4), 221. https://doi.org/10.3390/md19040221