
Electrochromic Properties and Electrochemical Behavior of Marennine, a Bioactive Blue-Green Pigment Produced by the Marine Diatom Haslea ostrearia

 , , , ,
, , , ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
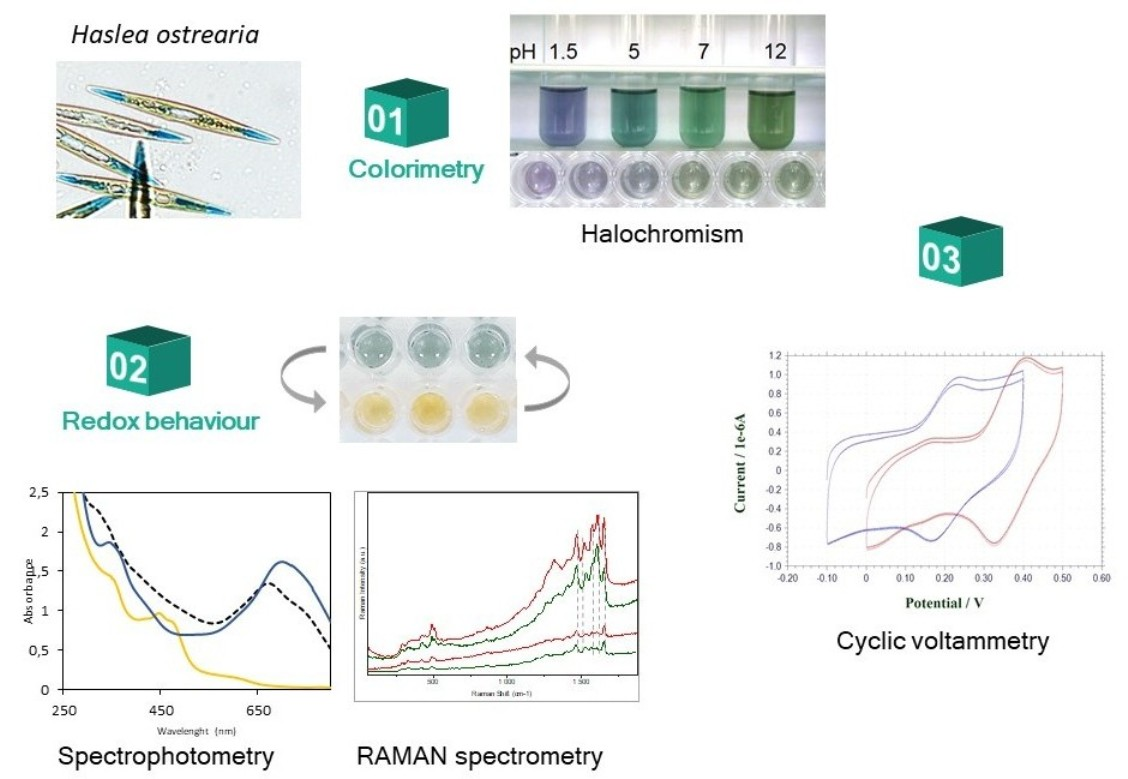
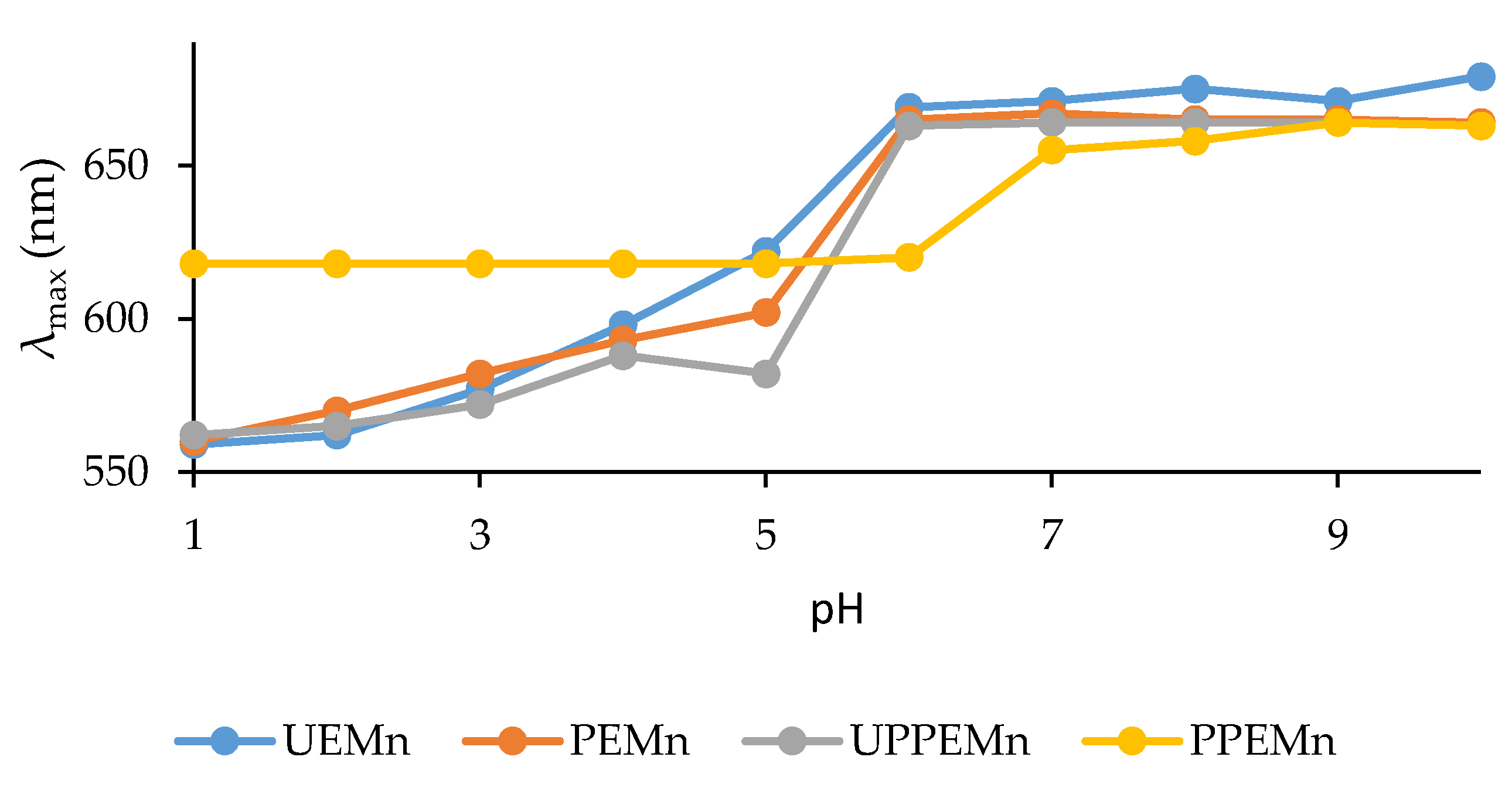
2.1. Study of Marennine Color at Different pH
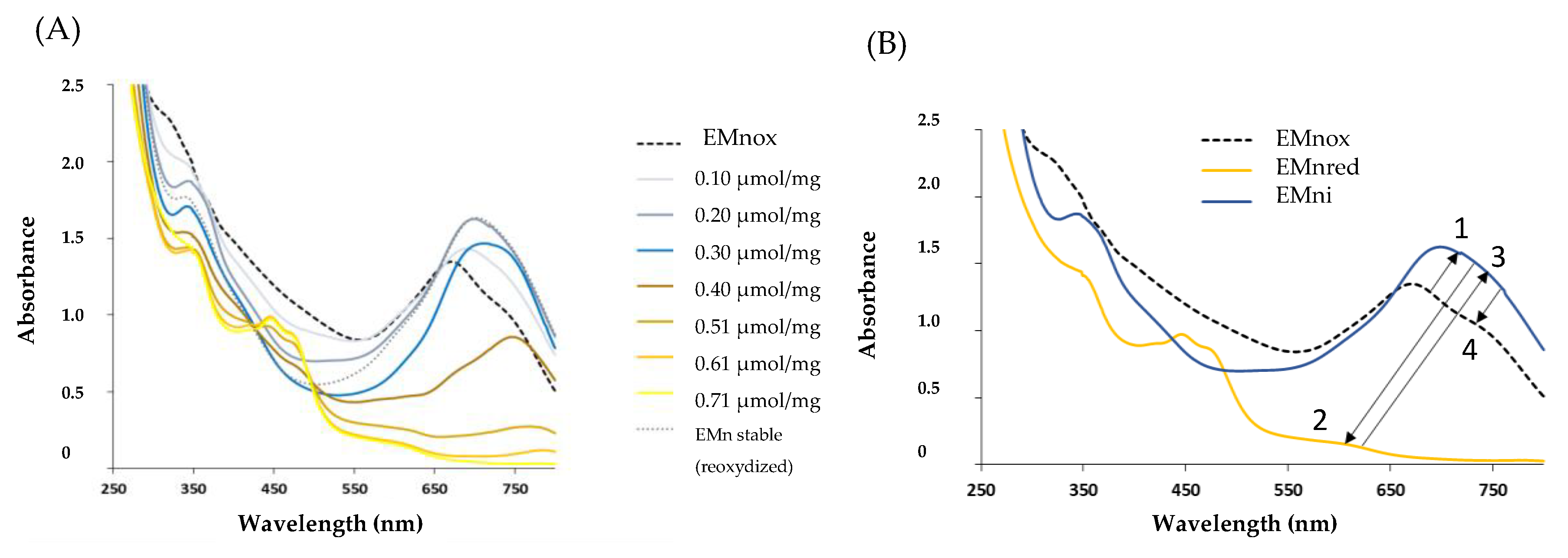
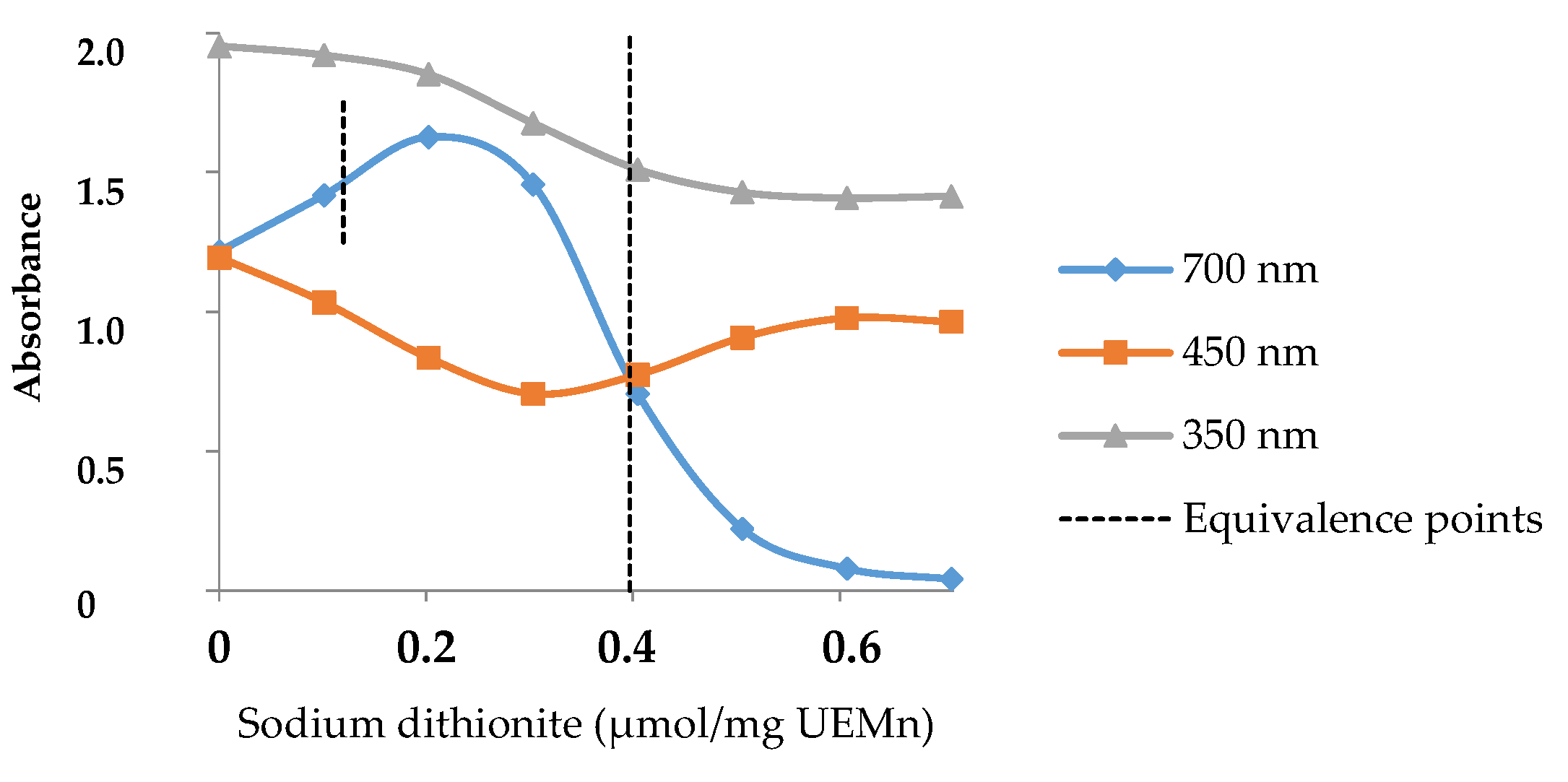
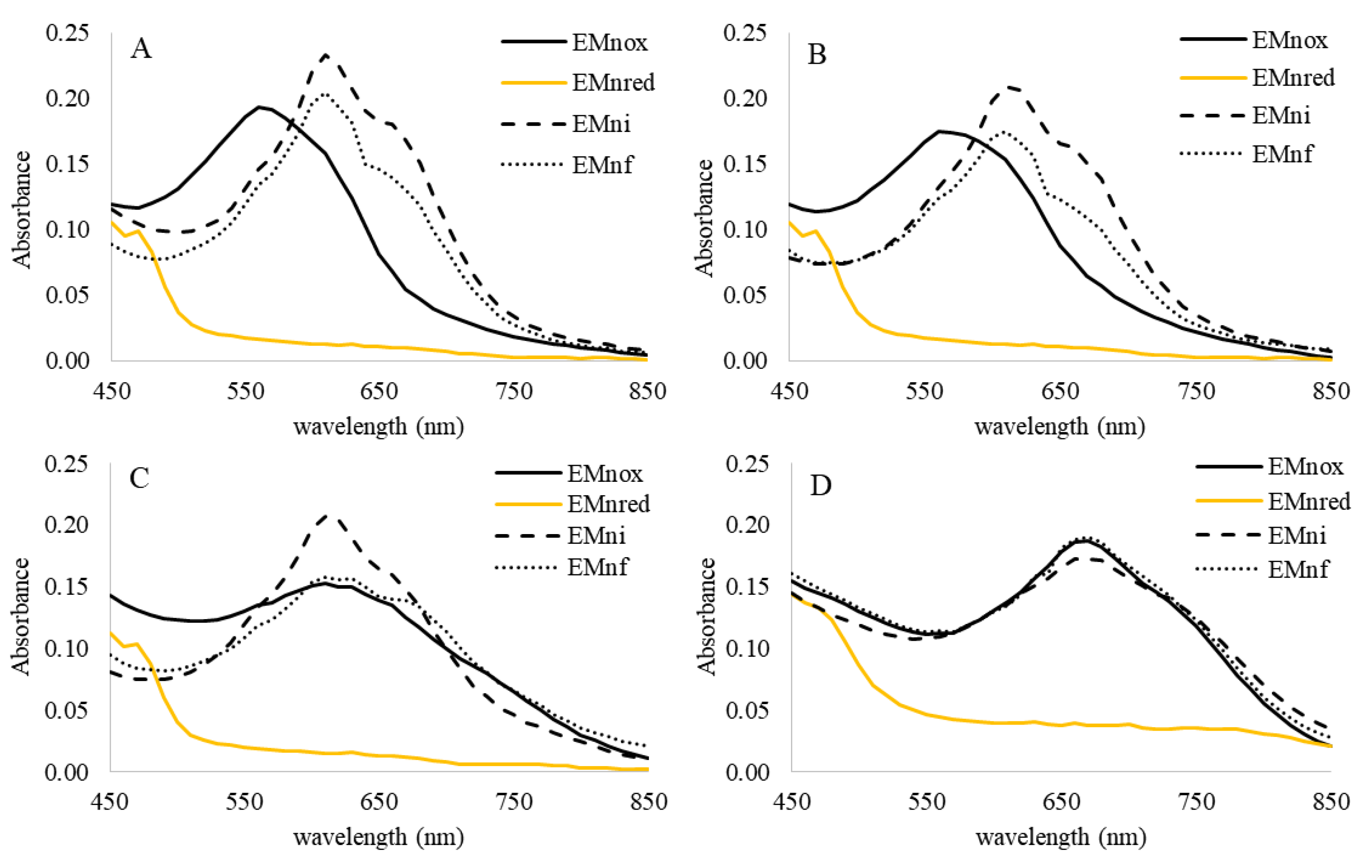
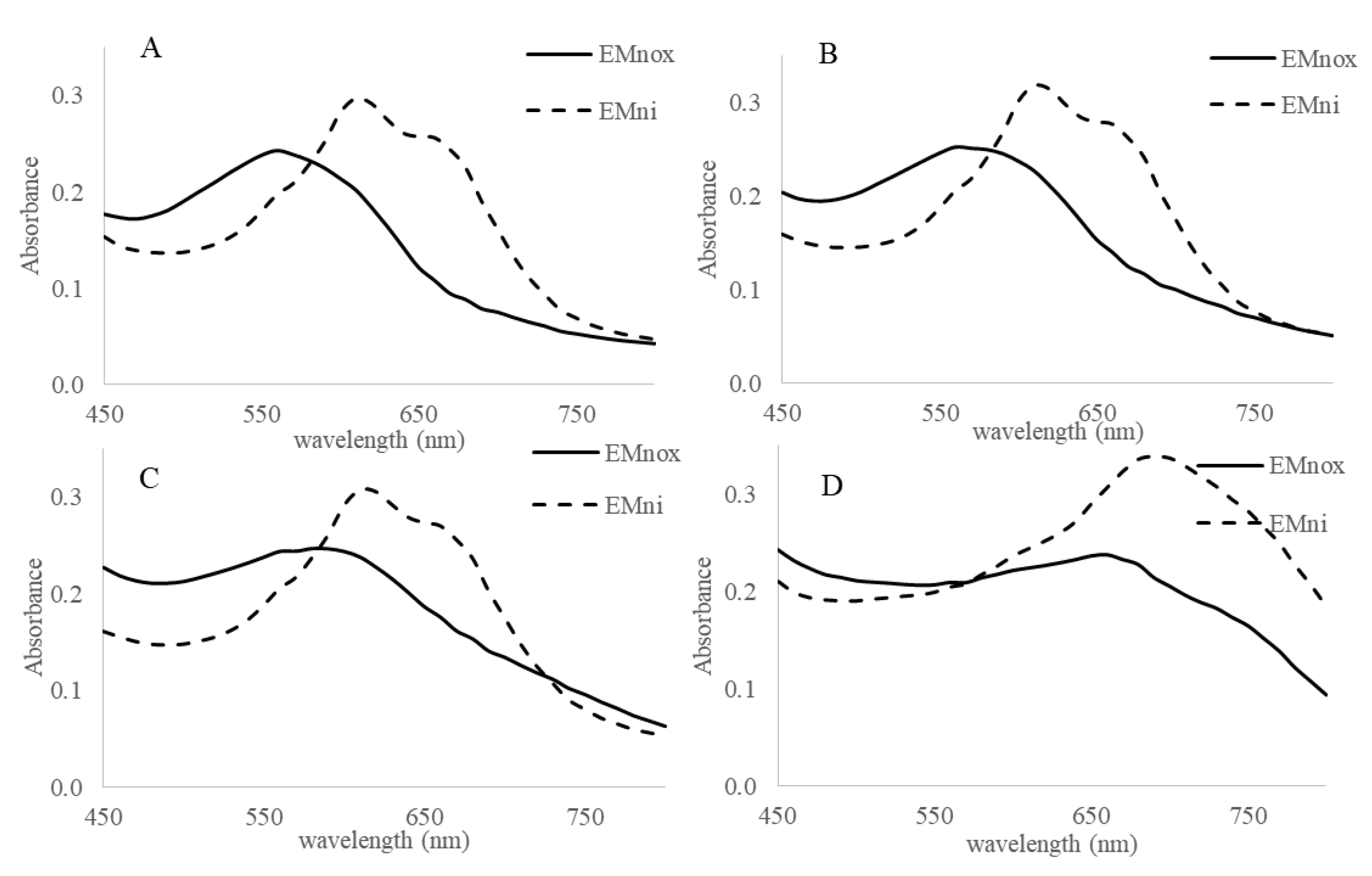
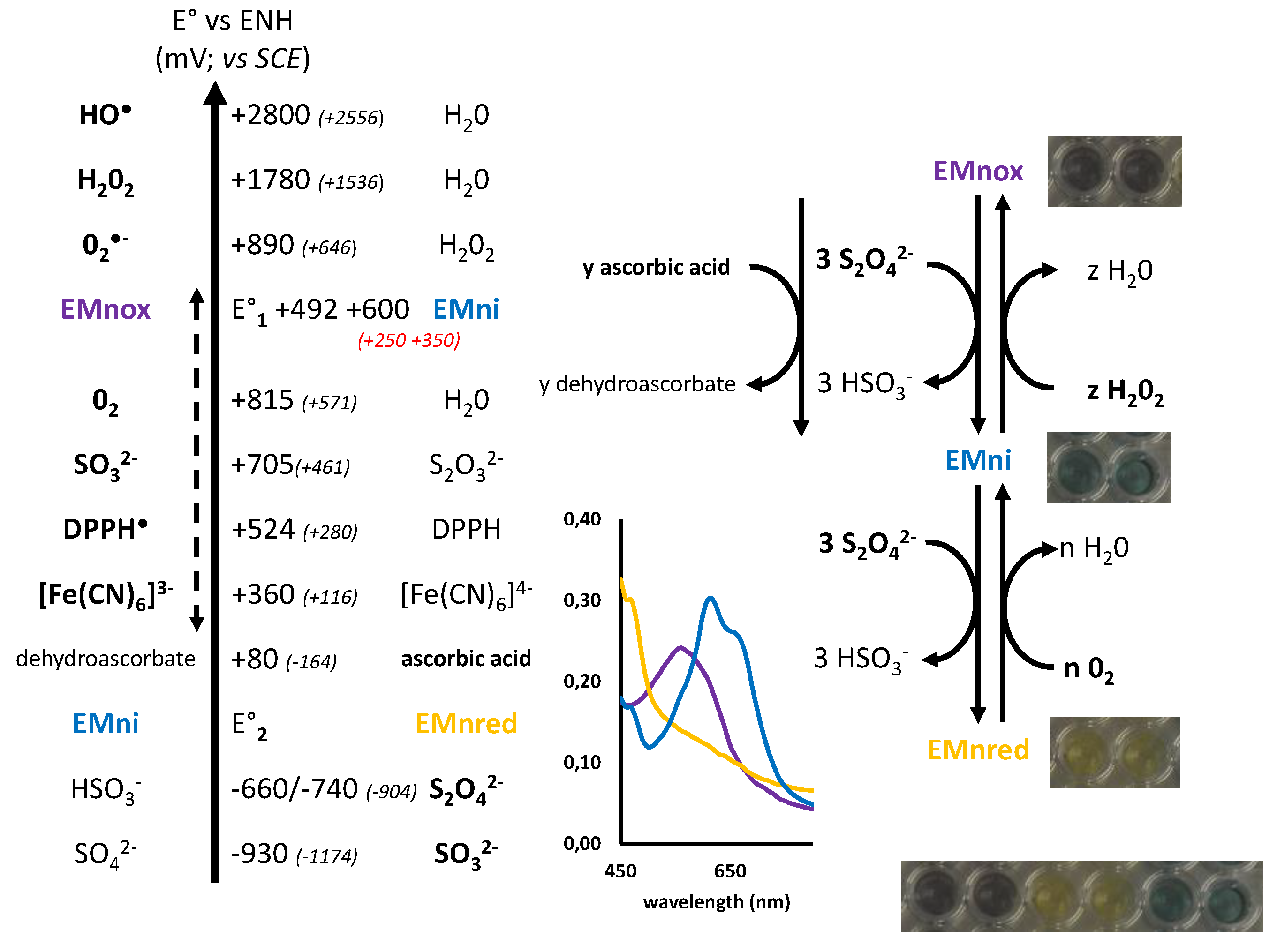
2.2. Marennine Redox Properties and Color Change
2.3. Marennine Reaction with Ascorbic Acid and Sodium Sulfite
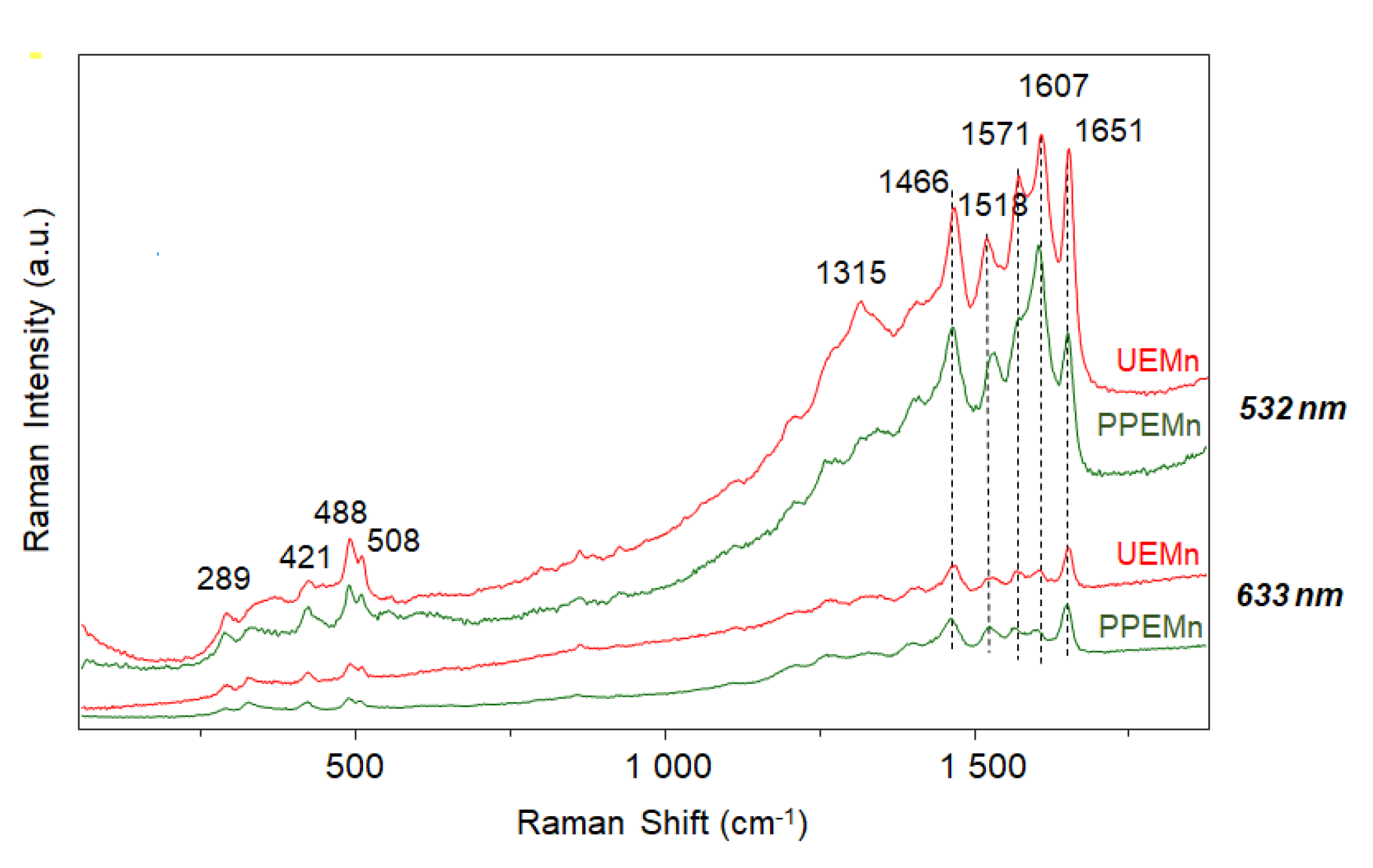
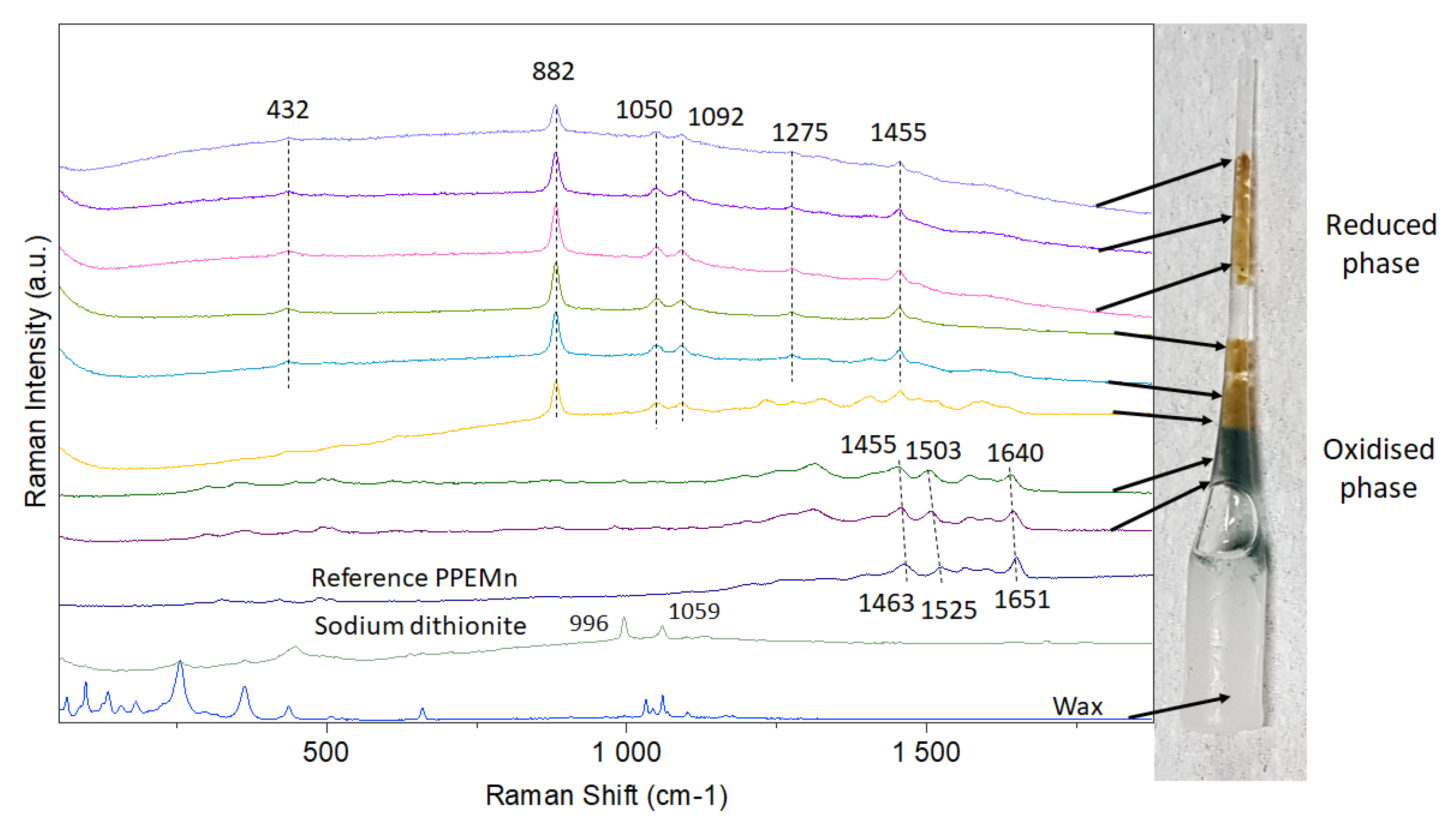
2.4. Raman Spectroscopy
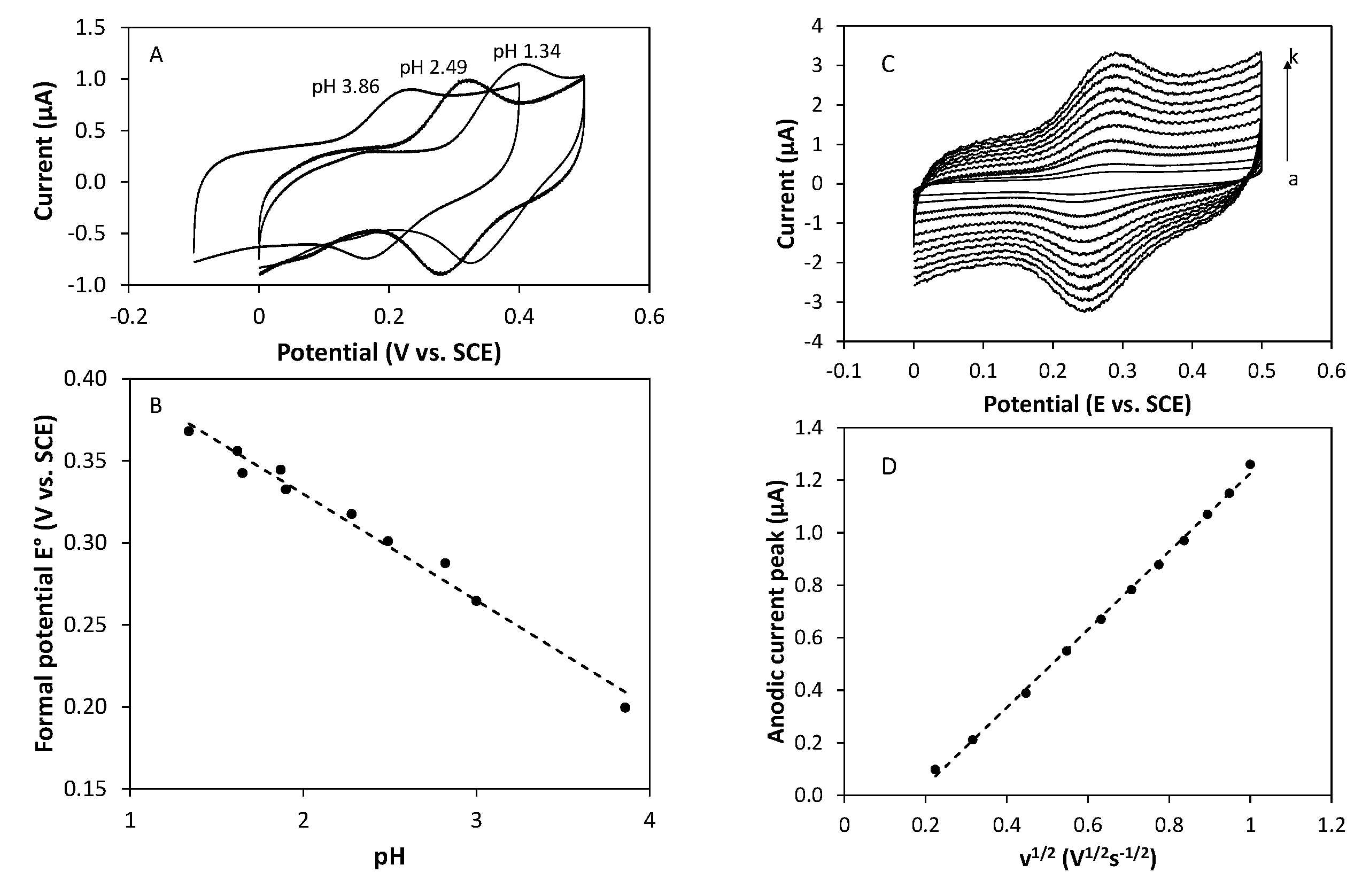
2.5. Electrochemical Behavior
3. Materials and Methods
3.1. Algal Production and Marennine Purification
3.2. Redox Experiments in Solution and Color Change
3.3. Resonance Raman Spectroscopy
3.4. Cyclic Voltammetry
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Puységur, M. Notice sur la cause du verdissement des huîtres. Rev. Marit. Coloniale 1880, 64, 248–256. [Google Scholar]
- Gastineau, R.; Prasetiya, F.S.; Falaise, C.; Cognie, B.; Decottignies, P.; Morançais, M.; Méléder, V.; Davidovich, N.; Turcotte, T.; Tremblay, R.; et al. Marennine-like pigments: Blue diatom or green oyster cult. In Blue Biotechnology: Production and Use of Marine Molecules; Stephane La Barre, S.S.B., Ed.; Wiley-VCH Verlag GmbH & Co. KGaA: Weinheim, Germany, 2018. [Google Scholar]
- Gaillon, B. Des huîtres vertes, et des causes de cette coloration. J. Phys. Chim. D’histoire Nat. Arts 1820, 91, 222–225. [Google Scholar]
- Bory, J.B. Navicule; Navicula, Encyclopédie Méthodique. Histoire Naturelle des Zoophytes, ou Animaux Rayonnés, Faisant Suite à L’histoire Naturelle des vers de Bruguièr; Mme Veuve Agasse: Paris, France, 1824; Volume: Tome 2. [Google Scholar]
- Simonsen, R. The diatom plankton of the Indian Ocean Expedition of R/V ‘Meteor’ 1964–1965. Meteor’ Forsch. Reihe 1974, 19, 1–107. [Google Scholar]
- Poulin, M.; Méléder, V.; Mouget, J.-L. Typification of the first blue pigmented diatom, Haslea ostrearia (Gaillon) Simonsen (Bacillariophyceae). Plant Ecol. Evol. 2019, 152, 402–408. [Google Scholar] [CrossRef]
- Lankester, E.R. On green oysters. Q. J. Microsc. Sci. 1886, 26, 71–94. [Google Scholar] [CrossRef]
- Pouvreau, J.B.; Housson, E.; Le Tallec, L.; Morançais, M.; Rincé, Y.; Fleurence, J.; Pondaven, P. Growth inhibition of several marine diatom species induced by the shading effect and allelopathic activity of marennine, a blue-green polyphenolic pigment of the diatom Haslea ostrearia (Gaillon/Bory) Simonsen. J. Exp. Mar. Biol. Ecol. 2007, 352, 212–225. [Google Scholar] [CrossRef]
- Pouvreau, J.B.; Taran, F.; Rosa, P.; Pin, S.; Fleurence, J.; Pondaven, P. Antioxidant and free radical scavenging properties of marennine, a blue-green polyphenolic pigment from the diatom Haslea ostrearia (Gaillon/Bory) Simonsen responsible for the natural greening of cultured oysters. J. Agric. Food Chem. 2008, 56, 6278–6286. [Google Scholar] [CrossRef] [PubMed]
- Gastineau, R.; Pouvreau, J.-B.; Hellio, C.; Morançais, M.; Fleurence, J.; Gaudin, P.; Bourgougnon, N.; Mouget, J.-L. Biological Activities of Purified Marennine, the Blue Pigment Responsible for the Greening of Oysters. J. Agric. Food Chem. 2012, 60, 3599–3605. [Google Scholar] [CrossRef] [PubMed]
- Prasetiya, F.S.; Safitri, I.; Widowati, I.; Cognie, B.; Decottignies, P.; Gastineau, R.; Morançais, M.; Windarto, E.; Tremblay, R.; Mouget, J.-L. Does allelopathy affect co-culturing Haslea ostrearia with other microalgae relevant to aquaculture? Environ. Boil. Fishes 2016, 28, 2241–2254. [Google Scholar] [CrossRef]
- Prasetiya, F.S.; Decottignies, P.; Tremblay, R.; Mouget, J.-L.; Sunarto, S.; Iskandar, I.; Dhahiyat, Y.; Cognie, B. Not only greening: The effects of marennine produced by Haslea ostrearia on physiological traits of three bivalve species. Aquac. Rep. 2020, 18, 100546. [Google Scholar] [CrossRef]
- Falaise, C.; Cormier, P.; Tremblay, R.; Audet, C.; Deschênes, J.-S.; Turcotte, F.; François, C.; Seger, A.; Hallegraeff, G.; Lindquist, N.; et al. Harmful or harmless: Biological effects of marennine on marine organisms. Aquat. Toxicol. 2019, 209, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Prasetiya, F.S.; Decottignies, P.; Tremblay, R.; Mouget, J.-L.; Cognie, B. Does culture supernatant of Haslea ostrearia containing marennine affect short-term physiological traits in the adult blue mussel Mytilus edulis? Aquac. Rep. 2019, 15, 100228. [Google Scholar] [CrossRef]
- Falaise, C.; James, A.; Travers, M.-A.; Zanella, M.; Badawi, M.; Mouget, J.-L. Complex Relationships between the Blue Pigment Marennine and Marine Bacteria of the Genus Vibrio. Mar. Drugs 2019, 17, 160. [Google Scholar] [CrossRef] [Green Version]
- Turcotte, F.; Mouget, J.-L.; Genard, B.; Lemarchand, K.; Deschênes, J.-S.; Tremblay, R. Prophylactic effect of Haslea ostrearia culture supernatant containing the pigment marennine to stabilize bivalve hatchery production. Aquat. Living Resour. 2016, 29, 401. [Google Scholar] [CrossRef] [Green Version]
- Martins, N.; Roriz, C.L.; Morales, P.; Barros, L.; Ferreira, I.C. Food colorants: Challenges, opportunities and current desires of agro-industries to ensure consumer expectations and regulatory practices. Trends Food Sci. Technol. 2016, 52, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Buchweitz, M. Natural solutions for blue colors in food. In Handbook on Natural Pigments in Food and Beverages; Woodhead Publishing: Cambridge, MA, USA, 2016; pp. 355–384. [Google Scholar]
- Research, A.M. Spirulina Market by Type (Arthrospira Platensis and Arthrospira Maxima), Application (Nutraceuticals, Cosmetics, Food & Beverages, Animal Feed, and Others), and Drug Formulation (Powder, Tablet & Capsule, Liquid, and Granule & Gelling Agent): Global Opportunity Analysis and Industry Forecast. Available online: https://www.alliedmarketresearch.com/spirulina-market (accessed on 12 January 2020).
- Gastineau, R.; Turcotte, F.; Pouvreau, J.-B.; Morançais, M.; Fleurence, J.; Windarto, E.; Prasetiya, F.S.; Arsad, S.; Jaouen, P.; Babin, M.; et al. Marennine, Promising Blue Pigments from a Widespread Haslea Diatom Species Complex. Mar. Drugs 2014, 12, 3161–3189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pouvreau, J.B.; Morançais, M.; Fleury, F.; Rosa, P.; Thion, L.; Cahingt, B.; Zal, F.; Fleurence, J.; Pondaven, P. Preliminary characterisation of the blue-green pigment “marennine” from the marine tychopelagic diatom Haslea ostrearia (Gaillon/Bory) Simonsen. J. Appl. Phycol. 2006, 18, 757–767. [Google Scholar] [CrossRef]
- Pouvreau, J.-B.; Morançais, M.; Massé, G.; Rosa, P.; Robert, J.-M.; Fleurence, J.; Pondaven, P. Purification of the blue-green pigment “marennine” from the marine tychopelagic diatom Haslea ostrearia (Gaillon/Bory) Simonsen. Environ. Boil. Fishes 2006, 18, 769–781. [Google Scholar] [CrossRef]
- Bocat, L. Sur la marennine de la diatomée bleue: Comparaison avec la phycocyanine. Comptes Rendus Séances Société Biol. Ses Fil. 1907, 62, 1073–1075. [Google Scholar]
- Newsome, A.G.; Culver, C.A.; Van Breemen, R.B. Nature’s Palette: The Search for Natural Blue Colorants. J. Agric. Food Chem. 2014, 62, 6498–6511. [Google Scholar] [CrossRef]
- Lambeth, D.O.; Palmer, G. The kinetics and mechanism of reduction of electron transfer proteins and other compounds of biological interest by dithionite. J. Biol. Chem. 1973, 248, 6095–6103. [Google Scholar] [CrossRef]
- de Carvalho, L.M.; Schwedt, G. Polarographic determination of dithionite and its decomposition products: Kinetic aspects, stabilizers, and analytical application. Anal. Chim. Acta 2001, 436, 293–300. [Google Scholar] [CrossRef]
- Kudrik, E.V.; Makarov, S.V.; Zahl, A.; van Eldik, R. Kinetics and mechanism of the iron phthalocyanine catalyzed reduction of nitrite by dithionite and sulfoxylate in aqueous solution. Inorg. Chem. 2005, 44, 6470–6475. [Google Scholar] [CrossRef] [PubMed]
- Srividya, N.; Paramasivan, G.; Seetharaman, K.; Ramamurthy, P. Two-step reduction of indigo carmine by dithionite: A stopped-flow study. J. Chem. Soc. Faraday Trans. 1994, 90, 2525–2530. [Google Scholar] [CrossRef]
- Mayhew, S.G. The redox potential of dithionite and SO− 2 from equilibrium reactions with flavodoxins, methyl viologen and hydrogen plus hydrogenase. Eur. J. Biochem. 1978, 85, 535–547. [Google Scholar] [CrossRef]
- Mimuro, M.; Füglistaller, P.; Rümbeli, R.; Zuber, H. Functional assignment of chromophores and energy transfer in C phycocyanin isolated from the thermophilic cyanobacterium Mastigocladus laminosus. Biochim. Biophys. Acta (BBA) Bioenerg. 1986, 848, 155–166. [Google Scholar] [CrossRef]
- Fouillaud, M.; Venkatachalam, M.; Girard-Valenciennes, E.; Caro, Y.; Dufossé, L. Anthraquinones and Derivatives from Marine-Derived Fungi: Structural Diversity and Selected Biological Activities. Mar. Drugs 2016, 14, 64. [Google Scholar] [CrossRef] [Green Version]
- Hedegaard, C.; Bardeau, J.-F.; Chateigner, D. Molluscan Shell Pigments: An in situ Resonance Raman Study. J. Molluscan Stud. 2006, 72, 157–162. [Google Scholar] [CrossRef]
- Trinkler, N.; Bardeau, J.-F.; Marin, F.; LaBonne, M.; Jolivet, A.; Crassous, P.; Paillard, C. Mineral phase in shell repair of Manila clam Venerupis philippinarum affected by brown ring disease. Dis. Aquat. Org. 2011, 93, 149–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Izumi, C.M.; Constantino, V.R.; Ferreira, A.M.C.; Temperini, M.L. Spectroscopic characterization of polyaniline doped with transition metal salts. Synth. Met. 2006, 156, 654–663. [Google Scholar] [CrossRef]
- Fatyeyeva, K.; Pud, A.A.; Bardeau, J.-F.; Tabellout, M. Structure–property relationship in aliphatic polyamide/polyaniline surface layered composites. Mater. Chem. Phys. 2011, 130, 760–768. [Google Scholar] [CrossRef]
- Cabrera-Alonso, R.; Guevara, E.; Ramírez-Elías, M.G.; Moncada, B.; González, F.J. Surface-enhanced Raman scattering of hydroquinone assisted by gold nanorods. J. Nanophotonics 2019, 13, 036006–036016. [Google Scholar] [CrossRef]
- Kubinyi, M.; Billes, F.; Grofcsik, A.; Keresztury, G. Vibrational spectra and normal coordinate analysis of phenol and hydroquinone. J. Mol. Struct. 1992, 266, 339–344. [Google Scholar] [CrossRef]
- Gastineau, R.; Davidovich, N.A.; Bardeau, J.-F.; Caruso, A.; Leignel, V.; Hardivillier, Y.; Jacquette, B.; Davidovich, O.I.; Rincé, Y.; Gaudin, P.; et al. Haslea karadagensis (Bacillariophyta): A second blue diatom, recorded from the Black Sea and producing a novel blue pigment. Eur. J. Phycol. 2012, 47, 469–479. [Google Scholar] [CrossRef] [Green Version]
- Gastineau, R.; Hansen, G.; Davidovich, N.A.; Davidovich, O.; Bardeau, J.-F.; Kaczmarska, I.; Ehrman, J.M.; Leignel, V.; Hardivillier, Y.; Jacquette, B.; et al. A new blue-pigmented hasleoid diatom, Haslea provincialis, from the Mediterranean Sea. Eur. J. Phycol. 2016, 51, 156–170. [Google Scholar] [CrossRef] [Green Version]
- Prasetiya, F.S.; Gastineau, R.; Poulin, M.; Lemieux, C.; Turmel, M.; Syakti, A.D.; Hardivillier, Y.; Widowati, I.; Risjani, Y.; Iskandar, I.; et al. Haslea nusantara (Bacillariophyceae), a new blue diatom from the Java Sea, Indonesia: Morphology, biometry and molecular characterization. Plant Ecol. Evol. 2019, 152, 188–202. [Google Scholar] [CrossRef]
- Bard, A.J.; Faulkner, L.R. Electrochemical Methods, Fundamentals and Applications, 2nd ed.; Wiley: New York, NY, USA, 2001. [Google Scholar]
- Mouget, J.-L.; Gastineau, R.; Davidovich, O.; Gaudin, P.; Davidovich, N. Light is a key factor in triggering sexual reproduction in the pennate diatom Haslea ostrearia. FEMS Microbiol. Ecol. 2009, 69, 194–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pouvreau, J.-B.; Morançais, M.; Fleurence, J.; Pondaven, P. Method for the quantification of the blue-green pigment “marennine” synthesized by the marine diatom Haslea ostrearia (Gaillon/Bory) Simonsen using HPLC gel-filtration and photodiode-array detection. Environ. Boil. Fishes 2006, 19, 263–270. [Google Scholar] [CrossRef]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Francezon, N.; Herbaut, M.; Bardeau, J.-F.; Cougnon, C.; Bélanger, W.; Tremblay, R.; Jacquette, B.; Dittmer, J.; Pouvreau, J.-B.; Mouget, J.-L.; et al. Electrochromic Properties and Electrochemical Behavior of Marennine, a Bioactive Blue-Green Pigment Produced by the Marine Diatom Haslea ostrearia. Mar. Drugs 2021, 19, 231. https://doi.org/10.3390/md19040231
Francezon N, Herbaut M, Bardeau J-F, Cougnon C, Bélanger W, Tremblay R, Jacquette B, Dittmer J, Pouvreau J-B, Mouget J-L, et al. Electrochromic Properties and Electrochemical Behavior of Marennine, a Bioactive Blue-Green Pigment Produced by the Marine Diatom Haslea ostrearia. Marine Drugs. 2021; 19(4):231. https://doi.org/10.3390/md19040231
Chicago/Turabian StyleFrancezon, Nellie, Mickaël Herbaut, Jean-François Bardeau, Charles Cougnon, William Bélanger, Réjean Tremblay, Boris Jacquette, Jens Dittmer, Jean-Bernard Pouvreau, Jean-Luc Mouget, and et al. 2021. "Electrochromic Properties and Electrochemical Behavior of Marennine, a Bioactive Blue-Green Pigment Produced by the Marine Diatom Haslea ostrearia" Marine Drugs 19, no. 4: 231. https://doi.org/10.3390/md19040231
APA StyleFrancezon, N., Herbaut, M., Bardeau, J. -F., Cougnon, C., Bélanger, W., Tremblay, R., Jacquette, B., Dittmer, J., Pouvreau, J. -B., Mouget, J. -L., & Pasetto, P. (2021). Electrochromic Properties and Electrochemical Behavior of Marennine, a Bioactive Blue-Green Pigment Produced by the Marine Diatom Haslea ostrearia. Marine Drugs, 19(4), 231. https://doi.org/10.3390/md19040231