
OVAT Analysis and Response Surface Methodology Based on Nutrient Sources for Optimization of Pigment Production in the Marine-Derived Fungus Talaromyces albobiverticillius 30548 Submerged Fermentation

 ,
, 
 ,
,  and
and 
Abstract
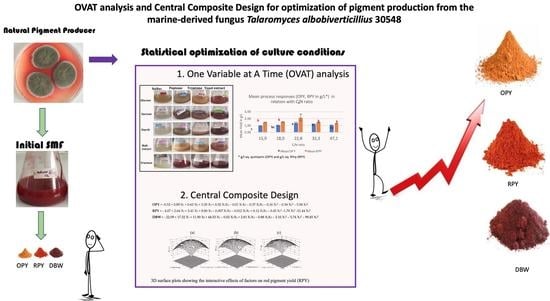
:
1. Introduction
2. Materials and Methods
2.1. Microorganism and Cultivation
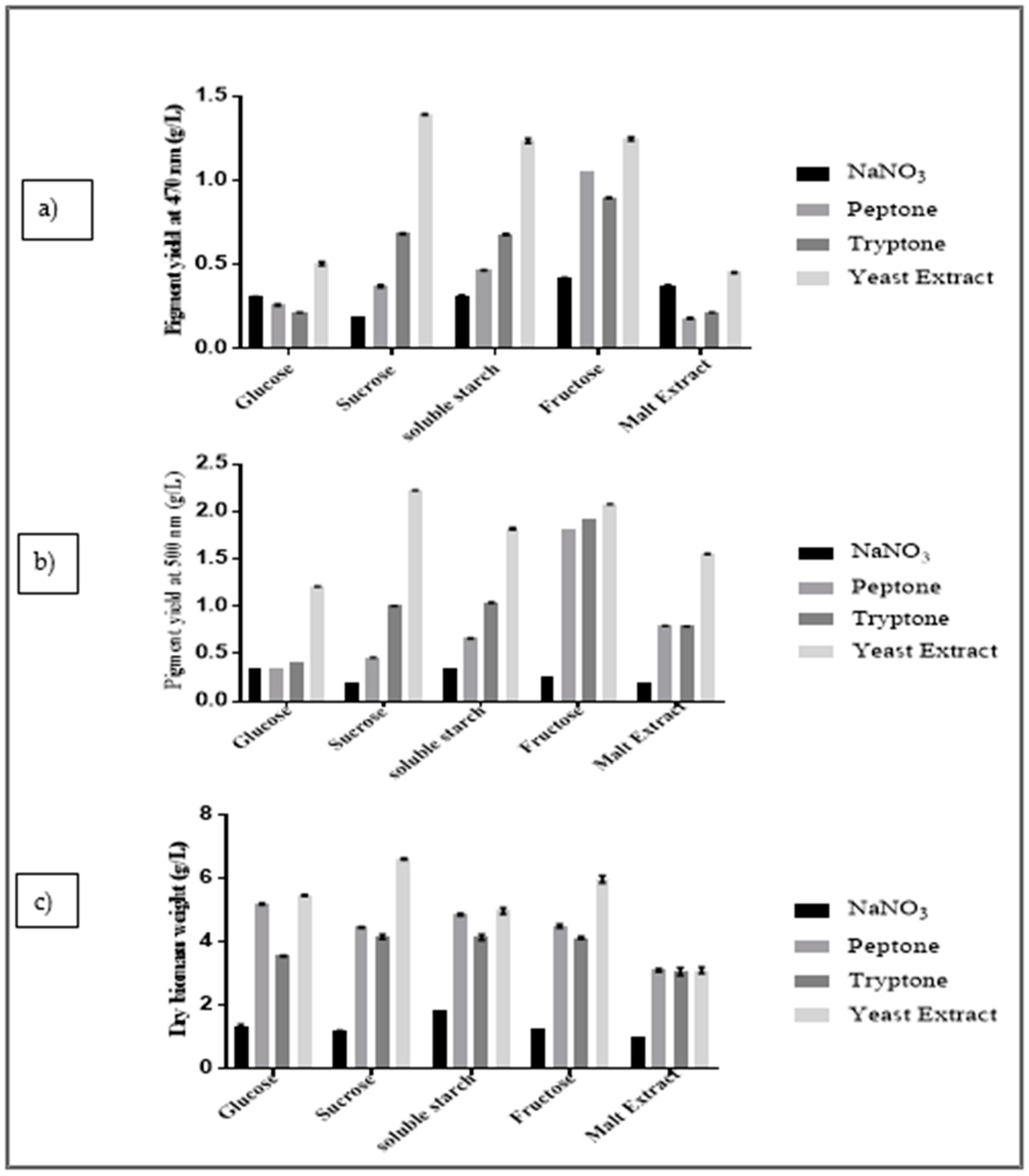
2.2. Primary Screening of Medium Components by OVAT Analysis
2.3. Submerged Fermentations
2.4. Quantification of Pigments
2.5. Dry Biomass Concentration
2.6. Optimization Using Central Composite Design (CCD) and Response Surface Modeling (RSM)
2.7. Statistical Analysis and Data Validation
2.8. C/N Ratio Calculation and Means Comparison
3. Results
3.1. Primary Screening of Media Components Using OVAT Analysis
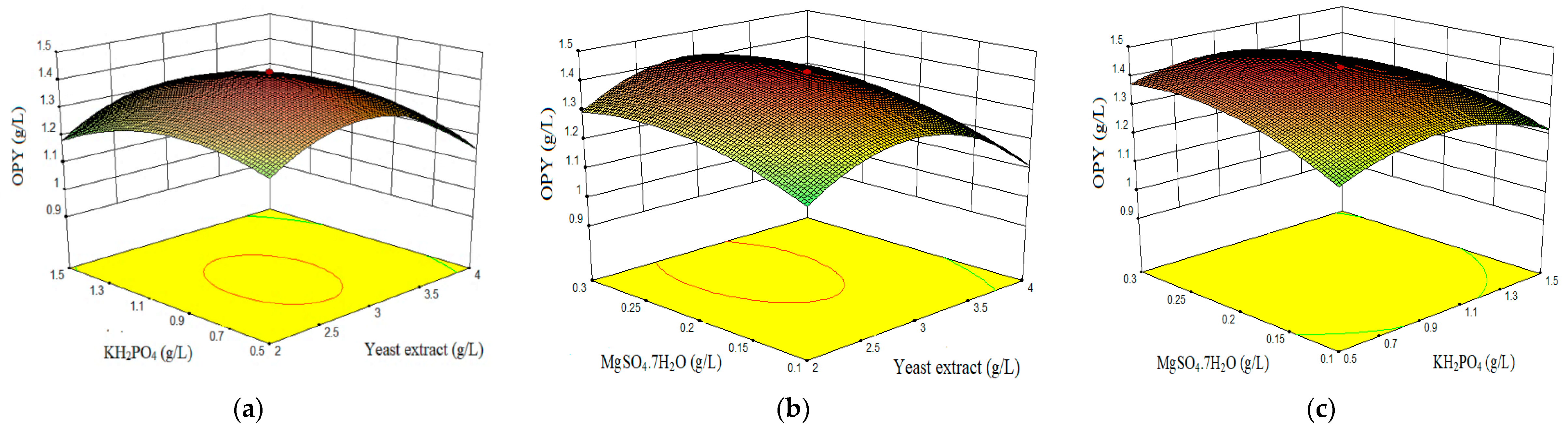
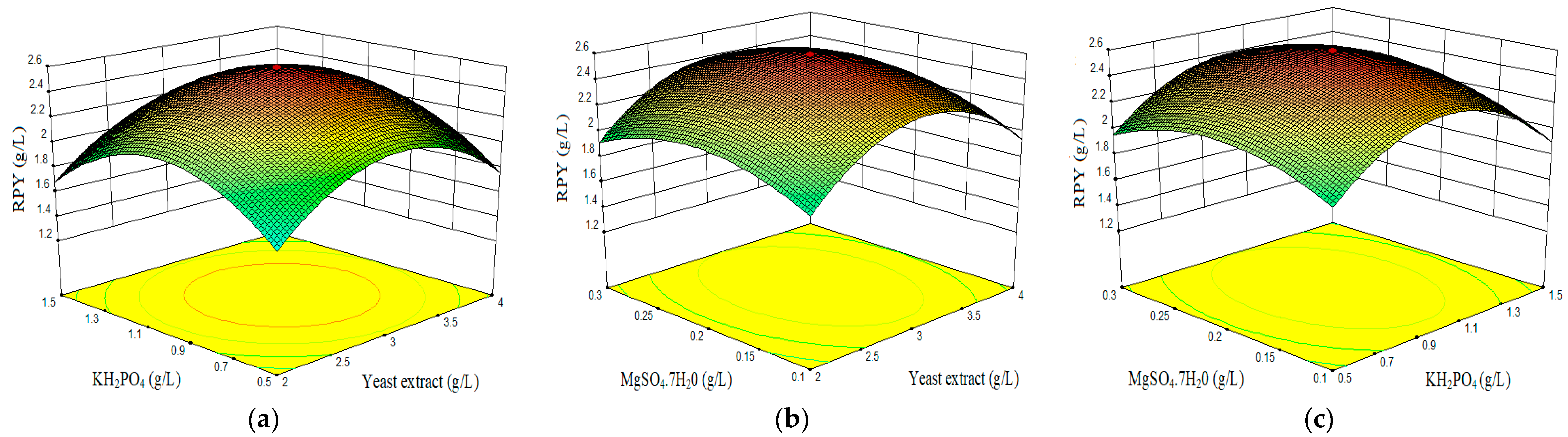
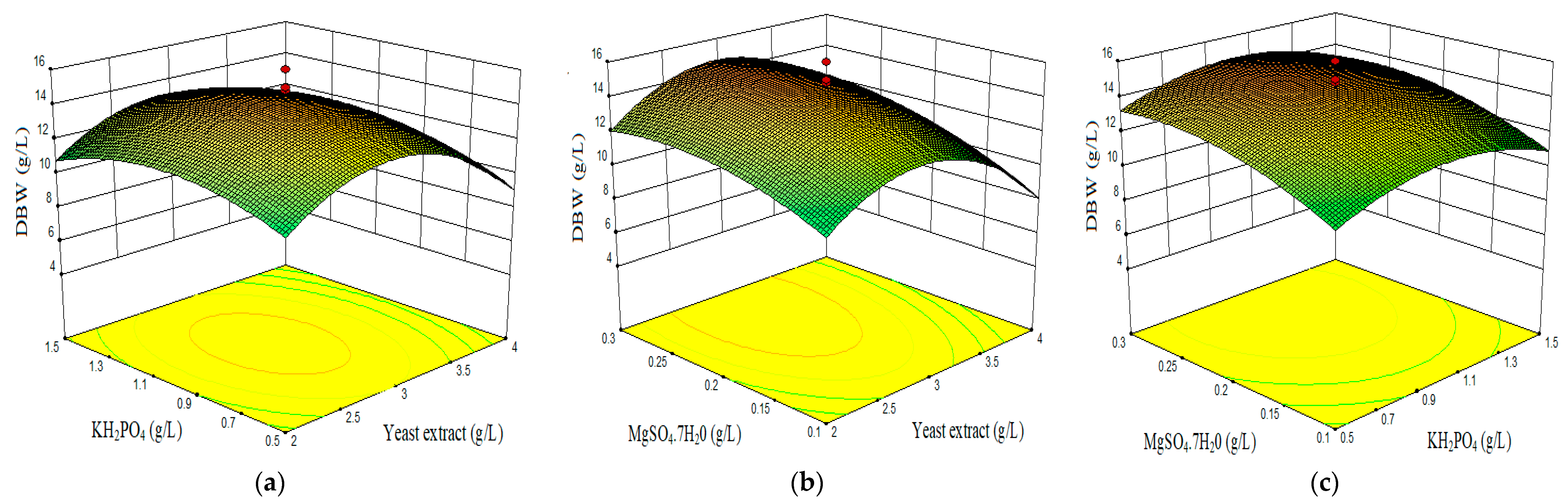
3.2. Optimization of Components Concentrations by Central Composite Design (CDD) and Response Surface Methodology (RSM)
- Development of second order polynomial models:
3.3. Analytical Validation
3.4. Effect of Process Variables and Responses (RSM)
- Combined effect of yeast extract, KH2PO4, and MgSO4·7H2O on orange pigment yield (OPY).
- Combined effect of yeast extract, KH2PO4, and MgSO4·7H2O on red pigment yield (RPY).
- Combined effect of yeast extract, KH2PO4, and MgSO4·7H2O on dry biomass weight (DBW).
3.5. Model Validation
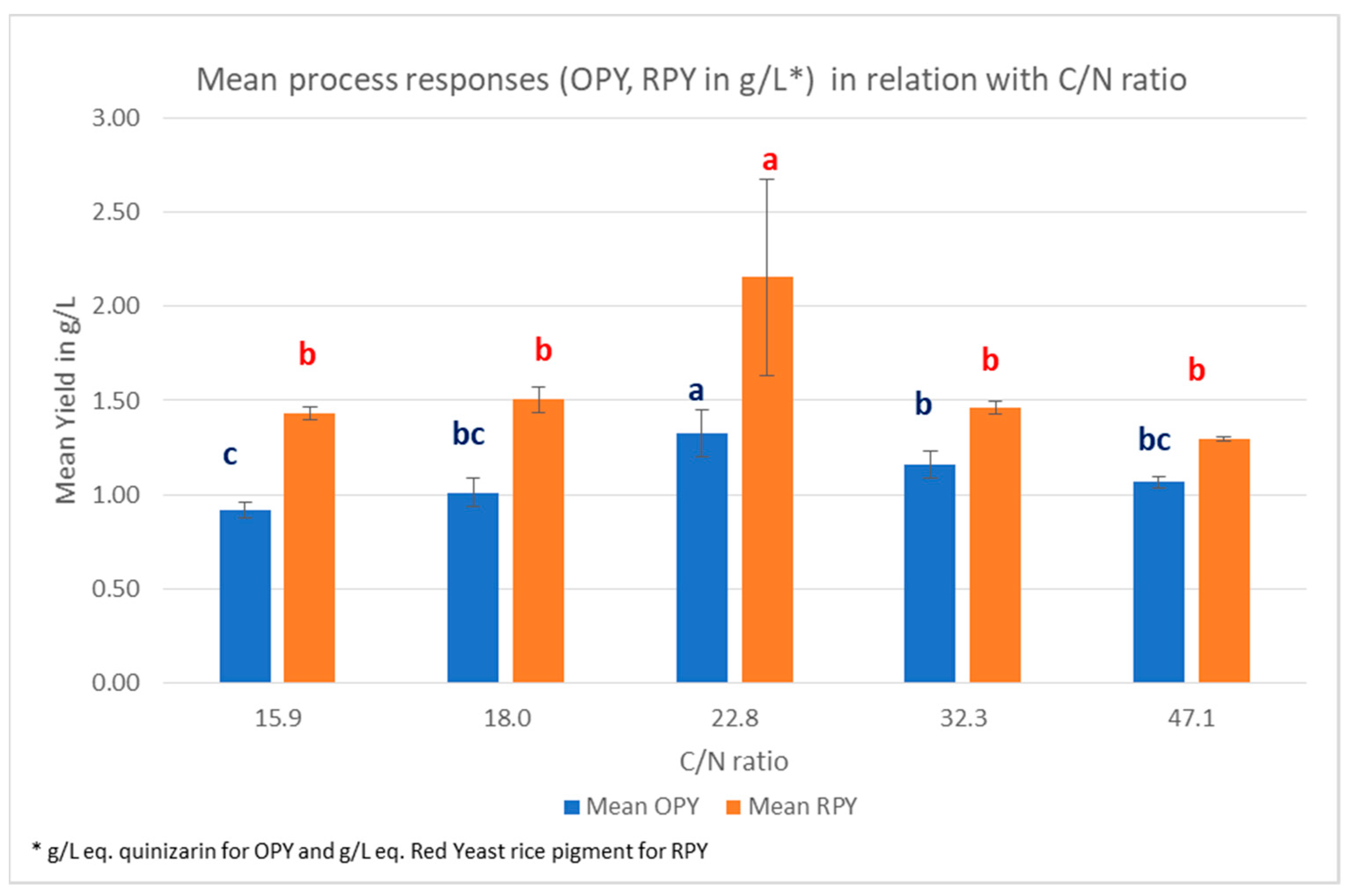
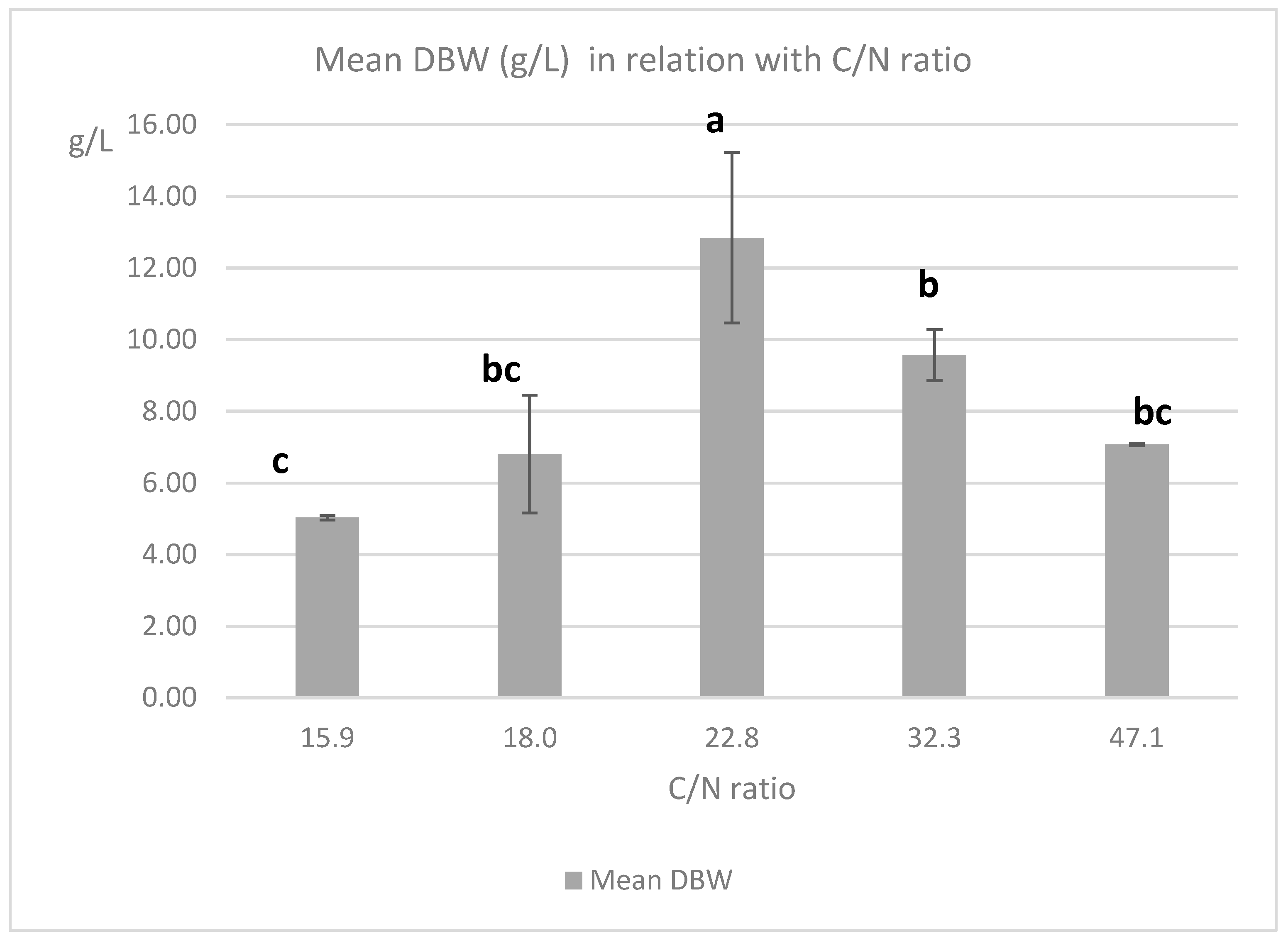
3.6. C/N Ration Influence in CCD Experiments
3.7. Effectiveness of OVAT and CDD on Process Optimization for Pigments and Biomass Production
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Joshi, V.K.; Attri, D.; Bala, A.; Bhushan, S. Microbial pigments. Indian J. Biotechnol. 2003, 2, 362–369. [Google Scholar]
- Tuli, H.S.; Chaudhary, P.; Beniwal, V.; Sharma, A.K. Microbial pigments as natural color sources: Current trends and future perspectives. J. Food Sci. Technol. 2015, 52, 4669–4678. [Google Scholar] [CrossRef] [Green Version]
- Downham, A.; Collins, P. Colouring our foods in the last and next millennium. Int. J. Food Sci. Technol. 2000, 35, 5–22. [Google Scholar] [CrossRef]
- Hallagan, J.B.; Allen, D.C.; Borzelleca, J.F. The safety and regulatory status of food, drug and cosmetics colour additives exempt from certification. Food Chem. Toxicol. 1995, 33, 515–528. [Google Scholar] [CrossRef]
- Wissgott, U.; Bortlik, K. Prospects for new natural food colorants. Trends Food Sci. Technol. 1996, 7, 298–302. [Google Scholar] [CrossRef]
- Potera, C. Diet and Nutrition: The Artificial Food Dye Blues. Environ. Health Perspect. 2010, 118, A428. [Google Scholar] [CrossRef] [Green Version]
- Dufossé, L. Microbial production of food grade pigments. Food Technol. Biotechnol. 2006, 44, 313–323. [Google Scholar]
- Malik, K.; Tokkas, J.; Goyal, S. Microbial pigments: A review. Int. J. Microb. Res. Technol. 2012, 1, 361–365. [Google Scholar]
- Jiang, Y.; Li, H.B.; Chen, F.; Hyde, K.D. Production potential of water-soluble Monascus red pigment by a newly isolated Penicillium sp. J. Agric. Technol. 2005, 1, 113–126. [Google Scholar]
- Dufossé, L.; Fouillaud, M.; Caro, Y.; Mapari, S.A.S.; Sutthiwong, N. Filamentous fungi are large-scale producers of pigments and colorants for the food industry. Curr. Opin. Biotechnol. 2014, 26, 56–61. [Google Scholar] [CrossRef] [PubMed]
- Mapari, S.A.; Thrane, U.; Meyer, A.S. Fungal polyketide azaphilone pigments as future natural food colorants? Trends Biotechnol. 2010, 28, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Pagano, M.C.; Dhar, P.P. Fungal pigments: An overview. In Fungal Biomolecules: Sources, Applications and Recent Developments; Gupta, V., Mach, R.L., Sreenivasaprasad, S., Eds.; John Wiley & Sons, Ltd.: Chichester, West Sussex, UK, 2015; pp. 173–181. [Google Scholar] [CrossRef]
- Calvo, A.M.; Wilson, R.A.; Bok, J.W.; Keller, N.P. Relationship between secondary metabolism and fungal development. Microbiol. Mol. Biol. Rev. 2002, 66, 447–459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gmoser, R.; Ferreira, J.A.; Lennartsson, P.R.; Taherzadeh, M.J. Filamentous ascomycetes fungi as a source of natural pigments. Fungal Biol. Biotechnol. 2017, 4, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morales-Oyervides, L.; Oliveira, J.C.; Sousa-Gallagher, M.J.; Méndez-Zavala, A.; Montañez, J.C. Quantitative assessment of the impact of the type of inoculum on the kinetics of cell growth, substrate consumption and pigment productivity by Penicillium purpurogenum GH2 in liquid culture with an integrated stochastic approach. Food Bioprod. Process. 2015, 96, 221–231. [Google Scholar] [CrossRef]
- Davies, J. Specialized microbial metabolites: Functions and origins. J. Antibiot. 2013, 66, 361–364. [Google Scholar] [CrossRef]
- Velíšek, J.; Cejpek, K. Pigments of higher fungi: A review. Czech. J. Food Sci. 2011, 29, 87–102. [Google Scholar] [CrossRef] [Green Version]
- Marmann, A.; Aly, A.H.; Lin, W.; Wang, B.; Proksch, P. Co-cultivation—A powerful emerging tool for enhancing the chemical diversity of microorganisms. Mar. Drugs 2014, 12, 1043–1065. [Google Scholar] [CrossRef] [Green Version]
- Schümann, J.; Hertweck, C. Advances in cloning, functional analysis and heterologous expression of fungal polyketide synthase genes. J. Biotechnol. 2006, 124, 690–703. [Google Scholar] [CrossRef]
- Chen, Y.; Feng, P.; Shang, Y.; Xu, Y.-J.; Wang, C. Biosynthesis of non-melanin pigment by a divergent polyketide synthase in Metarhizium robertsii. Fungal Genet. Biol. 2015, 81, 142–149. [Google Scholar] [CrossRef]
- Kurono, M.; Nakanishi, K.; Shindo, K.; Tada, M. Biosyntheses of Monascorubrin and Monascoflavin. Chem. Pharm. Bull. 1963, 11, 359–362. [Google Scholar] [CrossRef] [Green Version]
- Turner, W.B. Fungal Metabolites; Academic Press: London, UK; New York, NY, USA, 1971. [Google Scholar]
- Turner, W.; Aldridge, D. Fungal Metabolites, Volume II; Academic: Paris, France, 1983; pp. 257–259. [Google Scholar]
- Hajjaj, H.; Klaébé, A.; Loret, M.O.; Goma, G.; Blanc, P.J.; François, J. Biosynthetic pathway of citrinin in the filamentous fungus Monascus ruber as revealed by 13C nuclear magnetic resonance. Appl. Environ. Microbiol. 1999, 65, 311–314. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mapari, S.A.; Meyer, A.S.; Thrane, U.; Frisvad, J.C. Identification of potentially safe promising fungal cell factories for the production of polyketide natural food colorants using chemotaxonomic rationale. Microb. Cell Factories 2009, 8, 24. [Google Scholar] [CrossRef] [Green Version]
- Cho, Y.J.; Hwang, H.J.; Kim, S.W.; Song, C.H.; Yun, J.W. Effect of carbon source and aeration rate on broth rheology and fungal morphology during red pigment production by Paecilomyces sinclairii in a batch bioreactor. J. Biotechnol. 2002, 95, 13–23. [Google Scholar] [CrossRef]
- Duran, N.; Tixeira, M.F.S.; de Conti, R.; Esposito, E. Ecological-friendly pigments from fungi. Crit. Rev. Food Sci. Nutr. 2002, 42. [Google Scholar] [CrossRef]
- Engström, K.; Brishammar, S.; Svensson, C.; Bengtsson, M.; Andersson, R. Anthraquinones from some Drechslera species and Bipolaris sorokiniana. Mycol. Res. 1993, 97, 381–384. [Google Scholar] [CrossRef]
- Torres, F.A.E.; Zaccarim, B.R.; Lencastre Novaes, L.C.; Jozala, A.F.; Santos, C.A.; Teixeira, M.F.S.; Santos-Ebinuma, V.C. Natural colorants from filamentous fungi. Appl. Microbiol. Biotechnol. 2016, 100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiemann, P.; Willmann, A.; Straeten, M.; Kleigrewe, K.; Beyer, M.; Humpf, H.U.; Tudzynski, B. Biosynthesis of the red pigment bikaverin in Fusarium fujikuroi: Genes, their function and regulation. Mol. Microbiol. 2009, 72, 931–946. [Google Scholar] [CrossRef]
- Medentsev, A.G.; Arinbasarova, A.Y.; Akimenko, V.K. Biosynthesis of Naphthoquinone Pigments by Fungi of the Genus Fusarium. Appl. Biochem. Microbiol. 2005, 41, 503–507. [Google Scholar] [CrossRef]
- Lin, T.; Yakushijin, K.; Büchi, G.; Demain, A. Formation of water-solubleMonascus red pigments by biological and semi-synthetic processes. J. Ind. Microbiol. 1992, 9, 173–179. [Google Scholar] [CrossRef]
- Carvalho, J.C.; Pandey, A.; Babitha, S.; Soccol, C.R. Production of Monascus biopigments: An overview. Agro Food Ind. Hi Technol. 2003, 14, 37–43. [Google Scholar]
- Feng, Y.; Shao, Y.; Chen, F. Monascus pigments. Appl. Microbiol. Biotechnol. 2012, 96, 1421–1440. [Google Scholar] [CrossRef]
- Mukherjee, G.; Singh, S.K. Purification and characterization of a new red pigment from Monascus purpureus in submerged fermentation. Process. Biochem. 2011, 46. [Google Scholar] [CrossRef]
- Cho, Y.J.; Park, J.P.; Hwang, H.J.; Kim, S.W.; Choi, J.W.; Yun, J.W. Production of red pigment by submerged culture of Paecilomyces sinclairii. Lett. Appl. Microbiol. 2002, 35, 195–202. [Google Scholar] [CrossRef]
- Dong, Q.-L.; Guan, R.-J.; Xing, X.-Y.; Chen, J.-Y.; Yan, S.-Y. Optimization of pigment production from Paecilomyces lilacinus TD16 ant its antimicrobial activities. Mod. Chem. Ind. 2016, 8, 024. [Google Scholar]
- Méndez, A.; Pérez, C.; Montañéz, J.C.; Martínez, G.; Aguilar, C.N. Red pigment production by Penicillium purpurogenum GH2 is influenced by pH and temperature. J. Zhejiang Univ. Sci. B 2011, 12, 961–968. [Google Scholar] [CrossRef] [Green Version]
- Hailei, W.; Ping, L.; Yufeng, L.; Zhifang, R.; Gang, W. Overproduction of a potential red pigment by a specific self-immobilization biomembrane-surface liquid culture of Penicillium novae-zeelandiae. Bioprocess. Biosyst. Eng. 2012, 35. [Google Scholar] [CrossRef] [PubMed]
- Gunasekaran, S.; Poorniammal, R. Optimization of fermentation conditions for red pigment production from Penicillium sp. under submerged cultivation. Afr. J. Biotechnol. 2008, 7. [Google Scholar] [CrossRef] [Green Version]
- Li, X.L.; Cui, X.H.; Han, J.R. Sclerotial biomass and carotenoid yield of Penicillium sp. PT95 under oxidative growth conditions and in the presence of antioxidant ascorbic acid. J. Appl. Microbiol. 2006, 101. [Google Scholar] [CrossRef]
- Robinson, N.; Wood, K.; Hylands, P.J.; Gibson, T.M. Blue pigments of Penicillium herquei. J. Nat. Prod. 1992, 55. [Google Scholar] [CrossRef]
- Wang, P.-L.; Li, D.-Y.; Xie, L.-R.; Wu, X.; Hua, H.-M.; Li, Z.-L. Two new compounds from a marine-derived fungus Penicillium oxalicum. Nat. Prod. Res. 2014, 28, 290–293. [Google Scholar] [CrossRef]
- Frisvad, J.C.; Yilmaz, N.; Thrane, U.; Rasmussen, K.B.; Houbraken, J.; Samson, R.A. Talaromyces atroroseus, a New Species Efficiently Producing Industrially Relevant Red Pigments. PLoS ONE 2013, 8, e84102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yilmaz, N.; Houbraken, J.; Hoekstra, E.S.; Frisvad, J.C.; Visagie, C.M.; Samson, R.A. Delimitation and characterisation of Talaromyces purpurogenus and related species. Persoonia 2012, 29, 39–54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unagul, P.; Wongsa, P.; Kittakoop, P.; Intamas, S.; Srikitikulchai, P.; Tanticharoen, M. Production of red pigments by the insect pathogenic fungus Cordyceps unilateralis BCC 1869. J. Ind. Microbiol. Biotechnol. 2005, 32, 135–140. [Google Scholar] [CrossRef]
- Van Eijk, G.W.; Roeijmans, H.J. Averythrin-6-monomethyl ether, an anthraquinone pigment from Herpotrichia rhodosticta. Exp. Mycol. 1984, 8, 266–268. [Google Scholar] [CrossRef]
- Lin, C.-F. Isolation and cultural conditions of Monascus sp. for the production of pigment in a submerged culture. J. Ferment Technol. 1973, 51, 407–414. [Google Scholar]
- Lin, T.F.; Demain, A.L. Effect of nutrition of Monascus sp. on formation of red pigments. Appl. Microbiol. Biotechnol. 1991, 36, 70–75. [Google Scholar] [CrossRef]
- Carels, M.; Shepherd, D. The effect of changes in pH on phosphate and potassium uptake by Monascus rubiginosus ATCC 16367 in submerged shaken culture. Can. J. Microbiol. 1979, 25, 1484–1488. [Google Scholar] [CrossRef]
- Carels, M.; Shepherd, D. The effect of pH and amino acids on conidiation and pigment production of Monascus major ATCC 16362 and Monascus rubiginosus ATCC 16367 in submerged shaken culture. Can. J. Microbiol. 1978, 24, 1346–1357. [Google Scholar] [CrossRef]
- Wong, H.; Lin, Y.; Koehler, P. Regulation of growth and pigmentation of Monascus purpureus by carbon and nitrogen concentrations. Mycologia 1981, 73, 649–654. [Google Scholar] [CrossRef]
- Yoshimura, M.; Yamanaka, S.; Mitsugi, K.; Hirose, Y. Production of Monascus-pigment in a submerged culture. Agric. Biol. Chem. 1975, 39, 1789–1795. [Google Scholar] [CrossRef]
- Broder, C.; Koehler, P. Pigments produced by Monascus purpureus with regard to quality and quantity. J. Food Sci. 1980, 45, 567–569. [Google Scholar] [CrossRef]
- Su, Y.C. Fermentative production of anka-pigments (Monascus-pigments). Microbiol. Biotechnol. Lett. 1983, 11, 325. [Google Scholar]
- Souza, P.N.; Grigoletto, T.L.; de Moraes, L.A.; Abreu, L.M.; Guimaraes, L.H.; Santos, C.; Galvao, L.R.; Cardoso, P.G. Production and chemical characterization of pigments in filamentous fungi. Microbiology 2016, 162, 12–22. [Google Scholar] [CrossRef]
- Kalim, B.; Ali, N.M. Optimization of fermentation media and growth conditions for microbial xylanase production. 3 Biotech 2016, 6, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finkelstein, D.B. Biotechnology of Filamentous Fungi: Technology and Products; Elsevier Science: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Gonzalez, R.; Islas, L.; Obregon, A.-M.; Escalante, L.; Sanchez, S. Gentamicin formation in Micromonospora purpurea: Stimulatory effect of ammonium. J. Antibiot. 1995, 48, 479–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tasharrofi, N.; Adrangi, S.; Fazeli, M.; Rastegar, H.; Khoshayand, M.R.; Faramarzi, M.A. Optimization of chitinase production by Bacillus pumilus using Plackett-Burman design and response surface methodology. Iran. J. Pharm. Res. 2011, 10, 759. [Google Scholar] [PubMed]
- Joshi, S.; Yadav, S.; Desai, A.J. Application of response-surface methodology to evaluate the optimum medium components for the enhanced production of lichenysin by Bacillus licheniformis R2. Biochem. Eng. J. 2008, 41, 122–127. [Google Scholar] [CrossRef]
- Chen, D.; Xue, Y.; Chen, M.; Li, Z.; Wang, C. Optimization of submerged fermentation medium for citrinin-free monascin production by Monascus. Prep. Biochem. Biotechnol. 2016, 46, 772–779. [Google Scholar] [CrossRef]
- Ahmad, M.; Panda, B.P. Optimization of red pigment production by Monascus purpureus MTCC 369 under solid-state fermentation using response surface methodology. Songlanakarin J. Sci. Technol. 2014, 36, 439–444. [Google Scholar]
- Baneshi, F.; Azizi, M.; Saberi, M.; Farsi, M. Evaluation of pH, carbon source and temperature effect on the pigments production by Monascus purpureus in a liquid culture using response surface methodology. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 905–911. [Google Scholar]
- Silveira, S.T.; Daroit, D.J.; Brandelli, A. Pigment production by Monascus purpureus in grape waste using factorial design. Lwt-Food Sci. Technol. 2008, 41, 170–174. [Google Scholar] [CrossRef]
- Zhou, B.; Wang, J.; Pu, Y.; Zhu, M.; Liu, S.; Liang, S. Optimization of culture medium for yellow pigments production with Monascus anka mutant using response surface methodology. Eur. Food Res. Technol. 2009, 228, 895–901. [Google Scholar] [CrossRef]
- Venkatachalam, M.; Magalon, H.; Dufossé, L.; Fouillaud, M. Production of pigments from the tropical marine-derived fungi Talaromyces albobiverticillius: New resources for natural red-colored metabolites. J. Food Compos. Anal. 2018, 70, 35–48. [Google Scholar] [CrossRef]
- Venkatachalam, M.; Shum-Chéong-Sing, A.; Dufossé, L.; Fouillaud, M. Statistical optimization of the physico-chemical parameters for pigment production in submerged fermentation of Talaromyces albobiverticillius 30548. Microorganisms 2020, 8, 711. [Google Scholar] [CrossRef]
- Hernández-Rivera, J.; Méndez-Zavala, A.; Pérez-Berúmen, C.; Contreras-Esquivel, J.; Rodríguez-Herrera, R.; Aguilar, C. Culture conditions to improve the red pigment production by Penicillium purpurogenum GH2. In Advance in Food Science and Food Biotechnology in Developing Countries; Mexican Association for Food Science: Saltillo, Mexico, 2008; pp. 108–112. [Google Scholar]
- Chen, G.; Shi, K.; Song, D.; Quan, L.; Wu, Z. The pigment characteristics and productivity shifting in high cell density culture of Monascus anka mycelia. BMC Biotechnol. 2015, 15, 72. [Google Scholar] [CrossRef] [Green Version]
- Venkatachalam, M.; Zelena, M.; Cacciola, F.; Ceslova, L.; Girard-Valenciennes, E.; Clerc, P.; Dugo, P.; Mondello, L.; Fouillaud, M.; Rotondo, A.; et al. Partial characterization of the pigments produced by the marine-derived fungus Talaromyces albobiverticillius 30548. Towards a new fungal red colorant for the food industry. J. Food Comp. Anal. 2018, 67, 38–47. [Google Scholar] [CrossRef]
- Santos-Ebinuma, V.C.; Roberto, I.C.; Teixeira, M.F.S.; Pessoa, A., Jr. Improvement of submerged culture conditions to produce colorants by Penicillium purpurogenum. Braz. J. Microbiol. 2014, 45, 731–742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Velmurugan, P.; Lee, Y.H.; Venil, C.K.; Lakshmanaperumalsamy, P.; Chae, J.-C.; Oh, B.-T. Effect of light on growth, intracellular and extracellular pigment production by five pigment-producing filamentous fungi in synthetic medium. J. Biosci. Bioeng. 2010, 109, 346–350. [Google Scholar] [CrossRef] [PubMed]
- Bezerra, M.A.; Santelli, R.E.; Oliveira, E.P.; Villar, L.S.; Escaleira, L.A. Response surface methodology (RSM) as a tool for optimization in analytical chemistry. Talanta 2008, 76, 965–977. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, S.L.C.; Bruns, R.E.; Ferreira, H.S.; Matos, G.D.; David, J.M.; Brandão, G.C.; da Silva, E.G.P.; Portugal, L.A.; dos Reis, P.S.; Souza, A.S.; et al. Box-Behnken design: An alternative for the optimization of analytical methods. Anal. Chim. Acta 2007, 597, 179–186. [Google Scholar] [CrossRef]
- Thompson, K.A.; Summers, R.; Cook, S.M. Development and experimental validation of the composition and treatability of a new synthetic bathroom greywater (SynGrey). Environ. Sci. Water Res. Technol. 2017, 3, 1120–1131. [Google Scholar] [CrossRef]
- Prakash Maran, J.; Mekala, V.; Manikandan, S. Modeling and optimization of ultrasound-assisted extraction of polysaccharide from Cucurbita moschata. Carbohydr. Polym. 2013, 92, 2018–2026. [Google Scholar] [CrossRef]
- Balasubramani, P.; Palaniswamy, P.; Visvanathan, R.; Thirupathi, V.; Subbarayan, A.; Maran, J.P. Microencapsulation of garlic oleoresin using maltodextrin as wall material by spray drying technology. Int. J. Biol. Macromol. 2015, 72, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Prakash Maran, J.; Priya, B. Ultrasound-assisted extraction of polysaccharide from Nephelium lappaceum L. fruit peel. Int. J. Biol. Macromol. 2014, 70, 530–536. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Singh, S.K.; Tripathi, V.R.; Khare, S.K.; Garg, S.K. Comparative one-factor-at-a-time, response surface (statistical) and bench-scale bioreactor level optimization of thermoalkaline protease production from a psychrotrophic Pseudomonas putida SKG-1 isolate. Microb. Cell Factories 2011, 10, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, L.; Kwong, T.F.; Qian, P.-Y. Effect of culture conditions on mycelial growth, antibacterial activity, and metabolite profiles of the marine-derived fungus Arthrinium cf saccharicola. Appl. Microbiol. Biotechnol. 2006, 72, 1063–1073. [Google Scholar] [CrossRef]
- Bode, H.B.; Bethe, B.; Höfs, R.; Zeeck, A. Big effects from small changes: Possible ways to explore nature’s chemical diversity. ChemBioChem 2002, 3, 619–627. [Google Scholar] [CrossRef]
- Mapari, S.A.; Nielsen, K.F.; Larsen, T.O.; Frisvad, J.C.; Meyer, A.S.; Thrane, U. Exploring fungal biodiversity for the production of water-soluble pigments as potential natural food colorants. Curr. Opin. Biotechnol. 2005, 16, 231–238. [Google Scholar] [CrossRef]
- Çelik, E.; Çalık, P. Bioprocess Parameters and Oxygen Transfer Characteristics in β-Lactamase Production by Bacillus Species. Biotechnol. Prog. 2004, 20, 491–499. [Google Scholar] [CrossRef]
- Chadni, Z.; Rahaman, M.H.; Jerin, I.; Hoque, K.M.F.; Reza, M.A. Extraction and optimisation of red pigment production as secondary metabolites from Talaromyces verruculosus and its potential use in textile industries. Mycology 2017, 8, 48–57. [Google Scholar] [CrossRef] [Green Version]
- Lagashetti, A.C.; Dufossé, L.; Singh, S.K.; Singh, P.N. Fungal Pigments and Their Prospects in Different Industries. Microorganisms 2019, 7, 604. [Google Scholar] [CrossRef] [Green Version]
- Tseng, Y.; Chen, M.; Lin, C. Growth, pigment production and protease activity of Monascus purpureus as affected by salt, sodium nitrite, polyphosphate and various sugars. J. Appl. Microbiol. 2000, 88, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Sharmila, G.; Nidhi, B.; Muthukumaran, C. Sequential statistical optimization of red pigment production by Monascus purpureus (MTCC 369) using potato powder. Ind. Crop. Prod. 2013, 44, 158–164. [Google Scholar] [CrossRef]
- Silveira, S.T.; Daroit, D.J.; Sant’Anna, V.; Brandelli, A. Stability modeling of red pigments produced by Monascus purpureus in submerged cultivations with sugarcane bagasse. Food Bioprocess. Technol. 2013, 6, 1007–1014. [Google Scholar] [CrossRef]
- Zhou, Z.; Yin, Z.; Hu, X. Corncob hydrolysate, an efficient substrate for Monascus pigment production through submerged fermentation. Biotechnol. Appl. Biochem. 2014, 61, 716–723. [Google Scholar] [CrossRef]
- Babitha, S.; Soccol, C.R.; Pandey, A. Jackfruit seed-a novel substrate for the production of Monascus pigments through solid-state fermentation. Food Technol. Biotechnol. 2006, 44, 465–471. [Google Scholar]
- Kongruang, S. Growth kinetics of biopigment production by Thai isolated Monascus purpureus in a stirred tank bioreactor. J. Ind. Microbiol. Biotechnol. 2011, 38, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.-H.; Johns, M.R. Effect of carbon source on ethanol and pigment production by Monascus purpureus. Enzym. Microb. Technol. 1994, 16, 584–590. [Google Scholar] [CrossRef]
- Dey, D.; Banerjee, D.; Halder, S.; Pati, B. Optimization of Red Pigment Production by Endophytic Pezicula sp. BDF9/1 through OVAT and RSM Methodology. Int. J. Curr. Res. 2016, 8, 28194–28201. [Google Scholar]
- Kim, C.; Kim, S.; Hong, S. Production of red pigment by Serratia sp. KH-95 and its cultural properties. Korean J. Biotechnol. Bioeng. 1998, 13, 431–437. [Google Scholar]
- Stange, S.; Steudler, S.; Delenk, H.; Werner, A.; Walther, T.; Wagenführ, A. Influence of the nutrients on the biomass and pigment production of Chlorociboria aeruginascens. J. Fungi 2019, 5, 40. [Google Scholar] [CrossRef] [Green Version]
- Tudzynski, B. Nitrogen regulation of fungal secondary metabolism in fungi. Front. Microbiol. 2014, 5, 656. [Google Scholar] [CrossRef] [Green Version]
- Manan, M.; Mohamad, R.; Ariff, A. Monascus spp. a source of natural microbial color through fungal biofermentation. J. Microbiol. Exp. 2017, 5, 00148. [Google Scholar] [CrossRef] [Green Version]
- Arai, T.; Umemura, S.; Ota, T.; Ogihara, J.; Kato, J.; Kasumi, T. Effects of inorganic nitrogen sources on the production of pp-v [(10 z)-12-carboxyl-monascorubramine] and the expression of the nitrate assimilation gene cluster by Penicillium sp. AZ. Biosci. Biotechnol. Biochem. 2012, 76, 120–124. [Google Scholar] [CrossRef] [Green Version]
- Patrovsky, M.; Sinovska, K.; Branska, B.; Patakova, P. Effect of initial pH, different nitrogen sources, and cultivation time on the production of yellow or orange Monascus purpureus pigments and the mycotoxin citrinin. Food Sci. Nutr. 2019, 7, 3494–3500. [Google Scholar] [CrossRef] [Green Version]
- Poorniammal, R.; Gunasekaran, S.; Murugesan, R. Statistical optimization of culture medium for yellow pigment production by Thermomyces sp. J. Appl. Nat. Sci. 2015, 7, 203–210. [Google Scholar] [CrossRef] [Green Version]
- Yongsmith, B.; Krairak, S.; Chaisrisook, C. Fermentation of yellow pigments by cassava starch utilizing Monascus spp. Biotechnol Sustain. Util. Biol. Resour. Trop. 1998, 12, 235–244. [Google Scholar]
- Lakrod, K.; Chaisrisook, C.; Yongsmith, B.; Skinner, D. RAPD analysis of genetic variation within a collection of Monascus spp. isolated from red rice (ang-kak) and sofu. Mycol. Res. 2000, 104, 403–408. [Google Scholar] [CrossRef]
- Mostafa, M.E.; Abbady, M.S. Secondary metabolites and bioactivity of the Monascus pigments review article. Glob. J. Biotechnol. Biochem. 2014, 9, 1–13. [Google Scholar]
- Panagou, E.Z.; Katsaboxakis, C.Z.; Nychas, G.-J.E. Heat resistance of Monascus ruber ascospores isolated from thermally processed green olives of the Conservolea variety. Int. J. Food Microbiol. 2002, 76, 11–18. [Google Scholar] [CrossRef]
- Morales-Oyervides, L.; Ruiz-Sánchez, J.P.; Oliveira, J.C.; Sousa-Gallagher, M.J.; Morales-Martínez, T.K.; Albergamo, A.; Salvo, A.; Giuffrida, D.; Dufossé, L.; Montañez, J. Medium design from corncob hydrolyzate for pigment production by Talaromyces atroroseus GH2: Kinetics modeling and pigments characterization. Biochem. Eng. J. 2020, 161, 107698. [Google Scholar] [CrossRef]
- Jackson, M.A. Optimizing nutritional conditions for the liquid culture production of effective fungal biological control agents. J. Ind. Microbiol. Biotechnol. 1997, 19, 180–187. [Google Scholar] [CrossRef]
- Jackson, M.A.; Bothast, R.J. Carbon concentration and carbon-to-nitrogen ratio influence submerged-culture conidiation by the potential bioherbicide Colletotrichum truncatum NRRL 13737. Appl. Environ. Microbiol. 1990, 56, 3435–3438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gao, L.; Sun, M.H.; Liu, X.Z.; Che, Y.S. Effects of carbon concentration and carbon to nitrogen ratio on the growth and sporulation of several biocontrol fungi. Mycol. Res. 2007, 111, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Hajjaj, H.; Blanc, P.; Groussac, E.; Goma, G.; Uribelarrea, J.; Loubiere, P. Improvement of red pigment/citrinin production ratio as a function of environmental conditions by Monascus ruber. Biotechnol. Bioeng. 1999, 64, 497–501. [Google Scholar] [CrossRef]
- Chiu, S.-W.; Poon, Y.-K. Submerged production of Monascus pigments. Mycologia 1993, 85, 214–218. [Google Scholar] [CrossRef]
- Hajjaj, H.; Blanc, P.; Goma, G.; Francois, J. Sampling techniques and comparative extraction procedures for quantitative determination of intra-and extracellular metabolites in filamentous fungi. FEMS Microbiol. Lett. 1998, 164, 195–200. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variables | Symbol | Coded and Actual Values (in g/L) | ||||
|---|---|---|---|---|---|---|
| −α * | −1 | 0 | +1 | +α | ||
| Yeast extract | X1 | 1.32 | 2 | 3 | 4 | 4.68 |
| KH2PO4 | X2 | 0.16 | 0.5 | 1 | 1.5 | 1.84 |
| MgSO4·7H2O | X3 | 0.03 | 0.1 | 0.2 | 0.3 | 0.37 |
| Std Order | Run Order | Variables | (g/L) | Responses | |||
|---|---|---|---|---|---|---|---|
| X1 Yeast Extract | X2 KH2PO4 | X3 MgSO4·7H2O | OPY (g/L 1) | RPY (g/L 2) | DBW 3 (g/L) | ||
| 12 | 1 | 3 | 1.84 | 0.2 | 1.13 | 1.37 | 10.95 |
| 2 | 2 | 4 | 0.5 | 0.1 | 1.01 | 1.52 | 6.49 |
| 3 | 3 | 2 | 1.5 | 0.1 | 1.09 | 1.43 | 9.03 |
| 9 | 4 | 1.31 | 1 | 0.2 | 1.04 | 1.28 | 7.05 |
| 5 | 5 | 2 | 0.5 | 0.3 | 1.26 | 1.47 | 10.08 |
| 20 | 6 | 3 | 1 | 0.2 | 1.42 | 2.59 | 14.23 |
| 13 | 7 | 3 | 1 | 0.03 | 1.17 | 1.91 | 9.12 |
| 4 | 8 | 4 | 1.5 | 0.1 | 1.04 | 1.52 | 6.81 |
| 17 | 9 | 3 | 1 | 0.2 | 1.43 | 2.57 | 13.63 |
| 11 | 10 | 3 | 0.16 | 0.2 | 1.24 | 1.37 | 10.57 |
| 18 | 11 | 3 | 1 | 0.2 | 1.42 | 2.58 | 13.12 |
| 19 | 12 | 3 | 1 | 0.2 | 1.43 | 2.57 | 14.98 |
| 10 | 13 | 4.68 | 1 | 0.2 | 0.91 | 1.43 | 5.03 |
| 1 | 14 | 2 | 0.5 | 0.1 | 1.13 | 1.44 | 8.89 |
| 8 | 15 | 4 | 1.5 | 0.3 | 1.09 | 1.57 | 8.95 |
| 7 | 16 | 2 | 1.5 | 0.3 | 1.17 | 1.51 | 10.27 |
| 15 | 17 | 3 | 1 | 0.2 | 1.42 | 2.57 | 15.98 |
| 16 | 18 | 3 | 1 | 0.2 | 1.41 | 2.58 | 14.77 |
| 6 | 19 | 3 | 1 | 0.2 | 1.16 | 1.57 | 9.03 |
| 14 | 20 | 3 | 1 | 0.37 | 1.34 | 2.03 | 14.89 |
| Source | DF | Orange Pigment Yield OPY (g/L *) | |||
|---|---|---|---|---|---|
| Sum of Squares | Mean Square | F-Value | p-Value | ||
| Model | 9 | 0.52 | 0.058 | 51.09 | <0.0001 |
| A-Yeast extract | 1 | 0.024 | 0.024 | 208.28 | 0.0001 |
| B-KH2PO4 | 1 | 9.228 × 10−3 | 9.228 × 10−3 | 81.18 | 0.0001 |
| C-MgSO4·7H2O | 1 | 0.035 | 0.035 | 311.95 | 0.0001 |
| AB | 1 | 1.013 × 10−3 | 1.013 × 10−3 | 8.91 | 0.0137 |
| AC | 1 | 1.250 × 10−5 | 1.250 × 10−5 | 0.11 | 0.7470 |
| BC | 1 | 2.812 × 10−3 | 2.812 × 10−3 | 24.74 | 0.0006 |
| A2 | 1 | 0.36 | 0.36 | 3181.99 | <0.0001 |
| B2 | 1 | 0.10 | 0.10 | 898.39 | <0.0001 |
| C2 | 1 | 0.051 | 0.051 | 447.79 | <0.0001 |
| Residual | 10 | 1.137 × 10−3 | 1.137 × 10−4 | ||
| Lack of Fit | 5 | 8.534 × 10−4 | 1.707 × 10−4 | 3.01 | 0.1258 |
| Pure Error | 5 | 2.833 × 10−4 | 5.667 × 10−5 | ||
| Cor. Total | 19 | 0.52 | |||
| SD | 0.001 | ||||
| Mean | 1.22 | ||||
| CV% | 0.88 | ||||
| R2 | 0.9978 | ||||
| R2adj. | 0.9959 | ||||
| R2pred. | 0.9859 | ||||
| Adeq. Pre. | 68.726 | ||||
| Source | DF | Red Pigment Yield RPY (g/L *) | |||
|---|---|---|---|---|---|
| Sum of Squares | Mean Square | F-Value | p-Value | ||
| Model | 9 | 5.14 | 0.57 | 55.81 | <0.0001 |
| A-Yeast extract | 1 | 0.025 | 0.025 | 242.44 | <0.0001 |
| B-KH2PO4 | 1 | 6.590 × 10−5 | 6.590 × 10−5 | 0.64 | 0.04411 |
| C-MgSO4·7H2O | 1 | 0.012 | 0.012 | 121.27 | <0.0001 |
| AB | 1 | 1.125 × 10−4 | 1.125 × 10−4 | 1.10 | 0.3192 |
| AC | 1 | 1.250 × 10−5 | 1.250 × 10−5 | 0.12 | 0.7340 |
| BC | 1 | 3.125 × 10−4 | 3.125 × 10−4 | 3.05 | 0.1112 |
| A2 | 1 | 2.69 | 2.69 | 26252.50 | <0.0001 |
| B2 | 1 | 2.62 | 2.62 | 25611.75 | <0.0001 |
| C2 | 1 | 0.66 | 0.66 | 6473.14 | <0.0001 |
| Residual | 10 | 1.024 × 10−3 | 1.024 × 10−4 | ||
| Lack of Fit | 5 | 6.907 × 10−4 | 1.381 × 10−4 | 2.07 | 0.2215 |
| Pure Error | 5 | 3.333 × 10−4 | 6.667 × 10−5 | ||
| Cor. Total | 19 | 5.15 | |||
| SD | 0.001 | ||||
| Mean | 1.84 | ||||
| CV% | 0.55 | ||||
| R2 | 0.9998 | ||||
| R2adj. | 0.9996 | ||||
| R2pred. | 0.9988 | ||||
| Adeq. Pre. | 180.745 | ||||
| Source | DF | Dry Biomass Concentration (DBW g/L) | |||
|---|---|---|---|---|---|
| Sum of Squares | Mean Square | F-Value | p-Value | ||
| Model | 9 | 191.65 | 21.29 | 23.77 | <0.0001 |
| A-Yeast extract | 1 | 7.90 | 7.90 | 8.82 | 0.0141 |
| B-KH2PO4 | 1 | 0.11 | 0.11 | 0.12 | 0.7368 |
| C-MgSO4·7H2O | 1 | 20.70 | 20.70 | 23.10 | 0.0007 |
| AB | 1 | 1.013 × 10−3 | 1.013 × 10−3 | 1.130 × 10−3 | 0.9738 |
| AC | 1 | 0.63 | 0.63 | 0.71 | 0.4203 |
| BC | 1 | 0.015 | 0.015 | 0.017 | 0.8986 |
| A2 | 1 | 138.80 | 138.80 | 154.92 | <0.0001 |
| B2 | 1 | 29.66 | 29.66 | 33.11 | 0.0002 |
| C2 | 1 | 14.25 | 14.25 | 15.91 | 0.0026 |
| Residual | 10 | 8.96 | 0.90 | ||
| Lack of Fit | 5 | 3.75 | 0.75 | 0.72 | 0.6372 |
| Pure Error | 5 | 5.21 | 1.04 | ||
| Cor. Total | 19 | 200.61 | |||
| SD | 0.95 | ||||
| Mean | 10.69 | ||||
| CV% | 8.85 | ||||
| R2 | 0.9553 | ||||
| R2adj. | 0.9151 | ||||
| R2pred. | 0.8209 | ||||
| Adeq. Pre. | 15.026 | ||||
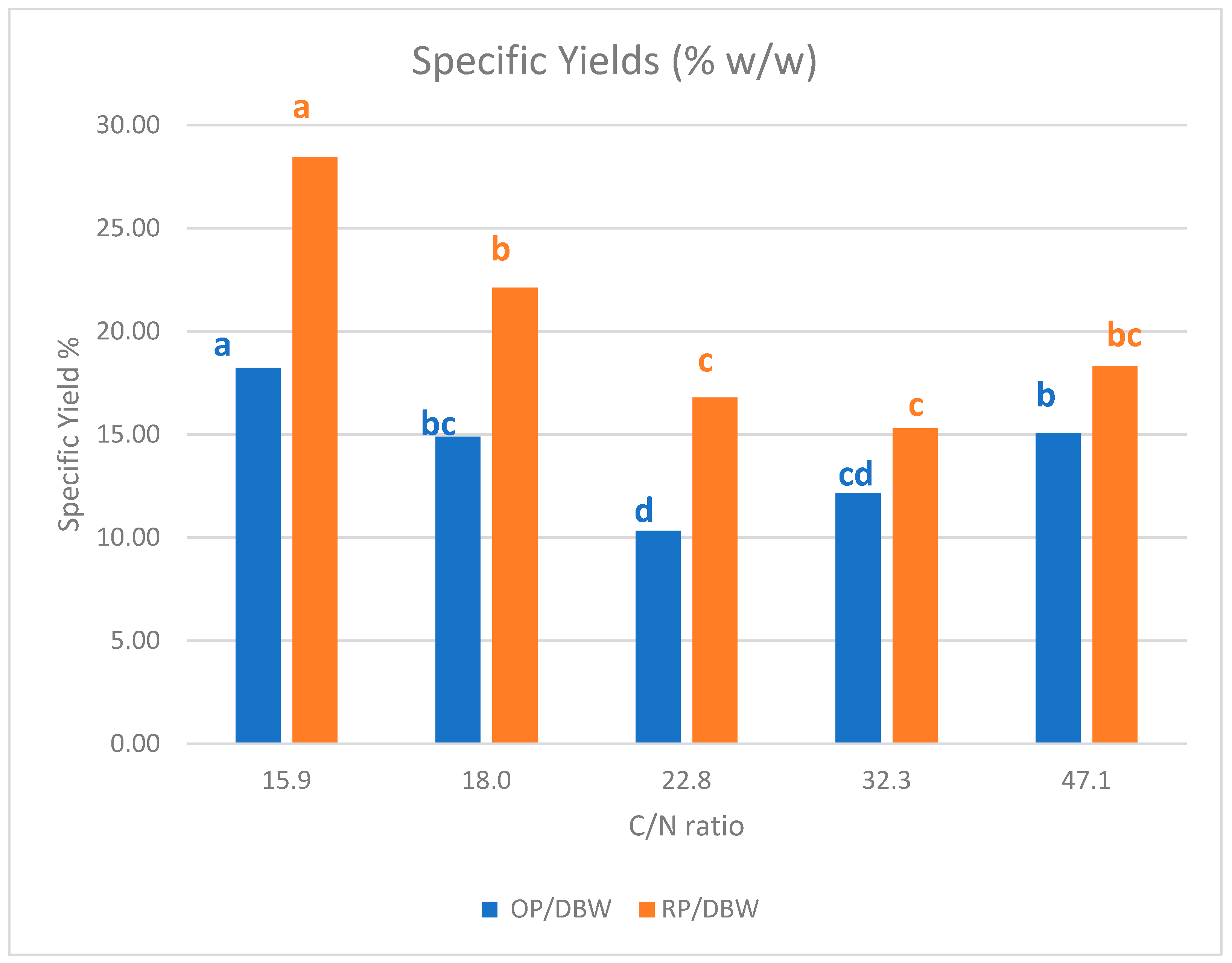
| C/N | n | Mean OPY 1 g/L | SD | Mean RPY 2 g/L | SD | Mean DBW 3 g/L | SD |
|---|---|---|---|---|---|---|---|
| 15.9 | 3 | 0.92 | 0.04 | 1.43 | 0.03 | 5.03 | 0.06 |
| 18 | 12 | 1.01 | 0.08 | 1.51 | 0.07 | 6.81 | 1.64 |
| 22.8 | 33 | 1.32 | 0.12 | 2.16 | 0.52 | 12.84 | 2.38 |
| 32.3 | 12 | 1.16 | 0.07 | 1.46 | 0.04 | 9.57 | 0.71 |
| 47.1 | 3 | 1.07 | 0.03 | 1.30 | 0.01 | 7.08 | 0.03 |
| C and N Sources | Min. Values g/L | Max. Values g/L | Incremental Factors | ||||||
| OPY | RPY | DBW | OPY | RPY | DBW | OPY | RPY | DBW | |
| 2 Factors Combination | |||||||||
| ME */Peptone | Sucrose/NaNO3 | ME */NaNO3 | Sucrose/YE ** | Sucrose/YE ** | Sucrose/YE ** | ||||
| OVAT optimization | 0.17 | 0.26 | 0.92 | 1.39 | 2.44 | 6.60 | ×8.2 | ×9.4 | ×7.2 |
| 4 Factors Concentrations(Sucrose/YE **/KH2PO4/MgSO4·7H2O) | |||||||||
| CCD optimization | 0.91 | 1.28 | 5.03 | 1.43 | 2.,59 | 15.98 | ×1.6 | ×2 | ×3.2 |
| Overall increase | ×8.4 | ×10 | ×17.4 | ||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Venkatachalam, M.; Shum-Chéong-Sing, A.; Caro, Y.; Dufossé, L.; Fouillaud, M. OVAT Analysis and Response Surface Methodology Based on Nutrient Sources for Optimization of Pigment Production in the Marine-Derived Fungus Talaromyces albobiverticillius 30548 Submerged Fermentation. Mar. Drugs 2021, 19, 248. https://doi.org/10.3390/md19050248
Venkatachalam M, Shum-Chéong-Sing A, Caro Y, Dufossé L, Fouillaud M. OVAT Analysis and Response Surface Methodology Based on Nutrient Sources for Optimization of Pigment Production in the Marine-Derived Fungus Talaromyces albobiverticillius 30548 Submerged Fermentation. Marine Drugs. 2021; 19(5):248. https://doi.org/10.3390/md19050248
Chicago/Turabian StyleVenkatachalam, Mekala, Alain Shum-Chéong-Sing, Yanis Caro, Laurent Dufossé, and Mireille Fouillaud. 2021. "OVAT Analysis and Response Surface Methodology Based on Nutrient Sources for Optimization of Pigment Production in the Marine-Derived Fungus Talaromyces albobiverticillius 30548 Submerged Fermentation" Marine Drugs 19, no. 5: 248. https://doi.org/10.3390/md19050248
APA StyleVenkatachalam, M., Shum-Chéong-Sing, A., Caro, Y., Dufossé, L., & Fouillaud, M. (2021). OVAT Analysis and Response Surface Methodology Based on Nutrient Sources for Optimization of Pigment Production in the Marine-Derived Fungus Talaromyces albobiverticillius 30548 Submerged Fermentation. Marine Drugs, 19(5), 248. https://doi.org/10.3390/md19050248