
Metabolomic Characterization of a cf. Neolyngbya Cyanobacterium from the South China Sea Reveals Wenchangamide A, a Lipopeptide with In Vitro Apoptotic Potential in Colon Cancer Cells

 , , , , ,
, , , , , 
 and
and
Abstract
:1. Introduction
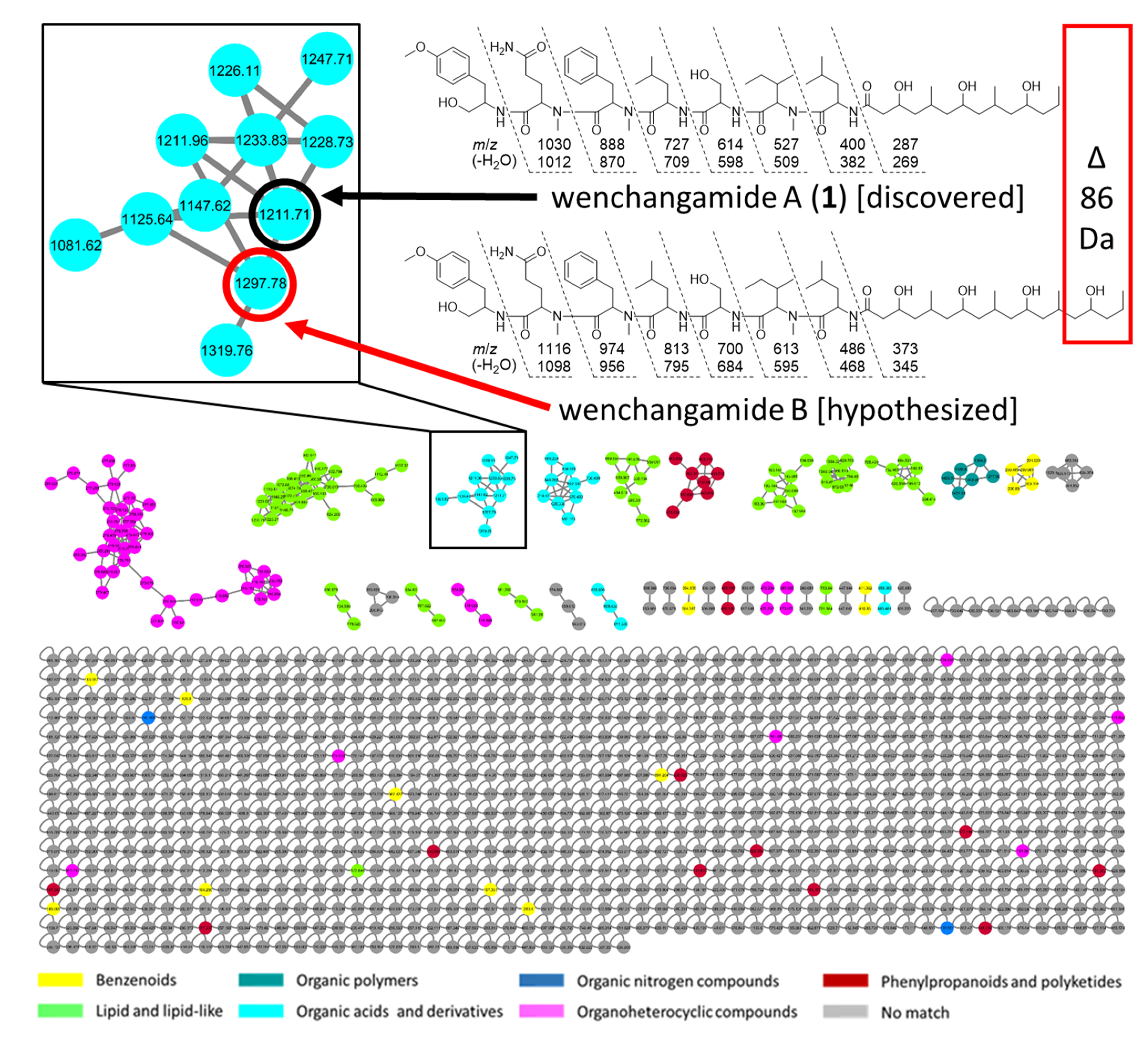
2. Results and Discussion
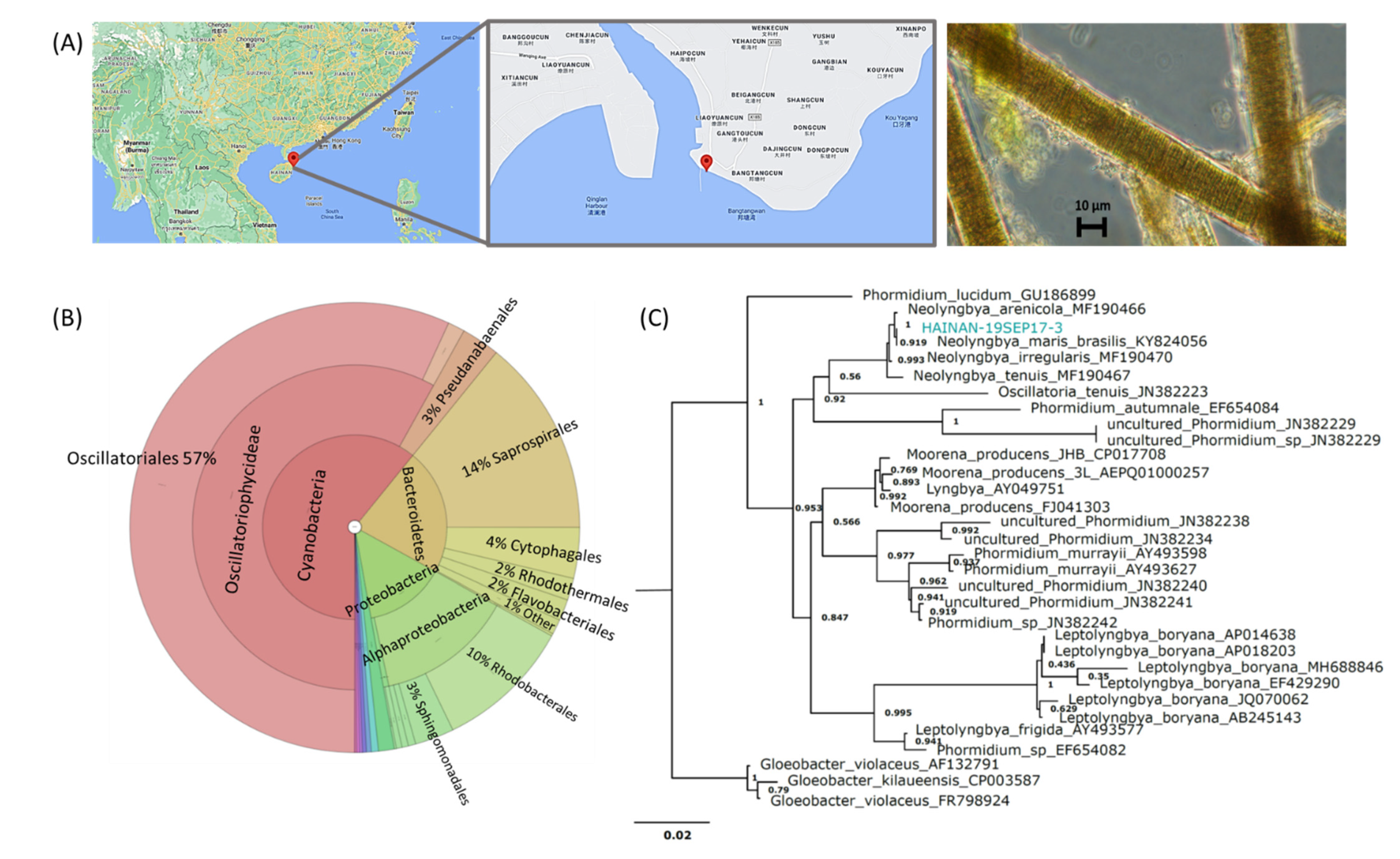
2.1. Sample Evaluation
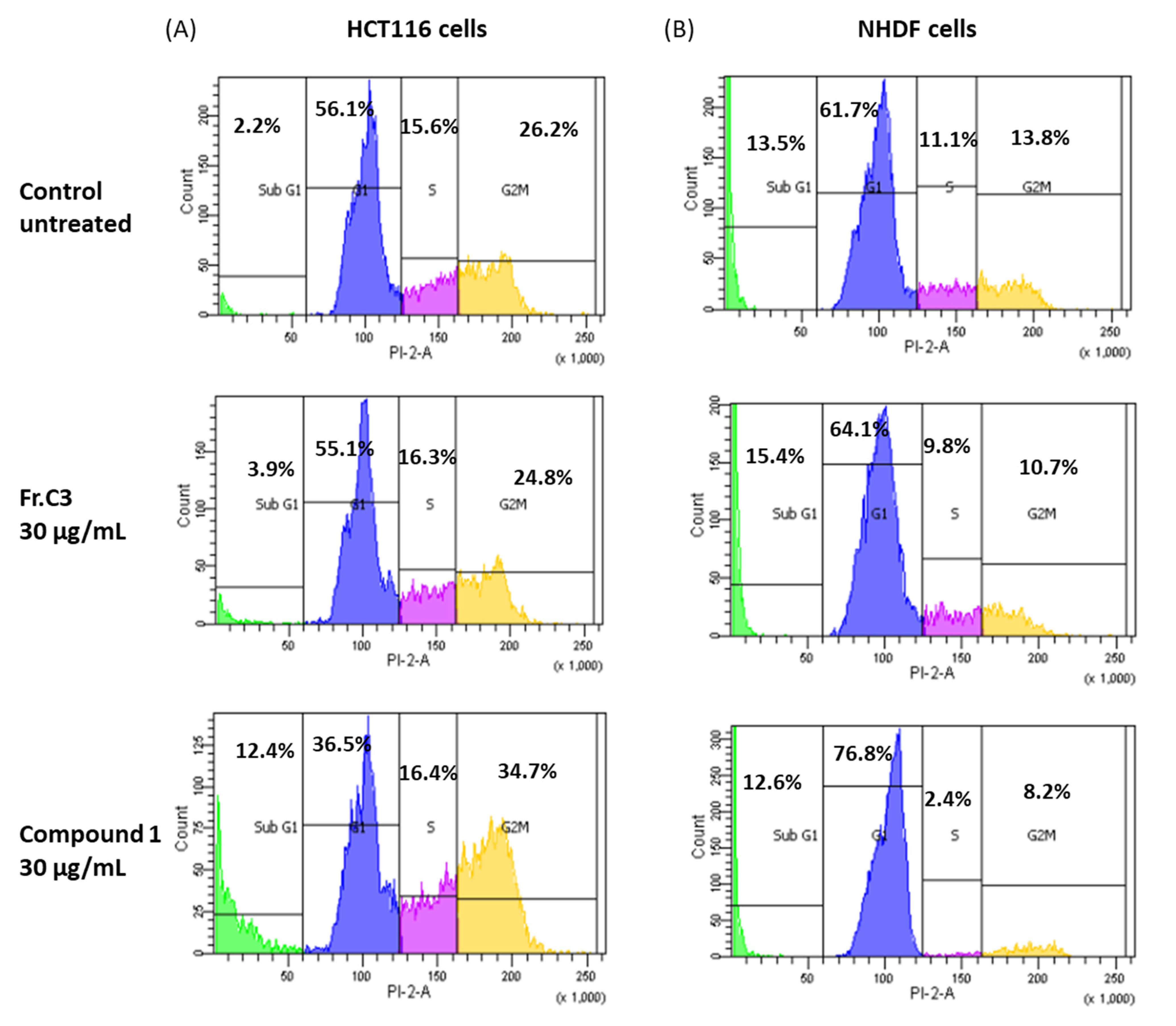
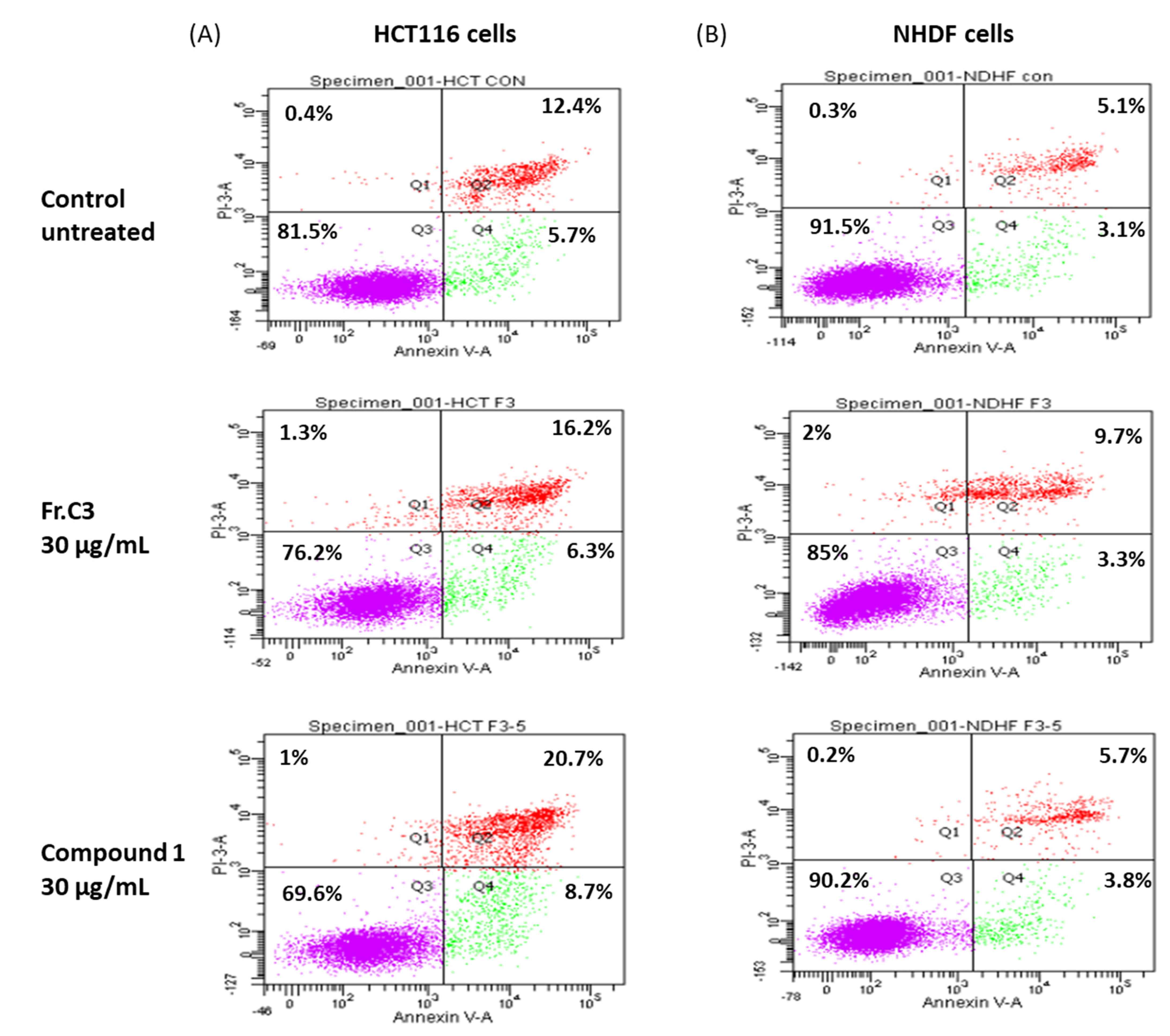
2.2. Inhibition Activity on Human Colon Cancer Cells In Vitro
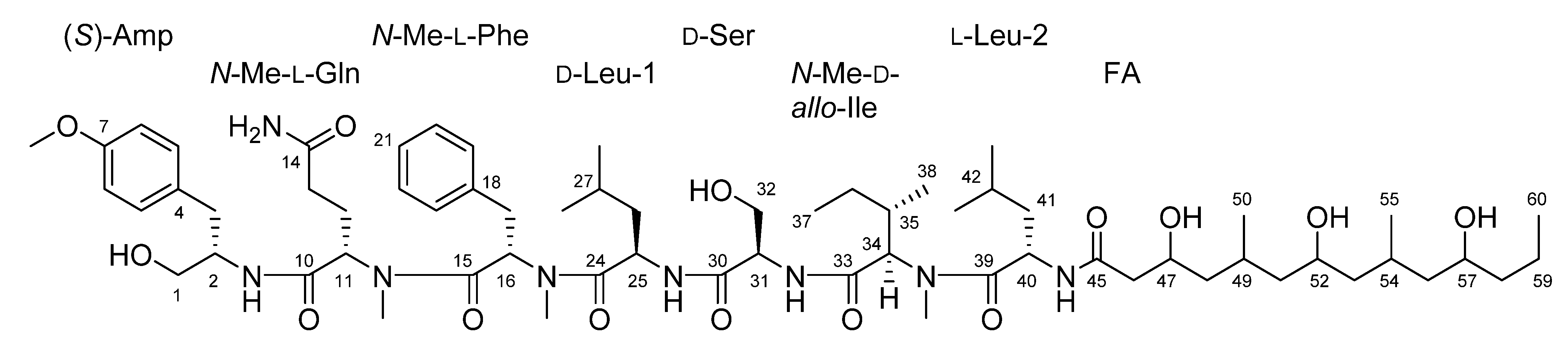
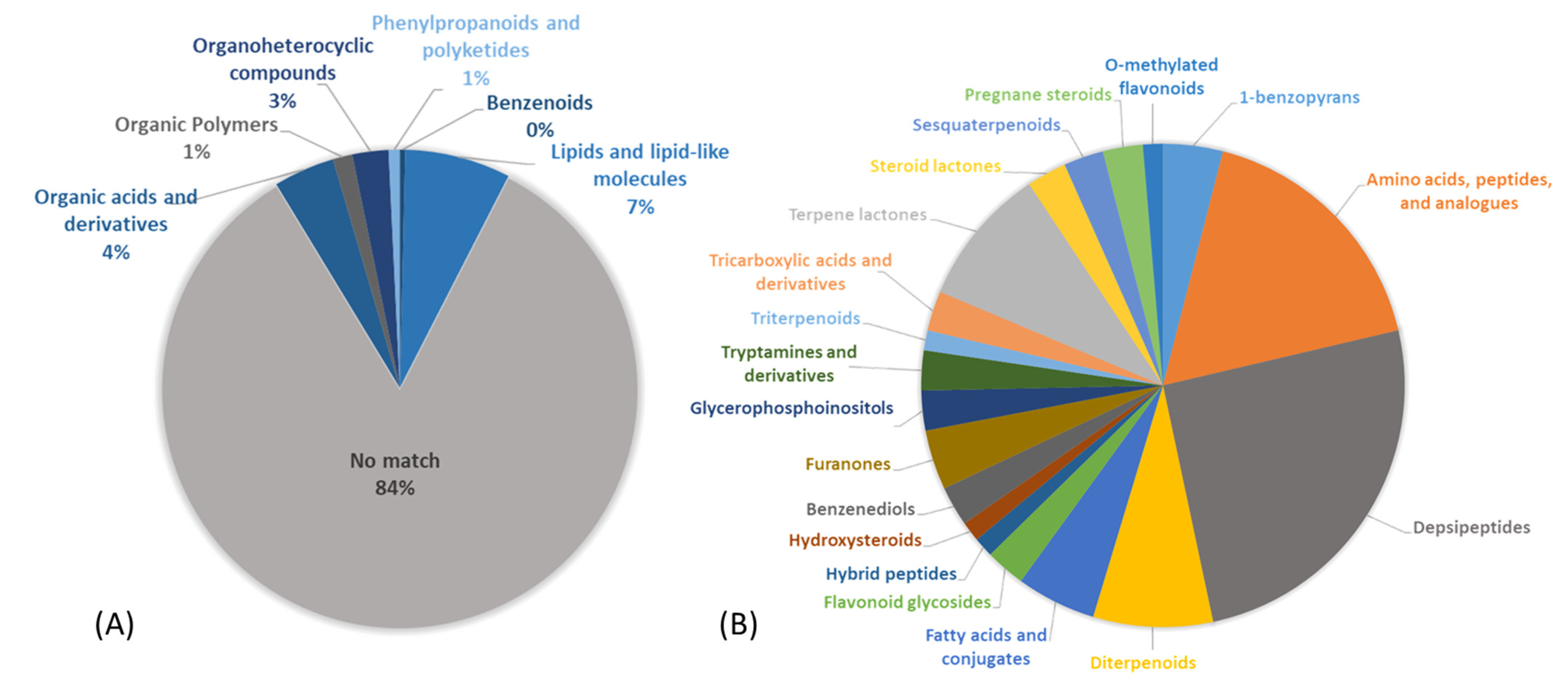
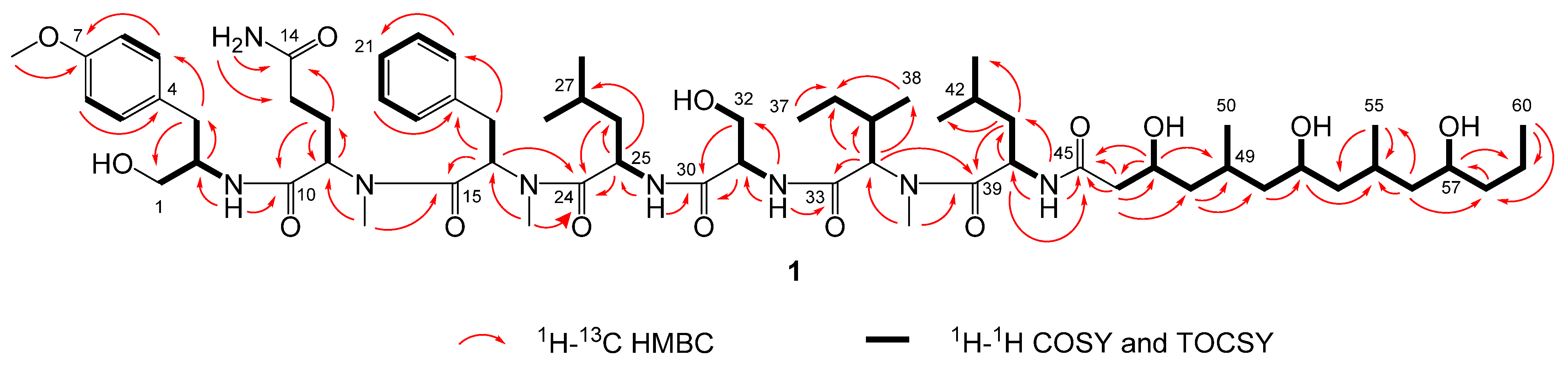
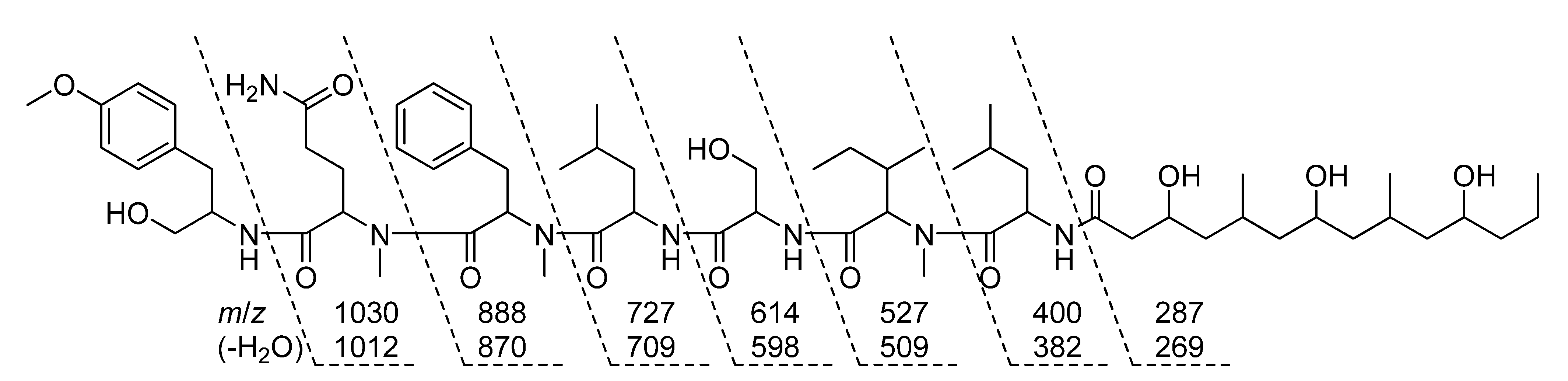
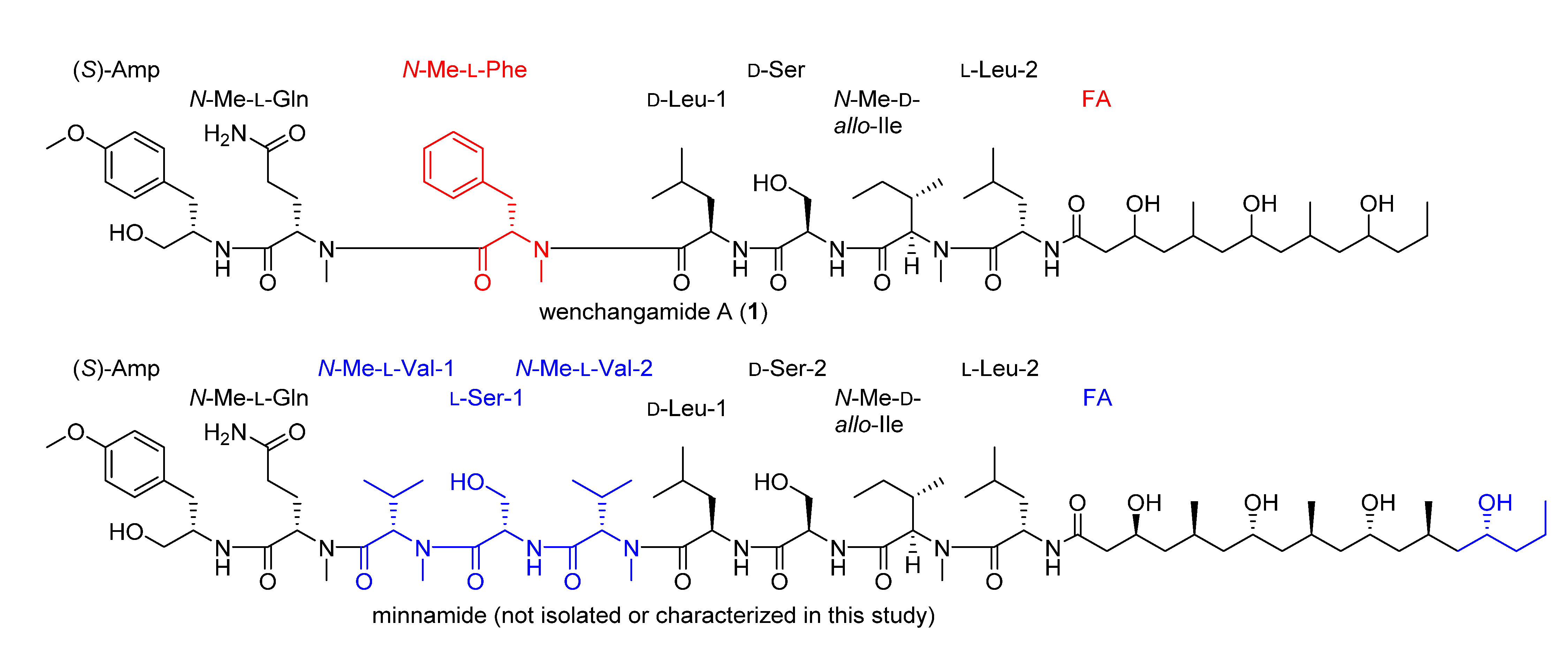
2.3. Natural Product Structure Elucidation
2.4. Additional Structure Hypothesis Generation
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Cyanobacterial Collection and Taxonomy
3.3. Extraction and Isolation
3.4. Isolated Materials (New Natural Products)
3.5. Determination of the Absolute Configuration of Wenchangamide A (1)
3.6. LC−MS Analysis and Molecular Networking Generation
3.7. Cell Culture
3.8. XTT Cell Proliferation Assay
3.9. Cell Cycle Analysis
3.10. Annexin-V/PI Double Staining
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salvador-Reyes, L.A.; Luesch, H. Biological targets and mechanisms of action of natural products from marine cyanobacteria. Nat. Prod. Rep. 2015, 32, 478–503. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demay, J.; Bernard, C.; Reinhardt, A.; Marie, B. Natural products from cyanobacteria: Focus on beneficial activities. Mar. Drugs 2019, 17, 320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, L.T.; Phyo, M.Y. Marine cyanobacteria: A source of lead compounds and their clinically-relevant molecular targets. Molecules 2020, 25, 2197. [Google Scholar] [CrossRef] [PubMed]
- Mayer, A.M.S.; Guerrero, A.J.; Rodríguez, A.D.; Taglialatela-Scafati, O.; Nakamura, F.; Fusetani, N. Marine pharmacology in 2014–2015: Marine compounds with antibacterial, antidiabetic, antifungal, anti-inflammatory, antiprotozoal, antituberculosis, antiviral, and anthelmintic activities; affecting the immune and nervous systems, and other miscellaneous mechanisms of action. Mar. Drugs 2020, 18, 5. [Google Scholar] [CrossRef] [Green Version]
- Jang, M.-H.; Ha, K.; Lucas, M.C.; Joo, G.-J.; Takamura, N. Changes in microcystin production by Microcystis aeruginosa exposed to phytoplanktivorous and omnivorous fish. Aquat. Toxicol. 2004, 68, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.; Gao, H.; Zhang, L.; Liang, H.; Zhu, X. Rapid evolution of tolerance to toxic Microcystis in two cladoceran grazers. Sci. Rep. 2016, 6, 25319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leao, T.; Castelão, G.; Korobeynikov, A.; Monroe, E.A.; Podell, S.; Glukhov, E.; Allen, E.E.; Gerwick, W.H.; Gerwick, L. Comparative genomics uncovers the prolific and distinctive metabolic potential of the cyanobacterial genus Moorea. Proc. Natl. Acad. Sci. USA 2017, 114, 3198–3203. [Google Scholar] [CrossRef] [Green Version]
- Moss, N.A.; Leao, T.; Glukhov, E.; Gerwick, L.; Gerwick, W.H. Collection, Culturing, and Genome Analyses of Tropical Marine Filamentous Benthic Cyanobacteria. In Methods in Enzymology; Moore, B.S., Ed.; Elsevier: Cambridge, UK, 2018; Volume 604, pp. 3–43. ISBN 9780128139592. [Google Scholar]
- Crnkovic, C.M.; May, D.S.; Orjala, J. The impact of culture conditions on growth and metabolomic profiles of freshwater cyanobacteria. J. Appl. Phycol. 2018, 30, 375–384. [Google Scholar] [CrossRef]
- Pye, C.R.; Bertin, M.J.; Lokey, R.S.; Gerwick, W.H.; Linington, R.G. Retrospective analysis of natural products provides insights for future discovery trends. Proc. Natl. Acad. Sci. USA 2017, 114, 5601–5606. [Google Scholar] [CrossRef] [Green Version]
- Hašler, P.; Dvořák, P.; Johansen, J.R.; Kitner, M.; Ondřej, V.; Poulíčková, A. Morphological and molecular study of epipelic filamentous genera Phormidium, Microcoleus and Geitlerinema (Oscillatoriales, Cyanophyta/cyanobacteria). Fottea-Olomouc 2012, 12, 341–356. [Google Scholar] [CrossRef] [Green Version]
- Stoyanov, P.; Moten, D.; Mladenov, R.; Dzhambazov, B.; Teneva, I. Phylogenetic relationships of some filamentous cyanoprokaryotic species. Evol. Bioinform. 2014, 10, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Nuryadi, H.; Sumimoto, S.; Teruya, T.; Suenaga, K.; Suda, S. Characterization of macroscopic colony-forming filamentous cyanobacteria from Okinawan coasts as potential sources of bioactive compounds. Mar. Biotechnol. 2020, 22, 824–835. [Google Scholar] [CrossRef]
- Anagnostidis, K. Nomenclatural changes in cyanoprokaryotic order Oscillatoriales. Preslia Praha 2001, 73, 359–375. [Google Scholar]
- Hoffmann, L.; Komárek, J.; Kaštovský, J. System of cyanoprokaryotes (cyanobacteria) state in 2004. Arch. Hydrobiol. Suppl. Algol. Stud. 2005, 117, 95–115. [Google Scholar] [CrossRef]
- Komárek, J.; Johansen, J.R. Filamentous Cyanobacteria. In Freshwater Algae of North America: Ecology and Classification, 1st ed.; Academic Press: Cambridge, MA, USA, 2003; pp. 117–196. ISBN 9780123858771. [Google Scholar]
- Komárek, J.; Johansen, J.R. Filamentous Cyanobacteria. In Freshwater Algae of North America: Ecology and Classification, 2nd ed.; Academic Press: Cambridge, MA, USA, 2015; pp. 135–235. ISBN 9780123858771. [Google Scholar]
- Komárek, J.; Kaštovský, J.; Mareš, J.; Johansen, J.R. Taxonomic classification of cyanoprokaryotes (cyanobacterial genera) 2014, using a polyphasic approach. Preslia 2014, 86, 295–335. [Google Scholar]
- Engene, N.; Paul, V.J.; Byrum, T.; Gerwick, W.H.; Thor, A.; Ellisman, M.H. Five chemically rich species of tropical marine cyanobacteria of the genus Okeania gen. nov. (Oscillatoriales, Cyanoprokaryota). J. Phycol. 2013, 49, 1095–1106. [Google Scholar] [CrossRef]
- Tronholm, A.; Engene, N. Moorena gen. nov., a valid name for “Moorea Engene & al.” nom. inval. (Oscillatoriaceae, Cyanobacteria). Not. Algarum 2019, 122, 122. [Google Scholar]
- Caires, T.A.; de Mattos Lyra, G.; Hentschke, G.S.; de Gusmão Pedrini, A.; Sant’Anna, C.L.; de Castro Nunes, J.M. Neolyngbya gen. nov. (Cyanobacteria, Oscillatoriaceae): A new filamentous benthic marine taxon widely distributed along the Brazilian coast. Mol. Phylogenet. Evol. 2018, 120, 196–211. [Google Scholar] [CrossRef]
- Caires, T.A.; da Silva, A.M.S.; Vasconcelos, V.M.; Affe, H.M.J.; de Souza Neta, L.C.; Boness, H.V.M.; Sant’Anna, C.L.; Nunes, J.M.C. Biotechnological potential of Neolyngbya (Cyanobacteria), a new marine benthic filamentous genus from Brazil. Algal Res. 2018, 36, 1–9. [Google Scholar] [CrossRef]
- Lydon, C.A.; Mathivathanan, L.; Sanchez, J.; dos Santos, L.A.H.; Sauvage, T.; Gunasekera, S.P.; Paul, V.J.; Berry, J.P. Eudesmacarbonate, a eudesmane-type sesquiterpene from a marine filamentous cyanobacterial mat (Oscillatoriales) in the Florida Keys. J. Nat. Prod. 2020, 83, 2030–2035. [Google Scholar] [CrossRef] [PubMed]
- Guan, H.; Wang, S. Algae. In Chinese Marine Materia Medica; Shanghai Scientific and Technical Publishers, China Ocean Press, and Chemical Industry Press: Shanghai, Beijing, China, 2009; pp. 37–304. ISBN 978-7-5323-9958-1. [Google Scholar]
- Titlyanov, E.A.; Titlyanova, T.V.; Li, X.; Huang, H. Chapter 2—Marine Plants of Coral Reefs. In Coral Reef Marine Plants of Hainan Island; Titlyanov, E.A., Titlyanova, T.V., Li, X., Huang, H., Eds.; Academic Press: Cambridge, MA, USA, 2017; pp. 5–39. ISBN 978-0-12-811963-1. [Google Scholar]
- Sun, W.; Wu, W.; Liu, X.; Zaleta-Pinet, D.A.; Clark, B.R. Bioactive compounds isolated from marine-derived microbes in China: 2009-2018. Mar. Drugs 2019, 17, 339. [Google Scholar] [CrossRef] [Green Version]
- Demarque, D.P.; Dusi, R.G.; de Sousa, F.D.M.; Grossi, S.M.; Silvério, M.R.S.; Lopes, N.P.; Espindola, L.S. Mass spectrometry-based metabolomics approach in the isolation of bioactive natural products. Sci. Rep. 2020, 10, 1051. [Google Scholar] [CrossRef] [Green Version]
- Luzzatto-Knaan, T.; Garg, N.; Wang, M.; Glukhov, E.; Peng, Y.; Ackermann, G.; Amir, A.; Duggan, B.M.; Ryazanov, S.; Gerwick, L.; et al. Digitizing mass spectrometry data to explore the chemical diversity and distribution of marine cyanobacteria and algae. eLife 2017, 6, e24214. [Google Scholar] [CrossRef]
- Luzzatto-Knaan, T.; Melnik, A.V.; Dorrestein, P.C. Mass spectrometry tools and workflows for revealing microbial chemistry. Analyst 2015, 140, 4949–4966. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.Y.; Sanchez, L.M.; Rath, C.M.; Liu, X.; Boudreau, P.D.; Bruns, N.; Glukhov, E.; Wodtke, A.; de Felicio, R.; Fenner, A.; et al. Molecular networking as a dereplication strategy. J. Nat. Prod. 2013, 76, 1686–1699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olivon, F.; Allard, P.-M.; Koval, A.; Righi, D.; Genta-Jouve, G.; Neyts, J.; Apel, C.; Pannecouque, C.; Nothias, L.-F.; Cachet, X.; et al. Bioactive natural products prioritization using massive multi-informational molecular networks. ACS Chem. Biol. 2017, 12, 2644–2651. [Google Scholar] [CrossRef]
- Nothias, L.F.; Nothias-Esposito, M.; da Silva, R.; Wang, M.; Protsyuk, I.; Zhang, Z.; Sarvepalli, A.; Leyssen, P.; Touboul, D.; Costa, J.; et al. Bioactivity-based molecular networking for the discovery of drug leads in natural product bioassay-guided fractionation. J. Nat. Prod. 2018, 81, 758–767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolfender, J.L.; Litaudon, M.; Touboul, D.; Queiroz, E.F. Innovative omics-based approaches for prioritisation and targeted isolation of natural products—New strategies for drug discovery. Nat. Prod. Rep. 2019, 36, 855–868. [Google Scholar] [CrossRef] [Green Version]
- Fox Ramos, A.E.; Evanno, L.; Poupon, E.; Champy, P.; Beniddir, M.A. Natural products targeting strategies involving molecular networking: Different manners, one goal. Nat. Prod. Rep. 2019, 36, 960–980. [Google Scholar] [CrossRef]
- Ernst, M.; Kang, K.B.; Caraballo-Rodríguez, A.M.; Nothias, L.F.; Wandy, J.; Wang, M.; Rogers, S.; Medema, M.H.; Dorrestein, P.C.; van der Hooft, J.J.J. MolNetEnhancer: Enhanced molecular networks by integrating metabolome mining and annotation tools. Metabolites 2019, 9, 144. [Google Scholar] [CrossRef] [Green Version]
- Cornet, L.; Bertrand, A.R.; Hanikenne, M.; Javaux, E.J.; Wilmotte, A.; Baurain, D. Metagenomic assembly of new (sub)polar cyanobacteria and their associated microbiome from non-axenic cultures. Microb. Genom. 2018, 4, 212. [Google Scholar] [CrossRef]
- Gris, B.; Treu, L.; Zampieri, R.M.; Caldara, F.; Romualdi, C.; Campanaro, S.; La Rocca, N. Microbiota of the therapeutic Euganean thermal muds with a focus on the main cyanobacteria species. Microorganisms 2020, 8, 1590. [Google Scholar] [CrossRef] [PubMed]
- Gogineni, V.; Hamann, M.T. Marine natural product peptides with therapeutic potential: Chemistry, biosynthesis, and pharmacology. Biochim. Biophys. Acta Gen. Subj. 2018, 1862, 81–196. [Google Scholar] [CrossRef]
- Gross, H.; König, G.M. Terpenoids from marine organisms: Unique structures and their pharmacological potential. Phytochem. Rev. 2006, 5, 115–141. [Google Scholar] [CrossRef]
- Sorokina, M.; Steinbeck, C. Review on natural products databases: Where to find data in 2020. J. Cheminform. 2020, 12, 1–51. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Fewer, D.P.; Holm, L.; Rouhiainen, L.; Sivonen, K. Atlas of nonribosomal peptide and polyketide biosynthetic pathways reveals common occurrence of nonmodular enzymes. Proc. Natl. Acad. Sci. USA 2014, 111, 9259–9264. [Google Scholar] [CrossRef] [Green Version]
- Naman, C.B.; Rattan, R.; Nikoulina, S.E.; Lee, J.; Miller, B.W.; Moss, N.A.; Armstrong, L.; Boudreau, P.D.; Debonsi, H.M.; Valeriote, F.A.; et al. Integrating molecular networking and biological assays to target the isolation of a cytotoxic cyclic octapeptide, samoamide A, from an American Samoan marine cyanobacterium. J. Nat. Prod. 2017, 80, 625–633. [Google Scholar] [CrossRef]
- Bar-Shalom, R.; Bergman, M.; Grossman, S.; Azzam, N.; Sharvit, L.; Fares, F. Inula viscosa extract inhibits growth of colorectal cancer cells in vitro and in vivo through induction of apoptosis. Front. Oncol. 2019, 9, 227. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Q.; Hirose, Y.; Yoshimi, N.; Murakami, A.; Koshimizu, K.; Ohigashi, H.; Sakata, K.; Matsumoto, Y.; Sayama, Y.; Mori, H. Further investigation of the modifying effect of various chemopreventive agents on apoptosis and cell proliferation in human colon cancer cells. J. Cancer Res. Clin. Oncol. 2002, 128, 539–546. [Google Scholar] [CrossRef]
- Sumimoto, S.; Kobayashi, M.; Sato, R.; Shinomiya, S.; Iwasaki, A.; Suda, S.; Teruya, T.; Inuzuka, T.; Ohno, O.; Suenaga, K. Minnamide A, a linear lipopeptide from the marine cyanobacterium Okeania hirsuta. Org. Lett. 2019, 21, 1187–1190. [Google Scholar] [CrossRef]
- Liang, X.; Matthew, S.; Chen, Q.Y.; Kwan, J.C.; Paul, V.J.; Luesch, H. Discovery and total synthesis of doscadenamide A: A quorum sensing signaling molecule from a marine cyanobacterium. Org. Lett. 2019, 21, 7274–7278. [Google Scholar] [CrossRef]
- Leber, C.A.; Naman, C.B.; Keller, L.; Almaliti, J.; Caro-Diaz, E.J.E.; Glukhov, E.; Joseph, V.; Sajeevan, T.P.; Reyes, A.J.; Biggs, J.S.; et al. Applying a chemogeographic strategy for natural product discovery from the marine cyanobacterium Moorena bouillonii. Mar. Drugs 2020, 18, 515. [Google Scholar] [CrossRef]
- Liang, X.; Chen, Q.-Y.; Seabra, G.M.; Matthew, S.; Kwan, J.C.; Li, C.; Paul, V.J.; Luesch, H. Bifunctional doscadenamides activate quorum sensing in gram-negative bacteria and synergize with TRAIL to induce apoptosis in cancer cells. J. Nat. Prod. 2021, 84, 779–789. [Google Scholar] [CrossRef]
- Ondov, B.D.; Bergman, N.H.; Phillippy, A.M. Interactive metagenomic visualization in a Web browser. BMC Bioinform. 2011, 12, 385. [Google Scholar] [CrossRef] [Green Version]
- Pruesse, E.; Peplies, J.; Glöckner, F.O. SINA: Accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 2012, 28, 1823–1829. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Moiety | Position | δC | Type | δH, Mult (J in Hz) | Moiety | Position | δC | Type | δH, Mult (J in Hz) |
|---|---|---|---|---|---|---|---|---|---|
| AMP | 1 | 62.7 | CH2 | 3.35, m c | N-Me-Ile | 33 | 169.4 | C | |
| 2 | 52.4 | CH | 3.91, ddd (9.4, 5.1, 4.9) | 34 | 59.9 | CH | 4.67, d (10.7) | ||
| 3 | 35.6 | CH2 | 2.77, dd (14.0, 5.1); 2.53, dd (14.0, 4.9) | 35 | 32.6 | CH | 1.9, m c | ||
| 4 | 130.9 | C | 36 | 25.5 | CH2 | 1.35, m c; 0.99, m | |||
| 5, 9 | 130.0 | CH | 7.04, d (8.6) | 37 | 11.3 | CH3 | 0.81, m c | ||
| 6, 8 | 113.5 | CH | 6.79, d (8.6) | 38 | 14.6 | CH3 | 0.68, d (6.7) | ||
| 7 | 157.6 | C | 34-N-Me | 30.5 | CH3 | 2.94, s | |||
| 7-O-Me | 55.0 c | CH3 | 3.71, s | ||||||
| 2-NH | 7.27, mc | Leu-2 | 39 | 173.0 | C | ||||
| 40 | 47.4 | CH | 4.72, m | ||||||
| N-Me-Gln | 10 | 169.2 | C | 41 | 40.5 | CH2 | 1.46, mc; 1.35, mc | ||
| 11 | 56.0 | CH | 4.84, dd (5.1, 10.4) c | 42 | 24.3 | CH | 1.61, m c | ||
| 12 | 23.8 | CH2 | 1.97, m; 1.61, m c | 43 | 23.1 | CH3 | 0.87, d (6.6) c | ||
| 13 | 31.3 | CH2 | 1.90, m c; 1.83, m c | 44 | 21.5 | CH3 | 0.87, d (6.6) c | ||
| 14 | 173.7 | C | 40-NH | 8.15, d (7.8) | |||||
| 14-NH2 | 7.28, mc; 6.71, br s | ||||||||
| 11-N-Me | 29.8 | CH3 | 2.42, s | FA | 45 | 171.0 | C | ||
| 46 | 43.8 | CH2 | 2.22, dd (14.0, 5.4) | ||||||
| N-Me-Phe | 15 | 170.3 | C | 2.13, dd (14.0, 7.4) | |||||
| 16 | 54.0 | CH | 5.62, dd (10.0, 5.9) | 47 | 65.7 | CH | 3.87, m | ||
| 17 | 34.6 | CH2 | 2.99 m; 2.94 m | 48 | 45.2 | CH2 | 1.23, m; 0.92, m c | ||
| 18 | 137.3 | C | 49 | 25.6 | CH | 1.76, m | |||
| 19, 23 | 129.5 | CH | 7.26, m | 50 | 20.3 | CH3 | 0.82, d (6.7) | ||
| 20, 22 | 128.0 | CH | 7.23, m | 51 | 45.3 | CH2 | 1.35, m c; 0.92, m c | ||
| 21 | 126.3 | CH | 7.17, t (6.7) | 52 | 64.9 | CH | 3.55, m | ||
| 16-N-Me | 30.1 | CH3 | 2.89, s | 53 | 46.7 | CH2 | 1.26, m c; 1.04, m | ||
| 54 | 25.3 | CH | 1.83, m c | ||||||
| Leu-1 | 24 | 171.4 | C | 55 | 19.2 | CH3 | 0.81, d (6.6) c | ||
| 25 | 46.9 | CH | 4.49, m | 56 | 45.7 | CH2 | 1.25 m c; 1.02 m c | ||
| 26 | 39.7c | CH2 | 0.95, m; 0.73, m | 57 | 67.0 | CH | 3.46, m c | ||
| 27 | 23.7 | CH | 1.12, m | 58 | 40.4 | CH2 | 1.46, m c; 1.26, m c | ||
| 28 | 23.0 | CH3 | 0.70, d (6.7) | 59 | 18.6 | CH2 | 1.35, m c; 1.26, m c | ||
| 29 | 21.7 | CH3 | 0.69, d (6.7) | 60 | 14.2 | CH2 | 0.85, t (6.8) | ||
| 25-NH | 8.04, d (7.9) | 47-OH | 4.53, m | ||||||
| 52-OH | 4.11, br s | ||||||||
| Ser | 30 | 170.0 | C | 57-OH | 4.17, br s | ||||
| 31 | 55.0 c | CH | 4.23, dt (7.8, 5.7) | ||||||
| 32 | 61.6 | CH2 | 3.43, m c | ||||||
| 31-NH | 7.53, d (7.8) | ||||||||
| 32-OH | 4.84, m c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, L.; Bar-Shalom, R.; Aharonovich, D.; Kurisawa, N.; Patial, G.; Li, S.; He, S.; Yan, X.; Iwasaki, A.; Suenaga, K.; et al. Metabolomic Characterization of a cf. Neolyngbya Cyanobacterium from the South China Sea Reveals Wenchangamide A, a Lipopeptide with In Vitro Apoptotic Potential in Colon Cancer Cells. Mar. Drugs 2021, 19, 397. https://doi.org/10.3390/md19070397
Ding L, Bar-Shalom R, Aharonovich D, Kurisawa N, Patial G, Li S, He S, Yan X, Iwasaki A, Suenaga K, et al. Metabolomic Characterization of a cf. Neolyngbya Cyanobacterium from the South China Sea Reveals Wenchangamide A, a Lipopeptide with In Vitro Apoptotic Potential in Colon Cancer Cells. Marine Drugs. 2021; 19(7):397. https://doi.org/10.3390/md19070397
Chicago/Turabian StyleDing, Lijian, Rinat Bar-Shalom, Dikla Aharonovich, Naoaki Kurisawa, Gaurav Patial, Shuang Li, Shan He, Xiaojun Yan, Arihiro Iwasaki, Kiyotake Suenaga, and et al. 2021. "Metabolomic Characterization of a cf. Neolyngbya Cyanobacterium from the South China Sea Reveals Wenchangamide A, a Lipopeptide with In Vitro Apoptotic Potential in Colon Cancer Cells" Marine Drugs 19, no. 7: 397. https://doi.org/10.3390/md19070397
APA StyleDing, L., Bar-Shalom, R., Aharonovich, D., Kurisawa, N., Patial, G., Li, S., He, S., Yan, X., Iwasaki, A., Suenaga, K., Zhu, C., Luo, H., Tian, F., Fares, F., Naman, C. B., & Luzzatto-Knaan, T. (2021). Metabolomic Characterization of a cf. Neolyngbya Cyanobacterium from the South China Sea Reveals Wenchangamide A, a Lipopeptide with In Vitro Apoptotic Potential in Colon Cancer Cells. Marine Drugs, 19(7), 397. https://doi.org/10.3390/md19070397