
Toxicity Screening of a Gambierdiscus australes Strain from the Western Mediterranean Sea and Identification of a Novel Maitotoxin Analogue

 ,
, 
 ,
,  , , ,
, , ,  and
and 
Abstract
:1. Introduction
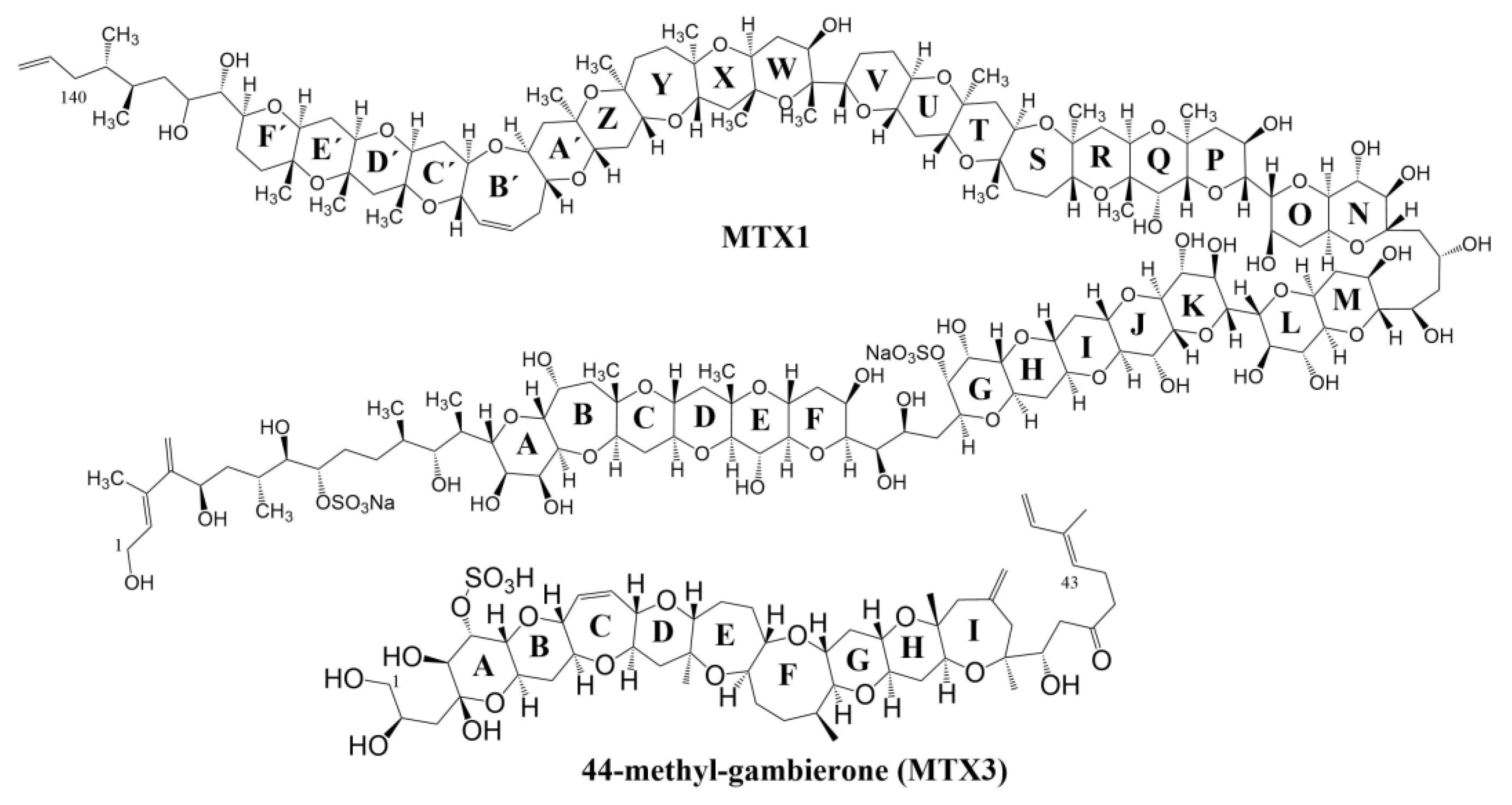
2. Results and Discussion
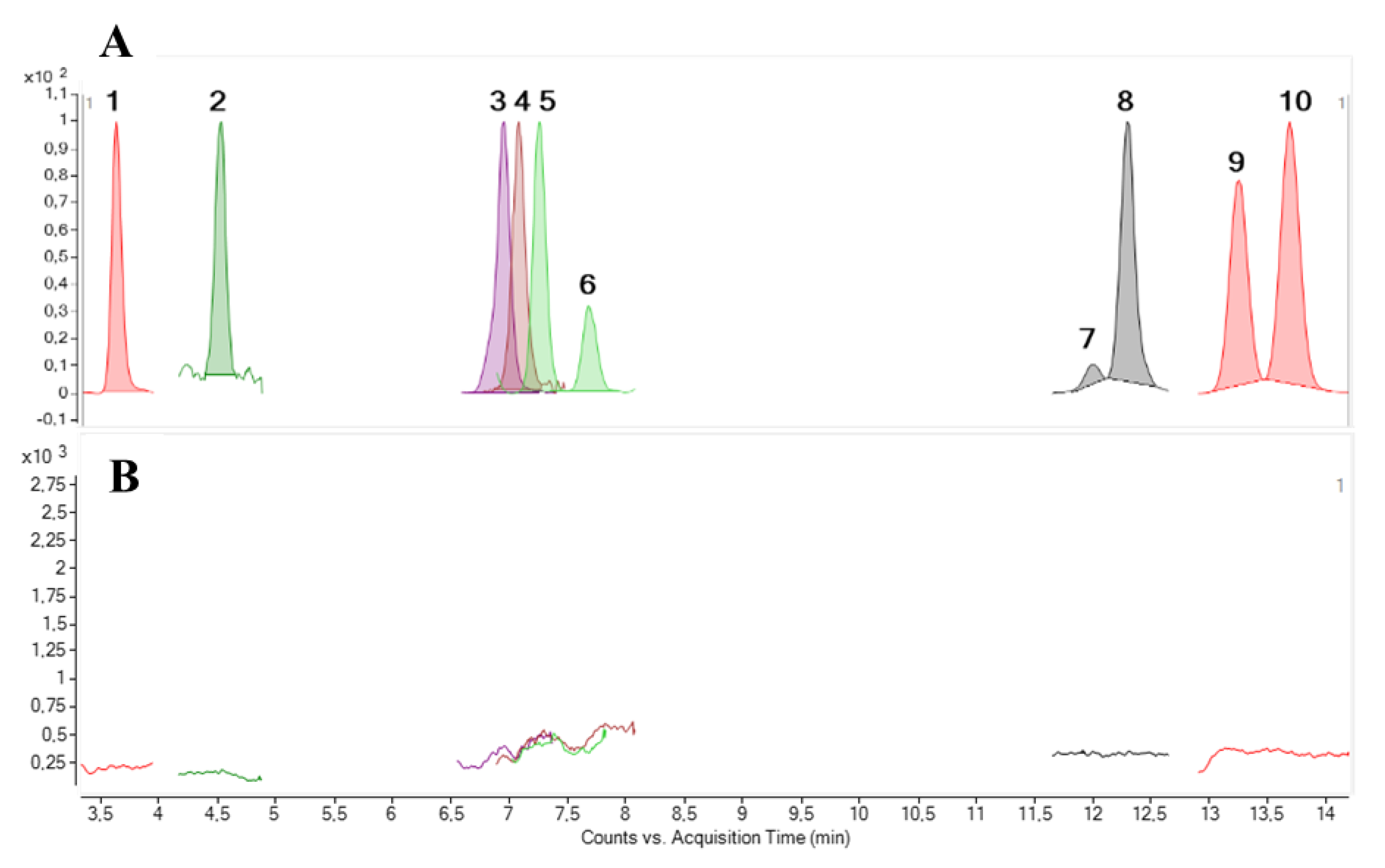
2.1. LC-MS/MS Analysis
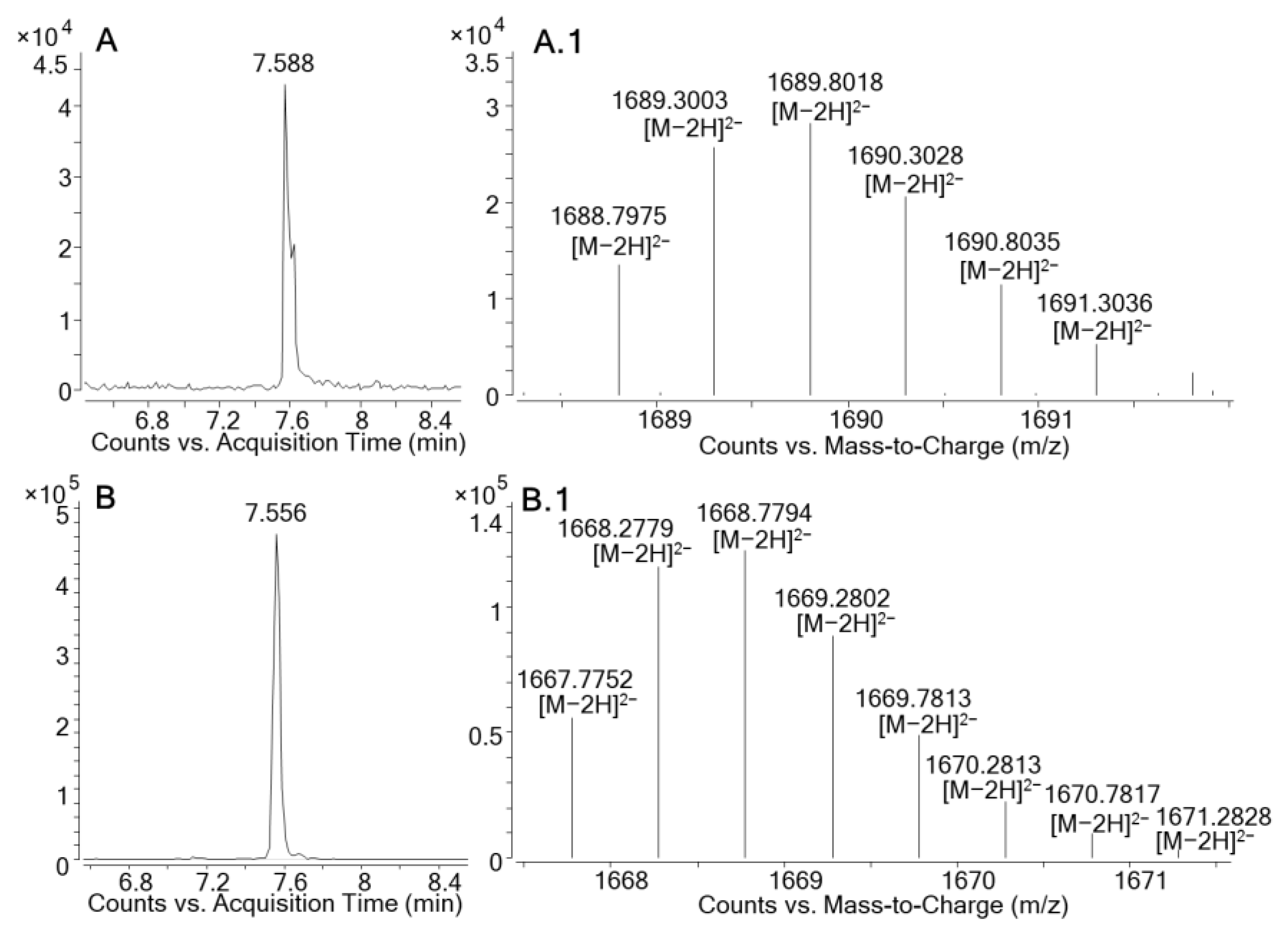
2.2. LC-HRMS Analysis
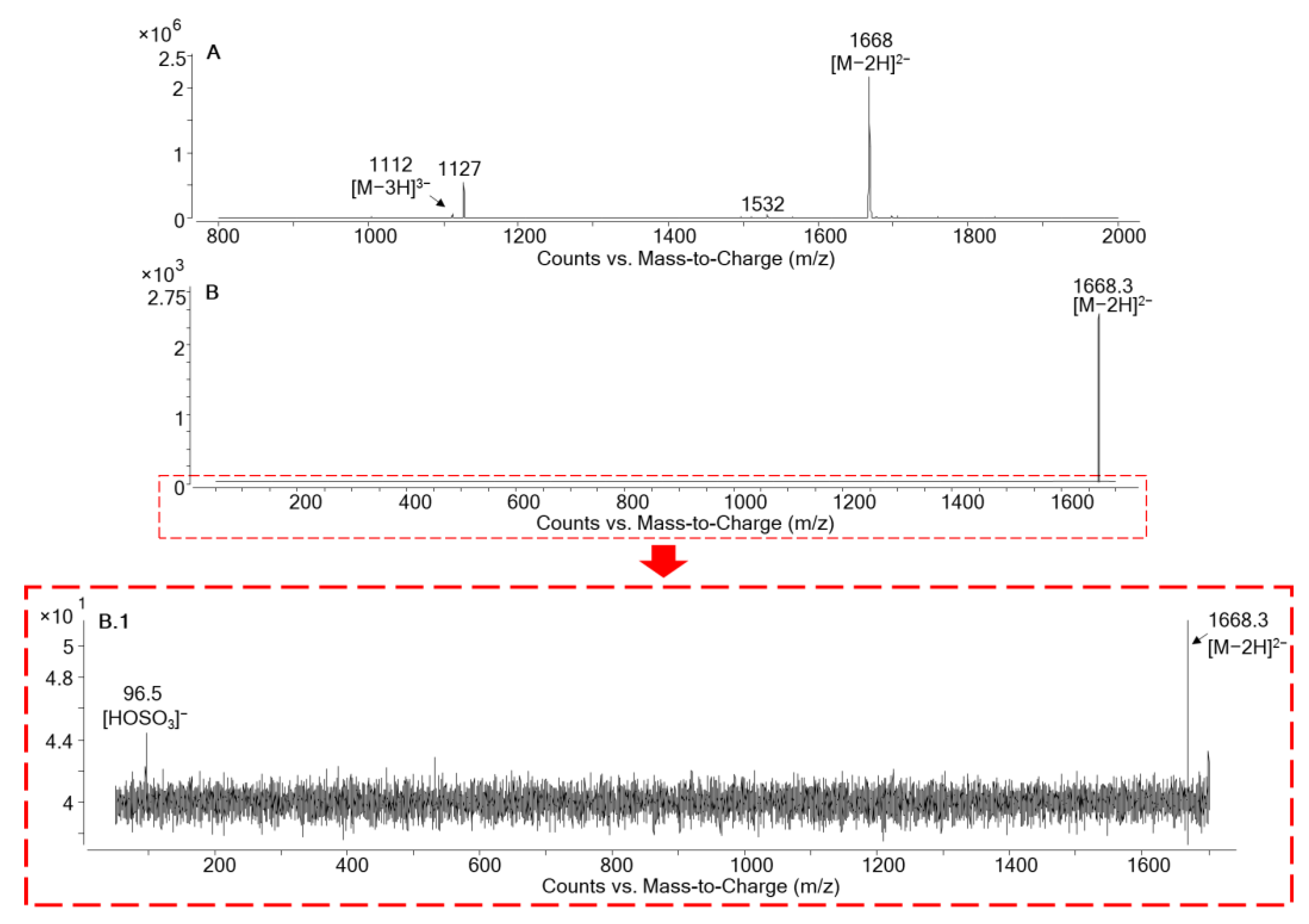
2.2.1. LC-HRMS in Negative Ionization Mode (ESI−)
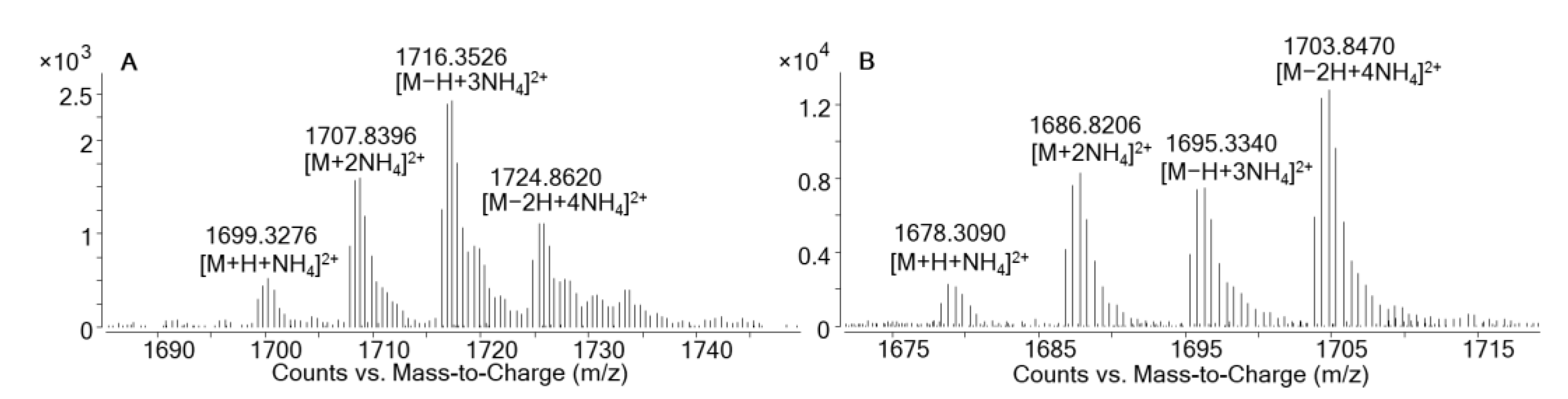
2.2.2. LC-HRMS in Positive Ionization Mode (ESI+)
2.2.3. Molecular Formula Determination for MTX5
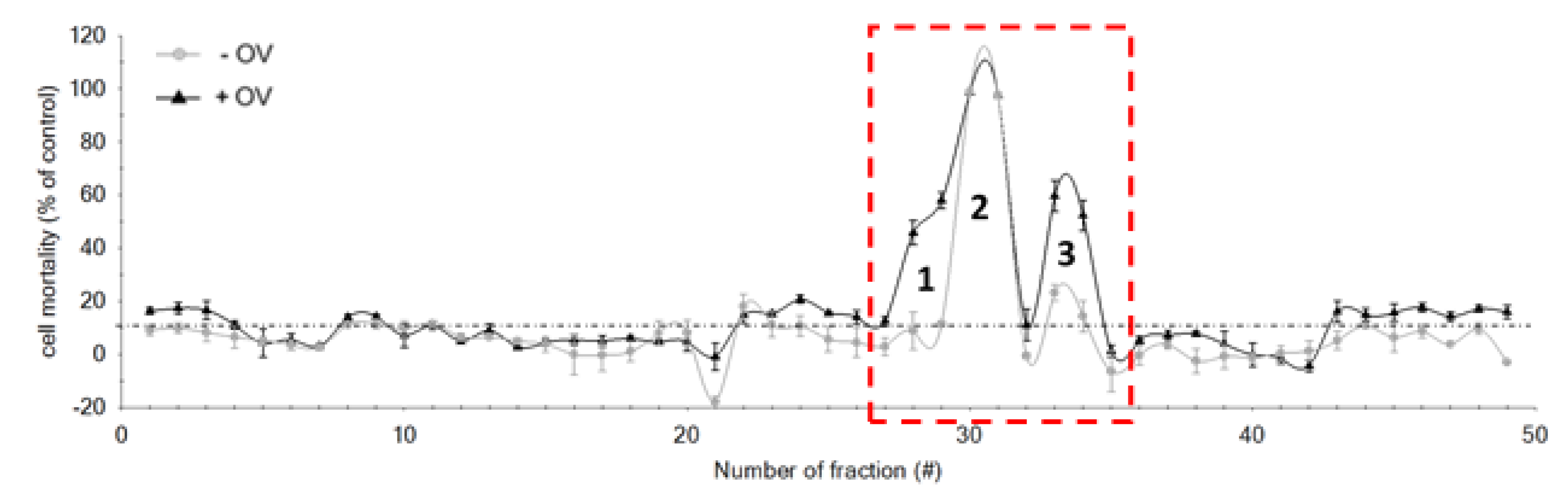
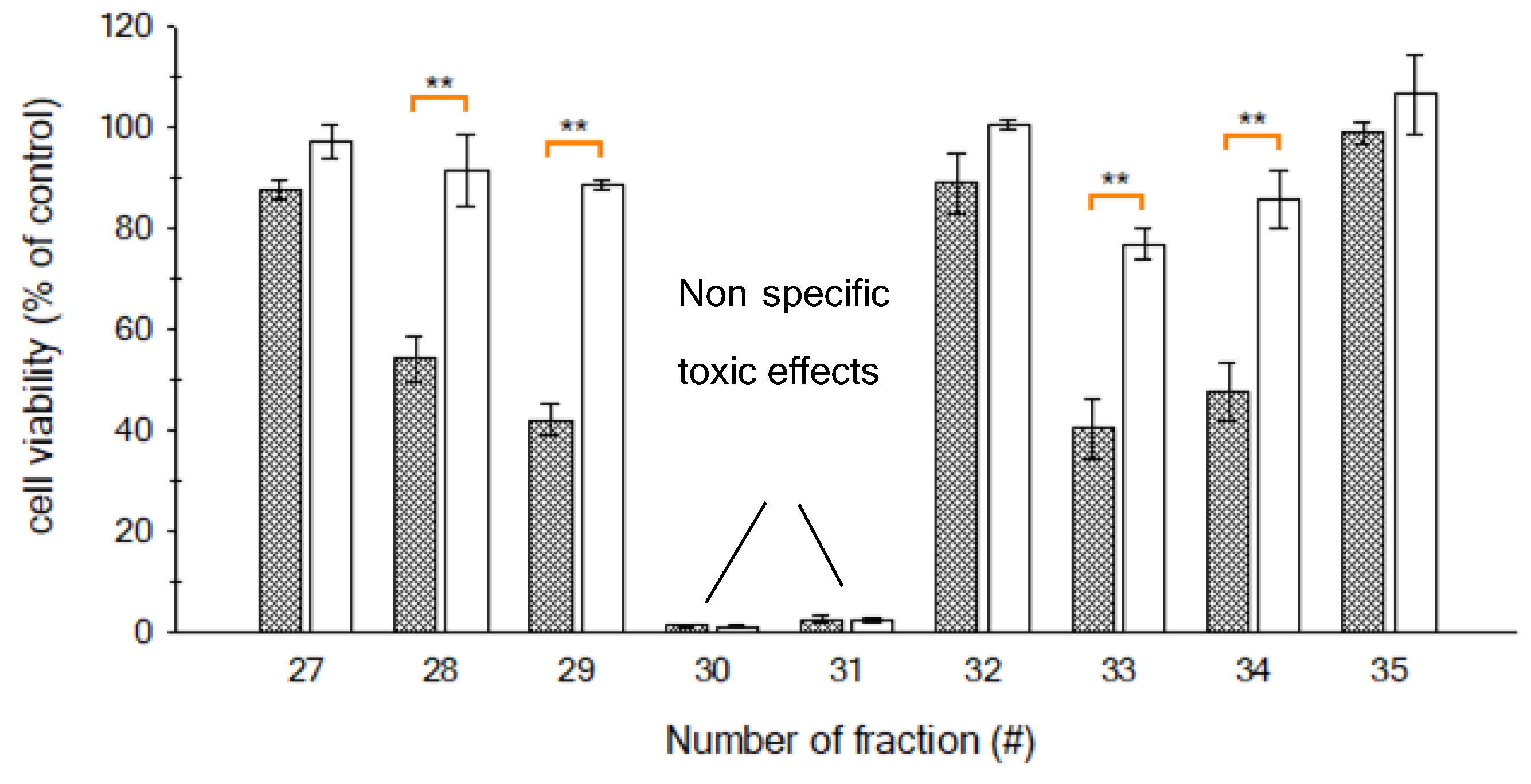
2.3. HPLC Fractionation
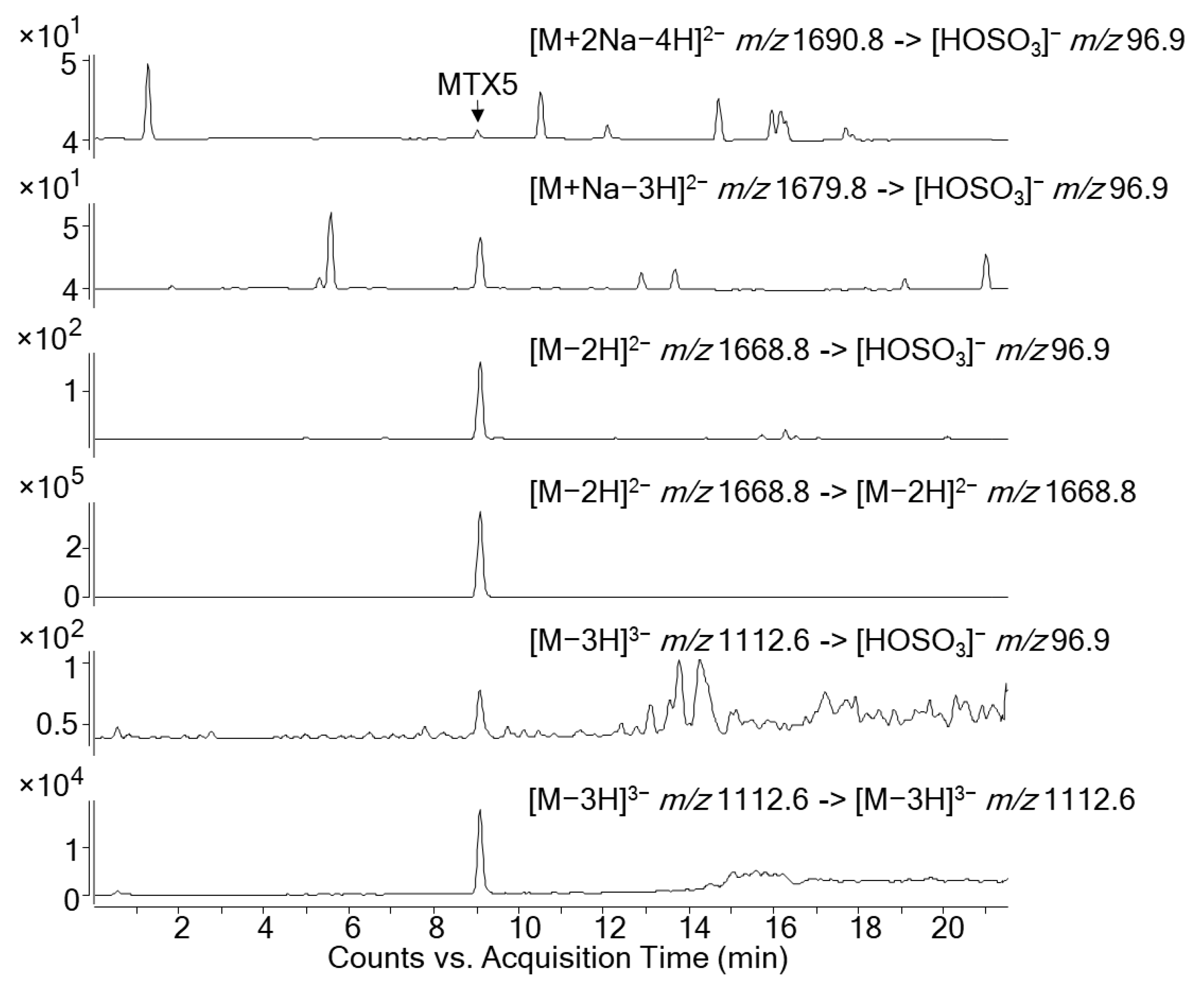
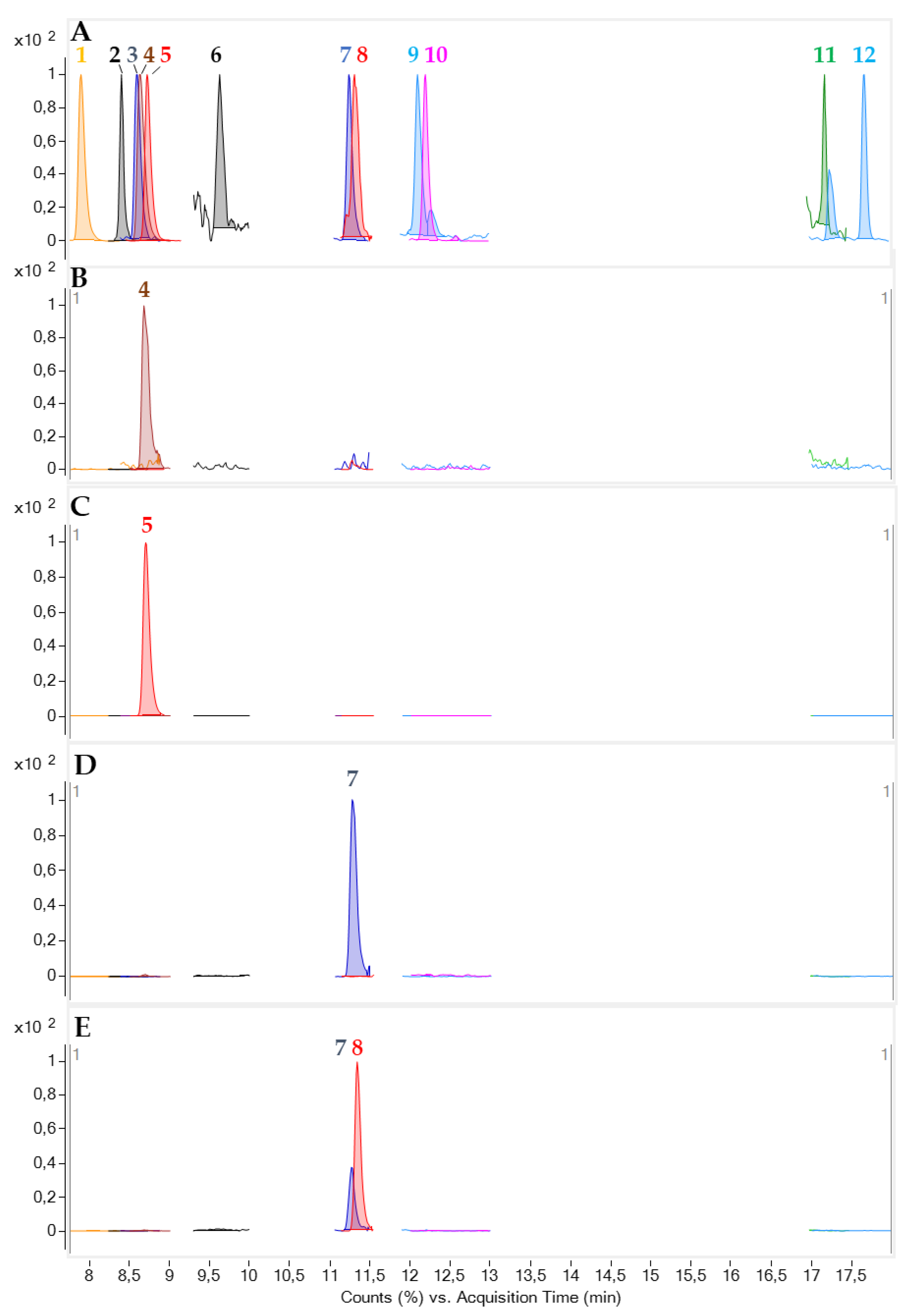
2.4. LC-MS/MS Analysis of the Toxic Fractions
3. Materials and Methods
3.1. Reference Materials and Chemicals
3.2. Gambierdiscus Australes Strain
3.3. Sample Pretreatment
3.4. HPLC-C18 Fractionation
3.5. N2a Assay
3.6. LC-HRMS (Agilent 6550 Q-Tof iFunnel)
3.7. LC-MS/MS (Agilent 6495 QQQ iFunnel)
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Yasumoto, T.; Nakajima, I.; Bagnis, R.; Adachi, R. Finding of a Dinoflagellate as a Likely Culprit of Ciguatera. Nippon. Suisan Gakkaishi 1977, 43, 1021–1026. [Google Scholar] [CrossRef] [Green Version]
- Yasumoto, T.; Murata, M. Marine Toxins. Chem. Rev. 1993, 93, 1897–1909. [Google Scholar] [CrossRef]
- Satake, M.; Murata, M.; Yasumoto, T. Gambierol: A New Toxic Polyether Compound Isolated from the Marine Dinoflagellate Gambierdiscus Toxicus. J. Am. Chem. Soc. 1993, 115, 361–362. [Google Scholar] [CrossRef]
- Watanabe, R.; Uchida, H.; Suzuki, T.; Matsushima, R.; Nagae, M.; Toyohara, Y.; Satake, M.; Oshima, Y.; Inoue, A.; Yasumoto, T. Gambieroxide, A Novel Epoxy Polyether Compound from the Dinoflagellate Gambierdiscus Toxicus GTP2 Strain. Tetrahedron 2013, 69, 10299–10303. [Google Scholar] [CrossRef]
- Rodríguez, I.; Genta-Jouve, G.; Alfonso, C.; Calabro, K.; Alonso, E.; Sánchez, J.A.; Alfonso, A.; Thomas, O.P.; Botana, L.M. Gambierone, A Ladder-Shaped Polyether from the Dinoflagellate Gambierdiscus Belizeanus. Org. Lett. 2015, 17, 2392–2395. [Google Scholar] [CrossRef] [PubMed]
- Nagai, H.; Murata, M.; Torigoe, K.; Satake, M.; Yasumoto, T. Gambieric Acids, New Potent Antifungal Substances with Unprecedented Polyether Structures from a Marine Dinoflagellate Gambierdiscus Toxicus. J. Org. Chem. 1992, 57, 5448–5453. [Google Scholar] [CrossRef]
- Lewis, R.J.; Holmes, M.J. Origin and Transfer of Toxins Involved in Ciguatera. Comp. Biochem. Physiol. C. 1993, 106, 615–628. [Google Scholar] [CrossRef]
- Oshiro, N.; Yogi, K.; Asato, S.; Sasaki, T.; Tamanaha, K.; Hirama, M.; Yasumoto, T.; Inafuku, Y. Ciguatera Incidence and Fish Toxicity in Okinawa, Japan. Toxicon 2010, 56, 656–661. [Google Scholar] [CrossRef]
- Hamilton, B.; Hurbungs, M.; Vernoux, J.-P.; Jones, A.; Lewis, R.J. Isolation and Characterisation of Indian Ocean Ciguatoxin. Toxicon 2002, 40, 685–693. [Google Scholar] [CrossRef]
- Vernoux, J.P.; Lewis, R.J. Isolation and Characterisation of Caribbean Ciguatoxins from the Horse-Eye Jack (Caranx latus). Toxicon 1997, 35, 889–900. [Google Scholar] [CrossRef]
- Ikehara, T.; Kuniyoshi, K.; Oshiro, N.; Yasumoto, T. Biooxidation of Ciguatoxins Leads to Species-Specific Toxin Profiles. Toxins 2017, 9, 205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Estevez, P.; Castro, D.; Pequeño-Valtierra, A.; Giraldez, J.; Gago-Martinez, A. Emerging Marine Biotoxins in Seafood from European Coasts: Incidence and Analytical Challenges. Foods 2019, 8, 149. [Google Scholar] [CrossRef] [Green Version]
- Costa, P.R.; Estevez, P.; Castro, D.; Soliño, L.; Gouveia, N.; Santos, C.; Rodrigues, S.M.; Leao, J.M.; Gago-Martínez, A. New Insights Into the Occurrence and Toxin Profile of Ciguatoxins in Selvagens Islands (Madeira, Portugal). Toxins 2018, 10, 524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodríguez, F.; Fraga, S.; Ramilo, I.; Rial, P.; Figueroa, R.I.; Riobó, P.; Bravo, I. Canary Islands (NE Atlantic) as a Biodiversity ‘Hotspot’ of Gambierdiscus: Implications for Future Trends of Ciguatera in the Area. Harmful Algae 2017, 67, 131–143. [Google Scholar] [CrossRef]
- Estevez, P.; Sibat, M.; Leão-Martins, J.M.; Reis Costa, P.; Gago-Martínez, A.; Hess, P. Liquid Chromatography Coupled to High-Resolution Mass Spectrometry for the Confirmation of Caribbean Ciguatoxin-1 as the Main Toxin Responsible for Ciguatera Poisoning Caused by Fish from European Atlantic Coasts. Toxins 2020, 12, 267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murata, M.; Yasumoto, T. The Structure Elucidation and Biological Activities of High Molecular Weight Algal Toxins: Maitotoxin, Prymnesins and Zooxanthellatoxins. Nat. Prod. Rep. 2000, 17, 293–314. [Google Scholar] [CrossRef] [PubMed]
- Kohli, G.S.; Papiol, G.G.; Rhodes, L.L.; Harwood, D.T.; Selwood, A.; Jerrett, A.; Murray, S.A.; Neilan, B.A. A Feeding Study to Probe the Uptake of Maitotoxin by Snapper (Pagrus auratus). Harmful Algae 2014, 37, 125–132. [Google Scholar] [CrossRef]
- Murata, M.; Naoki, H.; Matsunaga, S.; Satake, M.; Yasumoto, T. Structure and Partial Stereochemical Assignments for Maitotoxin, the Most Toxic and Largest Natural Non-Biopolymer. J. Am. Chem. Soc. 1994, 116, 7098–7107. [Google Scholar] [CrossRef]
- Holmes, M.J.; Lewis, R.J. Purification and Characterisation of Large and Small Maitotoxins from Cultured Gambierdiscus Toxicus. Nat. Toxins 1994, 2, 64–72. [Google Scholar] [CrossRef] [PubMed]
- Pisapia, F.; Sibat, M.; Herrenknecht, C.; Lhaute, K.; Gaiani, G.; Ferron, P.-J.; Fessard, V.; Fraga, S.; Nascimento, S.M.; Litaker, R.W.; et al. Maitotoxin-4, a Novel Mtx Analog Produced by Gambierdiscus Excentricus. Mar. Drugs 2017, 15, 220. [Google Scholar] [CrossRef] [PubMed]
- Mazzola, E.P.; Deeds, J.R.; Stutts, W.L.; Ridge, C.D.; Dickey, R.W.; White, K.D.; Williamson, R.T.; Martin, G.E. Elucidation and Partial NMR Assignment of Monosulfated Maitotoxins from the Caribbean. Toxicon 2019, 164, 44–50. [Google Scholar] [CrossRef]
- Lewis, R.J.; Holmes, M.J.; Alewood, P.F.; Jones, A. Lonspray Mass Spectrometry of Ciguatoxin-1, Maitotoxin-2 and -3, and Related Marine Polyether Toxins. Nat. Toxins 1994, 2, 56–63. [Google Scholar] [CrossRef]
- Murata, M.; Naoki, H.; Iwashita, T.; Matsunaga, S.; Sasaki, M.; Yokoyama, A.; Yasumoto, T. Structure of Maitotoxin. J. Am. Chem. Soc. 1993, 115, 2060–2062. [Google Scholar] [CrossRef]
- Murray, J.S.; Selwood, A.I.; Harwood, D.T.; Van Ginkel, R.; Puddick, J.; Rhodes, L.L.; Rise, F.; Wilkins, A.L. 44-Methylgambierone, a New Gambierone Analogue Isolated from Gambierdiscus Australes. Tetrahedron Lett. 2019, 60, 621–625. [Google Scholar] [CrossRef]
- Boente-Juncal, A.; Álvarez, M.; Antelo, Á.; Rodríguez, I.; Calabro, K.; Vale, C.; Thomas, O.P.; Botana, L.M. Structure Elucidation and Biological Evaluation of Maitotoxin-3, a Homologue Of Gambierone, from Gambierdiscus Belizeanus. Toxins 2019, 11, 79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, M.J.; Venables, B.; Lewis, R.J. Critical Review and Conceptual and Quantitative Models for the Transfer and Depuration of Ciguatoxins in Fishes. Toxins 2021, 13, 515. [Google Scholar] [CrossRef]
- Sanchez-Henao, J.A.; García-Álvarez, N.; Fernández, A.; Saavedra, P.; Sergent, F.S.; Padilla, D.; Acosta-Hernández, B.; Suárez, M.M.; Diogène, J.; Real, F. Predictive Score and Probability of CTX-Like Toxicity in Fish Samples from the Official Control of Ciguatera in the Canary Islands. Sci. Total Environ. 2019, 673, 576–584. [Google Scholar] [CrossRef]
- Sanchez-Henao, A.; García-Álvarez, N.; Sergent, F.S.; Estévez, P.; Gago-Martínez, A.; Martín, F.; Ramos-Sosa, M.; Fernández, A.; Diogène, J.; Real, F. Presence of Ctxs in Moray Eels and Dusky Groupers in the Marine Environment of the Canary Islands. Aquat. Toxicol. 2020, 221, 105427. [Google Scholar] [CrossRef] [PubMed]
- Fraga, S.; Rodríguez, F.; Caillaud, A.; Diogène, J.; Raho, N.; Zapata, M. Gambierdiscus excentricus sp. nov. (Dinophyceae), a Benthic Toxic Dinoflagellate from the Canary Islands (NE Atlantic Ocean). Harmful Algae 2011, 11, 10–22. [Google Scholar] [CrossRef] [Green Version]
- Reverté, L.; Toldrà, A.; Andree, K.B.; Fraga, S.; De Falco, G.; Campàs, M.; Diogène, J. Assessment of Cytotoxicity in Ten Strains of Gambierdiscus Australes from Macaronesian Islands by Neuro-2A Cell-Based Assays. J. Appl. Phycol. 2018, 30, 2447–2461. [Google Scholar] [CrossRef]
- Laza-Martinez, A.; David, H.; Riobó, P.; Miguel, I.; Orive, E. Characterization of a Strain of Fukuyoa paulensis (Dinophyceae) from the Western Mediterranean Sea. J. Eukaryot. Microbiol. 2015, 63, 481–497. [Google Scholar] [CrossRef] [Green Version]
- Bravo, I.; Rodriguez, F.; Ramilo, I.; Rial, P.; Fraga, S. Ciguatera-Causing Dinoflagellate Gambierdiscus spp. (Dinophyceae) In A Subtropical Region of North Atlantic Ocean (Canary Islands): Morphological Characterization and Biogeography. Toxins 2019, 11, 423. [Google Scholar] [CrossRef] [Green Version]
- Estevez, P.; Sibat, M.; Leão-Martins, J.M.; Tudó, A.; Rambla-Alegre, M.; Aligizaki, K.; Diogène, J.; Gago-Martinez, A.; Hess, P. Use of Mass Spectrometry to Determine the Diversity of Toxins Produced by Gambierdiscus and Fukuyoa Species from Balearic Islands and Crete (Mediterranean Sea) and the Canary Islands (Northeast Atlantic). Toxins 2020, 12, 305. [Google Scholar] [CrossRef]
- Tudó, À.; Toldrà, A.; Rey, M.; Todolí, I.; Andree, K.B.; Fernández-Tejedor, M.; Campàs, M.; Sureda, F.X.; Diogène, J. Gambierdiscus and Fukuyoa as Potential Indicators of Ciguatera Risk in the Balearic Islands. Harmful Algae 2020, 99, 101913. [Google Scholar] [CrossRef]
- Estevez, P.; Castro, D.; Pequeño-Valtierra, A.; Leao, J.M.; Vilariño, O.; Diogène, J.; Gago-Martínez, A. An Attempt To Characterize the Ciguatoxin Profile in Seriola Fasciata Causing Ciguatera Fish Poisoning in Macaronesia. Toxins 2019, 11, 221. [Google Scholar] [CrossRef] [Green Version]
- Estevez, P.; Castro, D.; Leao, J.M.; Yasumoto, T.; Dickey, R.; Gago-Martinez, A. Implementation of Liquid Chromatography Tandem Mass Spectrometry for the Analysis of Ciguatera Fish Poisoning in Contaminated Fish Samples from Atlantic Coasts. Food Chem. 2019, 280, 8–14. [Google Scholar] [CrossRef]
- Pisapia, F.; Sibat, M.; Watanabe, R.; Roullier, C.; Suzuki, T.; Hess, P.; Herrenknecht, C. Characterization of Maitotoxin-4 (MTX4) Using Electrospray Positive Mode Ionization High-Resolution Mass Spectrometry and UV Spectroscopy. Rapid Commun. Mass Spectrom. 2020, 34, E8859. [Google Scholar] [CrossRef]
- Patiny, L.; Borel, A. Chemcalc: A Building Block for Tomorrow’s Chemical Infrastructure. J. Chem. Inf. Model. 2013, 53, 1223–1228. [Google Scholar] [CrossRef] [Green Version]
- Lewis, R.J.; Inserra, M.; Vetter, I.; Holland, W.C.; Hardison, D.R.; Tester, P.A.; Litaker, R.W. Rapid Extraction and Identification of Maitotoxin and Ciguatoxin-Like Toxins from Caribbean and Pacific Gambierdiscus Using a New Functional Bioassay. PLoS ONE 2016, 11, e0160006. [Google Scholar] [CrossRef]
- Castro, D.; Manger, R.; Vilariño, O.; Gago-Martínez, A. Evaluation of Matrix Issues in the Applicability of the Neuro-2a Cell Based Assay on the Detection of CTX in Fish Samples. Toxins 2020, 12, 308. [Google Scholar] [CrossRef]
- Murray, J.S.; Boundy, M.J.; Selwood, A.I.; Harwood, D.T. Development of an LC-MS/MS Method To Simultaneously Monitor Maitotoxins and Selected Ciguatoxins in Algal Cultures and P-CTX-1B in Fish. Harmful Algae 2018, 80, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Moreiras, G.; Leão, J.M.; Gago-Martínez, A. Design of Experiments for the Optimization of Electrospray Ionization in the LC-MS/MS Analysis Of Ciguatoxins. J. Mass Spectrom. 2018, 53, 1059–1060. [Google Scholar] [CrossRef] [PubMed]
- Sibat, M.; Herrenknecht, C.; Darius, H.T.; Roué, M.; Chinain, M.; Hess, P. Detection of Pacific Ciguatoxins Using Liquid Chromatography Coupled to Either Low or High Resolution Mass Spectrometry (LC-MS/MS). J. Chromatogr. A 2018, 1571, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Yasumoto, T.; Igarashi, T.; Legrand, A.-M.; Cruchet, P.; Chinain, M.; Fujita, T.; Naoki, H. Structural Elucidation of Ciguatoxin Congeners by Fast-Atom Bombardment Tandem Mass Spectroscopy. J. Am. Chem. Soc. 2000, 122, 4988–4989. [Google Scholar] [CrossRef]
- Suzuki, T.; Ha, D.V.; Uesugi, A.; Uchida, H. Analytical Challenges to Ciguatoxins. Curr. Opin. Food Sci. 2017, 18, 37–42. [Google Scholar] [CrossRef]
- Estevez, P.; Leao, J.M.; Yasumoto, T.; Dickey, R.W.; Gago-Martinez, A. Caribbean Ciguatoxin-1 Stability Under Strongly Acidic Conditions: Characterisation of a New C-CTX1 Methoxy Congener. Food Addit. Contam. Part A 2020, 37, 519–529. [Google Scholar] [CrossRef]
- Nagai, H.; Torigoe, K.; Satake, M.; Murata, M.; Yasumoto, T.; Hirota, H. Gambieric Acids: Unprecedented Potent Antifungal Substances Isolated from Cultures of a Marine Dinoflagellate Gambierdiscus Toxicus. J. Am. Chem. Soc. 1992, 114, 1102–1103. [Google Scholar] [CrossRef]
- Nagai, H.; Mikami, Y.; Yazawa, K.; Gonoi, T.; Yasumoto, T. Biological Activities of Novel Polyether Antifungals, Gambieric Acids A and B from a Marine Dinoflagellate Gambierdiscus Toxicus. J. Antibiot. 1993, 46, 520–522. [Google Scholar] [CrossRef] [Green Version]
- Aecosan Website. Available online: http://www.aecosan.msssi.gob.es/aecosan/web/ciguatera/home/aecosan_home_ciguatera.htm (accessed on 25 March 2020).
- Satake, M.; Murata, M.; Yasumoto, T. The Structure of CTX3C, a Ciguatoxin Congener Isolated from Cultured Gambierdiscus Toxicus. Tetrahedron Lett. 1993, 34, 1975–1978. [Google Scholar] [CrossRef]
- Manger, R.L.; Leja, L.S.; Lee, S.Y.; Hungerford, J.M.; Hokama, Y.; Dickey, R.W.; Granade, H.R.; Lewis, R.; Yasumoto, T.; Wekell, M.M. Detection of Sodium Channel Toxins: Directed Cytotoxicity Assays of Purified Ciguatoxins, Brevetoxins, Saxitoxins, and Seafood Extracts. J. AOAC Int. 1995, 78, 521–527. [Google Scholar] [CrossRef]
- Mosmann, T. Rapid Colorimetric Assay for Cellular Growth and Survival: Application to Proliferation and Cytotoxicity Assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Manger, R.L.; Leja, L.S.; Lee, S.Y.; Hungerford, J.M.; Wekell, M.M. Tetrazolium-Based Cell Bioassay for Neurotoxins Active on Voltage-Sensitive Sodium Channels: Semiautomated Assay for Saxitoxins, Brevetoxins, and Ciguatoxins. Anal. Biochem. 1993, 214, 190–194. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
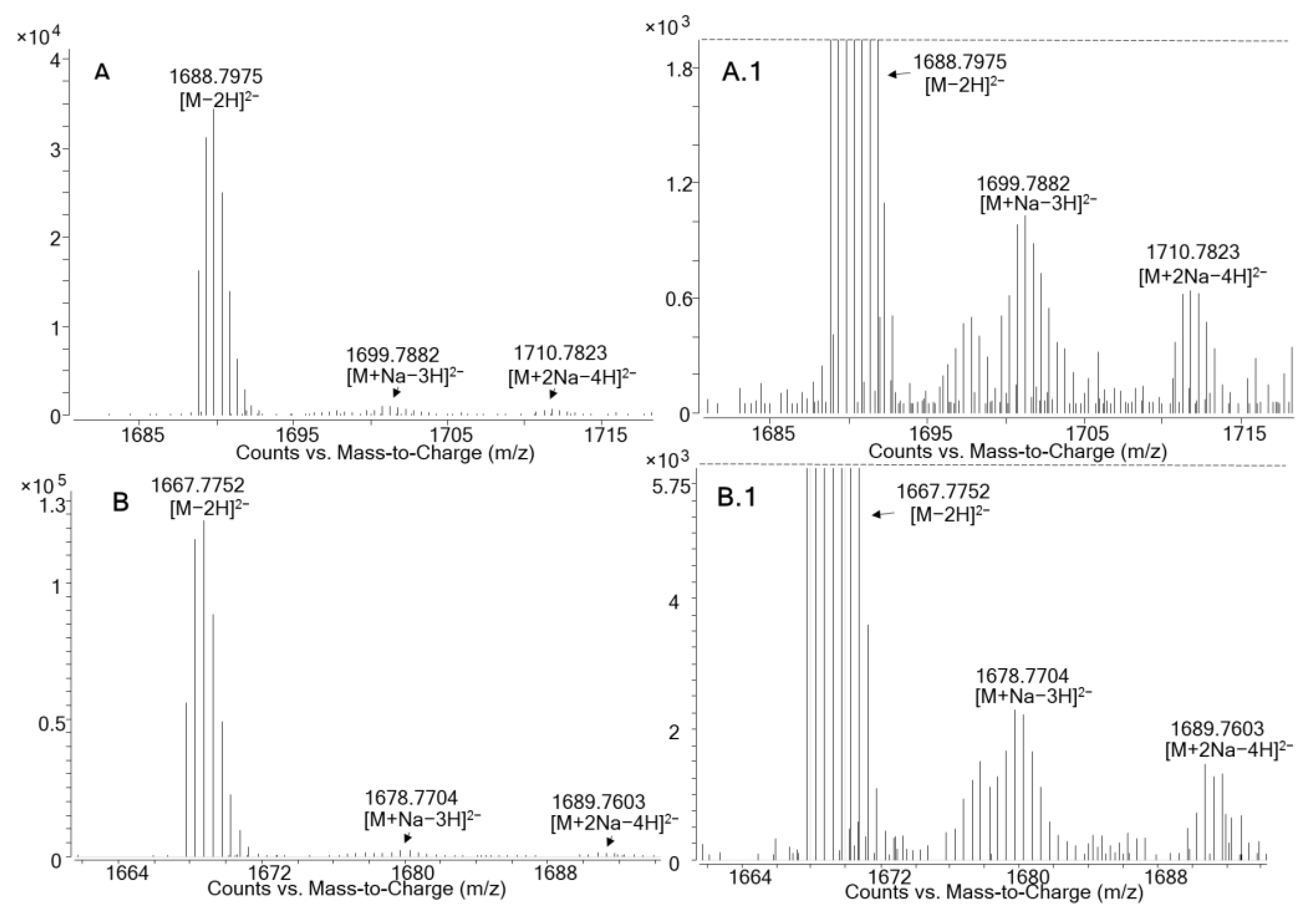
| MTX1 | New Maitotoxin Analogue | ||||
|---|---|---|---|---|---|
| Molecular Formula (M) | C164H258O68S2 | Unknown | |||
| Relative Molecular Weight (g/mol) | 3379.6172 | 3337.5649 | |||
| Retention Time (min) | 7.6 | 7.6 | |||
| Ion Species (Monoisotopic Mass) | Theoretical | Measured | Measured | ||
| ESI− | [M−2H]2− | 1688.8013 | 1688.7975 (Δppm: −2.3) | 1667.7752 | |
| [M+Na−3H]2− | 1699.7923 | 1699.7882 (Δppm: −2.4) | 1678.7704 | ||
| [M+2Na−4H]2− | 1710.7833 | 1710.7823(Δppm: −0.6) | 1689.7603 | ||
| [M−3H]3− | 1125.5318 | n.d. | n.d. | ||
| [M−4H]4− | 843.8970 | n.d. | n.d. | ||
| ESI+ | [M−2H+4NH4]2+ | 1724.8695 | 1724.8620 (Δppm: −4.4 ) | 1703.8470 | |
| [M−H+3NH4]2+ | 1716.3562 | 1716.3526 (Δppm: −2.1) | 1695.3340 | ||
| [M+2NH4]2+ | 1707.8430 | 1707.8396 (Δppm: −2.0) | 1686.8206 | ||
| [M+H+NH4]2+ | 1699.3297 | 1699.3276 (Δppm: −1.2) | 1678.3090 | ||
| Compound | Retention Time (min) | ESI | MRM Transitions Q1/Q3 (m/z) | CE (eV) | CAV (eV) | |
|---|---|---|---|---|---|---|
| MTX5 | 9.05 | - | [M−2H]2−/[M−2H]2− | 1668.8/1668.8 | 30 | 5 |
| [M−2H]2−/[HOSO3]− | 1668.8/96.9 | 60 | 5 | |||
| [M−3H]3−/[M−3H]3− | 1112.6/1112.6 | 30 | 5 | |||
| [M−3H]3−/[HOSO3]− | 1112.6/96.9 | 60 | 5 | |||
| [M+Na−3H]2− /[HOSO3]− | 1679.8/96.9 | 60 | 5 | |||
| [M+2Na−4H]2− /[HOSO3]− | 1690.8 /96.9 | 60 | 5 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Estevez, P.; Castro, D.; Leão-Martins, J.M.; Sibat, M.; Tudó, A.; Dickey, R.; Diogene, J.; Hess, P.; Gago-Martinez, A. Toxicity Screening of a Gambierdiscus australes Strain from the Western Mediterranean Sea and Identification of a Novel Maitotoxin Analogue. Mar. Drugs 2021, 19, 460. https://doi.org/10.3390/md19080460
Estevez P, Castro D, Leão-Martins JM, Sibat M, Tudó A, Dickey R, Diogene J, Hess P, Gago-Martinez A. Toxicity Screening of a Gambierdiscus australes Strain from the Western Mediterranean Sea and Identification of a Novel Maitotoxin Analogue. Marine Drugs. 2021; 19(8):460. https://doi.org/10.3390/md19080460
Chicago/Turabian StyleEstevez, Pablo, David Castro, José Manuel Leão-Martins, Manoëlla Sibat, Angels Tudó, Robert Dickey, Jorge Diogene, Philipp Hess, and Ana Gago-Martinez. 2021. "Toxicity Screening of a Gambierdiscus australes Strain from the Western Mediterranean Sea and Identification of a Novel Maitotoxin Analogue" Marine Drugs 19, no. 8: 460. https://doi.org/10.3390/md19080460
APA StyleEstevez, P., Castro, D., Leão-Martins, J. M., Sibat, M., Tudó, A., Dickey, R., Diogene, J., Hess, P., & Gago-Martinez, A. (2021). Toxicity Screening of a Gambierdiscus australes Strain from the Western Mediterranean Sea and Identification of a Novel Maitotoxin Analogue. Marine Drugs, 19(8), 460. https://doi.org/10.3390/md19080460