
Elemental Composition and Cell Mass Quantification of Cultured Thraustochytrids Unveil Their Large Contribution to Marine Carbon Pool






Abstract
:1. Introduction
2. Results
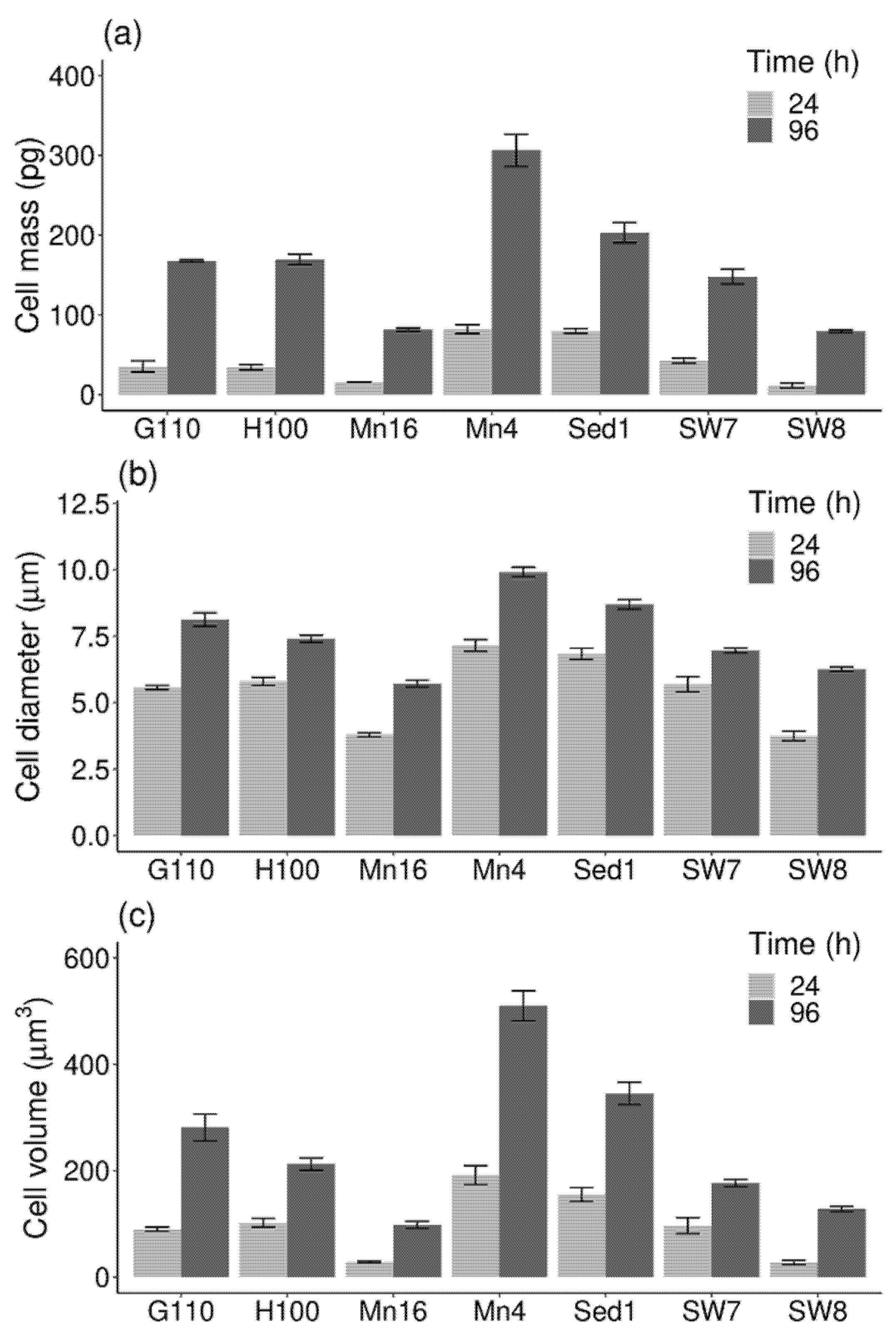
2.1. Cell Mass and Volume
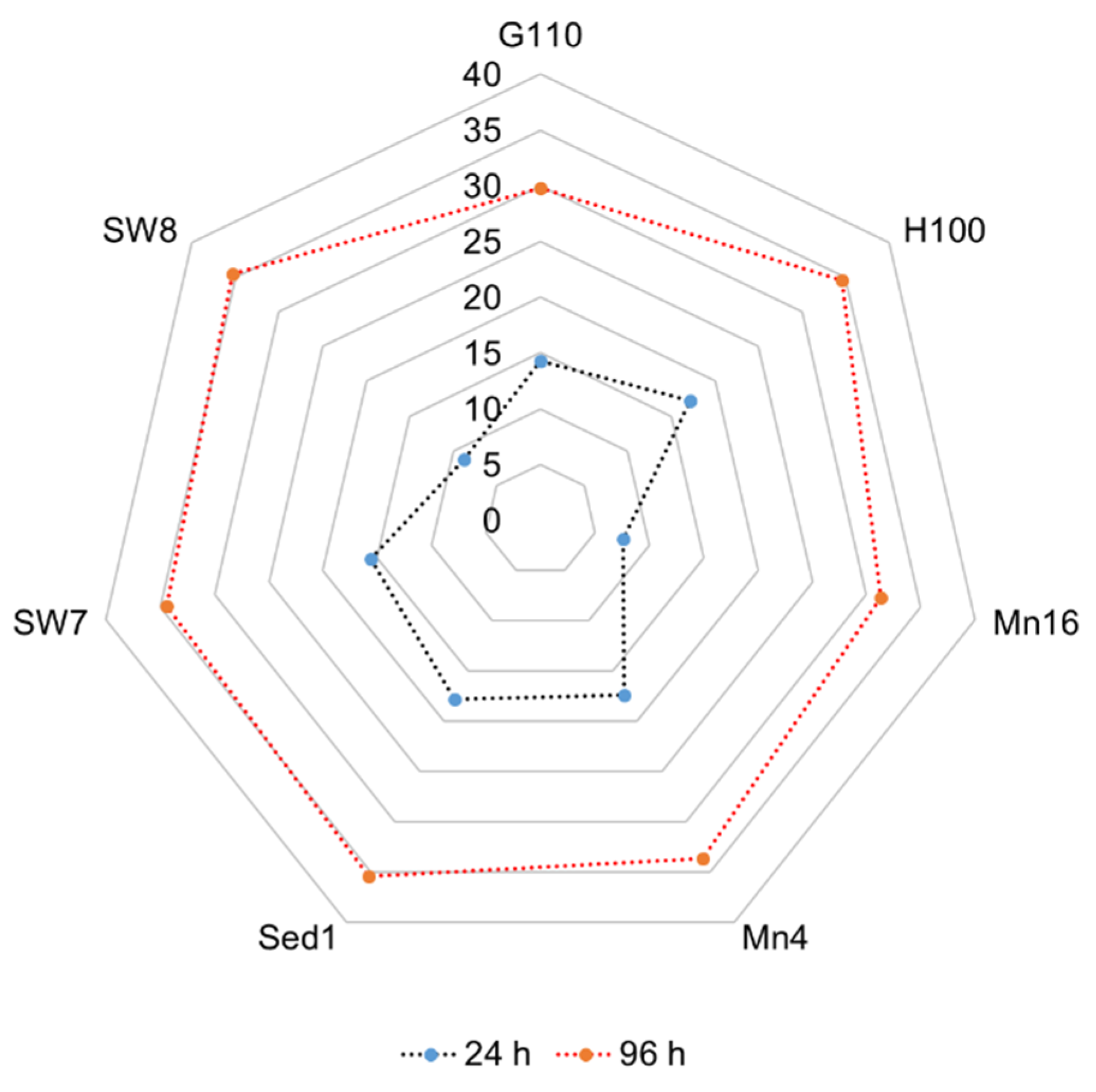
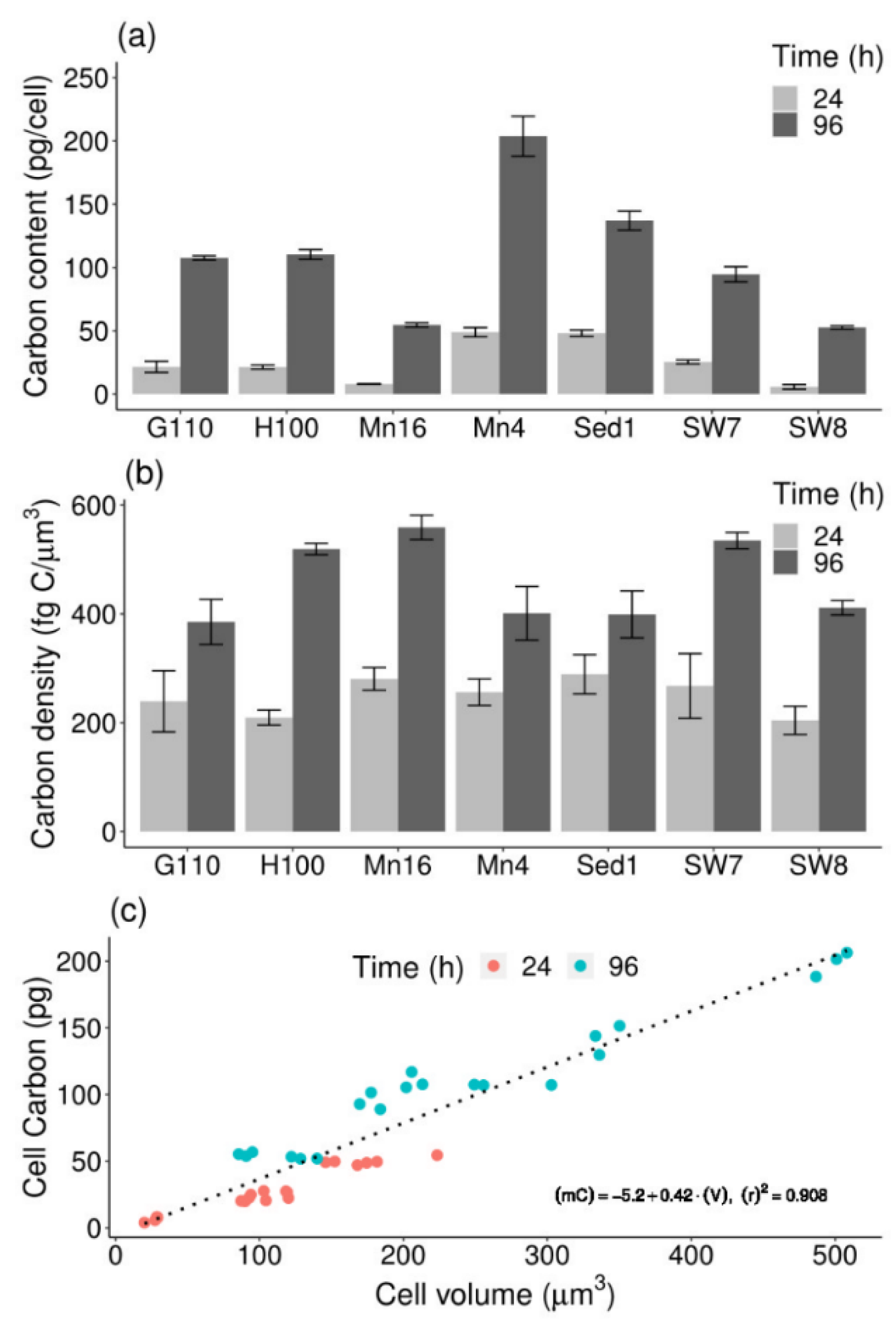
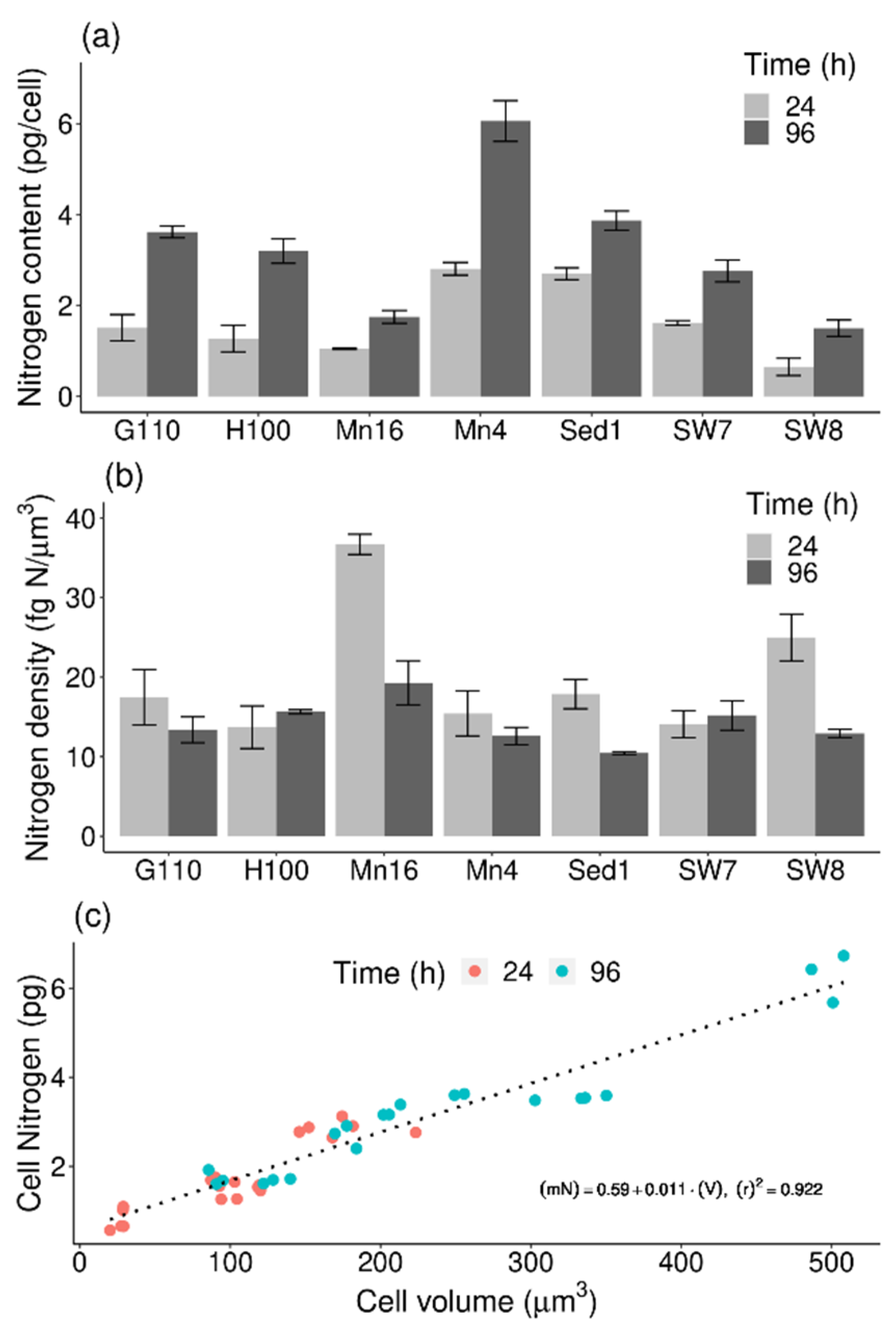
2.2. Elemental Composition
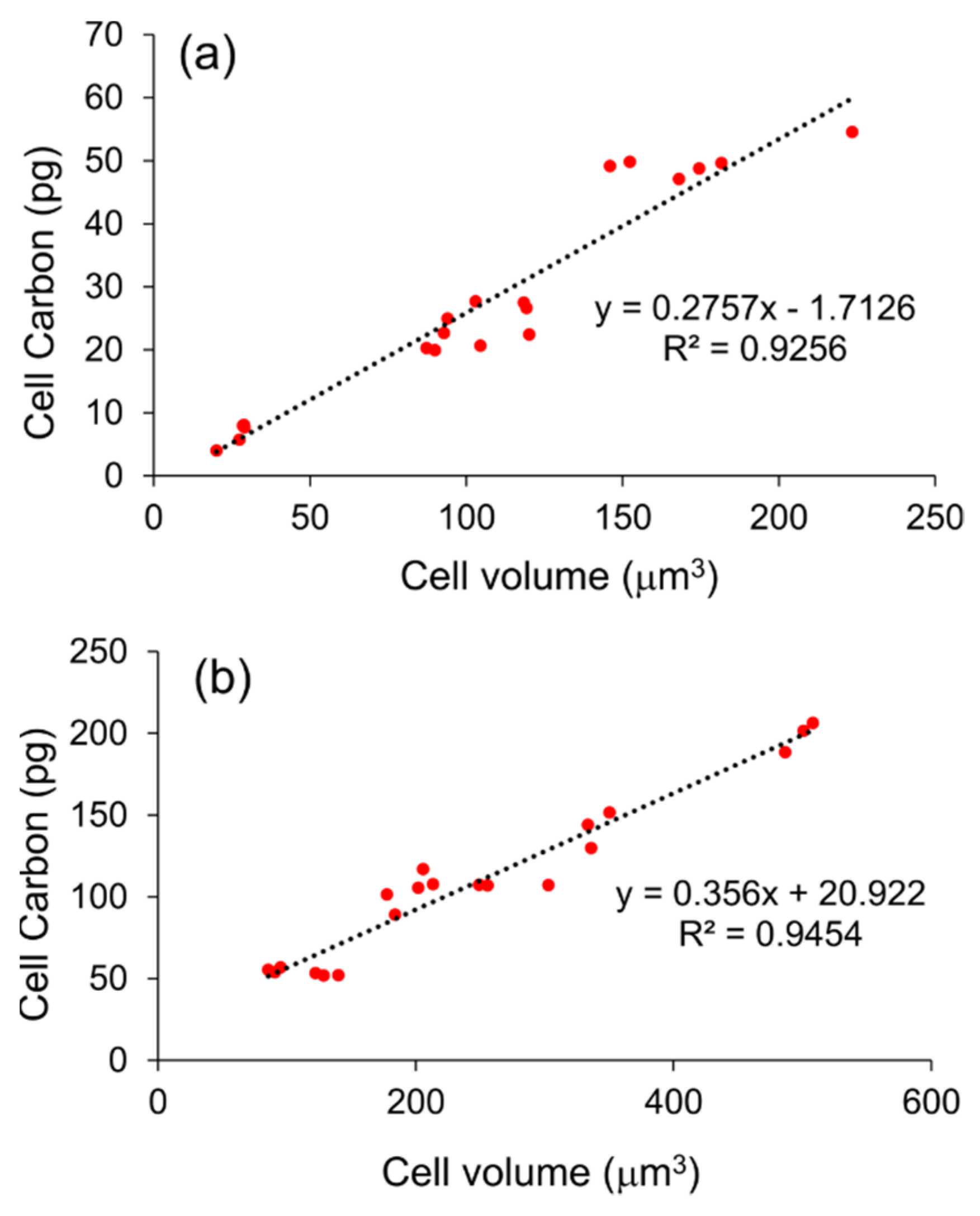
2.3. Cell Carbon-to-Volume Relationship
2.4. Cell Nitrogen-to-Volume Relationship
3. Discussion
4. Materials and Methods
4.1. Isolates and Culture Conditions
4.2. Analysis of Elemental Composition
4.3. Determination of Cell Count, Diameter, and Volume
4.4. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Bennett, R.M.; Honda, D.; Beakes, G.W.; Thines, M. Labyrinthulomycota. In Handbook of the Protists; Archibald, J.M., Simpson, A.G.B., Slamovits, C.H., Margulis, L., Melkonian, M., Chapman, D.J., Corliss, J.O., et al., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–36. [Google Scholar]
- Raghukumar, S. Ecology of the marine protists, the labyrinthulomycetes (thraustochytrids and labyrinthulids). Eur. J. Protistol. 2002, 38, 127–145. [Google Scholar] [CrossRef]
- Raghukumar, S. Speculations on niches occupied by fungi in the sea with relation to bacteria. Proc. Plant Sci. 1990, 100, 129–138. [Google Scholar]
- Fell, J.W.; Newell, S.Y. Biochemical and Molecular Methods for the Study of Marine Fungi. In Molecular Approaches to the Study of the Ocean; Cooksey, K.E., Ed.; Springer: Dordrecht, The Netherlands, 1998; pp. 259–283. [Google Scholar]
- Damare, V.; Raghukumar, S. Abundance of thraustochytrids and bacteria in the equatorial Indian Ocean, in relation to transparent exopolymeric particles (TEPs). FEMS Microbiol. Ecol. 2008, 65, 40–49. [Google Scholar] [CrossRef] [Green Version]
- Raghukumar, S.; Ramaiah, N.; Raghukumar, C. Dynamics of thraustochytrid protists in the water column of the Arabian sea. Aquat. Microb. Ecol. 2001, 24, 175–186. [Google Scholar] [CrossRef] [Green Version]
- Xie, N.; Sen, B.; Song, Z.; Zhao, Y.; Chen, Z.; Shi, W.; Zhang, Y.; Zhang, J.; Johnson, Z.I.; Wang, G. High phylogenetic diversity and abundance pattern of Labyrinthulomycete protists in the coastal waters of the Bohai Sea. Environ. Microbiol. 2018, 20, 3042–3056. [Google Scholar] [CrossRef] [PubMed]
- Bongiorni, L.; Mirto, S.; Pusceddu, A.; Danovaro, R. Response of benthic protozoa and thraustochytrid protists to fish farm impact in seagrass (Posidonia oceanica) and soft-bottom sediments. Microb. Ecol. 2005, 50, 268–276. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wang, X.; Liu, X.; Jiao, N.; Wang, G. Abundance and Novel Lineages of Thraustochytrids in Hawaiian Waters. Microb. Ecol. 2013, 66, 823. [Google Scholar] [CrossRef]
- Liu, Y.; Singh, P.; Liang, Y.; Li, J.; Xie, N.; Song, Z.; Daroch, M.; Leng, K.; Johnson, Z.I.; Wang, G. Abundance and Molecular Diversity of Thraustochytrids in Coastal Waters of Southern China. FEMS Microbiol. Ecol. 2017, 93, fix070. [Google Scholar] [CrossRef] [PubMed]
- Naganuma, T.; Takasugi, H.; Kimura, H. Abundance of thraustochytrids in coastal plankton. Mar. Ecol. Prog. Ser. 1998, 162, 105–110. [Google Scholar] [CrossRef] [Green Version]
- Kimura, H.; Fukuba, T.; Naganuma, T. Biomass of thraustochytrid protoctists in coastal water. Mar. Ecol. Prog. Ser. 1999, 189, 27–33. [Google Scholar] [CrossRef]
- Liu, X.; Sen, B.; Zhao, Y.; Bai, M.; He, Y.; Xie, Y.; Li, J.; Wang, G. Gradients of three coastal environments off the South China Sea and their impacts on the dynamics of heterotrophic microbial communities. Sci. Total Environ. 2019, 659, 499–506. [Google Scholar] [CrossRef] [PubMed]
- Ramaiah, N.; Raghukumar, S.; Mangesh, G.; Madhupratap, M. Seasonal variations in carbon biomass of bacteria, thraustochytrids and microzooplankton in the Northern Arabian Sea. Deep-Sea Res. Part II Top. Stud. Oceanogr. 2005, 52, 1910–1921. [Google Scholar] [CrossRef]
- Duan, Y.; Sen, B.; Xie, N.; Paterson, J.S.; Chen, Z.; Wang, G. Flow cytometry for rapid enumeration and biomass quantification of thraustochytrids in coastal seawaters. Microbes Environ. 2018, 33, 195–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bochdansky, A.B.; Clouse, M.A.; Herndl, G.J. Eukaryotic microbes, principally fungi and labyrinthulomycetes, dominate biomass on bathypelagic marine snow. ISME J. 2017, 11, 362–373. [Google Scholar] [CrossRef]
- Raghukumar, S. Thraustochytrid marine protists: Production of PUFAs and other emerging technologies. Mar. Biotechnol. 2008, 10, 631–640. [Google Scholar] [CrossRef]
- Aasen, I.M.; Ertesvåg, H.; Heggeset, T.M.B.; Liu, B.; Brautaset, T.; Vadstein, O.; Ellingsen, T.E. Thraustochytrids as production organisms for docosahexaenoic acid (DHA), squalene, and carotenoids. Appl. Microbiol. Biotechnol. 2016, 100, 4309–4321. [Google Scholar] [CrossRef]
- Byreddy, A.R. Thraustochytrids as an alternative source of omega-3 fatty acids, carotenoids and enzymes. Lipid Technol. 2016, 28, 68–70. [Google Scholar] [CrossRef]
- Bongiorni, L.; Pignataro, L.; Santangelo, G. Thraustochytrids (fungoid protists): An unexplored component of marine sediment microbiota. Sci. Mar. 2004, 68 (Suppl. 1), 43–48. [Google Scholar]
- Damare, V.S.; Raghukumar, S. Apparent grazing losses of Labyrinthulomycetes protists in oceanic and coastal waters: An experimental elucidation. Ecol. Res. 2015, 30, 403–414. [Google Scholar] [CrossRef]
- Raghukumar, S.; Damare, V.S. Increasing evidence for the important role of Labyrinthulomycetes in marine ecosystems. Bot. Mar. 2011, 54, 3–11. [Google Scholar] [CrossRef] [Green Version]
- Fagerbakke, K.M.; Heldal, M.; Norland, S. Content of carbon, nitrogen, oxygen, sulfur and phosphorus in native aquatic and cultured bacteria. Aquat. Microb. Ecol. 1996, 10, 15–27. [Google Scholar] [CrossRef]
- Egli, T.; Zinn, M. The concept of multiple-nutrient-limited growth of microorganisms and its application in biotechnological processes. Biotechnol. Adv. 2003, 22, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Maureaud, A.; Gascuel, D.; Colléter, M.; Palomares, M.L.D.; Du Pontavice, H.; Pauly, D.; Cheung, W.W.L. Global change in the trophic functioning of marine food webs. PLoS ONE 2017, 12, e0182826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Honda, D.; Yokochi, T.; Nakahara, T.; Erata, M.; Higashihara, T. Schizochytrium limacinum sp. nov., a new thraustochytrid from a mangrove area in the west Pacific Ocean. Mycol. Res. 1998, 102, 439–448. [Google Scholar] [CrossRef]
- Bagul, V.P.; Annapure, U.S. Isolation and characterization of docosahexaenoic acid-producing novel strain Aurantiochytrium sp. ICTFD5: A sterol with vitamin d-cholecalciferol, and cellulase and lipase producing thraustochytrid. Biores. Technol. Rep. 2021, 14, 100688. [Google Scholar]
- Caamaño, E.; Loperena, L.; Hinzpeter, I.; Pradel, P.; Gordillo, F.; Corsini, G.; Tello, M.; Lavín, P.; González, A.R. Isolation and molecular characterization of Thraustochytrium strain isolated from Antarctic Peninsula and its biotechnological potential in the production of fatty acids. Braz. J. Microbiol. 2017, 48, 671–679. [Google Scholar] [CrossRef] [PubMed]
- Raghukumar, S.; Schaumann, K. An Epifluorescence Microscopy Method for Direct Detection and Enumeration of the Fungi-Like Marine Protists, the Thraustochytrids. Limnol. Oceanogr. 1993, 38, 182–187. [Google Scholar]
- Liu, Y.; Singh, P.; Sun, Y.; Luan, S.; Wang, G. Culturable Diversity and Biochemical Features of Thraustochytrids from Coastal Waters of Southern China. Appl. Microbiol. Biotechnol. 2014, 98, 3241–3255. [Google Scholar] [CrossRef]
- Wang, Q.; Ye, H.; Xie, Y.; He, Y.; Sen, B.; Wang, G. Culturable Diversity and Lipid Production Profile of Labyrinthulomycete Protists Isolated from Coastal Mangrove Habitats of China. Mar. Drugs 2019, 17, 268. [Google Scholar] [CrossRef] [Green Version]
- Grob, C.; Ostrowski, M.; Holland, R.J.; Heldal, M.; Norland, S.; Erichsen, E.S.; Blindauer, C.; Martin, A.P.; Zubkov, M.V.; Scanlan, D.J. Elemental composition of natural populations of key microbial groups in Atlantic waters. Environ. Microbiol. 2013, 15, 3054–3064. [Google Scholar] [CrossRef] [Green Version]
- Vrede, K.; Heldal, M.; Norland, S.; Bratbak, G. Elemental composition (C, N, P) and cell volume of exponentially growing and nutrient-limited bacterioplankton. Appl. Environ. Microbiol. 2002, 68, 2965–2971. [Google Scholar] [CrossRef] [Green Version]
- Fukuda, R.; Ogawa, H.; Nagata, T.; Koike, I. Direct determination of carbon and nitrogen contents of natural bacterial assemblages in marine environments. Appl. Environ. Microbiol. 1998, 64, 3352–3358. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gefen, O.; Fridman, O.; Ronin, I.; Balaban, N.Q. Direct observation of single stationary-phase bacteria reveals a surprisingly long period of constant protein production activity. Proc. Natl. Acad. Sci. USA 2014, 111, 556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaishankar, J.; Srivastava, P. Molecular Basis of Stationary Phase Survival and Applications. Front. Microbiol. 2017, 8, 2000. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Growth Phase | Isolate | C (%) | N (%) | H (%) | S (%) | C:N:H:S |
|---|---|---|---|---|---|---|
| Exponential (24 h) | G110 | 61.10 ± 0.81 | 4.29 ± 0.05 | 9.28 ± 0.09 | 1.00 ± 0.02 | 61:4:9:1 |
| H100 | 62.43 ± 1.42 | 3.69 ± 0.50 | 9.47 ± 0.19 | 0.91 ± 0.03 | 62:4:9:1 | |
| Mn16 | 51.23 ± 0.17 | 6.73 ± 0.16 | 7.90 ± 0.03 | 1.21 ± 0.09 | 51:7:8:1 | |
| Mn4 | 59.56 ± 0.44 | 3.42 ± 0.06 | 8.57 ± 0.52 | 0.95 ± 0.19 | 60:3:9:1 | |
| Sed1 | 60.39 ± 0.05 | 3.38 ± 0.06 | 9.13 ± 0.01 | 1.10 ± 0.05 | 60:3:9:1 | |
| SW7 | 59.64 ± 1.47 | 3.82 ± 0.24 | 9.01 ± 0.20 | 1.09 ± 0.00 | 60:4:9:1 | |
| SW8 | 50.27 ± 1.02 | 5.71 ± 0.16 | 7.86 ± 0.12 | 1.82 ± 0.07 | 50:6:8:2 | |
| Stationary (96 h) | G110 | 64.28 ± 0.43 | 2.16 ± 0.10 | 13.90 ±0.22 | 0.34 ± 0.01 | 640:20:140:3 |
| H100 | 65.05 ± 0.27 | 1.89 ± 0.10 | 13.24 ±0.22 | 0.36 ± 0.01 | 325:10:65:2 | |
| Mn16 | 66.94 ± 1.07 | 2.14 ± 0.12 | 10.57 ±0.22 | 0.46 ± 0.01 | 134:4:22:1 | |
| Mn4 | 66.46 ± 0.92 | 1.98 ± 0.05 | 9.90 ±0.34 | 0.49 ± 0.01 | 132:4:20:1 | |
| Sed1 | 67.50 ± 0.90 | 1.91 ± 0.08 | 11.23 ±0.22 | 0.44 ± 0.01 | 340:10:55:2 | |
| SW7 | 64.01 ± 0.20 | 1.86 ± 0.05 | 11.90 ±0.22 | 0.41 ± 0.01 | 160:5:30:1 | |
| SW8 | 66.57 ± 0.65 | 1.90 ± 0.24 | 9.93 ±0.07 | 0.91 ± 0.07 | 670:20:100:9 |
| Isolate | Isolation Source | GenBank Accession | % Homology with Type Strain # |
|---|---|---|---|
| GXBH-110 (G110) | Mangrove leaves | MG429124.1 | 99.56 |
| HNHK-100 (H100) | Mangrove leaves | MG429118.1 | 99.18 |
| PKU#Mn16 (Mn16) | Mangrove leaves | JX847368.1 | 97.83 |
| PKU#Mn4 (Mn4) | Mangrove leaves | JX847360.1 | 98.80 |
| PKU#Sed1 (Sed1) | Sediment | JX847370.1 | 98.86 |
| PKU#SW7 (SW7) | Seawater | JX847377.1 | 98.75 |
| PKU#SW8 (SW8) | Seawater | JX847378.1 | 98.70 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sen, B.; Li, J.; Lu, L.; Bai, M.; He, Y.; Wang, G. Elemental Composition and Cell Mass Quantification of Cultured Thraustochytrids Unveil Their Large Contribution to Marine Carbon Pool. Mar. Drugs 2021, 19, 493. https://doi.org/10.3390/md19090493
Sen B, Li J, Lu L, Bai M, He Y, Wang G. Elemental Composition and Cell Mass Quantification of Cultured Thraustochytrids Unveil Their Large Contribution to Marine Carbon Pool. Marine Drugs. 2021; 19(9):493. https://doi.org/10.3390/md19090493
Chicago/Turabian StyleSen, Biswarup, Jiaqian Li, Lyu Lu, Mohan Bai, Yaodong He, and Guangyi Wang. 2021. "Elemental Composition and Cell Mass Quantification of Cultured Thraustochytrids Unveil Their Large Contribution to Marine Carbon Pool" Marine Drugs 19, no. 9: 493. https://doi.org/10.3390/md19090493
APA StyleSen, B., Li, J., Lu, L., Bai, M., He, Y., & Wang, G. (2021). Elemental Composition and Cell Mass Quantification of Cultured Thraustochytrids Unveil Their Large Contribution to Marine Carbon Pool. Marine Drugs, 19(9), 493. https://doi.org/10.3390/md19090493