
Structure of the 4-O-[1-Carboxyethyl]-d-Mannose-Containing O-Specific Polysaccharide of a Halophilic Bacterium Salinivibrio sp. EG9S8QL


 ,
,  ,
,
Abstract
:1. Introduction
2. Results
2.1. Strain Isolation and Identification
2.2. LPS Isolation and Characterization
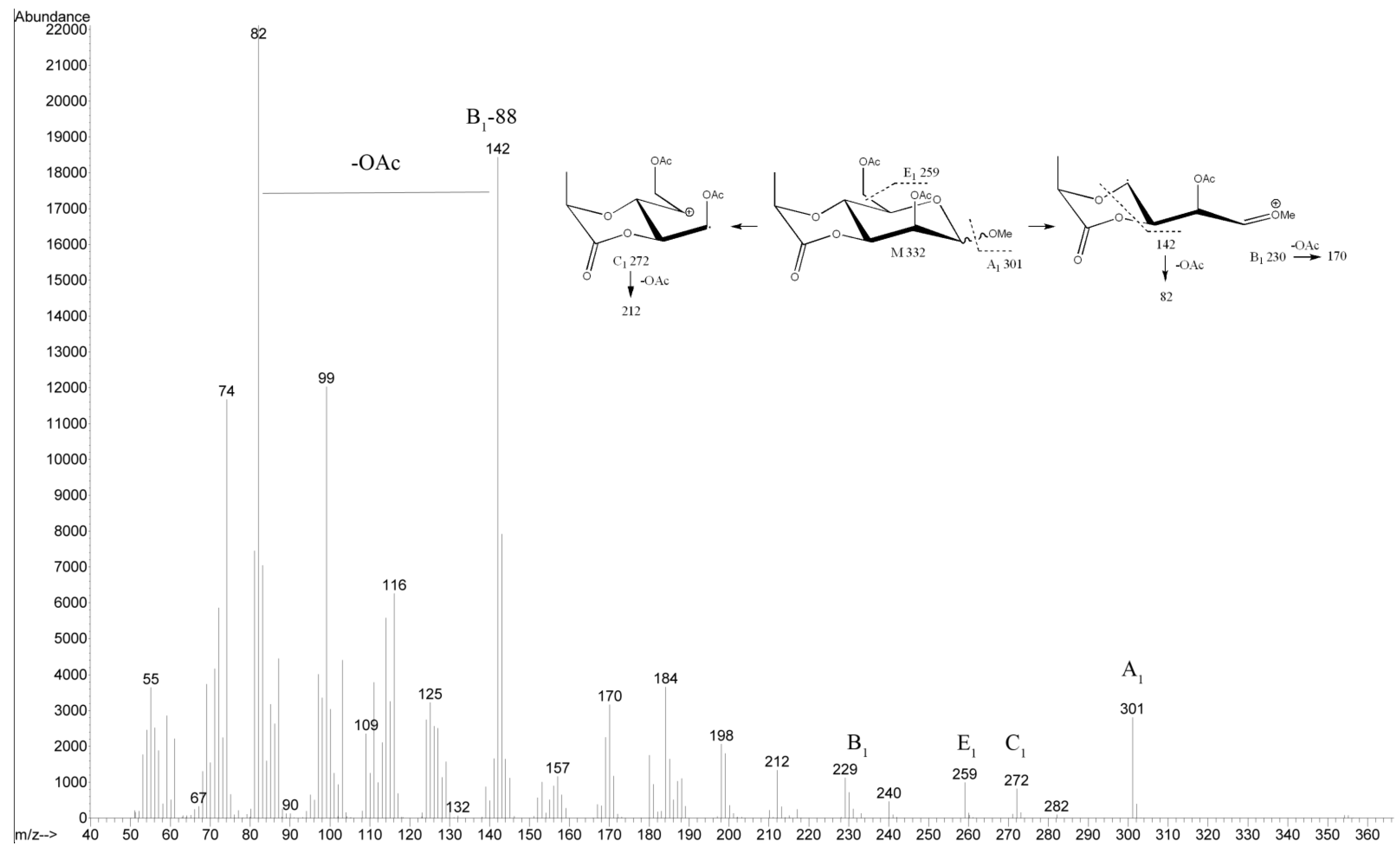
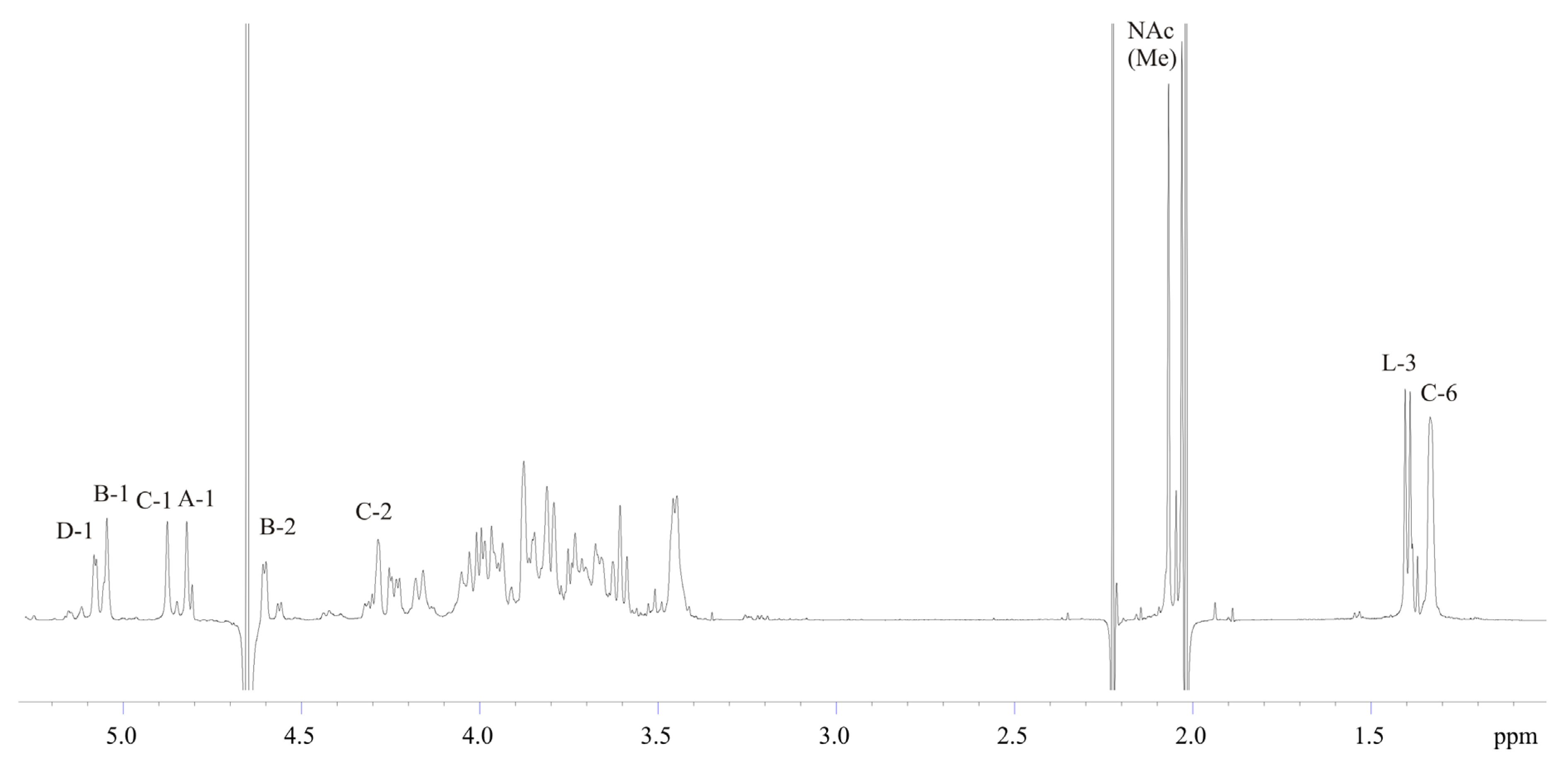
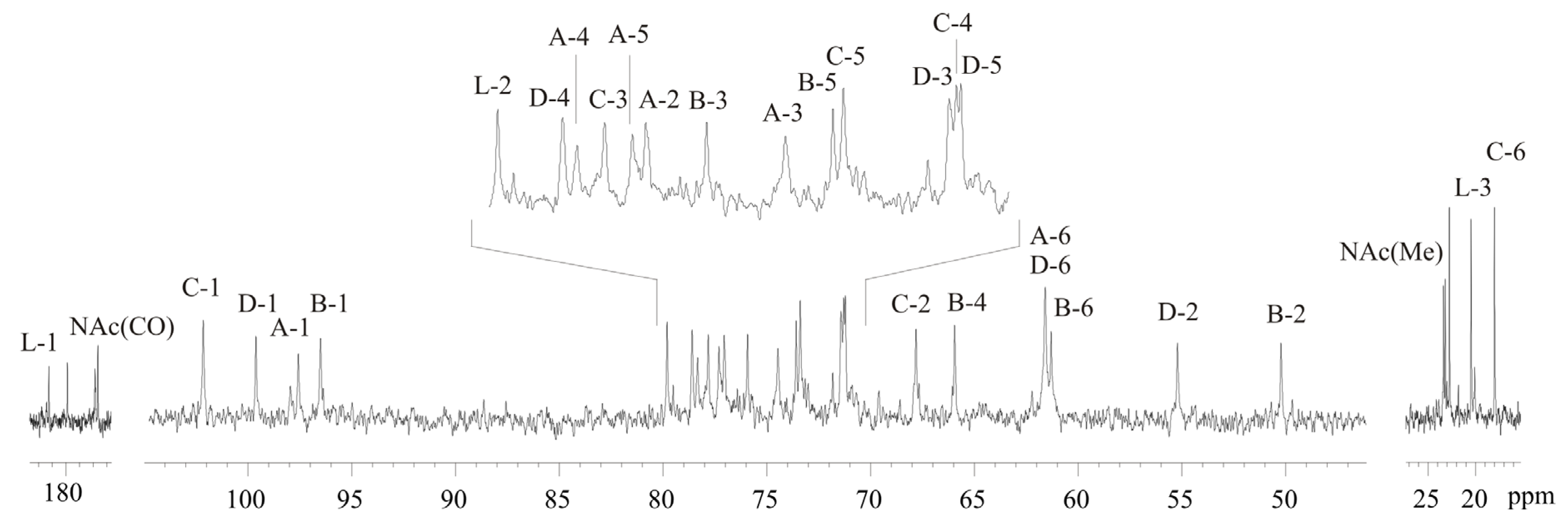
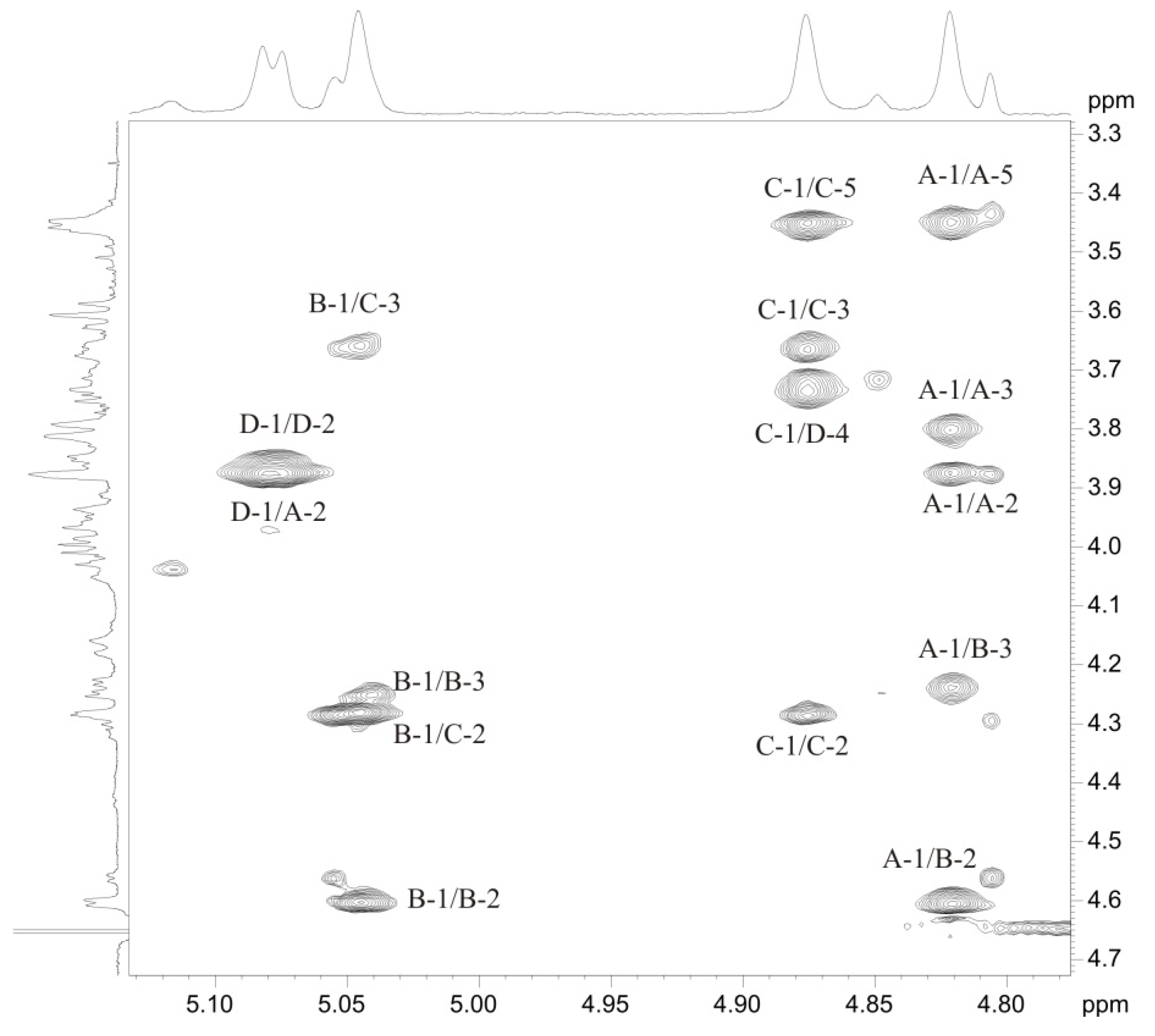
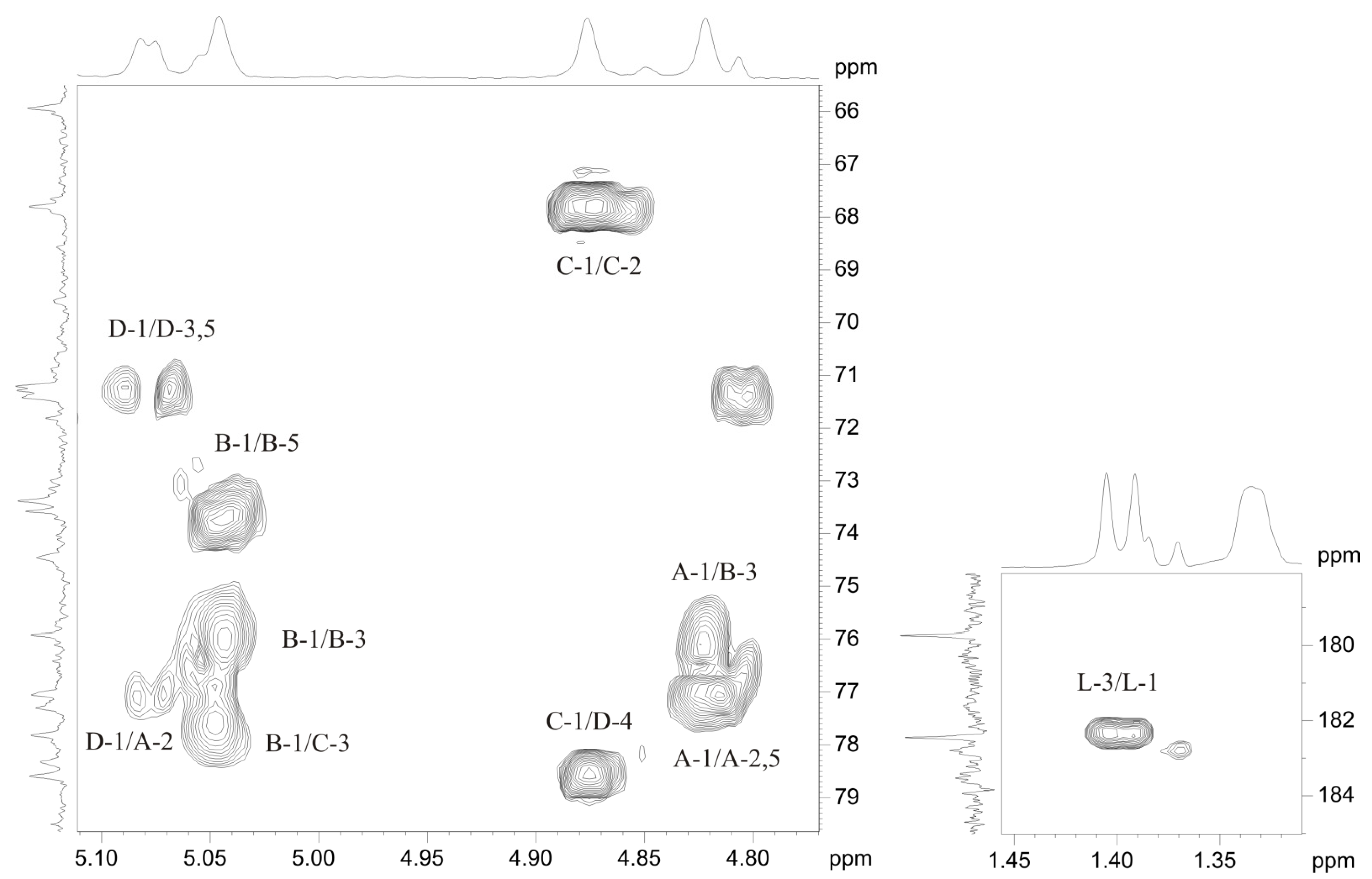
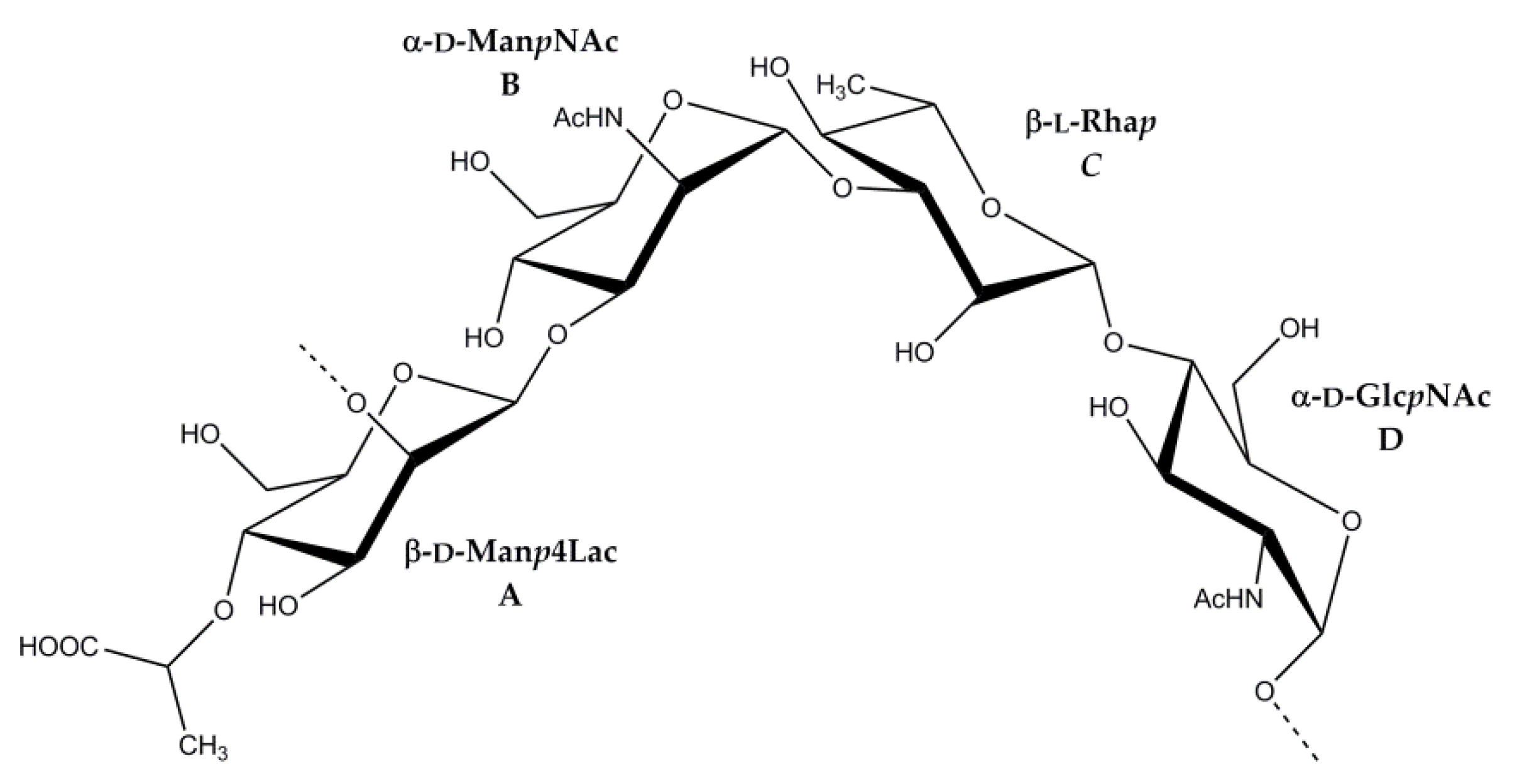
2.3. Structural Analysis of the OPS
3. Discussion
4. Materials and Methods
4.1. Sampling and Strain Isolation
4.2. Phenotypic and Genotypic Characterization of Strain EG9S8QL
4.3. Isolation of the Lipopolysaccharide and O-Specific Polysaccharide
4.4. Monosaccharide Analyses
4.5. NMR Spectroscopy
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Oren, A. Halophilic microbial communities and their environments. Curr. Opin. Biotechnol. 2015, 33, 119–124. [Google Scholar] [CrossRef] [PubMed]
- Perriss, S.J.; Laybourn-Parry, J. Microbial communities in saline lakes of the Vestfold Hills (eastern Antarctica). Polar Biol. 1997, 18, 135–144. [Google Scholar] [CrossRef]
- Hahn, M.W. The microbial diversity of inland waters. Curr. Opin. Biotechnol. 2006, 17, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Dillon, J.G.; McMath, L.M.; Trout, A.L. Seasonal changes in bacterial diversity in the Salton Sea. Hydrobiologia 2009, 632, 49–64. [Google Scholar] [CrossRef]
- Fernández-Delgado, M.; Suárez, P.; Giner, S.; Sanz, V.; Peña, J.; Sánchez, D.; García-Amado, M.A. Occurrence and virulence properties of Vibrio and Salinivibrio isolates from tropical lagoons of the southern Caribbean Sea. Antonie Leeuwenhoek 2017, 110, 833–841. [Google Scholar] [CrossRef]
- Zhang, H.; Zheng, S.; Ding, J.; Wang, O.; Liu, F. Spatial variation in bacterial community in natural wetland-river-sea ecosystems. J. Basic. Microbiol. 2017, 57, 536–546. [Google Scholar] [CrossRef]
- Huang, C.Y.; Garcia, J.L.; Patel, B.K.; Cayol, J.L.; Baresi, L.; Mah, R.A. Salinivibrio costicola subsp. vallismortis subsp. nov., a halotolerant facultative anaerobe from Death Valley, and emended description of Salinivibrio costicola. Int. J. Syst. Evol. Microbiol. 2000, 50, 615–622. [Google Scholar] [CrossRef] [Green Version]
- Lopez-Hermoso, C.; de la Haba, R.R.; Sanchez-Porro, C.; Ventosa, A. Salinivibrio kushneri sp. nov., a moderately halophilic bacterium isolated from salterns. Syst. Appl. Microbiol. 2018, 41, 159–166. [Google Scholar] [CrossRef]
- Chamroensaksri, N.; Tanasupawat, S.; Akaracharanya, A.; Visessanguan, W.; Kudo, T.; Itoh, T. Salinivibrio siamensis sp. nov., from fermented fish (pla-ra) in Thailand. Int. J. Syst. Evol. Microbiol. 2009, 59, 880–885. [Google Scholar] [CrossRef] [Green Version]
- Galisteo, C.; Sanchez-Porro, C.; de la Haba, R.R.; Lopez-Hermoso, C.; Fernandez, A.B.; Farias, M.E.; Ventosa, A. Characterization of Salinivibrio socompensis sp. nov., a new halophilic bacterium isolated from the high-altitude hypersaline lake Socompa, Argentina. Microorganisms 2019, 7, 241. [Google Scholar] [CrossRef] [Green Version]
- Genus Salinivibrio. 1996. Available online: https://bacterio.net/genus/salinivibrio (accessed on 5 August 2021).
- John, J.; Siva, V.; Richa, K.; Arya, A.; Kumar, A. Life in high salt concentrations with changing environmental conditions: Insights from genomic and phenotypic analysis of Salinivibrio sp. Microorganisms 2019, 7, 577. [Google Scholar] [CrossRef] [Green Version]
- John, J.; Dineshram, R.; Hemalatha, K.R.; Dhassiah, M.P.; Gopal, D.; Kumar, A. Bio-decolorization of synthetic dyes by a halophilic bacterium Salinivibrio sp. Front. Microbiol. 2020, 11, 594011. [Google Scholar] [CrossRef]
- Tao, G.B.; Tan, B.W.; Li, Z.J. Production of polyhydroxyalkanoates by a moderately halophilic bacterium of Salinivibrio sp. TGB10. Int. J. Biol. Macromol. 2021, 186, 574–579. [Google Scholar] [CrossRef]
- Van Thuoc, D.; Loan, T.T.; Tra, N.T. Accumulation of ectoines by halophilic bacteria isolated from fermented shrimp paste: An adaptation mechanism to salinity, temperature, and Ph stress. Curr. Microbiol. 2021, 78, 2355–2366. [Google Scholar] [CrossRef]
- Ventosa, A.; Nieto, J.J.; Oren, A. Biology of moderately halophilic aerobic bacteria. Microbiol. Mol. Biol. Rev. 1998, 62, 504–544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunde-Cimerman, N.; Plemenitas, A.; Oren, A. Strategies of adaptation of microorganisms of the three domains of life to high salt concentrations. FEMS Microbiol. Rev. 2018, 42, 353–375. [Google Scholar] [CrossRef] [PubMed]
- Raetz, C.R.; Whitfield, C. Lipopolysaccharide endotoxins. Annu. Rev. Biochem. 2002, 71, 635–700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carillo, S.; Pieretti, G.; Lindner, B.; Romano, I.; Nicolaus, B.; Lanzetta, R.; Parrilli, M.; Corsaro, M.M. Structural characterization of the core oligosaccharide isolated from the lipopolysaccharide of the haloalkaliphilic bacterium Salinivibrio sharmensis strain BAG(T). Carbohydr. Res. 2013, 368, 61–67. [Google Scholar] [CrossRef]
- Carillo, S.; Pieretti, G.; Lindner, B.; Romano, I.; Nicolaus, B.; Lanzetta, R.; Parrilli, M.; Corsaro, M.M. The Lipid A from the haloalkaliphilic bacterium Salinivibrio sharmensis strain BAG(T). Mar. Drugs 2013, 11, 184–193. [Google Scholar] [CrossRef] [Green Version]
- Toukach, P.V.; Egorova, K.S. Carbohydrate structure database merged from bacterial, archaeal, plant and fungal parts. Nucl. Acids Res. 2016, 44, D1229–D1236. [Google Scholar] [CrossRef]
- Westphal, O.; Jann, K. Bacterial lipopolysaccharides. Extraction with phenol-water and further applications of the procedure. Methods Carbohydr. Chem. 1965, 5, 83–91. [Google Scholar]
- Bock, K.; Pedersen, C. Carbon-13 nuclear magnetic resonance spectroscopy of monosaccharides. Adv. Carbohydr. Chem. Biochem. 1983, 41, 27–66. [Google Scholar]
- Lipkind, G.M.; Shashkov, A.S.; Knirel, Y.A.; Vinogradov, E.V.; Kochetkov, N.K. A computer-assisted structural analysis of regular polysaccharides on the basis of 13C-n.m.r. data. Carbohydr. Res. 1988, 175, 59–75. [Google Scholar] [CrossRef]
- Pellicer-Nacher, C.; Smets, B.F. Structure, composition, and strength of nitrifying membrane-aerated biofilms. Water Res. 2014, 57, 151–161. [Google Scholar] [CrossRef]
- Bogino, P.C.; de las Mercedes Oliva, M.; Sorroche, F.G.; Giordano, W. The role of bacterial biofilms and surface components in plant-bacterial associations. Int. J. Mol. Sci. 2013, 14, 15838–15859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bazaka, K.; Crawford, R.J.; Nazarenko, E.L.; Ivanova, E.P. Bacterial Extracellular Polysaccharides. In Advances in Experimental Medicine and Biology; Linke, D., Goldman, A., Eds.; Springer: Dordrecht, The Netherlands, 2011; Volume 715, pp. 213–226. [Google Scholar] [CrossRef]
- Casillo, A.; Lanzetta, R.; Parrilli, M.; Corsaro, M.M. Exopolysaccharides from marine and marine extremophilic bacteria: Structures, properties, ecological roles and applications. Mar. Drugs. 2018, 16, 69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leone, S.; Molinaro, A.; Dubery, I.; Lanzetta, R.; Parrilli, M. The O-specific polysaccharide structure from the lipopolysaccharide of the Gram-negative bacterium Raoultella terrigena. Carbohydr. Res. 2007, 342, 1514–1518. [Google Scholar] [CrossRef] [PubMed]
- Perepelov, A.V.; Senchenkova, S.N.; Shashkov, A.S.; Knirel, Y.A.; Liu, B.; Feng, L.; Wang, L. A completed structure of the O-polysaccharide from Shigella dysenteriae type 10. Rus. J. Bioorg. Chem. 2009, 35, 131–133. [Google Scholar] [CrossRef] [PubMed]
- Grue, M.R.; Parolis, H.; Parolis, L.A.S. Structure of the capsular antigen of Escherichia coli O8: K50: H. Carbohydr. Res. 1994, 258, 233–241. [Google Scholar] [CrossRef]
- Garozzo, D.; Impallomeni, G.; Spina, E.; Sturiale, L.; Cesàro, A.; Cescutti, P. Identification of N-acetylglucosamine and 4-O-[1-carboxyethyl]mannose in the exopolysaccharide from Cyanospira capsulate. Carbohydr. Res. 1995, 270, 97–106. [Google Scholar] [CrossRef]
- Kochetkov, N.K.; Sviridov, A.F.; Arifkhodzhaev, K.A.; Chizhov, O.S.; Shashkov, A.S. The structure of the extracellular polysaccharide from Mycobacterium lacticolum strain 121. Carbohydr. Res. 1979, 71, 193–203. [Google Scholar] [CrossRef]
- Shashkov, A.S.; Sviridov, A.F.; Arifkhodzhaev, K.A.; Chizhov, O.S.; Botvinko, I.V. Mycobacterial polysaccharides. Part 6. Carbon-13 NMR spectrum of an extracellular polysaccharide produced by Mycobacterium album B-88. Bioorganicheskaya Khimia. 1982, 8, 1252–1255. [Google Scholar]
- Ovchinnikova, O.G.; Rozalski, A.; Liu, B.; Knirel, Y.A. O-antigens of bacteria of the genus Providencia: Structure, serology, genetics, and biosynthesis. Biochem. Mosc. 2013, 78, 798–817. [Google Scholar] [CrossRef] [PubMed]
- Knirel, Y.A.; Perepelov, A.V.; Kondakova, A.N.; Senchenkova, S.N.; Sidorczyk, Z.; Rozalski, A.; Kaza, W. Structure and serology of O-antigens as the basis for classification of Proteus strains. Innate Immun. 2011, 17, 70–96. [Google Scholar] [CrossRef] [PubMed]
- Sehgal, S.N.; Gibbons, N.E. Effect of some metal ions on the growth of Halobacterium cutirubrum. Can. J. Microbiol. 1960, 6, 165–169. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, I.M.; Konnova, S.A.; Sigida, E.N.; Lyubun, E.V.; Muratova, A.Y.; Fedonenko, Y.P.; Elbanna, K. Bioremediation potential of a halophilic Halobacillus sp. strain, EG1HP4QL: Exopolysaccharide production, crude oil degradation, and heavy metal tolerance. Extremophiles 2020, 24, 157–166. [Google Scholar] [CrossRef]
- Gregersen, T. Rapid method for distinction of Gram-negative from Gram-positive bacteria. Eur. J. App. Microbiol. Biotech. 1978, 5, 123–127. [Google Scholar] [CrossRef]
- Cowan, S.T.; Steel, K.J. Manual for Identification of Medical Bacteria; Cambrige University Press: London, UK, 1965. [Google Scholar]
- Smibert, R.M.; Krieg, N.R. Phenotypic characterization. In Methods for General and Molecular Bacteriology; Gerhard, P., Murray, R.G.E., Wood, W.A., Krieg, N.R., Eds.; American Society for Microbiology: Washington, DC, USA, 1994; pp. 607–654. [Google Scholar]
- Chen, Y.G.; Cui, X.L.; Pukall, R.; Li, H.M.; Yang, Y.L.; Xu, L.H.; Wen, M.L.; Peng, Q.; Jiang, C.L. Salinicoccus kunmingensis sp. nov., a moderately halophilic bacterium isolated from a salt mine in Yunnan, south-west China. Int. J. Syst. Evol. Microbiol. 2007, 57, 2327–2332. [Google Scholar] [CrossRef]
- Weisburg, W.G.; Barns, S.M.; Pelletier, D.A.; Lane, D.J. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 1991, 173, 697–703. [Google Scholar] [CrossRef] [Green Version]
- Elbanna, K.; Ibrahim, I.M.; Revol-Junelles, A.M. Purification and characterization of halo-alkali-thermophilic protease from Halobacterium sp. strain HP25 isolated from raw salt, Lake Qarun, Fayoum, Egypt. Extremophiles 2015, 19, 763–774. [Google Scholar] [CrossRef] [PubMed]
- Hitchcock, P.J.; Brown, T.M. Morphological heterogeneity among Salmonella lipopolysaccharide chemotypes in silver-stain polyacrylamide gels. J. Bacteriol. 1983, 154, 269–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, C.M.; Frasch, C.E. A sensitive silver stain for detecting lipopolysaccharides in polyacrylamide gels. Anal. Biochem. 1982, 119, 115–119. [Google Scholar] [CrossRef]
- Sawardeker, J.S.; Sloneker, J.H.; Jeanes, A. Quantitative determination of monosaccharides as their alditol acetates by gas liquid chromatography. Anal. Chem. 1965, 37, 1602–1603. [Google Scholar] [CrossRef]
- Leontein, K.; Lindberg, B.; Lönngren, J. Assignment of absolute configuration of sugars by g.l.c. of their acetylated glycosides formed from chiral alcohols. Carbohydr. Res. 1978, 62, 359–362. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Residue | H-1 C-1 3J1,2 | H-2 C-2 3J2,3 | H-3 C-3 3J3,4 | H-4 C-4 3J4,5 | H-5 C-5 3J5,6 | H-6(a, b) C-6 |
|---|---|---|---|---|---|---|
| →2)-β-d-Manp4Lac-(1→, A | 4.82 97.6 <2 | 3.88 77.1 ~4 | 3.80 74.5 ~10 | 3.60 78.3 ~10 | 3.45 77.3 ~6 | 3.76, 3.93 61.6 |
| →3)-α-d-ManpNAc-(1→, B | 5.05 96.5 <2 | 4.60 50.2 ~4 | 4.24 75.9 ~10 | 3.61 65.9 ~10 | 3.96 73.6 ~6 | 3.82, 3.84 61.3 |
| →3)-β-l-Rhap-(1→, C | 4.88 102.2 <2 | 4.29 67.8 ~4 | 3.67 77.8 ~10 | 3.46 71.3 ~6 | 3.46 73.4 ~6 | 1.33 18.0 |
| →4)-α-d-GlcpNAc-(1→, D | 5.07 99.6 3.7 | 3.87 55.2 ~10 | 3.99 71.4 ~10 | 3.74 78.6 ~10 | 4.17 71.2 ~10 | 3.69, 4.04 61.6 |
| Lac, L | 182.5 | 4.00 79.8 | 1.40 20.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sigida, E.N.; Ibrahim, I.M.; Kokoulin, M.S.; Abulreesh, H.H.; Elbanna, K.; Konnova, S.A.; Fedonenko, Y.P. Structure of the 4-O-[1-Carboxyethyl]-d-Mannose-Containing O-Specific Polysaccharide of a Halophilic Bacterium Salinivibrio sp. EG9S8QL. Mar. Drugs 2021, 19, 508. https://doi.org/10.3390/md19090508
Sigida EN, Ibrahim IM, Kokoulin MS, Abulreesh HH, Elbanna K, Konnova SA, Fedonenko YP. Structure of the 4-O-[1-Carboxyethyl]-d-Mannose-Containing O-Specific Polysaccharide of a Halophilic Bacterium Salinivibrio sp. EG9S8QL. Marine Drugs. 2021; 19(9):508. https://doi.org/10.3390/md19090508
Chicago/Turabian StyleSigida, Elena N., Ibrahim M. Ibrahim, Maxim S. Kokoulin, Hussein H. Abulreesh, Khaled Elbanna, Svetlana A. Konnova, and Yulia P. Fedonenko. 2021. "Structure of the 4-O-[1-Carboxyethyl]-d-Mannose-Containing O-Specific Polysaccharide of a Halophilic Bacterium Salinivibrio sp. EG9S8QL" Marine Drugs 19, no. 9: 508. https://doi.org/10.3390/md19090508
APA StyleSigida, E. N., Ibrahim, I. M., Kokoulin, M. S., Abulreesh, H. H., Elbanna, K., Konnova, S. A., & Fedonenko, Y. P. (2021). Structure of the 4-O-[1-Carboxyethyl]-d-Mannose-Containing O-Specific Polysaccharide of a Halophilic Bacterium Salinivibrio sp. EG9S8QL. Marine Drugs, 19(9), 508. https://doi.org/10.3390/md19090508