
Low Molecular Weight, 4-O-Sulfation, and Sulfation at Meta-Fucose Positively Promote the Activities of Sea Cucumber Fucoidans on Improving Insulin Resistance in HFD-Fed Mice



 and
and
Abstract
:1. Introduction
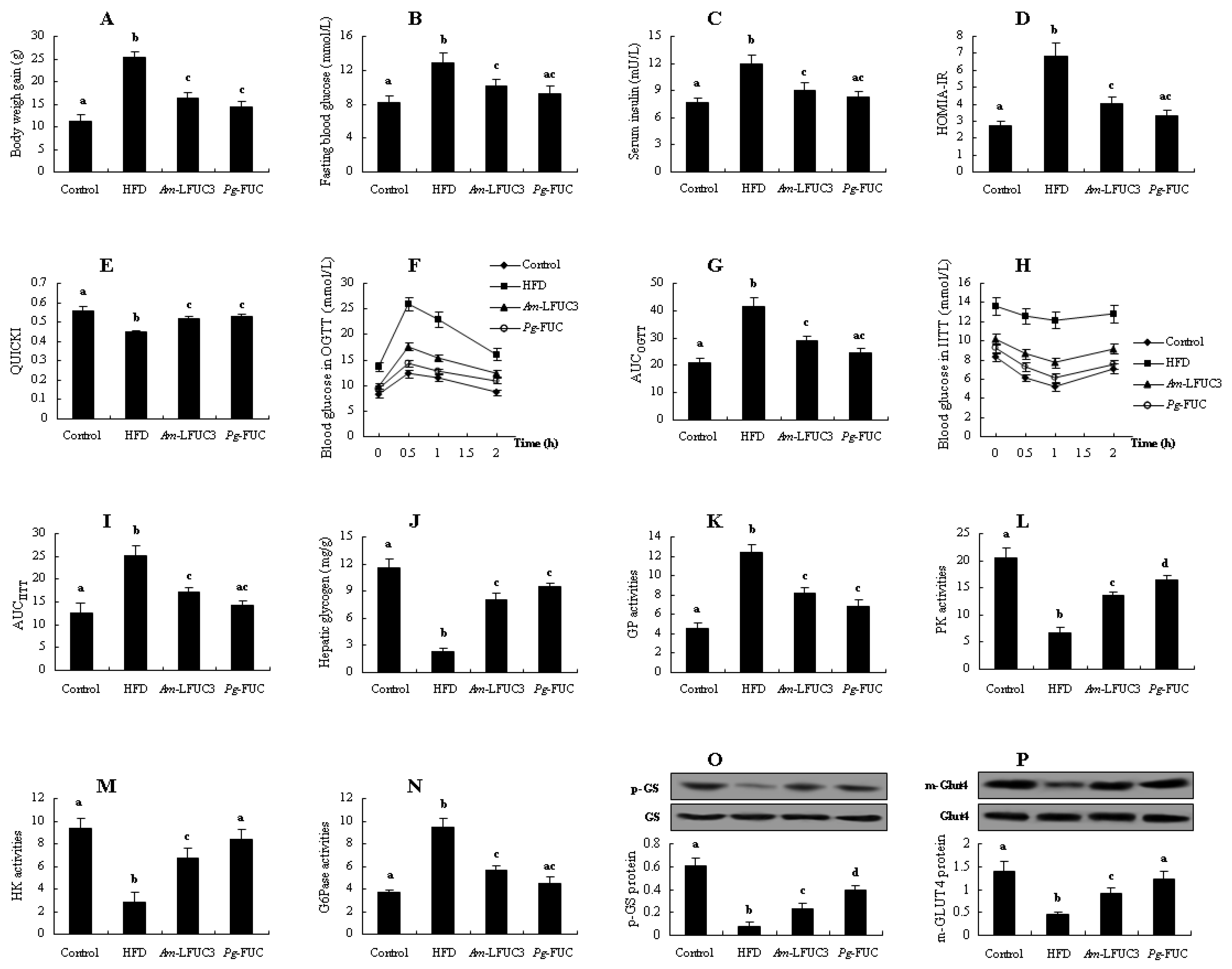
2. Results
2.1. Effects of Fucoidans with Different Mw on Insulin Resistance
2.2. Effects of Fucoidans with Different Cs on Insulin Resistance
2.3. Effects of Sulfation at Ortho-, or Meta-fucose Positions on Insulin Resistance
2.4. Effects of 2-O-Sulfation or 4-O-Sulfation on Insulin Resistance
3. Discussion
4. Materials and Methods
4.1. Preparation of Fucoidans
4.2. Low-Mw Fucoidans Preparation
4.3. Animal Experiments
4.4. OGTT and IITT
4.5. Insulin Stimulation and Plasma Membrane Preparation
4.6. Blood Glucose and Plasma Insulin Analysis
4.7. Hepatic Glycogen Content Detection
4.8. Glucose Metabolism Enzymes Test
4.9. Western Blotting
4.10. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Saravanakumar, K.; Park, S.; Sathiyaseelan, A.; Kim, K.; Cho, S.; Mariadoss, A.V.A.M.; Wang, M. Metabolite profiling of methanolic extract of gardenia jaminoides by LC-MS/MS and GC-MS and its anti-diabetic, and anti-oxidant activities. Pharmaceuticals 2021, 14, 102. [Google Scholar] [CrossRef] [PubMed]
- Ren, L.; Zhou, X.; Huang, X.; Wang, C.; Li, Y. The IRS/PI3K/Akt signaling pathway mediates olanzapine-induced hepatic insulin resistance in male rats. Life Sci. 2019, 217, 229–236. [Google Scholar] [CrossRef]
- Bao, S.; Wu, Y.L.; Wang, X.; Han, S.; Cho, S.; Ao, W.; Nan, J.X. Agriophyllum oligosaccharides ameliorate hepatic injury in type 2 diabetic db/db mice targeting INS-R/IRS-2/PI3K/AKT/PPAR-gamma/Glut4 signal pathway. J. Ethnopharmacol. 2020, 257, 112863. [Google Scholar] [CrossRef]
- Zhang, H.; Hui, J.; Yang, J.; Deng, J.; Fan, D. Eurocristatine, a plant alkaloid from Eurotium cristatum, alleviates insulin resistance in db/db diabetic mice via activation of PI3K/AKT signaling pathway. Eur. J. Pharmacol. 2020, 887, 173557. [Google Scholar] [CrossRef]
- Cao, C.; Li, C.; Chen, Q.; Huang, Q.; Pérez, M.E.M.; Fu, X. Physicochemical characterization, potential antioxidant and hypoglycemic activity of polysaccharide from Sargassum pallidum. Int. J. Biol. Macromol. 2019, 138, 1009–1017. [Google Scholar] [CrossRef]
- Subash-Babu, P.; Ignacimuthu, S.; Alshatwi, A.A. Nymphayol increases glucose-stimulated insulin secretion by RIN-5F cells and GLUT4-mediated insulin sensitization in type 2 diabetic rat liver. Chem. Biol. Interact. 2015, 226, 72–81. [Google Scholar] [PubMed]
- Barazzoni, R.; Gortan Cappellari, G.; Ragni, M.; Nisoli, E. Insulin resistance in obesity: An overview of fundamental alterations. Eat. Weight Disord. 2018, 23, 149–157. [Google Scholar] [CrossRef] [PubMed]
- Saravanakumar, K.; Park, S.; Mariadoss, A.V.A.; Sathiyaseelan, A.; Veeraraghavan, V.P.; Kim, K.; Wang, M. Chemical composition, antioxidant, and anti-diabetic activities of ethyl acetate fraction of Stachys riederi var. japonica (Miq.) in streptozotocin-induced type 2 diabetic mice. Food Chem. Toxicol. 2021, 155, 112374. [Google Scholar] [CrossRef]
- Gao, X.; Tian, Y.; Randell, E.; Zhou, H.; Sun, G. Unfavorable associations between serum trimethylamine N-oxide and L-carnitine levels with components of metabolic syndrome in the newfoundland population. Front. Endocrinol. (Lausanne) 2019, 10, 168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariadoss, A.V.A.M.; Park, S.; Saravanakumar, K.; Sathiyaseelan, A.; Wang, M. Ethyl acetate fraction of Helianthus tuberosus L. induces anti-diabetic, and wound-healing activities in insulin-resistant human liver cancer and mouse fibroblast cells. Antioxidants 2021, 10, 99. [Google Scholar] [CrossRef]
- Zheng, Y.; Ley, S.; Hu, F. Global aetiology and epidemiology of type 2 diabetes mellitus and its complications. Nat. Rev. Endocrinol. 2018, 14, 88–98. [Google Scholar] [CrossRef]
- Guo, X.; Ye, X.; Sun, Y.; Wu, D.; Wu, N.; Hu, Y.; Chen, S. Ultrasound effects on the degradation kinetics, structure, and antioxidant activity of sea cucumber fucoidan. J. Agric. Food Chem. 2014, 62, 1088–1095. [Google Scholar] [CrossRef]
- Zhang, M.; Chen, L.; Liu, Y.; Chen, M.; Zhang, S.; Kong, D. Sea cucumber Cucumaria frondosa fucoidan inhibits osteosarcoma adhesion and migration by regulating cytoskeleton remodeling. Oncol. Rep. 2020, 44, 469–476. [Google Scholar] [CrossRef]
- Li, C.; Niu, Q.; Li, S.; Zhang, X.; Liu, C.; Cai, C.; Li, G.; Yu, G. Fucoidan from sea cucumber Holothuria polii: Structural elucidation and stimulation of hematopoietic activity. Int. J. Biol. Macromol. 2020, 154, 1123–1131. [Google Scholar] [CrossRef]
- Li, Q.; Jiang, S.; Shi, W.; Qi, X.; Song, W.; Mou, J.; Yang, J. Structure characterization, antioxidant and immunoregulatory properties of a novel fucoidan from the sea cucumber Stichopus chloronotus. Carbohyd. Polym. 2020, 231, 115767. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Chang, Y.; Xue, C.; Wang, J.; Shen, J. Gastric protective activities of sea cucumber fucoidans with different molecular weight and chain conformations: A structure–activity relationship investigation. J. Agric. Food Chem. 2018, 66, 8615–8622. [Google Scholar] [CrossRef]
- Zhu, Y.; Tian, Y.; Wang, N.; Chang, Y.; Xue, C.; Wang, J. Structure-function relationship analysis of fucoidan from sea cucumber [Holothuria tubulosa] on ameliorating metabolic inflammation. J. Food Biochem. 2021, 45, e13500. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Wang, J.; Wang, J.; Yang, H.; Yan, X.; Su, L. Fucoidan from Acaudina molpadioides improves insulin resistance by altering gut microbiota dysfunction. J. Funct. Food. 2019, 57, 59–67. [Google Scholar] [CrossRef]
- Wang, J.; Hu, S.; Jiang, W.; Song, W.; Cai, L.; Wang, J. Fucoidan from sea cucumber may improve hepatic inflammatory response and insulin resistance in mice. Int. Immunopharmacol. 2016, 31, 15–23. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, J.; Zhao, Y.; Hu, S.; Shi, D.; Xue, C. Fucoidan from sea cucumber Cucumaria frondosa exhibits anti-hyperglycemic effects in insulin resistant mice via activating the PI3K/PKB pathway and GLUT4. J. Biosci. Bioeng. 2016, 121, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Zhao, F.; Liu, Q.; Cao, J.; Xu, Y.; Pei, Z.; Fan, H.; Yuan, Y.; Shen, X.; Li, C. A sea cucumber [Holothuria leucospilota] polysaccharide improves the gut microbiome to alleviate the symptoms of type 2 diabetes mellitus in Goto-Kakizaki rats. Food Chem. Toxicol. 2020, 135, 110886. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Li, J.; Mao, G.; Yan, L.; Hu, Y.; Ye, X.; Tian, D.; Linhardt, R.; Chen, S. Effect of the sulfation pattern of sea cucumber-derived fucoidan oligosaccharides on modulating metabolic syndromes and gut microbiota dysbiosis caused by HFD in mice. J. Funct. Food. 2019, 55, 193–210. [Google Scholar] [CrossRef]
- Yu, L.; Xue, C.; Chang, Y.; Xu, X.; Ge, L.; Liu, G.; Wang, Y. Structure elucidation of fucoidan composed of a novel tetrafucose repeating unit from sea cucumber Thelenota ananas. Food Chem. 2014, 146, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.; Xia, G.; Wang, J.; Wang, Y.; Li, Z.; Xue, C. Fucoidan from sea cucumber protects against high-fat high-sucrose diet-induced hyperglycaemia and insulin resistance in mice. J. Funct. Food. 2014, 10, 128–138. [Google Scholar] [CrossRef]
- Wright, C.M.; Bezabhe, W.; Fitton, J.H.; Stringer, D.N.; Bereznicki, L.R.E.; Peterson, G.M. Effect of a fucoidan extract on insulin resistance and cardiometabolic markers in obese, nondiabetic subjects: A randomized, controlled trial. J. Altern. Complement. Med. 2019, 25, 346–352. [Google Scholar] [CrossRef]
- Zhu, Z.; Zhu, B.; Sun, Y.; Ai, C.; Wang, L.; Wen, C.; Yang, J.; Song, S.; Liu, X. Sulfated polysaccharide from sea cucumber and its depolymerized derivative prevent obesity in association with modification of gut microbiota in high-fat diet-fed mice. Mol. Nutr. Food Res. 2018, 62, 1800446. [Google Scholar] [CrossRef]
- Yang, C.; Chung, D.; Shin, I.S.; Lee, H.; Kim, J.; Lee, Y.; You, S. Effects of molecular weight and hydrolysis conditions on anticancer activity of Fucoidans from sporophyll of Undaria pinnatifida. Int. J. Biol. Macromol. 2008, 43, 433–437. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.; Wang, J.; Jin, W.; Zhang, H.; Zhang, Q. Degradation of Laminaria japonica fucoidan by hydrogen peroxide and antioxidant activities of the degradation products of different molecular weights. Carbohyd. Polym. 2012, 87, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wu, N.; Chen, Y.; Tan, J.; Wang, J.; Geng, L.; Qin, Y.; Zhang, Q. Degradation of different molecular weight fucoidans and their inhibition of TGF-β1 induced epithelial–mesenchymal transition in mouse renal tubular epithelial cells. Int. J. Biol. Macromol. 2020, 151, 545–553. [Google Scholar] [CrossRef]
- Li, B.; Zhao, R.; Wei, X. Anticoagulant activity of fucoidan form Hizikia fusiforme. Agro Food Ind. Hi-Tech. 2008, 19, 22–24. [Google Scholar]
- Cho, M.; Lee, B.; You, S. Relationship between oversulfation and conformation of low and high molecular weight fucoidans and evaluation of their in vitro anticancer activity. Molecules 2010, 16, 291–297. [Google Scholar] [CrossRef] [Green Version]
- Koyanagi, S.; Tanigawa, N.; Nakagawa, H.; Soeda, S.; Shimeno, H. Oversulfation of fucoidan enhances its anti-angiogenic and antitumor activities. Biochem. Pharmacol. 2003, 65, 173–179. [Google Scholar] [CrossRef]
- Li, S.; Li, J.; Zhi, Z.; Hu, Y.; Ge, J.; Ye, X.; Tian, D.; Linhardt, R.J.; Chen, S. 4-O-Sulfation in sea cucumber fucodians contribute to reversing dyslipidiaemia caused by HFD. Int. J. Biol. Macromol. 2017, 99, 96–104. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Li, J.; Mao, G.; Wu, T.; Hu, Y.; Ye, X.; Tian, D.; Linhardt, R.J.; Chen, S. A fucoidan from sea cucumber Pearsonothuria graeffei with well-repeated structure alleviates gut microbiota dysbiosis and metabolic syndromes in HFD-fed mice. Food Funct. 2018, 9, 5371–5380. [Google Scholar] [CrossRef]
- Mandal, R.; Mateu, C.G.; Chattopadhyay, K.; Pujol, C.A.; Damonte, E.B.; Ray, B. Structural features and antiviral activity of sulphated fucans from the brown seaweed Cystoseira indica. Antivir. Chem. Chemother. 2007, 18, 153–162. [Google Scholar] [CrossRef] [Green Version]
- Pereira, M.S.; Melo, F.R.; Mourão, P.A.S. Is there a correlation between structure and anticoagulant action of sulfated galactans and sulfated fucans? Glycobiology 2002, 12, 573–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonseca, R.J.C.; Santos, G.R.C.; Mourão, P.A.S. Effects of polysaccharides enriched in 2,4-disulfated fucose units on coagulation, thrombosis and bleeding Practical and conceptual implications. Thromb. Haemost. 2009, 102, 829–836. [Google Scholar] [CrossRef]
- Chevolot, L.; Mulloy, B.; Ratiskol, J.; Foucault, A.; Colliec-Jouault, S. A disaccharide repeat unit is the major structure in fucoidans from two species of brown algae. Carbohydr. Res. 2001, 330, 529–535. [Google Scholar] [CrossRef]
- Chang, Y.; Hu, Y.; Yu, L.; McClements, D.; Xu, X.; Liu, G.; Xue, C. Primary structure and chain conformation of fucoidan extracted from sea cucumber Holothuria tubulosa. Carbohydr. Polym. 2016, 136, 1091–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdelmageed, M.E.; Shehatou, G.S.G.; Suddek, G.M.; Salem, H.A. Protocatechuic acid improves hepatic insulin resistance and restores vascular oxidative status in type-2 diabetic rats. Environ. Toxicol. Pharmacol. 2021, 83, 103577. [Google Scholar] [CrossRef]
- Hu, S.; Du, M.; Su, L.; Yang, H. Phosphatidylserine from Portunus trituberculatus eggs alleviates insulin resistance and alters the gut microbiota in high-fat-diet-fed mice. Mar. Drug. 2020, 18, 483. [Google Scholar] [CrossRef] [PubMed]
- Mirzaalian, Y.; Nourian, M.; Gholamalizadeh, M.; Doaei, S.; Hatami, M.; Hassanzadeh, A.; Askari, G.; Farahi, R. The association of quantitative insulin sensitivity indices [HOMA-IR and QUICKI] with anthropometric and cardiometabolic indicators in adolescents. Arch. Med. Sci. Atheroscler. Dis. 2019, 4, e32. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | HFD | Ta-LFUC | Ib-FUC | Ta-FUC | Am-FUC | ||
|---|---|---|---|---|---|---|---|
| Body weight gain (g) | 11.16 ± 1.47 a | 25.43 ± 1.25 b | 23.04 ± 1.33 b | 15.94 ± 1.41 d | 24.37 ± 1.72 b | 21.68 ± 1.39 c | |
| Fasting blood glucose (mmol/L) | 8.25 ± 0.77 a | 12.89 ± 1.13 b | 12.49 ± 0.66 b | 9.61 ± 0.59 d | 13.24 ± 0.71 b | 11.57 ± 0.84 b | |
| Serum insulin (mU/L) | 7.63 ± 0.60 a | 12.01 ± 0.96 b | 11.12 ± 0.78 b | 8.75 ± 0.80 d | 11.87 ± 0.78 b | 10.26 ± 0.58 c | |
| HOMIA-IR | 2.76 ± 0.21 a | 6.85 ± 0.74 b | 6.16 ± 0.63 b | 3.82 ± 0.31 d | 6.95 ± 0.58 b | 5.22 ± 0.50 c | |
| QUICKI | 0.56 ± 0.02 a | 0.45 ± 0.00 b | 0.47 ± 0.02 b | 0.52 ± 0.01 d | 0.46 ± 0.01 b | 0.47 ± 0.01 b | |
| OGTT | Blood glucose at 0 h (mmol/L) | 8.22 ± 0.64 a | 13.57 ± 0.90 b | 12.26 ± 0.64 b | 9.33 ± 0.68 d | 13.30 ± 0.77 b | 11.42± 0.71 c |
| Blood glucose at 0.5 h (mmol/L) | 12.24 ± 0.79 a | 25.88 ± 1.25 b | 24.25 ± 0.79 b | 15.56 ± 0.77 d | 24.94 ± 0.96 b | 22.15 ± 0.87 c | |
| Blood glucose at 1 h (mmol/L) | 11.47 ± 0.77 a | 22.87 ± 1.43 b | 20.04 ± 0.51 c | 13.68 ± 0.72 d | 22.51 ± 0.76 b | 19.02 ± 0.64 c | |
| Blood glucose at 2 h (mmol/L) | 8.53 ± 0.60 a | 16.07 ± 1.19 b | 14.17 ± 0.71 c | 11.85 ± 0.64 d | 15.72 ± 0.84 b | 14.64 ± 0.81 c | |
| AUCOGTT | 21.04 ± 1.67 a | 41.52 ± 3.21 b | 37.08 ± 2.56 c | 26.30 ± 1.81 d | 40.54 ± 3.06 b | 35.16 ± 2.24 c | |
| IITT | Blood glucose at 0 h (mmol/L) | 8.27 ± 0.45 a | 13.57 ± 0.96 b | 12.51 ± 0.80 b | 9.53 ± 0.58 d | 13.17 ± 0.86 b | 11.60 ± 0.56 c |
| Blood glucose at 0.5 h (mmol/L) | 6.09 ± 0.38 a | 12.56 ± 0.78 b | 11.66 ± 0.73 b | 8.01 ± 0.73 d | 12.42 ± 0.85 b | 9.81 ± 0.63 c | |
| Blood glucose at 1 h (mmol/L) | 5.19 ± 0.42 a | 12.13 ± 0.87 b | 11.17 ± 0.70 b | 7.30 ± 0.69 d | 11.89 ± 0.89 b | 9.38 ± 0.66 c | |
| Blood glucose at 2 h (mmol/L) | 7.07 ± 0.53 a | 12.80 ± 0.95 b | 11.86 ± 0.81 b | 8.42 ± 0.61 d | 12.55 ± 0.78 b | 10.63 ± 0.57 c | |
| AUCIITT | 12.54 ± 1.83 a | 25.17 ± 2.26 b | 23.27 ± 1.49 b | 16.07 ± 0.26 d | 24.70 ± 1.70 b | 20.16 ± 1.24 c | |
| Hepatic glycogen (mg/g) | 11.59 ± 0.98 a | 2.34 ± 0.30 b | 3.17 ± 0.25 b | 8.77 ± 0.42 d | 2.88 ± 0.57 b | 5.35 ± 0.47 c | |
| Hepatic GP activities | 4.58 ± 0.51 a | 12.46 ± 0.73 b | 11.24 ± 0.39 b | 8.08 ± 0.34 d | 11.61 ± 0.53 b | 9.91 ± 0.45 c | |
| Hepatic PK activities | 20.44 ± 1.97 a | 6.76 ± 0.89 b | 7.25 ± 0.83 b | 15.20 ± 0.93 d | 6.57 ± 0.88 b | 9.57 ± 0.60 c | |
| Hepatic HK activities | 9.39 ± 0.86 a | 2.82 ± 0.84 b | 3.35 ± 0.93 b | 7.53 ± 0.88 d | 3.04 ± 0.75 b | 4.65 ± 0.31 c | |
| Hepatic G6Pase activities | 3.67 ± 0.24 a | 9.45 ± 0.83 b | 8.62 ± 0.85 b | 5.14 ± 0.77 d | 9.38 ± 0.55 b | 7.13 ± 0.70 c | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, S.; Chen, S.; Zhu, H.; Du, M.; Jiang, W.; Liu, Y.; Gao, X.; Su, L.; Xu, Y. Low Molecular Weight, 4-O-Sulfation, and Sulfation at Meta-Fucose Positively Promote the Activities of Sea Cucumber Fucoidans on Improving Insulin Resistance in HFD-Fed Mice. Mar. Drugs 2022, 20, 37. https://doi.org/10.3390/md20010037
Hu S, Chen S, Zhu H, Du M, Jiang W, Liu Y, Gao X, Su L, Xu Y. Low Molecular Weight, 4-O-Sulfation, and Sulfation at Meta-Fucose Positively Promote the Activities of Sea Cucumber Fucoidans on Improving Insulin Resistance in HFD-Fed Mice. Marine Drugs. 2022; 20(1):37. https://doi.org/10.3390/md20010037
Chicago/Turabian StyleHu, Shiwei, Sichun Chen, Hongli Zhu, Mengyu Du, Wei Jiang, Yu Liu, Xiang Gao, Laijin Su, and Yangli Xu. 2022. "Low Molecular Weight, 4-O-Sulfation, and Sulfation at Meta-Fucose Positively Promote the Activities of Sea Cucumber Fucoidans on Improving Insulin Resistance in HFD-Fed Mice" Marine Drugs 20, no. 1: 37. https://doi.org/10.3390/md20010037
APA StyleHu, S., Chen, S., Zhu, H., Du, M., Jiang, W., Liu, Y., Gao, X., Su, L., & Xu, Y. (2022). Low Molecular Weight, 4-O-Sulfation, and Sulfation at Meta-Fucose Positively Promote the Activities of Sea Cucumber Fucoidans on Improving Insulin Resistance in HFD-Fed Mice. Marine Drugs, 20(1), 37. https://doi.org/10.3390/md20010037