
Cloning and Characterization of a Novel Alginate Lyase from Paenibacillus sp. LJ-23

Abstract
:1. Introduction
2. Results and Discussions
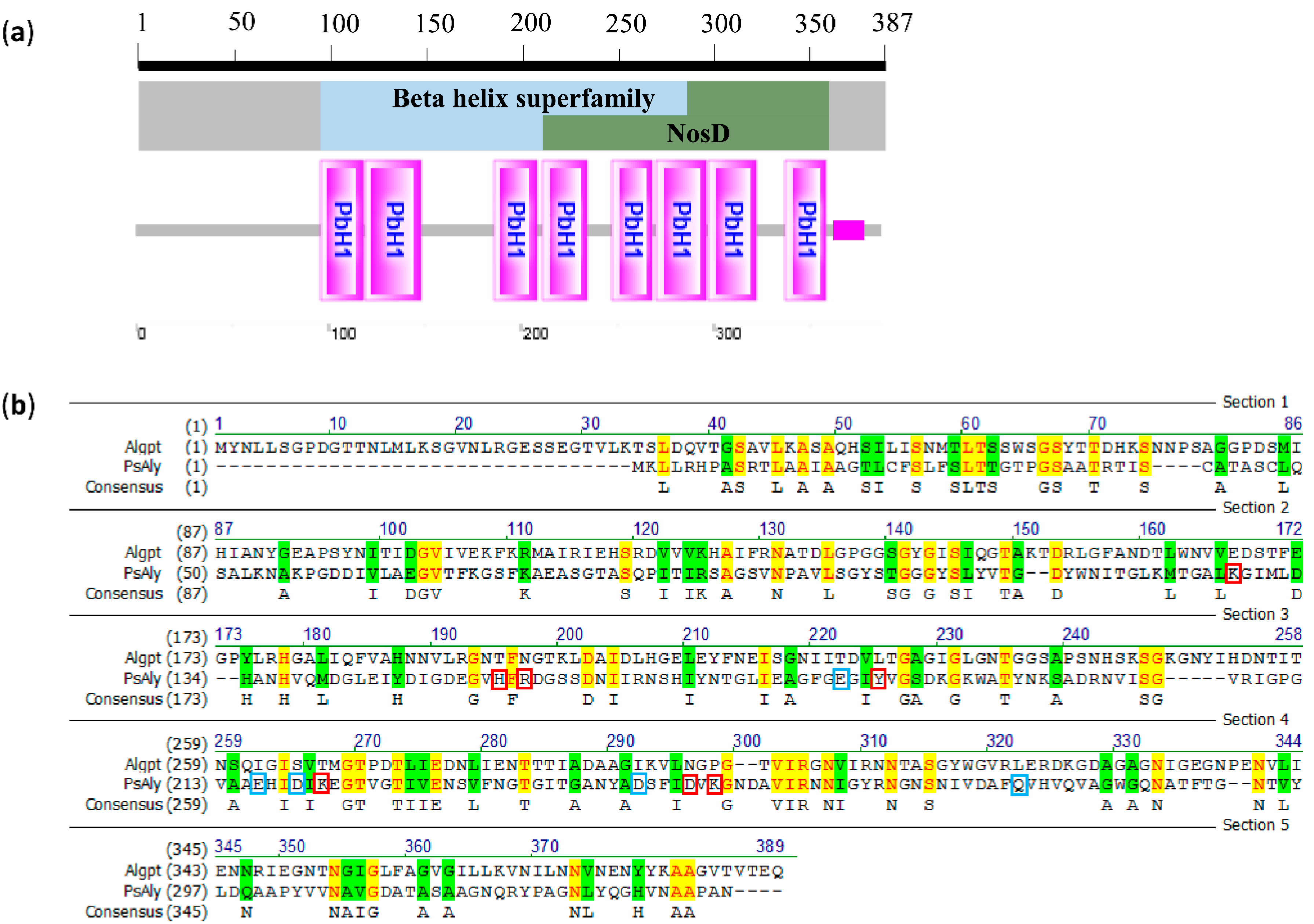
2.1. Cloning and Sequence Analysis of Algpt
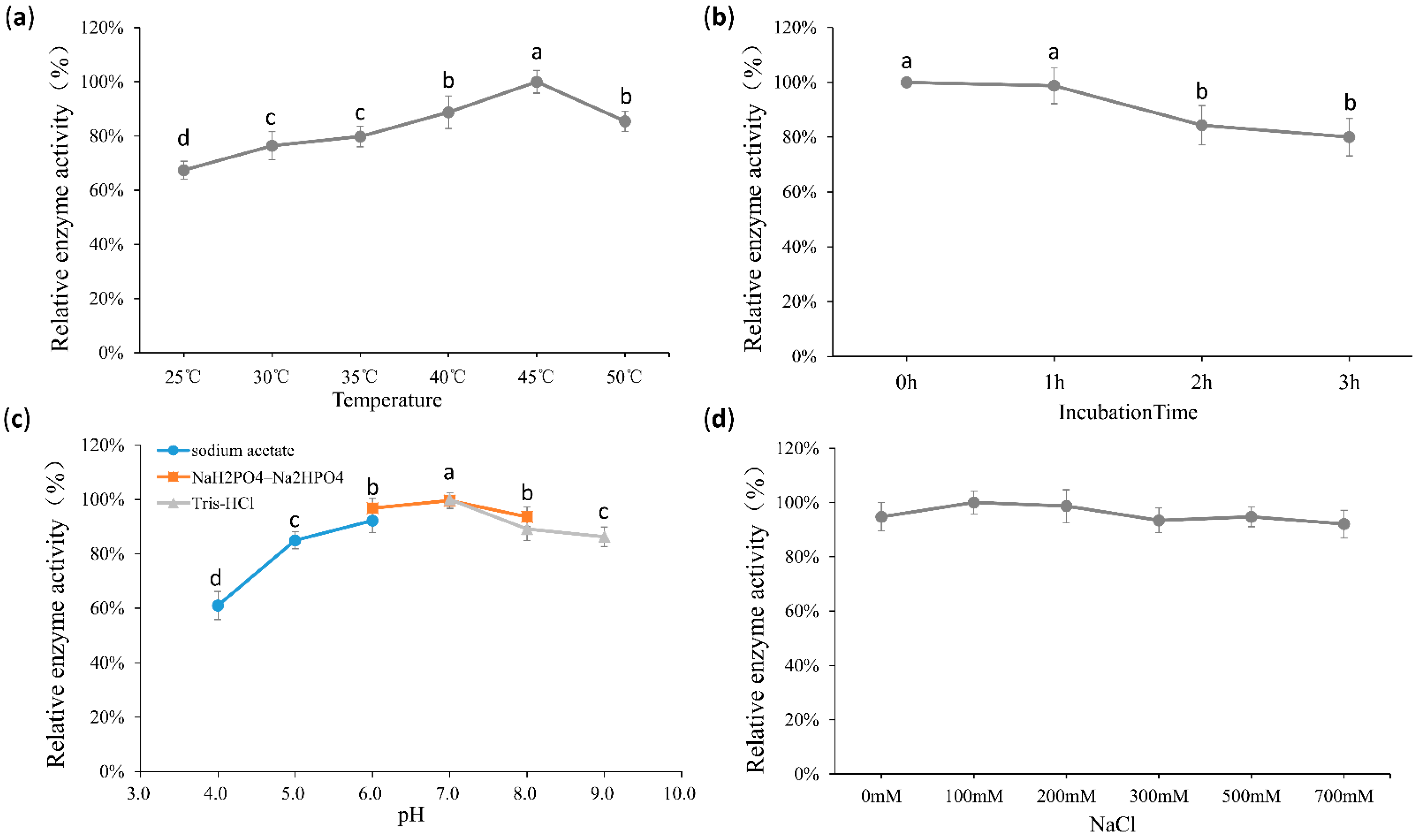
2.2. Biochemical Properties of Algpt
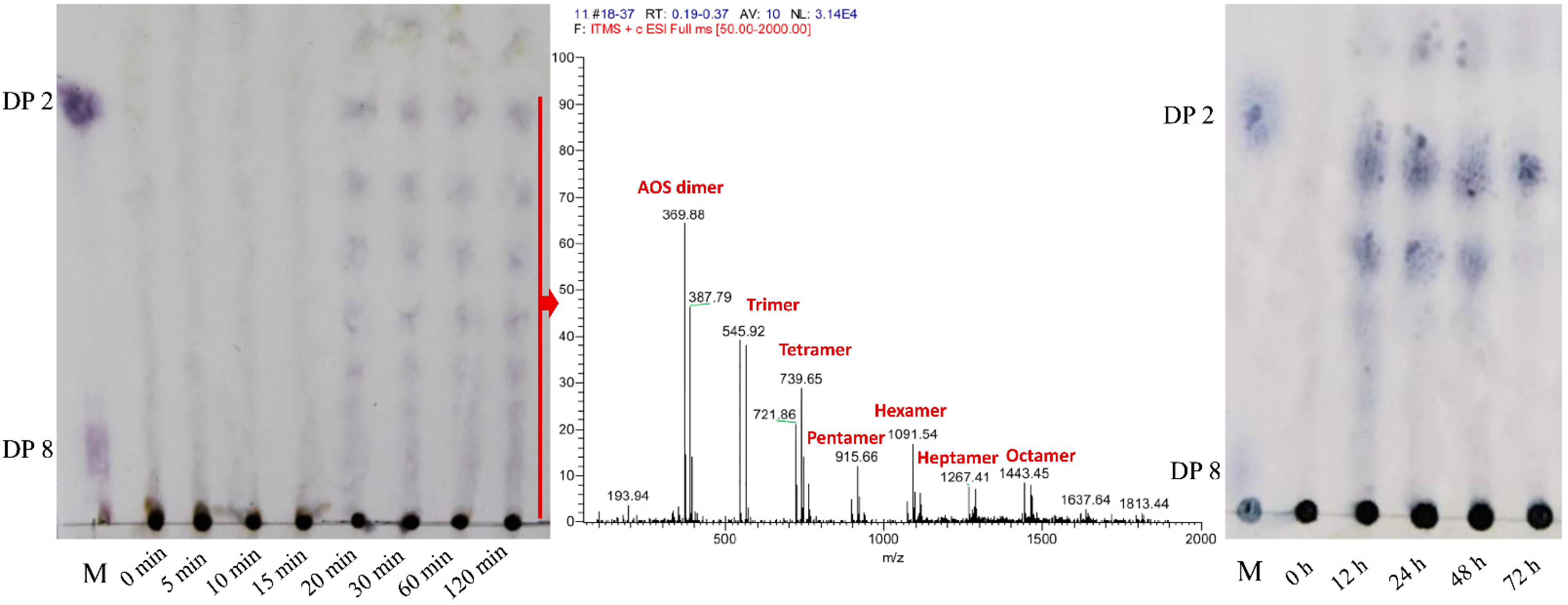
2.3. Analysis of the Enzymatic Hydrolysate
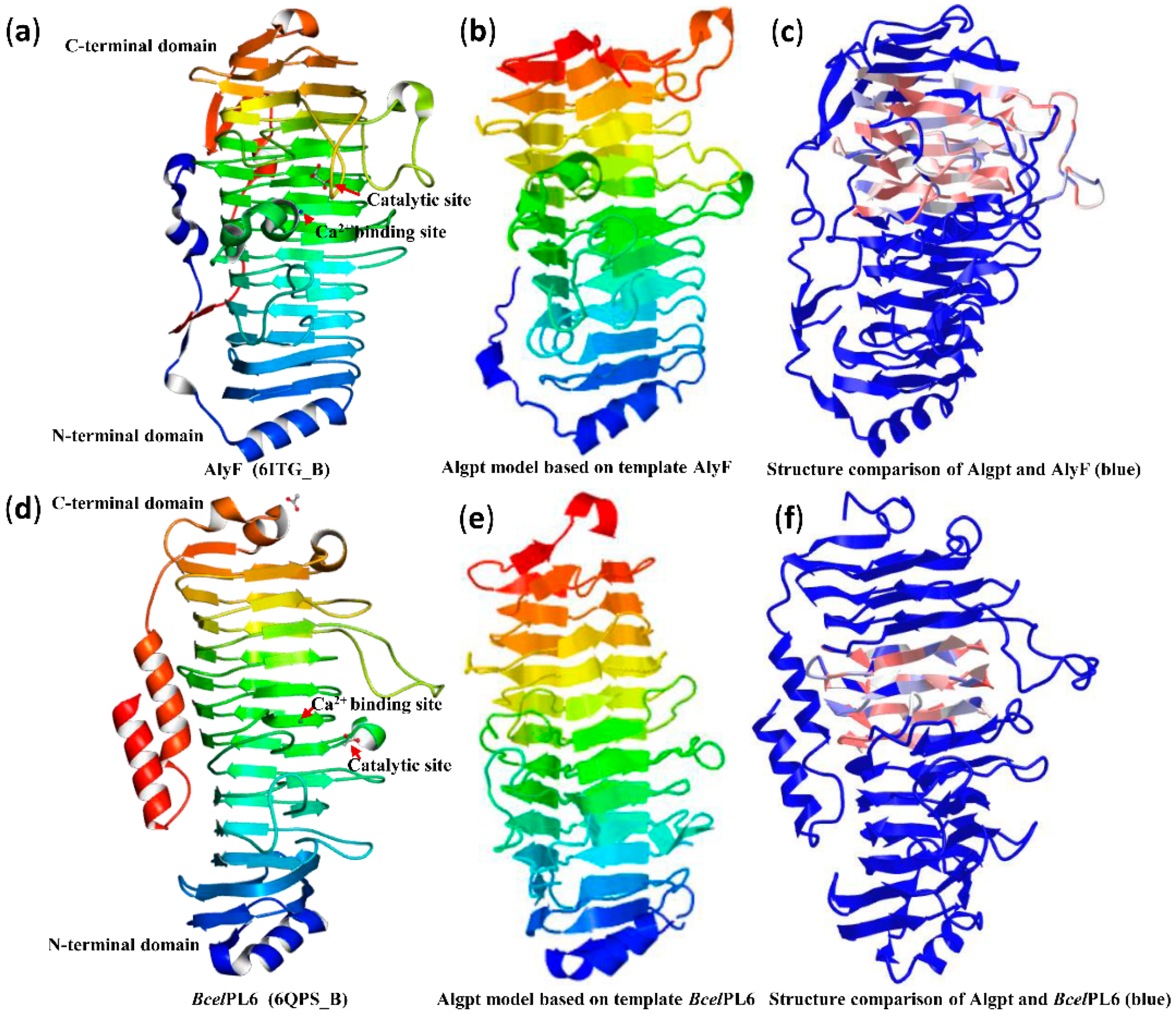
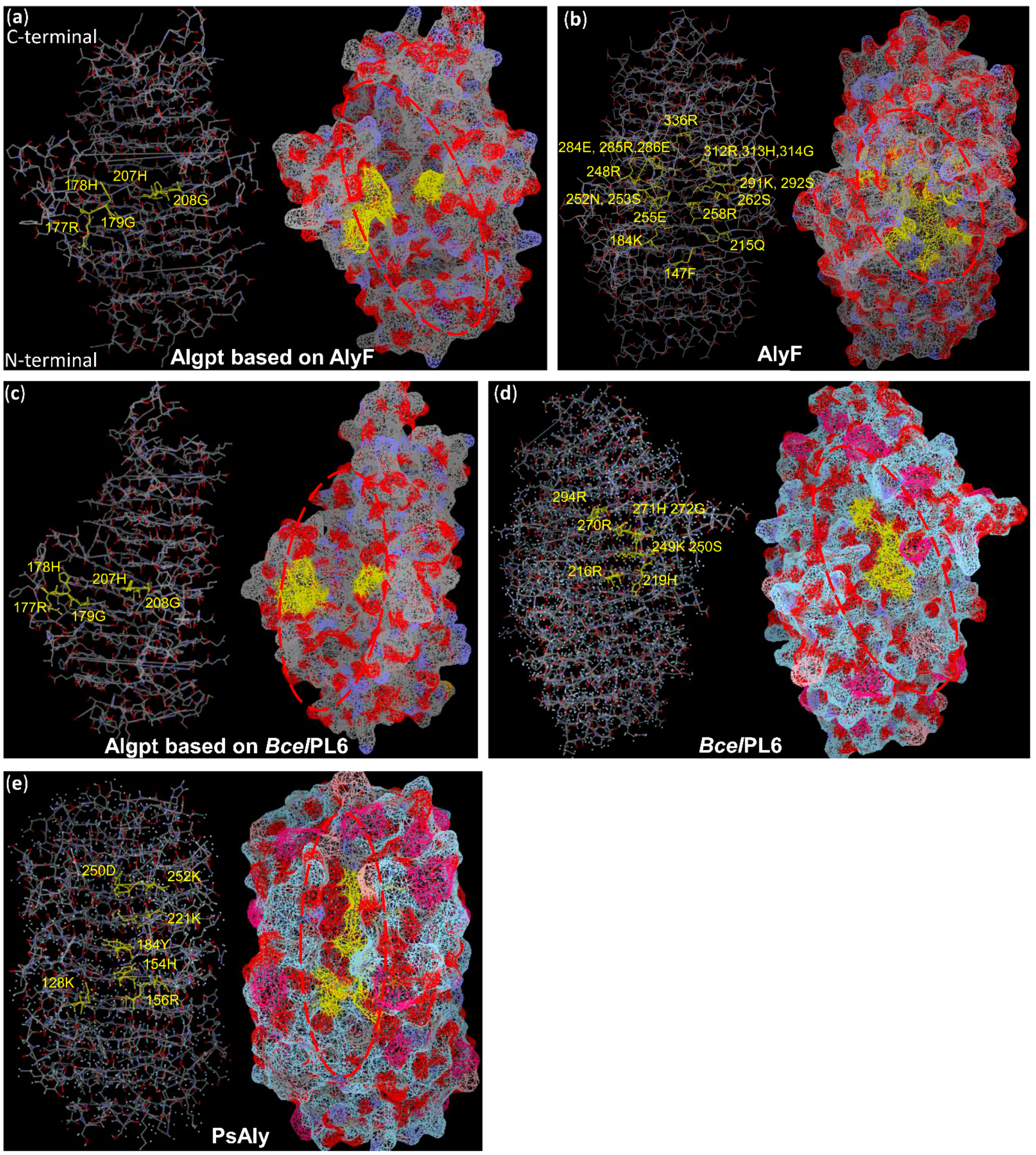
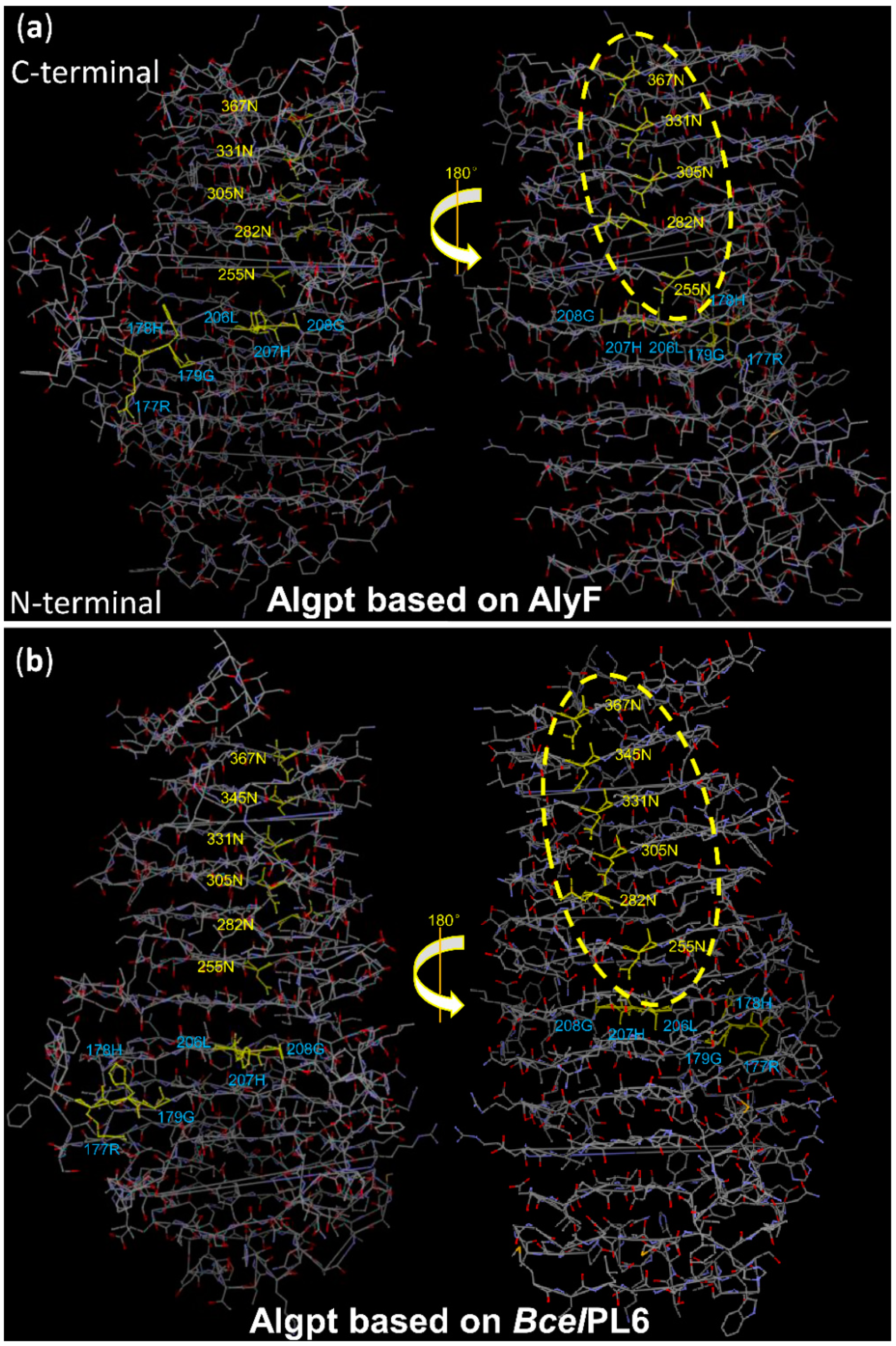
2.4. Three-Dimensional Structure Comparison and Prediction of Novel Enzyme
3. Conclusions
4. Materials and Methods
4.1. Materials
4.2. Screening and Identification of Strain LJ-23
4.3. Sequence Analysis of Algpt
4.4. Cloning and Recombinant Expression of the Alginate Lyase from Paenibacillus sp. LJ-23
4.5. Enzymatic Activity Assay
4.6. Biochemical Characterization of the Recombinant Alginate Lyase
4.7. Detection and Analysis of the Enzymatic Depolymerized Products
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sari-Chmayssem, N.; Taha, S.; Mawlawi, H.; Guégan, J.-P.; Jeftić, J.; Benvegnu, T. Extracted and depolymerized alginates from brown algae Sargassum vulgare of Lebanese origin: Chemical, rheological, and antioxidant properties. J. Appl. Phycol. 2015, 28, 1915–1929. [Google Scholar] [CrossRef]
- Synytsya, A.; Čopíková, J.; Kim, W.J.; Park, Y.I. Cell wall polysaccharides of marine algae. In Springer Handbook of Marine Biotechnology; Kim, S.K., Ed.; Springer: Berlin/Heidelberg, Germany, 2015; pp. 543–590. [Google Scholar]
- Zhu, B.; Yin, H. Alginate lyase: Review of major sources and classification, properties, structure-function analysis and applications. Bioengineered 2015, 6, 125–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donati, I.; Paoletti, S. Material properties of alginates. In Alginates: Biology and Applications; Rehm, B., Ed.; Springer: Berlin/Heidelberg, Germany, 2009; pp. 1–53. [Google Scholar]
- Lee, K.Y.; Mooney, D.J. Alginate: Properties and biomedical applications. Prog. Polym. Sci. 2012, 37, 106–126. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.; Ni, F.; Xiong, Q.; Yao, Z. Marine oligosaccharides originated from seaweeds: Source, preparation, structure, physiological activity and applications. Crit. Rev. Food Sci. Nutr. 2020, 61, 60–74. [Google Scholar] [CrossRef]
- Trincone, A. Short bioactive marine oligosaccharides: Diving into recent literature. Curr. Biotechnol. 2015, 4, 212–222. [Google Scholar] [CrossRef]
- Fan, Y.; Li, Y.; Zhang, J.; Ding, X.; Cui, J.; Wang, G.; Wang, Z.; Wang, L. Alginate enhances memory properties of antitumor CD8+ T cells by promoting cellular antioxidation. ACS Biomater. Sci. Eng. 2019, 5, 4717–4725. [Google Scholar] [CrossRef] [PubMed]
- Szekalska, M.; Wroblewska, M.; Trofimiuk, M.; Basa, A.; Winnicka, K. Alginate oligosaccharides affect mechanical properties and antifungal activity of alginate buccal films with posaconazole. Mar. Drugs 2019, 17, 692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, R.M.; Nguyen, N.T.; Lee, S.M.; Kim, Y.H.; Min, J. Alginate oligosaccharides can maintain activities of lysosomes under low pH condition. Sci. Rep. 2021, 11, 11504. [Google Scholar] [CrossRef] [PubMed]
- Jiang, F.; Yang, L.; Wang, S.; Ying, X.; Ling, J.; Ouyang, X.-k. Fabrication and characterization of zein-alginate oligosaccharide complex nanoparticles as delivery vehicles of curcumin. J. Mol. Liq. 2021, 342, 116937. [Google Scholar] [CrossRef]
- Stokniene, J.; Powell, L.C.; Aarstad, O.A.; Aachmann, F.L.; Rye, P.D.; Hill, K.E.; Thomas, D.W.; Ferguson, E.L. Bi-functional alginate oligosaccharide-polymyxin conjugates for improved treatment of multidrug-resistant Gram-negative bacterial infections. Pharmaceutics 2020, 12, 1080. [Google Scholar] [CrossRef]
- Wong, T.Y.; Preston, L.A.; Schiller, N.L. Alginate lyase: Review of major sources and enzyme characteristics, structure-function analysis, biological roles, and applications. Annu. Rev. Microbiol. 2000, 54, 289–340. [Google Scholar] [CrossRef] [PubMed]
- Dou, W.; Wei, D.; Li, H.; Li, H.; Rahman, M.M.; Shi, J.; Xu, Z.; Ma, Y. Purification and characterisation of a bifunctional alginate lyase from novel Isoptericola halotolerans CGMCC 5336. Carbohydr. Polym. 2013, 98, 1476–1482. [Google Scholar] [CrossRef]
- Sawant, S.S.; Salunke, B.K.; Kim, B.S. A rapid, sensitive, simple plate assay for detection of microbial alginate lyase activity. Enzym. Microb. Technol. 2015, 77, 8–13. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Chen, L.; Zhang, Z.; Wang, X.; Qin, S.; Yan, P. Screening of alginate lyase-excreting microorganisms from the surface of brown algae. AMB Express 2017, 7, 74. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Hu, F.; Wang, M.; Zhu, B.; Ni, F.; Yao, Z. Elucidation of degradation pattern and immobilization of a novel alginate lyase for preparation of alginate oligosaccharides. Int. J. Biol. Macromol. 2020, 146, 579–587. [Google Scholar] [CrossRef]
- Stender, E.G.P.; Andersen, C.D.; Fredslund, F.; Holck, J.; Solberg, A.; Teze, D.; Peters, G.H.J.; Christensen, B.E.; Aachmann, F.L.; Welner, D.H.; et al. Structural and functional aspects of mannuronic acid-specific PL6 alginate lyase from the human gut microbe Bacteroides cellulosilyticus. J. Biol. Chem. 2019, 294, 17915–17930. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, F.; Wang, P.; Zhang, Y.Z.; Chen, X.L. Diversity of three-dimensional structures and catalytic mechanisms of alginate lyases. Appl. Environ. Microbiol. 2018, 84, e02040-17. [Google Scholar] [CrossRef] [Green Version]
- Kurakake, M.; Kitagawa, Y.; Okazaki, A.; Shimizu, K. Enzymatic properties of alginate lyase from Paenibacillus sp. S29. Appl. Biochem. Biotechnol. 2017, 183, 1455–1464. [Google Scholar] [CrossRef]
- Wantuch, P.L.; Jella, S.; Duke, J.A.; Mousa, J.J.; Henrissat, B.; Glushka, J.; Avci, F.Y. Characterization of the beta-glucuronidase Pn3Pase as the founding member of glycoside hydrolase family GH169. Glycobiology 2021, 31, 266–274. [Google Scholar] [CrossRef]
- Yadav, S.; Pandey, A.K.; Dubey, S.K. Molecular modeling, docking and simulation dynamics of beta-glucosidase reveals high-efficiency, thermo-stable, glucose tolerant enzyme in Paenibacillus lautus BHU3 strain. Int. J. Biol. Macromol. 2021, 168, 371–382. [Google Scholar] [CrossRef]
- Zhang, W.; Ma, J.; Yan, Q.; Jiang, Z.; Yang, S. Biochemical characterization of a novel acidic chitinase with antifungal activity from Paenibacillus xylanexedens Z2–4. Int. J. Biol. Macromol. 2021, 182, 1528–1536. [Google Scholar] [CrossRef] [PubMed]
- Chhe, C.; Uke, A.; Baramee, S.; Tachaapaikoon, C.; Pason, P.; Waeonukul, R.; Ratanakhanokchai, K.; Kosugi, A. Characterization of a thermophilic facultatively anaerobic bacterium Paenibacillus sp. strain DA-C8 that exhibits xylan degradation under anaerobic conditions. J. Biotechnol. 2021, 342, 64–71. [Google Scholar] [CrossRef]
- Zhu, J.; Mo, K.; Zheng, Z.; Wang, Z.; Hu, Y.; Zou, X.; Gu, H.; Wang, H.; Bao, S.; Huang, H. Paenibacillus algicola sp. nov., a novel alginate lyase-producing marine bacterium. Int. J. Syst. Evol. Microbiol. 2020, 70, 5087–5092. [Google Scholar] [CrossRef]
- Itoh, T.; Nakagawa, E.; Yoda, M.; Nakaichi, A.; Hibi, T.; Kimoto, H. Structural and biochemical characterisation of a novel alginate lyase from Paenibacillus sp. str. FPU-7. Sci. Rep. 2019, 9, 14870. [Google Scholar] [CrossRef]
- Yamasaki, M.; Ogura, K.; Hashimoto, W.; Mikami, B.; Murata, K. A structural basis for depolymerization of alginate by polysaccharide lyase family-7. J. Mol. Biol. 2005, 352, 11–21. [Google Scholar] [CrossRef]
- Belik, A.; Silchenko, A.; Malyarenko, O.; Rasin, A.; Kiseleva, M.; Kusaykin, M.; Ermakova, S. Two new alginate lyases of PL7 and PL6 families from polysaccharide-degrading bacterium Formosa algae KMM 3553T: Structure, properties, and products analysis. Mar. Drugs 2020, 18, 130. [Google Scholar] [CrossRef] [Green Version]
- Xu, F.; Dong, F.; Wang, P.; Cao, H.Y.; Li, C.Y.; Li, P.Y.; Pang, X.H.; Zhang, Y.Z.; Chen, X.L. Novel molecular insights into the catalytic mechanism of marine bacterial alginate lyase AlyGC from polysaccharide lyase family 6. J. Biol. Chem. 2017, 292, 4457–4468. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.; Hu, F.; Zhu, B.; Sun, Y.; Yao, Z. Biochemical characterization and elucidation of action pattern of a novel polysaccharide lyase 6 family alginate lyase from marine bacterium Flammeovirga sp. NJ-04. Mar. Drugs 2019, 17, 323. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.I.; Choi, S.H.; Lee, E.Y.; Kim, H.S. Molecular cloning, purification, and characterization of a novel polyMG-specific alginate lyase responsible for alginate MG block degradation in Stenotrophomas maltophilia KJ-2. Appl. Microbiol. Biotechnol. 2012, 95, 1643–1653. [Google Scholar] [CrossRef] [PubMed]
- Mathieu, S.; Henrissat, B.; Labre, F.; Skjak-Braek, G.; Helbert, W. Functional exploration of the polysaccharide lyase family PL6. PLoS ONE 2016, 11, e0159415. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Tan, H.; Qin, Y.; Xu, Q.; Du, Y.; Yin, H. Characterization of a new endo-type alginate lyase from Vibrio sp. W13. Int. J. Biol. Macromol. 2015, 75, 330–337. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Zhou, J.; Li, X.; Peng, Q.; Lu, H.; Du, Y. Characterization of a new alginate lyase from newly isolated Flavobacterium sp. S20. J. Ind. Microbiol. Biotechnol. 2013, 40, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Liu, Z.; Jiang, C.; Li, L.; Xue, C.; Mao, X. Biochemical characterization and degradation pattern analysis of a novel PL-6 alginate lyase from Streptomyces coelicolor A3(2). Food Chem. 2020, 323, 126852. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Q.; Zhang, K.; Shi, Y.; Li, W.; Diao, X.; Liu, W. Structural insights into a novel Ca2+-independent PL-6 alginate lyase from Vibrio OU02 identify the possible subsites responsible for product distribution. Biochim. Biophys. Acta Gen. Subj. 2019, 1863, 1167–1176. [Google Scholar] [CrossRef]
- Zeng, J.; An, D.; Jiao, C.; Xiao, Q.; Weng, H.; Yang, Q.; Xiao, A. Cloning, expression, and characterization of a new pH- and heat-stable alginate lyase from Pseudoalteromonas carrageenovora ASY5. J. Food Biochem. 2019, 43, e12886. [Google Scholar] [CrossRef]
- Li, S.; Wang, L.; Han, F.; Gong, Q.; Yu, W. Cloning and characterization of the first polysaccharide lyase family 6 oligoalginate lyase from marine Shewanella sp. Kz7. J. Biochem. 2016, 159, 77–86. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Wang, L.; Chen, X.; Zhao, W.; Sun, M.; Han, Y. Cloning, expression, and biochemical characterization of two new oligoalginate lyases with synergistic degradation capability. Mar. Biotechnol. 2018, 20, 75–86. [Google Scholar] [CrossRef]
- Wang, M.; Chen, L.; Li, Y.; Chen, L.; Liu, Z.; Wang, X.; Yan, P.; Qin, S. Responses of soil microbial communities to a short-term application of seaweed fertilizer revealed by deep amplicon sequencing. Appl. Soil Ecol. 2018, 125, 288–296. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, Y.H.; Yin, H.; Wang, W.X.; Zhao, X.M.; Du, Y.G. Alginate oligosaccharides enhanced Triticum aestivum L. tolerance to drought stress. Plant Physiol. Biochem. 2013, 62, 33–40. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, H.; Yin, H.; Wang, W.; Zhao, X.; Du, Y. Nitric oxide mediates alginate oligosaccharides-induced root development in wheat (Triticum aestivum L.). Plant Physiol. Biochem. 2013, 71, 49–56. [Google Scholar] [CrossRef]
- Zhang, Y.; Yin, H.; Zhao, X.; Wang, W.; Du, Y.; He, A.; Sun, K. The promoting effects of alginate oligosaccharides on root development in Oryza sativa L. mediated by auxin signaling. Carbohydr. Polym. 2014, 113, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Zhang, L.; Yu, X.; Wang, S.; Xu, C.; Yin, H.; Wang, S. Alginate oligosaccharide attenuates alpha2,6-sialylation modification to inhibit prostate cancer cell growth via the hippo/YAP pathway. Cell Death Dis. 2019, 10, 374. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Feng, Z.; Feng, W.; Hu, T.; Guan, H.; Mao, Y. AOS ameliorates monocrotaline-induced pulmonary hypertension by restraining the activation of P-selectin/p38MAPK/NF-κB pathway in rats. Biomed. Pharmacother. 2019, 109, 1319–1326. [Google Scholar] [CrossRef]
- Huang, W.; Matte, A.; Li, Y.; Kim, Y.S.; Linhardt, R.J.; Su, H.; Cygler, M. Crystal structure of chondroitinase B from Flavo-bacterium heparinum and its complex with a disaccharide product at 1.7 Å resolution. J. Mol. Biol. 1999, 294, 1257–1269. [Google Scholar] [CrossRef] [PubMed]
- Luis, A.S.; Briggs, J.; Zhang, X.; Farnell, B.; Ndeh, D.; Labourel, A.; Baslé, A.; Cartmell, A.; Terrapon, N.; Stott, K.; et al. Dietary pectic glycans are degraded by coordinated enzyme pathways in human colonic Bacteroides. Nat. Microbiol. 2018, 3, 210–219. [Google Scholar] [CrossRef] [PubMed]
- Yoder, M.D.; Lietzke, S.E.; Jurnak, F. Unusual structural features in the parallel β-helix in pectate lyases. Structure 1993, 1, 241–251. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Alginate Lyase | Source | Temperature 1 | pH | NaCl (mM) | Family | Reference |
|---|---|---|---|---|---|---|
| Algb | Vibrio sp. W13 | 30 °C, 20 °C–30 °C | 8.0, 7.0–9.0 | 300, 200–600 | PL-7 | [33] |
| Alg2A | Flavobacterium sp. S20 | 40 °C, 40 °C–50 °C | 8.5, 8.0–9.5 | N.D. 2 | PL-7 | [34] |
| AlyPL6 | Pedobacter hainanensis NJ-02 | 45 °C, 30 °C–50 °C | 9.0, 9.0–10.0 | N.D. | PL-6 | [17] |
| OUC-ScCD6 | Streptomyces ecolicolor A3(2) | 50 °C, 30 °C–60 °C | 9.0, 9.0–10.0 | N.D. | PL-6 | [35] |
| FsAlyPL6 | Flammeovirga sp. NJ-04 | 45 °C, 40 °C–50 °C | 9.0, 8.0–9.0 | N.D. | PL-6 | [29] |
| BcelPL6 | Bacteroides cellulosilyticus CRE21 | 30 °C | 7.5, 7.5–8.0 | 150, 150–250 | PL-6 | [18] |
| AlgF | Vibrio splendidus OU02 | 30 °C, 25 °C–35 °C | 7.5, 7.0–8.0 | 200, 50–200 | PL-6 | [36] |
| Algpt | Paenibacillus sp. LJ-23 | 45 °C, 30 °C–50 °C | 7.0, 6.0–9.0 | 0–700 | PL-6 | This study |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, M.; Chen, L.; Lou, Z.; Yuan, X.; Pan, G.; Ren, X.; Wang, P. Cloning and Characterization of a Novel Alginate Lyase from Paenibacillus sp. LJ-23. Mar. Drugs 2022, 20, 66. https://doi.org/10.3390/md20010066
Wang M, Chen L, Lou Z, Yuan X, Pan G, Ren X, Wang P. Cloning and Characterization of a Novel Alginate Lyase from Paenibacillus sp. LJ-23. Marine Drugs. 2022; 20(1):66. https://doi.org/10.3390/md20010066
Chicago/Turabian StyleWang, Mingpeng, Lei Chen, Zhengyu Lou, Xueting Yuan, Guiping Pan, Xiaoyan Ren, and Pengyu Wang. 2022. "Cloning and Characterization of a Novel Alginate Lyase from Paenibacillus sp. LJ-23" Marine Drugs 20, no. 1: 66. https://doi.org/10.3390/md20010066
APA StyleWang, M., Chen, L., Lou, Z., Yuan, X., Pan, G., Ren, X., & Wang, P. (2022). Cloning and Characterization of a Novel Alginate Lyase from Paenibacillus sp. LJ-23. Marine Drugs, 20(1), 66. https://doi.org/10.3390/md20010066