


Fucoxanthin Production of Microalgae under Different Culture Factors: A Systematic Review

 , , and
, , and
Abstract
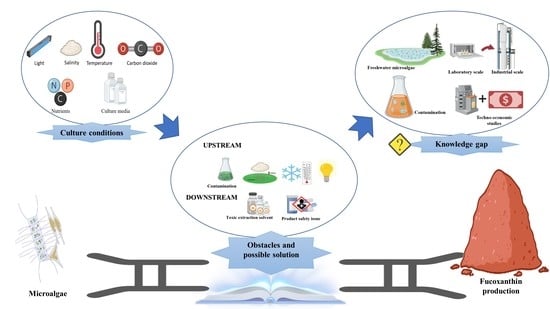
:
1. Introduction
2. Results
3. Discussion
3.1. Microalgae as Fucoxanthin Source
3.2. Effect of Light
3.3. Effect of Nutrients
3.4. Effect of Culture Media
3.5. Effect of Salinity
3.6. Effect of Temperature
3.7. Effect of Carbon Dioxide
3.8. Effect of Oxidative Stress
4. Potential Applications of Fucoxanthin and Current Fucoxanthin-Based Products
5. Obstacles and Possible Solutions
6. Knowledge Gap and Future Directions
7. Materials and Methods
7.1. Literature Search Strategy
7.2. Screening and Eligibility Criteria
7.3. Data Extraction
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Krinsky, N.I.; Johnson, E.J. Carotenoid Actions and Their Relation to Health and Disease. Mol. Aspects Med. 2005, 26, 459–516. [Google Scholar] [CrossRef] [PubMed]
- Muthuirulappan, S.; Francis, S.P. Anti-Cancer Mechanism and Possibility of Nano-Suspension Formulation for a Marine Algae Product Fucoxanthin. Asian Pacific J. Cancer Prev. 2013, 14, 2213–2216. [Google Scholar] [CrossRef] [PubMed]
- Joel, J. Global Fucoxanthin Market 2016 Industry Trends, Sales, Supply, Demand, Analysis and Forecast to 2021; Analysis and Forcast: New York, NY, USA, 2016. [Google Scholar]
- Wang, H.; Zhang, Y.; Chen, L.; Cheng, W.; Liu, T. Combined Production of Fucoxanthin and EPA from Two Diatom Strains Phaeodactylum tricornutum and Cylindrotheca fusiformis Cultures. Bioprocess Biosyst. Eng. 2018, 41, 1061–1071. [Google Scholar] [CrossRef] [PubMed]
- Marella, T.K.; Tiwari, A. Marine Diatom Thalassiosira weissflogii Based Biorefinery for Co-Production of Eicosapentaenoic Acid and Fucoxanthin. Bioresour. Technol. 2020, 307, 123245. [Google Scholar] [CrossRef]
- Khaw, Y.S.; Yusoff, F.M.; Tan, H.T.; Noor Mazli, N.A.I.; Nazarudin, M.F.; Shaharuddin, N.A.; Omar, A.R. The Critical Studies of Fucoxanthin Research Trends from 1928 to June 2021: A Bibliometric Review. Mar. Drugs 2021, 19, 606. [Google Scholar] [CrossRef]
- Wang, S.; Wu, S.; Yang, G.; Pan, K.; Wang, L.; Hu, Z. A Review on the Progress, Challenges and Prospects in Commercializing Microalgal Fucoxanthin. Biotechnol. Adv. 2021, 53, 107865. [Google Scholar] [CrossRef]
- Seth, K.; Kumar, A.; Rastogi, R.P.; Meena, M.; Vinayak, V. Bioprospecting of Fucoxanthin from Diatoms—Challenges and Perspectives. Algal Res. 2021, 60, 102475. [Google Scholar] [CrossRef]
- Pajot, A.; Hao Huynh, G.; Picot, L.; Marchal, L.; Nicolau, E. Fucoxanthin from Algae to Human, an Extraordinary Bioresource: Insights and Advances in up and Downstream Processes. Mar. Drugs 2022, 20, 222. [Google Scholar] [CrossRef]
- Mohibbullah, M.; Haque, M.N.; Sohag, A.A.M.; Hossain, M.T.; Zahan, M.S.; Uddin, M.J.; Hannan, M.A.; Moon, I.S.; Choi, J.-S. A Systematic Review on Marine Algae-Derived Fucoxanthin: An Update of Pharmacological Insights. Mar. Drugs 2022, 20, 279. [Google Scholar] [CrossRef]
- Qiu, S.; Shen, Y.; Wu, Z.; Zhang, X.; Ge, S. Effects of Algae Subtype and Extraction Condition on Extracted Fucoxanthin Antioxidant Property: A 20-Year Meta-Analysis. Algal Res. 2021, 53, 102161. [Google Scholar] [CrossRef]
- Zhang, H.; Gong, P.; Cai, Q.; Zhang, C.; Gao, B. Maximizing Fucoxanthin Production in Odontella aurita by Optimizing the Ratio of Red and Blue Light-Emitting Diodes in an Auto-Controlled Internally Illuminated Photobioreactor. Bioresour. Technol. 2022, 344, 126260. [Google Scholar] [CrossRef]
- Mousavi Nadushan, R.; Hosseinzade, I. Optimization of Production and Antioxidant Activity of Fucoxanthin from Marine Haptophyte Algae, Isochrysis galbana. Iran. J. Fish. Sci. 2020, 19, 2901–2908. [Google Scholar]
- Xia, S.; Gao, B.; Fu, J.; Xiong, J.; Zhang, C. Production of Fucoxanthin, Chrysolaminarin, and Eicosapentaenoic Acid by Odontella aurita under Different Nitrogen Supply Regimes. J. Biosci. Bioeng. 2018, 126, 723–729. [Google Scholar] [CrossRef]
- Mao, X.; Chen, S.H.Y.; Lu, X.; Yu, J.; Liu, B. High Silicate Concentration Facilitates Fucoxanthin and Eicosapentaenoic Acid (EPA) Production under Heterotrophic Condition in the Marine Diatom Nitzschia laevis. Algal Res. 2020, 52, 102086. [Google Scholar] [CrossRef]
- Kanamoto, A.; Kato, Y.; Yoshida, E.; Hasunuma, T.; Kondo, A. Development of a Method for Fucoxanthin Production Using the Haptophyte Marine Microalga Pavlova sp. OPMS 30543. Mar. Biotechnol. 2021, 23, 331–341. [Google Scholar] [CrossRef]
- Butler, T.O.; Padmaperuma, G.; Lizzul, A.M.; McDonald, J.; Vaidyanathan, S. Towards a Phaeodactylum tricornutum Biorefinery in an Outdoor UK Environment. Bioresour. Technol. 2022, 344, 126320. [Google Scholar] [CrossRef]
- Tokushima, H.; Inoue-Kashino, N.; Nakazato, Y.; Masuda, A.; Ifuku, K.; Kashino, Y. Advantageous Characteristics of the Diatom Chaetoceros gracilis as a Sustainable Biofuel Producer. Biotechnol. Biofuels 2016, 9, 235. [Google Scholar] [CrossRef]
- Hao, T.B.; Yang, Y.F.; Balamurugan, S.; Li, D.W.; Yang, W.D.; Li, H.Y. Enrichment of f/2 Medium Hyperaccumulates Biomass and Bioactive Compounds in the Diatom Phaeodactylum tricornutum. Algal Res. 2020, 47, 101872. [Google Scholar] [CrossRef]
- Gérin, S.; Delhez, T.; Corato, A.; Remacle, C.; Franck, F. A Novel Culture Medium for Freshwater Diatoms Promotes Efficient Photoautotrophic Batch Production of Biomass, Fucoxanthin, and Eicosapentaenoic Acid. J. Appl. Phycol. 2020, 32, 1581–1596. [Google Scholar] [CrossRef]
- Erdoğan, A.; Karataş, A.B.; Demirel, Z.; Dalay, M.C. Purification of Fucoxanthin from the Diatom Amphora capitellata by Preparative Chromatography after Its Enhanced Productivity via Oxidative Stress. J. Appl. Phycol. 2022, 34, 301–309. [Google Scholar] [CrossRef]
- Petrushkina, M.; Gusev, E.; Sorokin, B.; Zotko, N.; Mamaeva, A.; Filimonova, A.; Kulikovskiy, M.; Maltsev, Y.; Yampolsky, I.; Guglya, E. Fucoxanthin Production by Heterokont Microalgae. Algal Res. 2017, 24, 387–393. [Google Scholar] [CrossRef]
- Mohamadnia, S.; Tavakoli, O.; Faramarzi, M.A. Enhancing Production of Fucoxanthin by the Optimization of Culture Media of the Microalga Tisochrysis lutea. Aquaculture 2021, 533, 736074. [Google Scholar] [CrossRef]
- Beuzenberg, V.; Goodwin, E.O.; Puddick, J.; Romanazzi, D.; Adams, S.L.; Packer, M.A. Optimising Conditions for Growth and Xanthophyll Production in Continuous Culture of Tisochrysis lutea Using Photobioreactor Arrays and Central Composite Design Experiments. New Zeal. J. Bot. 2017, 55, 64–78. [Google Scholar] [CrossRef]
- Xia, S.; Wang, K.; Wan, L.; Li, A.; Hu, Q.; Zhang, C. Production, Characterization, and Antioxidant Activity of Fucoxanthin from the Marine Diatom Odontella aurita. Mar. Drugs 2013, 11, 2667–2681. [Google Scholar] [CrossRef]
- Premaratne, M.; Liyanaarachchi, V.C.; Nimarshana, P.H.V.; Ariyadasa, T.U.; Malik, A.; Attalage, R.A. Co-Production of Fucoxanthin, Docosahexaenoic Acid (DHA) and Bioethanol from the Marine Microalga Tisochrysis lutea. Biochem. Eng. J. 2021, 176, 108160. [Google Scholar] [CrossRef]
- Yi, Z.; Su, Y.; Cherek, P.; Nelson, D.R.; Lin, J.; Rolfsson, O.; Wu, H.; Salehi-Ashtiani, K.; Brynjolfsson, S.; Fu, W. Combined Artificial High-Silicate Medium and LED Illumination Promote Carotenoid Accumulation in the Marine Diatom Phaeodactylum tricornutum. Microb. Cell Fact. 2019, 18, 209. [Google Scholar] [CrossRef]
- Parkes, R.; Archer, L.; Gee, D.M.; Smyth, T.J.; Gillespie, E.; Touzet, N. Differential Responses in EPA and Fucoxanthin Production by the Marine Diatom Stauroneis sp. under Varying Cultivation Conditions. Biotechnol. Prog. 2021, 37, e3197. [Google Scholar] [CrossRef]
- Ishika, T.; Moheimani, N.R.; Bahri, P.A.; Laird, D.W.; Blair, S.; Parlevliet, D. Halo-Adapted Microalgae for Fucoxanthin Production: Effect of Incremental Increase in Salinity. Algal Res. 2017, 28, 66–73. [Google Scholar] [CrossRef]
- Yang, R.; Wei, D. Improving Fucoxanthin Production in Mixotrophic Culture of Marine Diatom Phaeodactylum tricornutum by LED Light Shift and Nitrogen Supplementation. Front. Bioeng. Biotechnol. 2020, 8, 820. [Google Scholar] [CrossRef]
- Sun, Z.; Wang, X.; Liu, J. Screening of Isochrysis Strains for Simultaneous Production of Docosahexaenoic Acid and Fucoxanthin. Algal Res. 2019, 41, 101545. [Google Scholar] [CrossRef]
- Gao, F.; Teles, I.; Wijffels, R.H.; Barbosa, M.J. Process Optimization of Fucoxanthin Production with Tisochrysis lutea. Bioresour. Technol. 2020, 315, 123894. [Google Scholar] [CrossRef]
- McClure, D.D.; Luiz, A.; Gerber, B.; Barton, G.W.; Kavanagh, J.M. An Investigation into the Effect of Culture Conditions on Fucoxanthin Production Using the Marine Microalgae Phaeodactylum tricornutum. Algal Res. 2018, 29, 41–48. [Google Scholar] [CrossRef]
- Guo, B.; Liu, B.; Yang, B.; Sun, P.; Lu, X.; Liu, J.; Chen, F. Screening of Diatom Strains and Characterization of Cyclotella cryptica as a Potential Fucoxanthin Producer. Mar. Drugs 2016, 14, 125. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Sun, H.; Wu, T.; Fu, Y.; He, Y.; Mao, X. Storage Carbon Metabolism of Isochrysis zhangjiangensis under Different Light Intensities and Its Application for Co-Production of Fucoxanthin and Stearidonic Acid. Bioresour. Technol. 2019, 282, 94–102. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.; Sun, H.; Zhao, W.; Cheng, K.; Chen, F.; Liu, B. A Hetero-Photoautotrophic Two-Stage Cultivation Process for Production of Fucoxanthin by the Marine Diatom Nitzschia laevis. Mar. Drugs 2018, 16, 219. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Woolschot, S.; Cabanelas, I.T.D.; Wijffels, R.H.; Barbosa, M.J. Light Spectra as Triggers for Sorting Improved Strains of Tisochrysis lutea. Bioresour. Technol. 2021, 321, 124434. [Google Scholar] [CrossRef] [PubMed]
- Kanazawa, K.; Ozaki, Y.; Hashimoto, T.; Das, S.K.; Matsushita, S.; Hirano, M.; Okada, T.; Komoto, A.; Mori, N.; Nakatsuka, M. Commercial-Scale Preparation of Biofunctional Fucoxanthin from Waste Parts of Brown Sea Algae Laminalia japonica. Food Sci. Technol. Res. 2008, 14, 573. [Google Scholar] [CrossRef]
- Foo, S.C.; Yusoff, F.M.; Ismail, M.; Basri, M.; Yau, S.K.; Khong, N.M.H.; Chan, K.W.; Ebrahimi, M. Antioxidant Capacities of Fucoxanthin-Producing Algae as Influenced by Their Carotenoid and Phenolic Contents. J. Biotechnol. 2017, 241, 175–183. [Google Scholar] [CrossRef]
- Kim, S.M.; Jung, Y.J.; Kwon, O.N.; Cha, K.H.; Um, B.H.; Chung, D.; Pan, C.H. A Potential Commercial Source of Fucoxanthin Extracted from the Microalga Phaeodactylum tricornutum. Appl. Biochem. Biotechnol. 2012, 166, 1843–1855. [Google Scholar] [CrossRef]
- Kim, S.M.; Kang, S.-W.; Kwon, O.-N.; Chung, D.; Pan, C.-H. Fucoxanthin as a Major Carotenoid in Isochrysis Aff. galbana: Characterization of Extraction for Commercial Application. J. Korean Soc. Appl. Biol. Chem. 2012, 55, 477–483. [Google Scholar] [CrossRef]
- Pasquet, V.; Chérouvrier, J.R.; Farhat, F.; Thiéry, V.; Piot, J.M.; Bérard, J.B.; Kaas, R.; Serive, B.; Patrice, T.; Cadoret, J.P. Study on the Microalgal Pigments Extraction Process: Performance of Microwave Assisted Extraction. Process Biochem. 2011, 46, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Derwenskus, F.; Metz, F.; Gille, A.; Schmid-Staiger, U.; Briviba, K.; Schließmann, U.; Hirth, T. Pressurized Extraction of Unsaturated Fatty Acids and Carotenoids from Wet Chlorella vulgaris and Phaeodactylum tricornutum Biomass Using Subcritical Liquids. Gcb Bioenergy 2019, 11, 335–344. [Google Scholar] [CrossRef]
- Medina, E.; Cerezal, P.; Morales, J.; Ruiz-Domínguez, M.C. Fucoxanthin from Marine Microalga Isochrysis galbana: Optimization of Extraction Methods with Organic Solvents. Dyna 2019, 86, 174–178. [Google Scholar] [CrossRef]
- Khoo, K.S.; Ooi, C.W.; Chew, K.W.; Chia, S.R.; Foo, S.C.; Ng, H.S.; Show, P.L. Extraction of Fucoxanthin from Chaetoceros calcitrans by Electropermeabilization-Assisted Liquid Biphasic Flotation System. J. Chromatogr. A 2022, 1668, 462915. [Google Scholar] [CrossRef]
- Khoo, K.S.; Ooi, C.W.; Chew, K.W.; Foo, S.C.; Show, P.L. Bioprocessing of Chaetoceros calcitrans for the Recovery of Fucoxanthin Using CO2-Based Alkyl Carbamate Ionic Liquids. Bioresour. Technol. 2021, 322, 124520. [Google Scholar] [CrossRef]
- Sun, P.; Wong, C.-C.; Li, Y.; He, Y.; Mao, X.; Wu, T.; Ren, Y.; Chen, F. A Novel Strategy for Isolation and Purification of Fucoxanthinol and Fucoxanthin from the Diatom Nitzschia laevis. Food Chem. 2019, 277, 566–572. [Google Scholar] [CrossRef]
- Gao, F.; Teles, I.; Ferrer-Ledo, N.; Wijffels, R.H.; Barbosa, M.J. Production and High Throughput Quantification of Fucoxanthin and Lipids in Tisochrysis lutea Using Single-Cell Fluorescence. Bioresour. Technol. 2020, 318, 124104. [Google Scholar] [CrossRef]
- Bayu, A.; Rachman, A.; Noerdjito, D.R.; Putra, M.Y.; Widayatno, W.B. High-Value Chemicals from Marine Diatoms: A Biorefinery Approach. In IOP Conference Series: Earth and Environmental Science; IOP Publishing: West Java, Indonesia, 2020; Volume 460, p. 12012. [Google Scholar]
- Jensen, A. The Effect of Seaweed Carotenoids on Egg Yolk Coloration. Poult. Sci. 1963, 42, 912–916. [Google Scholar] [CrossRef]
- Mok, I.K.; Yoon, J.R.; Pan, C.H.; Kim, S.M. Development, Quantification, Method Validation, and Stability Study of a Novel Fucoxanthin-Fortified Milk. J. Agric. Food Chem. 2016, 64, 6196–6202. [Google Scholar] [CrossRef]
- Mok, I.K.; Lee, J.K.; Kim, J.H.; Pan, C.H.; Kim, S.M. Fucoxanthin Bioavailability from Fucoxanthin-Fortified Milk: In Vivo and in Vitro Study. Food Chem. 2018, 258, 79–86. [Google Scholar] [CrossRef]
- Kang, S.Y.; Kang, H.; Lee, J.E.; Jo, C.S.; Moon, C.B.; Ha, J.; Hwang, J.S.; Choi, J. Antiaging Potential of Fucoxanthin Concentrate Derived from Phaeodactylum tricornutum. J. Cosmet. Sci. 2020, 71, 53–64. [Google Scholar]
- Rodríguez-Luna, A.; Ávila-Román, J.; González-Rodríguez, M.L.; Cózar, M.J.; Rabasco, A.M.; Motilva, V.; Talero, E. Fucoxanthin-Containing Cream Prevents Epidermal Hyperplasia and UVB-Induced Skin Erythema in Mice. Mar. Drugs 2018, 16, 378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.J.; Nam, G.W. Sunscreen Boosting Effect by Solid Lipid Nanoparticles-Loaded Fucoxanthin Formulation. Cosmetics 2020, 7, 14. [Google Scholar] [CrossRef]
- Abidov, M.; Ramazanov, Z.; Seifulla, R.; Grachev, S. The Effects of XanthigenTM in the Weight Management of Obese Premenopausal Women with Non-alcoholic Fatty Liver Disease and Normal Liver Fat. Diabetes Obes. Metab. 2010, 12, 72–81. [Google Scholar] [CrossRef]
- Lai, C.S.; Tsai, M.L.; Badmaev, V.; Jimenez, M.; Ho, C.T.; Pan, M.H. Xanthigen Suppresses Preadipocyte Differentiation and Adipogenesis through Down-Regulation of PPARγ and C/EBPs and Modulation of SIRT-1, AMPK, and FoxO Pathways. J. Agric. Food Chem. 2012, 60, 1094–1101. [Google Scholar] [CrossRef]
- Molina, D.; de Carvalho, J.C.; Júnior, A.I.M.; Faulds, C.; Bertrand, E.; Soccol, C.R. Biological Contamination and Its Chemical Control in Microalgal Mass Cultures. Appl. Microbiol. Biotechnol. 2019, 103, 9345–9358. [Google Scholar] [CrossRef]
- Li, W.; Zhang, T.; Tang, X.; Wang, B. Oomycetes and Fungi: Important Parasites on Marine Algae. Acta Oceanol. Sin. 2010, 29, 74–81. [Google Scholar] [CrossRef]
- Huang, Y.; Liu, J.; Wang, H.; Gao, Z. Treatment Potential of a Synergistic Botanical Pesticide Combination for Rotifer Extermination during Outdoor Mass Cultivation of Spirulina platensis. Algal Res. 2014, 6, 139–144. [Google Scholar] [CrossRef]
- He, Q.; Zhang, H.; Ma, M.; He, Y.; Jia, J.; Hu, Q.; Gong, Y. Critical Assessment of Protozoa Contamination and Control Measures in Mass Culture of the Diatom Phaeodactylum tricornutum. Bioresour. Technol. 2022, 359, 127460. [Google Scholar] [CrossRef]
- Zhu, Z.; Jiang, J.; Fa, Y. Overcoming the Biological Contamination in Microalgae and Cyanobacteria Mass Cultivations for Photosynthetic Biofuel Production. Molecules 2020, 25, 5220. [Google Scholar] [CrossRef]
- Ishika, T.; Laird, D.W.; Bahri, P.A.; Moheimani, N.R. Co-Cultivation and Stepwise Cultivation of Chaetoceros muelleri and Amphora sp. for Fucoxanthin Production under Gradual Salinity Increase. J. Appl. Phycol. 2019, 31, 1535–1544. [Google Scholar] [CrossRef]
- Bechet, Q.; Shilton, A.; Fringer, O.B.; Munoz, R.; Guieysse, B. Mechanistic Modeling of Broth Temperature in Outdoor Photobioreactors. Environ. Sci. Technol. 2010, 44, 2197–2203. [Google Scholar] [CrossRef] [PubMed]
- Mata, T.M.; Martins, A.A.; Caetano, N.S. Microalgae for Biodiesel Production and Other Applications: A Review. Renew. Sustain. Energy Rev. 2010, 14, 217–232. [Google Scholar] [CrossRef]
- Ras, M.; Steyer, J.-P.; Bernard, O. Temperature Effect on Microalgae: A Crucial Factor for Outdoor Production. Rev. Environ. Sci. Bio/Technol. 2013, 12, 153–164. [Google Scholar] [CrossRef]
- Derwenskus, F.; Weickert, S.; Lewandowski, I.; Schmid-Staiger, U.; Hirth, T. Economic Evaluation of Up-and Downstream Scenarios for the Co-Production of Fucoxanthin and Eicosapentaenoic Acid with Phaeodactylum tricornutum Using Flat-Panel Airlift Photobioreactors with Artificial Light. Algal Res. 2020, 51, 102078. [Google Scholar] [CrossRef]
- Gao, F.; Cabanelas, I.T.D.; Wijffels, R.H.; Barbosa, M.J. Fucoxanthin and Docosahexaenoic Acid Production by Cold-Adapted Tisochrysis lutea. New Biotechnol. 2022, 66, 16–24. [Google Scholar] [CrossRef]
- Foo, S.C.; Yusoff, F.M.; Ismail, M.; Basri, M.; Khong, N.M.H.; Chan, K.W.; Yau, S.K. Efficient Solvent Extraction of Antioxidant-Rich Extract from a Tropical Diatom, Chaetoceros calcitrans (Paulsen) Takano 1968. Asian Pac. J. Trop. Biomed. 2015, 5, 796–801. [Google Scholar] [CrossRef]
- Delbrut, A.; Albina, P.; Lapierre, T.; Pradelles, R.; Dubreucq, E. Fucoxanthin and Polyunsaturated Fatty Acids Co-Extraction by a Green Process. Molecules 2018, 23, 874. [Google Scholar] [CrossRef]
- Leong, H.Y.; Ooi, C.W.; Law, C.L.; Julkifle, A.L.; Ling, T.C.; Show, P.L. Application of Liquid Biphasic Flotation for Betacyanins Extraction from Peel and Flesh of Hylocereus polyrhizus and Antioxidant Activity Evaluation. Sep. Purif. Technol. 2018, 201, 156–166. [Google Scholar] [CrossRef]
- Grobbelaar, J.U. Quality Control and Assurance: Crucial for the Sustainability of the Applied Phycology Industry. J. Appl. Phycol. 2003, 15, 209–215. [Google Scholar] [CrossRef]
- Mohammad Faisal, H.; Mamoon, R.; Thomas, B.; Mohammed, A.; Winnie, W.; Kolawole Adeshina, K.; Rajjit, S.; Michael, J.; Shamly, A. Evaluation of Fucoxanthin Content in Popular Weight Loss Supplements: The Case for Stricter Regulation of Dietary Supplements. J. Obes. Weight. Medicat. 2019, 5, 31. [Google Scholar] [CrossRef]
- Merlo, S.; Gabarrell Durany, X.; Pedroso Tonon, A.; Rossi, S. Marine Microalgae Contribution to Sustainable Development. Water 2021, 13, 1373. [Google Scholar] [CrossRef]
- Ozkan, A.; Kinney, K.; Katz, L.; Berberoglu, H. Reduction of Water and Energy Requirement of Algae Cultivation Using an Algae Biofilm Photobioreactor. Bioresour. Technol. 2012, 114, 542–548. [Google Scholar] [CrossRef]
- Pugazhendhi, A.; Nagappan, S.; Bhosale, R.R.; Tsai, P.-C.; Natarajan, S.; Devendran, S.; Al-Haj, L.; Ponnusamy, V.K.; Kumar, G. Various Potential Techniques to Reduce the Water Footprint of Microalgal Biomass Production for Biofuel—A Review. Sci. Total Environ. 2020, 749, 142218. [Google Scholar] [CrossRef]
- Wen, X.; Du, K.; Wang, Z.; Peng, X.; Luo, L.; Tao, H.; Xu, Y.; Zhang, D.; Geng, Y.; Li, Y. Effective Cultivation of Microalgae for Biofuel Production: A Pilot-Scale Evaluation of a Novel Oleaginous Microalga Graesiella sp. WBG-1. Biotechnol. Biofuels 2016, 9, 123. [Google Scholar] [CrossRef]
- Gao, F.; Sá, M.; Cabanelas, I.T.D.; Wijffels, R.H.; Barbosa, M.J. Improved Fucoxanthin and Docosahexaenoic Acid Productivities of a Sorted Self-Settling Tisochrysis lutea Phenotype at Pilot Scale. Bioresour. Technol. 2021, 325, 124725. [Google Scholar] [CrossRef]
- Méndez, C.; Uribe, E. Control of Brachionus sp. and Amoeba sp. in Cultures of Arthrospira sp. Lat. Am. J. Aquat. Res. 2012, 40, 553–561. [Google Scholar] [CrossRef]
- Ma, M.; Gong, Y.; Hu, Q. Identification and Feeding Characteristics of the Mixotrophic Flagellate Poterioochromonas malhamensis, a Microalgal Predator Isolated from Outdoor Massive Chlorella Culture. Algal Res. 2018, 29, 142–153. [Google Scholar] [CrossRef]
- Gong, Y.; Patterson, D.J.; Li, Y.; Hu, Z.; Sommerfeld, M.; Chen, Y.; Hu, Q. Vernalophrys algivore Gen. Nov., sp. Nov.(Rhizaria: Cercozoa: Vampyrellida), a New Algal Predator Isolated from Outdoor Mass Culture of Scenedesmus dimorphus. Appl. Environ. Microbiol. 2015, 81, 3900–3913. [Google Scholar] [CrossRef]
- Yuan, D.; Zhan, X.; Wang, M.; Wang, X.; Feng, W.; Gong, Y.; Hu, Q. Biodiversity and Distribution of Microzooplankton in Spirulina (Arthrospira) platensis Mass Cultures throughout China. Algal Res. 2018, 30, 38–49. [Google Scholar] [CrossRef]
- Dryden, R.C.; Wright, S.J.L. Predation of Cyanobacteria by Protozoa. Can. J. Microbiol. 1987, 33, 471–482. [Google Scholar] [CrossRef]
- Banu, J.R.; Kavitha, S.; Gunasekaran, M.; Kumar, G. Microalgae Based Biorefinery Promoting Circular Bioeconomy-Techno Economic and Life-Cycle Analysis. Bioresour. Technol. 2020, 302, 122822. [Google Scholar] [CrossRef]
- Bessette, A.P.; Teymouri, A.; Martin, M.J.; Stuart, B.J.; Resurreccion, E.P.; Kumar, S. Life Cycle Impacts and Techno-Economic Implications of Flash Hydrolysis in Algae Processing. ACS Sustain. Chem. Eng. 2018, 6, 3580–3588. [Google Scholar] [CrossRef]
- Subhash, G.V.; Rajvanshi, M.; Kumar, G.R.K.; Sagaram, U.S.; Prasad, V.; Govindachary, S.; Dasgupta, S. Challenges in Microalgal Biofuel Production: A Perspective on Techno Economic Feasibility under Biorefinery Stratagem. Bioresour. Technol. 2022, 343, 126155. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Microalgae | Type | Factor | Tested Parameter | Highest Bc (g/L) | Optimised Condition for BC | Highest Fx Content (% DW) | Optimised Condition for Fx Content | Reference |
|---|---|---|---|---|---|---|---|---|
| Mallomonas sp. SBV13 | F | Light | 10–226 µmol/m2/s | 3.75 | 226 µmol/m2/s | ≈2.50 | 24 µmol/m2/s | Petrushkina et al. [12] |
| Isochrysis sp. CCMP1324 | M | Light | 30, 60, 120 µmol/m2/s | ≈2.6 | 60 µmol/m2/s | ≈1.30 | 30 µmol/m2/s | Sun et al. [13] |
| Nitrogen | 25, 50, 100 mg/L | ≈2.75 | 100 mg/L | ≈1.35 | 100 mg/L | |||
| Phosphorus | 1.13, 2.25, 4.50 mg/L | ≈2.75 | 4.50 mg/L | ≈1.35 | 4.50 mg/L | |||
| Tisochrysis lutea | M | Light | 50, 150, 300, 500 µmol/m2/s | 1.91 | 300 µmol/m2/s | 0.52 | 50 µmol/m2/s | Gao et al. [14] |
| Temperature | 16.5, 20, 25, 30 °C | 1.81 | 30 °C | ≈0.21 | 25 °C | |||
| Phaeodactylum tricornutum | M | Light | 100, 150, 210 µmol/m2/s | 0.29 | 150 µmol/m2/s | 4.28 | 100 µmol/m2/s | McClure et al. [15] |
| Culture media | f/2, 10 × f/2, f/2 + 10 × nitrate | 0.59 | 10 × f/2 | 5.92 | f/2 + 10 × nitrate | |||
| Carbon dioxide | 0, 1 and 2% | 0.45 | 0% | 2.32 | 1% | |||
| P. tricornutum | M | Light | 30, 70, 120, 180 µmol/m2/s | 1.56 | 70 µmol/m2/s | 0.75 | 30 µmol/m2/s | Wang et al. [4] |
| Nitrogen | 0, 75, 150, 300 mg/L | 1.61 | 300 mg/L | 0.54 | 300 mg/L | |||
| Salinity | 5, 10, 20, 30 ‰ | 1.49 | 20‰ | 0.74 | 20‰ | |||
| Cylindrotheca fusiformis | M | Light | 30, 70, 120, 180 µmol/m2/s | 1.38 | 120 µmol/m2/s | 0.65 | 30 µmol/m2/s | |
| Nitrogen | 0, 75, 150, 300 mg/L | 1.52 | 300 mg/L | 0.61 | 300 mg/L | |||
| Salinity | 5, 10, 20, 30‰ | 1.64 | 30‰ | 0.58 | 10‰ | |||
| Cyclotella cryptica CCMP333 | M | Light | 10, 20, 30, 40 µmol/m2/s | ≈1.25 | 30 µmol/m2/s | 1.08 | 10 µmol/m2/s | Guo et al. [16] |
| Combined | Presence of light and nitrate | 1.72 | Light and nitrate | 1.29 | Light and nitrate | |||
| Isochrysis zhangjiangensis | M | Light | 40, 80, 120, 180, 300 µmol/m2/s | 2.4 | 300 µmol/m2/s | 2.33 | 40 µmol/m2/s | Li et al. [17] |
| Carbon dioxide | 0, 2, 5% | 1.35 | 5% | 2.32 | 5% | |||
| Nitzschia laevis | M | Light | 0, 10, 20, 30, 40, 60 µmol/m2/s | 2.22 | 0 µmol/m2/s | 1.11 | 10 µmol/m2/s | Lu et al. [18] |
| M | Light | BL: WL (0:1, 1:1, 1:0) | NA | NA | 1.20 | BL:WL,1:1 | ||
| T. lutea | M | Light | BL, RL, GL, BL + RL, BL + GL, BL + RL + GL | 0.38 | BL + RL + GL | 1.68 | BL + GL | Gao et al. [19] |
| Odontella aurita | M | Light | RL, BL, WL | 3.87 | RL | 1.52 | RL | Zhang et al. [20] |
| Light | RL:BL (1:9, 2:8, 3:7, 4:6, 5:5, 6:4, 7:3, 8:2, 9:1) | 5.65 | RL:BL, 8:2 | 1.62 | RL:BL, 8:2 | |||
| Isochrysis galbana | M | Nitrogen | 2, 4, 8, 12 mM | a 7 × 106 | 4 mM | 1.81 | 4 mM | Nadushan and Hosseinzade [21] |
| M | Salinity | 25, 35‰ | ≈a 5.9 × 106 | 35‰ | 1.20 | 35‰ | ||
| Thalassiosira weissflogii | M | Nitrogen | 0, 37.5., 75, 150, 300 mg/L | a 3.5 × 106 | 75 mg/L | ≈0.70 | 300 mg/L | Marella and Tiwari [5] |
| Phosphorus | 0, 2.5, 5, 10, 20 mg/L | a 3.6 × 106 | 10 mg/L | ≈0.64 | 10 mg/L | |||
| Silicate | 0, 35, 70, 140, 280 mg/L | ≈a 3.9 × 106 | 280 mg/L | 0.75 | 280 mg/L | |||
| Combined | Light (100,300 µmol/m2/s) and light (BL, RL, WL) | a 9.5 × 106 | 300 µmol/m2/s and BL | ≈1.00 | 100 µmol/m2/s and BL | |||
| O. aurita | M | Nitrogen | ILN, IHN, SN | 5.84 | IHN | 2.33 | SN | Xia et al. [22] |
| Navicula laevis | M | Silicate | 0, 64, 240, 480, 960 mg/L | 2.41 | 480 mg/L | ≈1.40 | 240 mg/L | Mao et al. [23] |
| Pavlova sp. OPMS 30543 | M | Nitrogen | sodium nitrate, potassium nitrate, urea, ammonium chloride | 1.8 | Potassium nitrate | 1.27 | Sodium nitrate | Kanamoto et al. [24] |
| Carbon | glucose, methanol, sodium acetate, sodium bicarbonate | 1.79 | Sodium acetate | 0.73 | Methanol | |||
| Culture media | 2X Daigo IMK, f/2, Walne | 0.92 | 2X Daigo IMK | 0.26 | 2X Daigo IMK | |||
| P. tricornutum | M | Culture media | Cell-Hi F2P, JWP, WP, FloraMicroBloom, f/2 | 0.45 | Cell-Hi F2P | 1.33 | Cell-Hi F2P | Butler et al. [25] |
| Chaetoceros gracilis | M | Nitrogen | Urea, sewage water, liquid fertiliser | NA | NA | 1.95 mg/L | Urea | Tokushima et al. [26] |
| Culture media | Daigo IMK, f/2 | a 5.22 × 106 | Daigo IMK | 2.2 mg/L | Daigo IMK | |||
| P. tricornutum | M | Nitrogen | 0.5 g/L YE, 1 g/L Try, 0.5 g/L YE + 1 g/L Try | ≈0.24 | 0.5 g/L YE + 1 g/L Try | ≈7.5 mg/L | 0.5 g/L YE + 1 g/L Try | Hao et al. [27] |
| Sellaphora minima | F | Culture media | FDMed, Guillard and Lorenzen’s WC, Modified COMBO | 1.71 | FDMed | 0.75 | FDMed | Gerin et al. [28] |
| Nitzchia palea | F | 1.19 | FDMed | 0.55 | FDMed | |||
| Amphora capitellata | M | Oxidative stress | 0.1 mM hydrogen peroxide + 0.1 mM Fe2+, 0.1 mM sodium hypochlorite + 0.1 mM Fe2+, 0.1 mM hydrogen peroxide + 0.1 mM sodium hypochlorite | ≈0.64 | 0.1 mM hydrogen peroxide + 0.1 mM sodium hypochlorite | 4.18 | 0.1 mM hydrogen peroxide + 0.1 mM sodium hypochlorite | Erdogan et al. [29] |
| T. lutea | M | Salinity | 25–45 g/L | a 4.34 × 108 | 36.27 g/L | 7.94 | 36.27 g/L | Mohamadnia et al. [30] |
| Nitrate | 0–0.300 g/L | a 4.34 × 108 | 0.16 g/L | 7.94 | 0.16 g/L | |||
| Glucose | 0.50–6.50 g/L | a 4.34 × 108 | 3.90 g/L | 7.94 | 3.90 g/L | |||
| T. lutea | M | Temperature | 19–35 °C | NA | NA | 0.09 | 25 °C | Beuzenberg et al. [31] |
| Light | 40–1000 µmol/m2/s | NA | NA | 0.09 | 76 µmol/m2/s | |||
| pH | 6.7–8.5 | NA | NA | 0.09 | 7.4 | |||
| O. aurita | M | Combined | Light (100, 300 µmol/m2/s) and nitrate (6, 18 mM) | 6.36 | 300 µmol/m2/s and 18 mM | 2.08 | 100 µmol/m2/s and 18 mM | Xia et al. [32] |
| T. lutea | M | Combined | Light (50, 100, 150 µmol/m2/s) and nitrate (882, 2646 µM) | 0.60 | 150 µmol/m2/s and 2646 µM | 1.51 | 50 µmol/m2/s and 2646 µM | Premaratne et al. [33] |
| P. tricornutum | M | Combined | Light (128, 204, 255 µmol/m2/s) and silicate (0.3, 3.0 mM) | NA | NA | ≈0.75 | 204 µmol/m2/s and 0.3 mM | Yi et al. [34] |
| Stauroneis sp. | M | Combined | Light (WL, BL, RL, GL) and nutrient (f/2 without silicate, f/2 without nitrate, normal seawater, f/2) | NA | NA | 0.59 | BL and f/2 | Parkes et al. [35] |
| Chrysotila carterae | M | Salinity | 35–125‰ | NA | NA | 0.10 | 35‰ | Ishika et al. [36] |
| Chaetoceros muelleri | M | Salinity | 35–125‰ | NA | NA | 0.29 | 45‰ | |
| P. tricornutum | M | Salinity | 35–125‰ | NA | NA | 0.19 | 45‰ | |
| T. lutea | M | Salinity | 35–125‰ | NA | NA | 0.21 | 45‰ | |
| Amphora sp. | M | Salinity | 35–125‰ | NA | NA | 0.15 | 75‰ | |
| Navicula sp. | M | Salinity | 35–125‰ | NA | NA | 0.12 | 85‰ | |
| P. tricornutum | M | Light | 10, 20, 30, 40, 50, 100, 150, 200 µmol/m2/s | 4.80 | 20 µmol/m2/s | 1.60 | 20 µmol/m2/s | Yang and Wei [37] |
| Light | RL:BL (0:1, 6:1, 1:1, 1:2, 1:0) | 5.53 | RL:BL (6:1) | 1.63 | RL:BL (0:1) |
| Microalgae | Fucoxanthin Content (% DW) | Reference |
|---|---|---|
| Chaetoceros calcitrans | 0.51 | Foo et al. [39] |
| Isochrysis galbana | 0.22 | |
| Skeletonema costatum | 0.04 | |
| Odontella sinensis | 0.12 | |
| Phaeodactylum tricornutum | 0.01 | |
| P. tricornutum | 1.57 | Kim et al. [40] |
| P.tricornutum | 0.86 | Kim et al. [41] |
| C. gracilis | 0.22 | |
| I. galbana | 0.60 | |
| Isochrysis aff. galbana | 1.82 | |
| Nitzschia sp. | 0.49 | |
| Cylindrotheca closterium | 0.52 | Pasquet et al. [42] |
| P. tricornutum | 1.86 | Derwenskus et al. [43] |
| I. galbana | 0.63 | Medina et al. [44] |
| Chaetoceros calcitrans | 1.61 | Khoo et al. [45] |
| C. calcitrans | 1.75 | Khoo et al. [46] |
| N. laevis | 0.17 | Sun et al. [47] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khaw, Y.S.; Yusoff, F.M.; Tan, H.T.; Noor Mazli, N.A.I.; Nazarudin, M.F.; Shaharuddin, N.A.; Omar, A.R.; Takahashi, K. Fucoxanthin Production of Microalgae under Different Culture Factors: A Systematic Review. Mar. Drugs 2022, 20, 592. https://doi.org/10.3390/md20100592
Khaw YS, Yusoff FM, Tan HT, Noor Mazli NAI, Nazarudin MF, Shaharuddin NA, Omar AR, Takahashi K. Fucoxanthin Production of Microalgae under Different Culture Factors: A Systematic Review. Marine Drugs. 2022; 20(10):592. https://doi.org/10.3390/md20100592
Chicago/Turabian StyleKhaw, Yam Sim, Fatimah Md Yusoff, Hui Teng Tan, Nur Amirah Izyan Noor Mazli, Muhammad Farhan Nazarudin, Noor Azmi Shaharuddin, Abdul Rahman Omar, and Kazutaka Takahashi. 2022. "Fucoxanthin Production of Microalgae under Different Culture Factors: A Systematic Review" Marine Drugs 20, no. 10: 592. https://doi.org/10.3390/md20100592
APA StyleKhaw, Y. S., Yusoff, F. M., Tan, H. T., Noor Mazli, N. A. I., Nazarudin, M. F., Shaharuddin, N. A., Omar, A. R., & Takahashi, K. (2022). Fucoxanthin Production of Microalgae under Different Culture Factors: A Systematic Review. Marine Drugs, 20(10), 592. https://doi.org/10.3390/md20100592