
Sea Cucumber Saponins Derivatives Alleviate Hepatic Lipid Accumulation Effectively in Fatty Acids-Induced HepG2 Cells and Orotic Acid-Induced Rats

 ,
,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
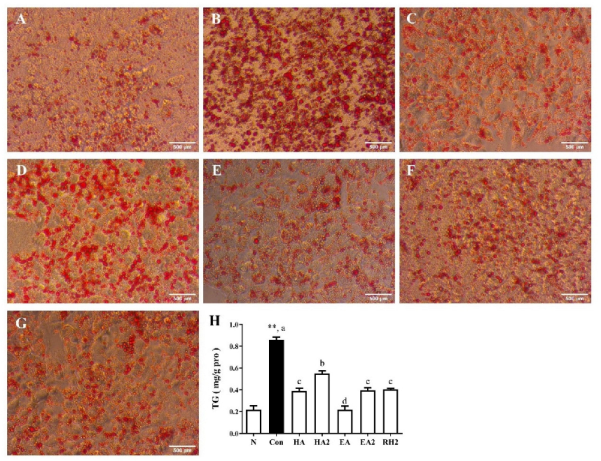
2.1. Effects of Saponins on Lipid Accumulation in HepG2 Cells
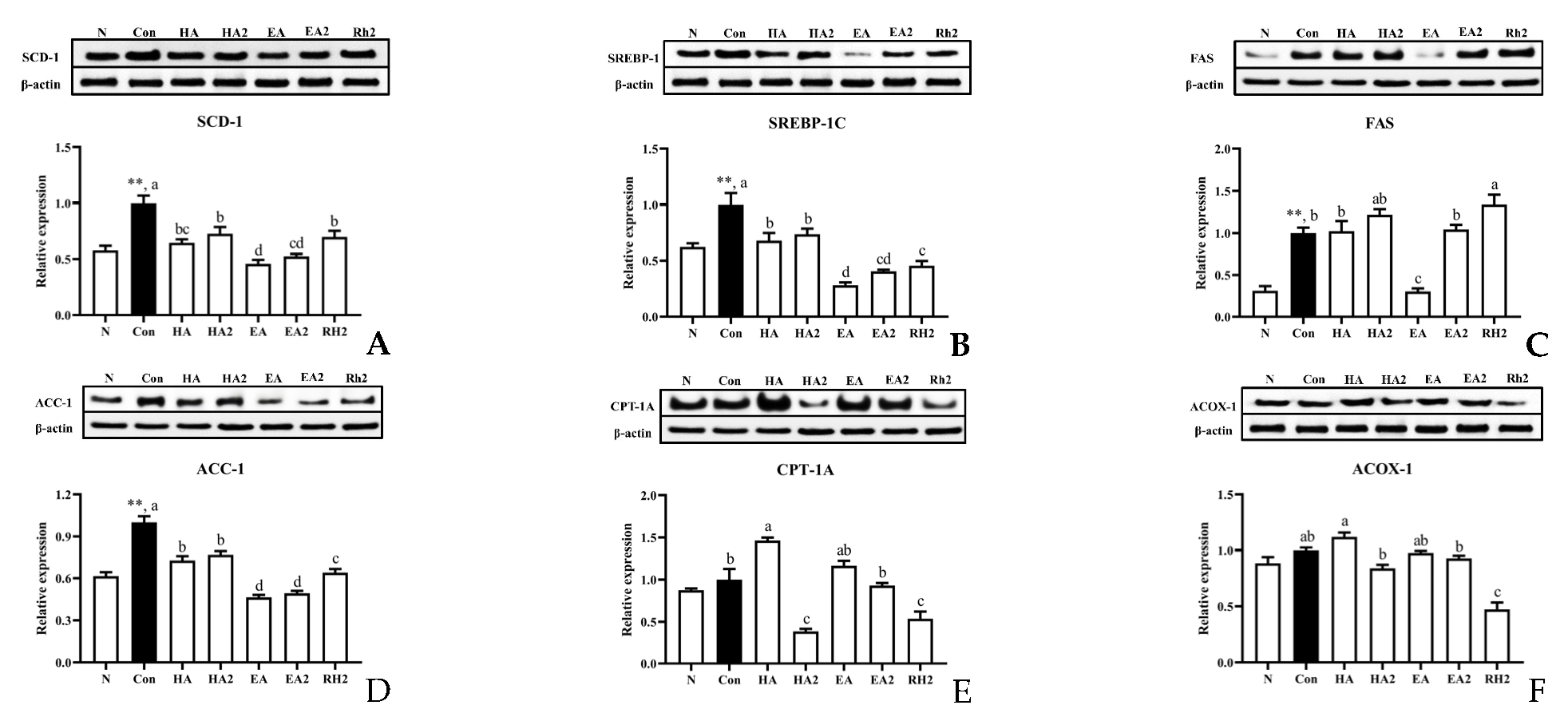
2.2. Effects of Saponins on Lipogenesis and Fatty Acid β-Oxidation in HepG2 Cells
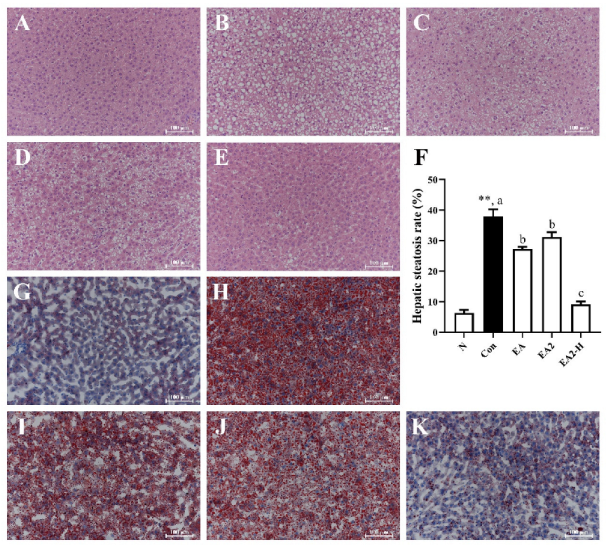
2.3. Effects of EA and EA2 on Liver Histopathological Features of Rats
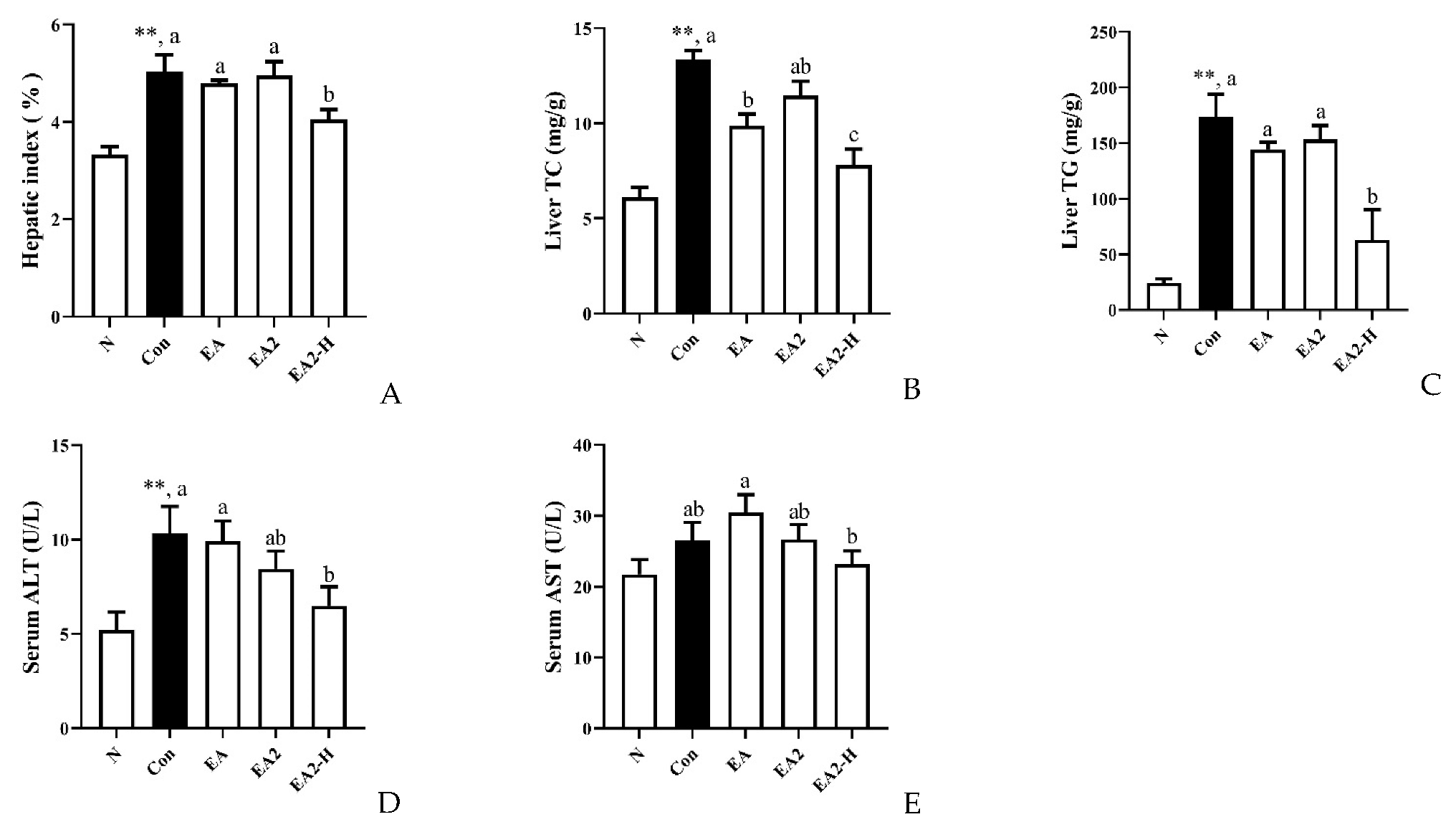
2.4. Effects of EA and EA2 on Lipid Metabolism and Liver Injury in Rats
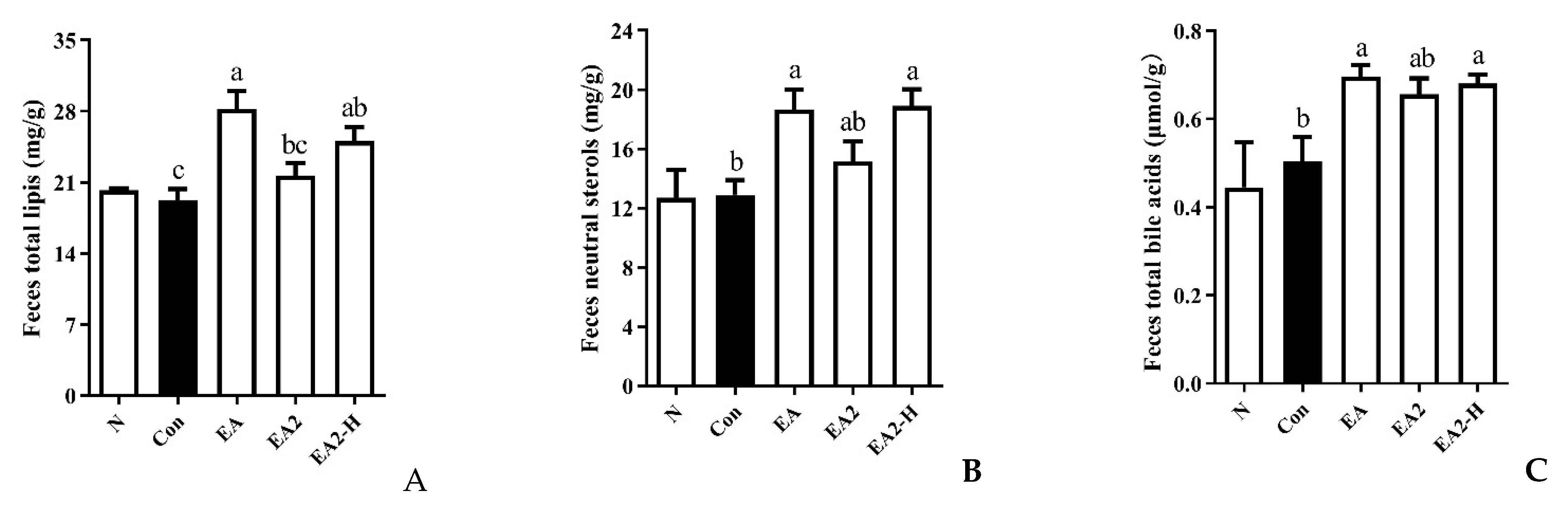
2.5. Effects of EA and EA2 on Lipids Excretion in Rats
2.6. Effects of EA and EA2 on the Expression of Genes Related to Fatty Acids and Cholesterol Metabolism
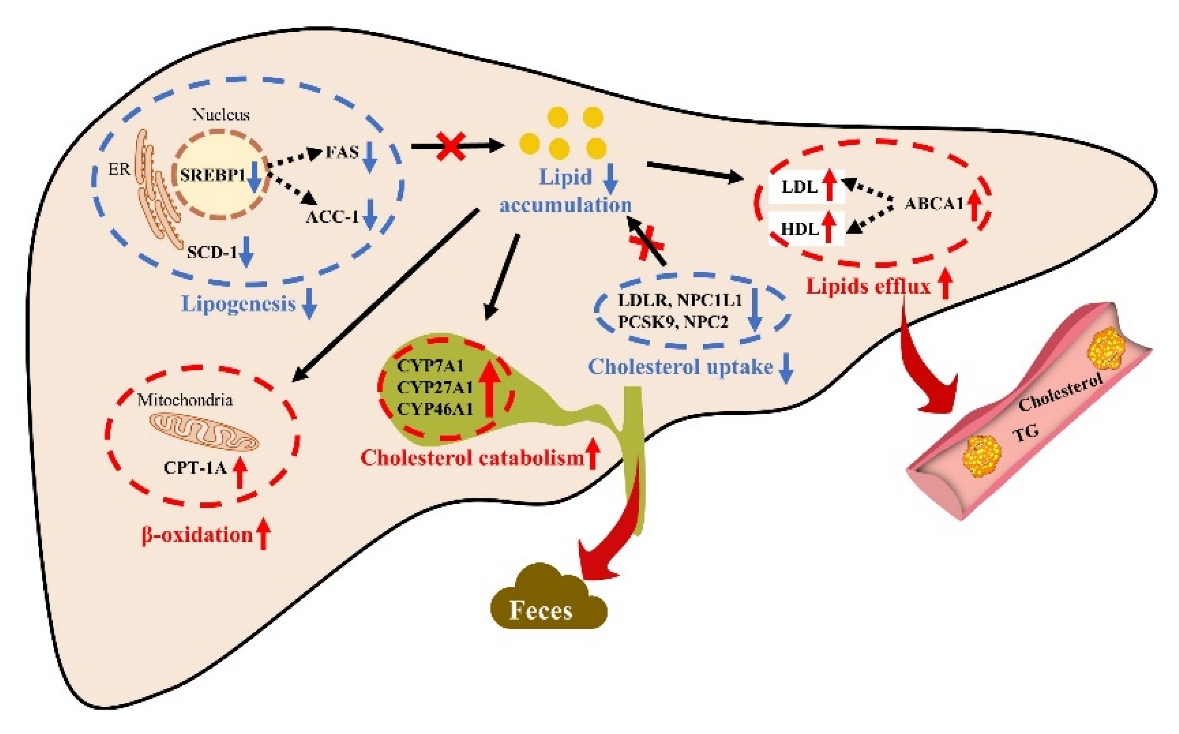
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Preparation of Sea Cucumber Saponins and Their Derivatives
4.3. Experimental Design
4.4. Cell Culture
4.5. Cell Viability Analysis
4.6. Oil Red O Staining
4.7. Western Blotting Analysis
4.8. Animals and treatments
4.9. Histological Analysis
4.10. Lipids Extraction
4.11. Determination of Lipids in Liver and Serum
4.12. Determination of Lipids in Feces
4.13. RNA-Seq Transcriptome Analysis
4.14. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Wang, F.S.; Fan, J.G.; Zhang, Z.; Gao, B.; Wang, H.Y. The global burden of liver disease: The major impact of China. Hepatology 2014, 60, 2099–2108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Byrne, C.D.; Targher, G. NAFLD: A multisystem disease. J. Hepatol. 2015, 62 (Suppl. S1), S47–S64. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, F.; Zhou, J.; Wang, W.; Zhang, X.J.; Ji, Y.X.; Zhang, P.; She, Z.G.; Zhu, L.; Cai, J.; Li, H. Unexpected Rapid Increase in the Burden of NAFLD in China From 2008 to 2018: A Systematic Review and Meta-Analysis. Hepatology 2019, 70, 1119–1133. [Google Scholar] [CrossRef] [PubMed]
- Neuschwander-Tetri, B.A. Therapeutic Landscape for NAFLD in 2020. Gastroenterology 2020, 158, 1984–1998.e3. [Google Scholar] [CrossRef]
- Ding, L.; Xu, Z.J.; Shi, H.H.; Xue, C.H.; Huang, Q.R.; Yanagita, T.; Wang, Y.M.; Zhang, T.T. Sterol sulfate alleviates atherosclerosis via mediating hepatic cholesterol metabolism in ApoE(-/-) mice. Food Funct. 2021, 12, 4887–4896. [Google Scholar] [CrossRef]
- Zhu, Q.; Lin, L.; Zhao, M. Sulfated fucan/fucosylated chondroitin sulfate-dominated polysaccharide fraction from low-edible-value sea cucumber ameliorates type 2 diabetes in rats: New prospects for sea cucumber polysaccharide based-hypoglycemic functional food. Int. J. Biol. Macromol. 2020, 159, 34–45. [Google Scholar] [CrossRef]
- Zhao, Y.C.; Xue, C.H.; Zhang, T.T.; Wang, Y.M. Saponins from Sea Cucumber and Their Biological Activities. J. Agric. Food Chem. 2018, 66, 7222–7237. [Google Scholar] [CrossRef]
- Song, S.; Cong, P.; Xu, J.; Li, G.; Liu, X.; Li, Z.; Xue, C.; Xue, Y.; Wang, Y. Absorption and pharmacokinetic study of two sulphated triterpenoid saponins in rat after oral and intravenous administration of saponin extracts of Pearsonothuria graeffei by HPLC-MS. J. Funct. Foods 2016, 25, 62–69. [Google Scholar] [CrossRef]
- Van Dyck, S.; Gerbaux, P.; Flammang, P. Qualitative and quantitative saponin contents in five sea cucumbers from the Indian ocean. Mar. Drugs 2010, 8, 173–189. [Google Scholar] [CrossRef]
- Meng, J.; Hu, X.; Zhang, T.; Dong, P.; Li, Z.; Xue, C.; Chang, Y.; Wang, Y. Saponin from sea cucumber exhibited more significant effects than ginsenoside on ameliorating high fat diet-induced obesity in C57BL/6 mice. MedChemComm 2018, 9, 725–734. [Google Scholar] [CrossRef]
- Chen, J.L.; Song, S.S.; Sun, S.H.; Li, X.M.; Liu, Y.J.; Li, Z.J.; Xue, C.H. Enzymatic hydrolysis of sea cucumber saponin Echinoside A and identification of its product structure and hemolytic activity. Sci. Technol. Food Ind. 2018, 39, 6. [Google Scholar]
- Soltani, M.; Parivar, K.; Baharara, J.; Kerachian, M.A.; Asili, J. Hemolytic and cytotoxic properties of saponin purified from Holothuria leucospilota sea cucumber. Rep. Biochem. Mol. Biol. 2014, 3, 43–50. [Google Scholar] [PubMed]
- Song, S.; Zhang, L.; Cao, J.; Xiang, G.; Cong, P.; Dong, P.; Li, Z.; Xue, C.; Xue, Y.; Wang, Y. Characterization of Metabolic Pathways and Absorption of Sea Cucumber Saponins, Holothurin A and Echinoside A, In Vitro and In Vivo. J. Food Sci. 2017, 82, 1961–1967. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Virgous, C.; Si, H. How Does Ginsenoside Rh2 Mitigate Adipogenesis in Cultured Cells and Obese Mice? Molecules 2020, 25, 2412. [Google Scholar] [CrossRef]
- Lu, H.; Yuan, X.; Zhang, Y.; Han, M.; Liu, S.; Han, K.; Liang, P.; Cheng, J. HCBP6 deficiency exacerbates glucose and lipid metabolism disorders in non-alcoholic fatty liver mice. Biomed. Pharmacother. 2020, 129, 110347. [Google Scholar] [CrossRef]
- Guo, J.; Fang, W.; Sun, L.; Lu, Y.; Dou, L.; Huang, X.; Tang, W.; Yu, L.; Li, J. Ultraconserved element uc.372 drives hepatic lipid accumulation by suppressing miR-195/miR4668 maturation. Nat. Commun. 2018, 9, 612. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.; Zhang, Y.; Chai, L.; Wang, H. Histological changes, lipid metabolism and oxidative stress in the liver of Bufo gargarizans exposed to cadmium concentrations. Chemosphere 2017, 179, 337–346. [Google Scholar] [CrossRef]
- Bedossa, P.; Poitou, C.; Veyrie, N.; Bouillot, J.L.; Basdevant, A.; Paradis, V.; Tordjman, J.; Clement, K. Histopathological algorithm and scoring system for evaluation of liver lesions in morbidly obese patients. Hepatology 2012, 56, 1751–1759. [Google Scholar] [CrossRef]
- Wang, H.; Hu, L.; Li, L.; Wu, X.; Fan, Z.; Zhang, C.; Wang, J.; Jia, J.; Wang, S. Inorganic nitrate alleviates the senescence-related decline in liver function. Sci. China Life Sci. 2018, 61, 24–34. [Google Scholar] [CrossRef]
- Pottenger, L.A.; Getz, G.S. Serum lipoprotein accumulation in the livers of orotic acid-fed rats. J. Lipid Res. 1971, 12, 450–459. [Google Scholar] [CrossRef]
- Wang, Y.M.; Hu, X.Q.; Xue, Y.; Li, Z.J.; Yanagita, T.; Xue, C.H. Study on possible mechanism of orotic acid-induced fatty liver in rats. Nutrition 2011, 27, 571–575. [Google Scholar] [CrossRef]
- Alaupovic, P. The concept of apolipoprotein-defined lipoprotein families and its clinical significance. Curr. Atheroscler. Rep. 2003, 5, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Schneider, K.M.; Candels, L.S.; Hov, J.R.; Myllys, M.; Hassan, R.; Schneider, C.V.; Wahlström, A.; Mohs, A.; Zühlke, S.; Liao, L.; et al. Gut microbiota depletion exacerbates cholestatic liver injury via loss of FXR signalling. Nat. Metab. 2021, 3, 1228–1241. [Google Scholar] [CrossRef] [PubMed]
- Han, J.; Li, E.; Chen, L.; Zhang, Y.; Wei, F.; Liu, J.; Deng, H.; Wang, Y. The CREB coactivator CRTC2 controls hepatic lipid metabolism by regulating SREBP1. Nature 2015, 524, 243–246. [Google Scholar] [CrossRef] [PubMed]
- Vander Heiden, M.G.; Cantley, L.C.; Thompson, C.B. Understanding the Warburg effect: The metabolic requirements of cell proliferation. Science 2009, 324, 1029–1033. [Google Scholar] [CrossRef] [Green Version]
- Carrella, M.; Björkhem, I.; Gustafsson, J.A.; Einarsson, K.; Hellström, K. The metabolism of steroids in the fatty liver induced by orotic acid feeding. Biochem. J. 1976, 158, 89–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, J.; Yang, H.; Song, B.L. Mechanisms and regulation of cholesterol homeostasis. Nat. Rev. Mol. Cell Biol. 2020, 21, 225–245. [Google Scholar] [CrossRef]
- Hu, X.Q.; Wang, Y.M.; Wang, J.F.; Xue, Y.; Li, Z.J.; Nagao, K.; Yanagita, T.; Xue, C.H. Dietary saponins of sea cucumber alleviate orotic acid-induced fatty liver in rats via PPARalpha and SREBP-1c signaling. Lipids Health Dis. 2010, 9, 25. [Google Scholar] [CrossRef] [Green Version]
- Dong, P.; Xue, C.H.; Du, Q.Z. Separation of two main triterpene glycosides from sea cucumber Pearsonothuria graeffei by high-speed countercurrent chromatography. Acta Chromatogr. 2008, 20, 269–276. [Google Scholar] [CrossRef]
- Kumar, P.; Nagarajan, A.; Uchil, P.D. Analysis of Cell Viability by the MTT Assay. Cold Spring Harb. Protoc. 2018, 6, 469–471. [Google Scholar] [CrossRef]
- Dhami-Shah, H.; Vaidya, R.; Udipi, S.; Raghavan, S.; Abhijit, S.; Mohan, V.; Balasubramanyam, M.; Vaidya, A. Picroside II attenuates fatty acid accumulation in HepG2 cells via modulation of fatty acid uptake and synthesis. Clin. Mol. Hepatol. 2018, 24, 77–87. [Google Scholar] [CrossRef] [PubMed]
- Ricchi, M.; Odoardi, M.R.; Carulli, L.; Anzivino, C.; Ballestri, S.; Pinetti, A.; Fantoni, L.I.; Marra, F.; Bertolotti, M.; Banni, S.; et al. Differential effect of oleic and palmitic acid on lipid accumulation and apoptosis in cultured hepatocytes. J. Gastroenterol. Hepatol. 2009, 24, 830–840. [Google Scholar] [CrossRef]
- Durschlag, R.P.; Robinson, J.L. Orotic acid-induced metabolic changes in the rat. J. Nutr. 1980, 110, 816–821. [Google Scholar] [CrossRef] [PubMed]
- Lou, Q.M.; Wang, Y.M.; Liu, X.F.; Xue, C.H. Lipid profile and fatty acid compositions in body wall of Apostichopus japonicus (selenka). J. Food Biochem. 2012, 36, 317–321. [Google Scholar] [CrossRef]
- Folch, J.; Lees, M.; Sloane Stanley, G.H. A simple method for the isolation and purification of total lipides from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar] [CrossRef]
- Ding, L.; Zhang, L.; Shi, H.; Xue, C.; Yanagita, T.; Zhang, T.; Wang, Y. EPA-enriched ethanolamine plasmalogen alleviates atherosclerosis via mediating bile acids metabolism. J. Funct. Foods 2020, 66, 103824. [Google Scholar] [CrossRef]
- Evrard, E.; Janssen, G. Gas-liquid chromatographic determination of human fecal bile acids. J. Lipid Res. 1968, 9, 226–236. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, X.; Zeng, B.; Wen, L.; Zhao, Y.; Li, Z.; Xue, C.; Zhang, T.; Wang, Y. Sea Cucumber Saponins Derivatives Alleviate Hepatic Lipid Accumulation Effectively in Fatty Acids-Induced HepG2 Cells and Orotic Acid-Induced Rats. Mar. Drugs 2022, 20, 703. https://doi.org/10.3390/md20110703
Li X, Zeng B, Wen L, Zhao Y, Li Z, Xue C, Zhang T, Wang Y. Sea Cucumber Saponins Derivatives Alleviate Hepatic Lipid Accumulation Effectively in Fatty Acids-Induced HepG2 Cells and Orotic Acid-Induced Rats. Marine Drugs. 2022; 20(11):703. https://doi.org/10.3390/md20110703
Chicago/Turabian StyleLi, Xiaoyue, Beibei Zeng, Lu Wen, Yingcai Zhao, Zhaojie Li, Changhu Xue, Tiantian Zhang, and Yuming Wang. 2022. "Sea Cucumber Saponins Derivatives Alleviate Hepatic Lipid Accumulation Effectively in Fatty Acids-Induced HepG2 Cells and Orotic Acid-Induced Rats" Marine Drugs 20, no. 11: 703. https://doi.org/10.3390/md20110703
APA StyleLi, X., Zeng, B., Wen, L., Zhao, Y., Li, Z., Xue, C., Zhang, T., & Wang, Y. (2022). Sea Cucumber Saponins Derivatives Alleviate Hepatic Lipid Accumulation Effectively in Fatty Acids-Induced HepG2 Cells and Orotic Acid-Induced Rats. Marine Drugs, 20(11), 703. https://doi.org/10.3390/md20110703