

Never, Ever Make an Enemy… Out of an Anemone: Transcriptomic Comparison of Clownfish Hosting Sea Anemone Venoms


Abstract
:1. Introduction
2. Results
2.1. Transcriptome Assemblies
2.2. Putative Toxins Inferred from Transcriptomes across Clades
2.2.1. Stichodactylina
2.2.2. Heteractina
2.2.3. Entacmaea
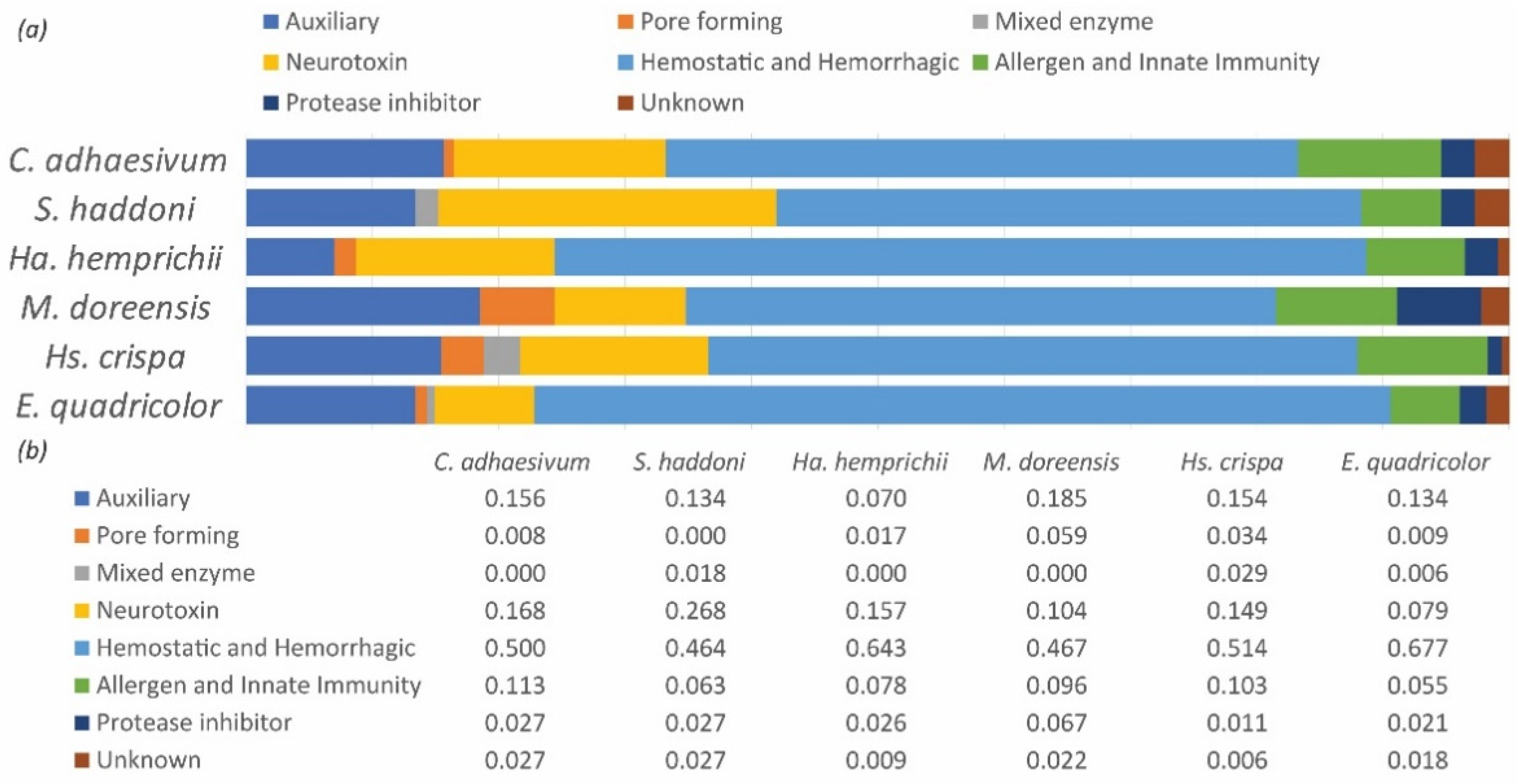
2.3. Diversity of Toxins within Functional Groups
2.3.1. Hemostatic and Hemorrhagic Toxins
2.3.2. Neurotoxins/Protease Inhibitors
2.3.3. Auxiliary Venom
2.3.4. Mixed Enzyme
2.3.5. Pore Forming Toxins
2.3.6. Allergens and Innate Immunity
2.3.7. Venom Transcrits of Unknown Function
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Casewell, N.R.; Wüster, W.; Vonk, F.J.; Harrison, R.A.; Fry, B.G. Complex Cocktails: The Evolutionary Novelty of Venoms. Trends Ecol. Evol. 2013, 28, 219–229. [Google Scholar] [CrossRef]
- Moran, Y.; Genikhovich, G.; Gordon, D.; Wienkoop, S.; Zenkert, C.; Özbek, S.; Technau, U.; Gurevitz, M. Neurotoxin Localization to Ectodermal Gland Cells Uncovers an Alternative Mechanism of Venom Delivery in Sea Anemones. Proc. R. Soc. B Biol. Sci. 2012, 279, 1351–1358. [Google Scholar] [CrossRef] [Green Version]
- Rachamim, T.; Morgenstern, D.; Aharonovich, D.; Brekhman, V.; Lotan, T.; Sher, D. The Dynamically Evolving Nematocyst Content of an Anthozoan, a Scyphozoan, and a Hydrozoan. Mol. Biol. Evol. 2015, 32, 740–753. [Google Scholar] [CrossRef] [Green Version]
- Daly, M. Functional and genetic diversity of toxins in sea anemones. In Evolution of Venomous Animals and Their Toxins; Gopalakrishnakone, P., Malhotra, A., Eds.; Toxinology; Springer: Dordrecht, The Netherlands, 2016; pp. 1–18. ISBN 978-94-007-6727-0. [Google Scholar]
- Fautin, D.G. Structural Diversity, Systematics, and Evolution of Cnidae. Toxicon 2009, 54, 1054–1064. [Google Scholar] [CrossRef] [PubMed]
- Schlesinger, A.; Zlotkin, E.; Kramarsky-Winter, E.; Loya, Y. Cnidarian Internal Stinging Mechanism. Proc. R. Soc. B Biol. Sci. 2009, 276, 1063–1067. [Google Scholar] [CrossRef] [Green Version]
- Nevalainen, T.J.; Peuravuori, H.J.; Quinn, R.J.; Llewellyn, L.E.; Benzie, J.A.H.; Fenner, P.J.; Winkel, K.D. Phospholipase A2 in Cnidaria. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2004, 139, 731–735. [Google Scholar] [CrossRef]
- Jouiaei, M.; Sunagar, K.; Gross, A.F.; Scheib, H.; Alewood, P.F.; Moran, Y.; Fry, B.G. Evolution of an Ancient Venom: Recognition of a Novel Family of Cnidarian Toxins and the Common Evolutionary Origin of Sodium and Potassium Neurotoxins in Sea Anemone. Mol. Biol. Evol. 2015, 32, 1598–1610. [Google Scholar] [CrossRef] [Green Version]
- Suggett, D.J.; Hall-Spencer, J.M.; Rodolfo-Metalpa, R.; Boatman, T.G.; Payton, R.; Tye Pettay, D.; Johnson, V.R.; Warner, M.E.; Lawson, T. Sea Anemones May Thrive in a High CO2 World. Glob. Change Biol. 2012, 18, 3015–3025. [Google Scholar] [CrossRef] [PubMed]
- Daly, M.; Rack, F.; Zook, R. Edwardsiella andrillae, a New Species of Sea Anemone from Antarctic Ice. PLoS ONE 2013, 8, e83476. [Google Scholar] [CrossRef] [Green Version]
- Menezes, C.; Thakur, N.L. Sea Anemone Venom: Ecological Interactions and Bioactive Potential. Toxicon 2022, 208, 31–46. [Google Scholar] [CrossRef]
- Sachkova, M.Y.; Macrander, J.; Surm, J.M.; Aharoni, R.; Menard-Harvey, S.S.; Klock, A.; Leach, W.B.; Reitzel, A.M.; Moran, Y. Some like It Hot: Population-Specific Adaptations in Venom Production to Abiotic Stressors in a Widely Distributed Cnidarian. BMC Biol. 2020, 18, 121. [Google Scholar] [CrossRef]
- LaJeunesse, T.C.; Parkinson, J.E.; Gabrielson, P.W.; Jeong, H.J.; Reimer, J.D.; Voolstra, C.R.; Santos, S.R. Systematic Revision of Symbiodiniaceae Highlights the Antiquity and Diversity of Coral Endosymbionts. Curr. Biol. 2018, 28, 2570–2580. [Google Scholar] [CrossRef] [Green Version]
- Goffredi, S.K.; Motooka, C.; Fike, D.A.; Gusmão, L.C.; Tilic, E.; Rouse, G.W.; Rodríguez, E. Mixotrophic Chemosynthesis in a Deep-Sea Anemone from Hydrothermal Vents in the Pescadero Basin, Gulf of California. BMC Biol. 2021, 19, 8. [Google Scholar] [CrossRef]
- Williams, J.D.; McDermott, J.J. Hermit Crab Biocoenoses: A Worldwide Review of the Diversity and Natural History of Hermit Crab Associates. J. Exp. Mar. Biol. Ecol. 2004, 305, 1–128. [Google Scholar] [CrossRef]
- Mercier, A.; Hamel, J.-F. Nature and Role of Newly Described Symbiotic Associations between a Sea Anemone and Gastropods at Bathyal Depths in the NW Atlantic. J. Exp. Mar. Biol. Ecol. 2008, 358, 57–69. [Google Scholar] [CrossRef]
- Sanamyan, N.P.; Sanamyan, K.E.; Tabachnik, K.R. The First Species of Actiniaria, Spongiactis japonica gen.n., sp.n. (Cnidaria: Anthozoa), an Obligate Symbiont of a Glass Sponge. Invertebr. Zool. 2012, 9, 127–141. [Google Scholar] [CrossRef]
- Izumi, T.; Ise, Y.; Yanagi, K.; Shibata, D.; Ueshima, R. First Detailed Record of Symbiosis Between a Sea Anemone and Homoscleromorph Sponge, With a Description of Tempuractis rinkai Gen. et Sp. Nov. (Cnidaria: Anthozoa: Actiniaria: Edwardsiidae). Zoolog. Sci. 2018, 35, 188–198. [Google Scholar] [CrossRef] [Green Version]
- Vader, W.; Tandberg, A.H.S. Amphipods and Sea Anemones, an Update. J. Crustac. Biol. 2020, 40, 872–878. [Google Scholar] [CrossRef]
- Fautin, D.G. The Anemonefish Symbiosis: What Is Known and What Is Not. Symbiosis 1991, 10, 23–26. [Google Scholar]
- Dunn, D.F. The Clownfish Sea Anemones: Stichodactylidae (Coelenterata: Actiniaria) and Other Sea Anemones Symbiotic with Pomacentrid Fishes. Trans. Am. Philos. Soc. 1981, 71, 3–115. [Google Scholar] [CrossRef]
- Godwin, J.; Fautin, D.G. Defense of Host Actinians by Anemonefishes. Copeia 1992, 1992, 902–908. [Google Scholar] [CrossRef]
- Holbrook, S.J.; Schmitt, R.J. Growth, Reproduction and Survival of a Tropical Sea Anemone (Actiniaria): Benefits of Hosting Anemonefish. Coral Reefs 2005, 24, 67–73. [Google Scholar] [CrossRef]
- Porat, D.; Chadwick-Furman, N.E. Effects of Anemonefish on Giant Sea Anemones: Ammonium Uptake, Zooxanthella Content and Tissue Regeneration. Mar. Freshw. Behav. Physiol. 2005, 38, 43–51. [Google Scholar] [CrossRef]
- Alan Verde, E.; Cleveland, A.; Lee, R.W. Nutritional Exchange in a Tropical Tripartite Symbiosis II: Direct Evidence for the Transfer of Nutrients from Host Anemone and Zooxanthellae to Anemonefish. Mar. Biol. 2015, 162, 2409–2429. [Google Scholar] [CrossRef]
- Litsios, G.; Sims, C.A.; Wüest, R.O.; Pearman, P.B.; Zimmermann, N.E.; Salamin, N. Mutualism with Sea Anemones Triggered the Adaptive Radiation of Clownfishes. BMC Evol. Biol. 2012, 12, 212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCord, C.L.; Nash, C.M.; Cooper, W.J.; Westneat, M.W. Phylogeny of the Damselfishes (Pomacentridae) and Patterns of Asymmetrical Diversification in Body Size and Feeding Ecology. PLoS ONE 2021, 16, e0258889. [Google Scholar] [CrossRef] [PubMed]
- Frédérich, B.; Sorenson, L.; Santini, F.; Slater, G.J.; Alfaro, M.E. Iterative Ecological Radiation and Convergence during the Evolutionary History of Damselfishes (Pomacentridae). Am. Nat. 2013, 181, 94–113. [Google Scholar] [CrossRef]
- Elliott, J.K.; Lougheed, S.C.; Bateman, B.; McPhee, L.K.; Boag, P.T. Molecular Phylogenetic Evidence for the Evolution of Specialization in Anemonefishes. Proc. R. Soc. Lond. B Biol. Sci. 1999, 266, 677–685. [Google Scholar] [CrossRef] [Green Version]
- Titus, B.M.; Benedict, C.; Laroche, R.; Gusmão, L.C.; Van Deusen, V.; Chiodo, T.; Meyer, C.P.; Berumen, M.L.; Bartholomew, A.; Yanagi, K.; et al. Phylogenetic Relationships among the Clownfish-Hosting Sea Anemones. Mol. Phylogenet. Evol. 2019, 139, 106526. [Google Scholar] [CrossRef]
- Lubbock, R. The Clownfish/Anemone Symbiosis: A Problem of Cellular Recognition. Parasitology 1981, 82, 159–173. [Google Scholar] [CrossRef]
- Allen, G.R. Anemonefishes and Their Amazing Partnership. Aust. Nat. Hist. 1975, 18, 274–277. [Google Scholar]
- Mebs, D. Chemical Biology of the Mutualistic Relationships of Sea Anemones with Fish and Crustaceans. Toxicon 2009, 54, 1071–1074. [Google Scholar] [CrossRef]
- Mebs, D. Anemonefish Symbiosis: Vulnerability and Resistance of Fish to the Toxin of the Sea Anemone. Toxicon 1994, 32, 1059–1068. [Google Scholar] [CrossRef] [PubMed]
- Elliott, J.K.; Mariscal, R.N. Coexistence of Nine Anemonefish Species: Differential Host and Habitat Utilization, Size and Recruitment. Mar. Biol. 2001, 138, 23–36. [Google Scholar] [CrossRef]
- Lubbock, R.; Smith, D.C. Why Are Clownfishes Not Stung by Sea Anemones? Proc. R. Soc. Lond. B Biol. Sci. 1980, 207, 35–61. [Google Scholar] [CrossRef]
- Mariscal, R.N. The Nature of the Symbiosis between Indo-Pacific Anemone Fishes and Sea Anemones. Mar. Biol. 1970, 6, 58–65. [Google Scholar] [CrossRef]
- Davenport, D.; Norris, K.S. Observations on the Symbiosis of the Sea Anemone Stoichactis and the Pomacentrid Fish, Amphiprion percula. Biol. Bull. 1958, 115, 397–410. [Google Scholar] [CrossRef]
- Elliott, J.K.; Mariscal, R.N.; Roux, K.H. Do Anemonefishes Use Molecular Mimicry to Avoid Being Stung by Host Anemones? J. Exp. Mar. Biol. Ecol. 1994, 179, 99–113. [Google Scholar] [CrossRef]
- Elliott, J.K.; Mariscal, R.N. Acclimation or Innate Protection of Anemonefishes from Sea Anemones? Copeia 1997, 1997, 284–289. [Google Scholar] [CrossRef]
- Marcionetti, A.; Rossier, V.; Roux, N.; Salis, P.; Laudet, V.; Salamin, N. Insights into the Genomics of Clownfish Adaptive Radiation: Genetic Basis of the Mutualism with Sea Anemones. Genome Biol. Evol. 2019, 11, 869–882. [Google Scholar] [CrossRef] [Green Version]
- Nedosyko, A.M.; Young, J.E.; Edwards, J.W.; Silva, K.B. da Searching for a Toxic Key to Unlock the Mystery of Anemonefish and Anemone Symbiosis. PLoS ONE 2014, 9, e98449. [Google Scholar] [CrossRef]
- Norton, R.S.; Pennington, M.W.; Wulff, H. Potassium Channel Blockade by the Sea Anemone Toxin ShK for the Treatment of Multiple Sclerosis and Other Autoimmune Diseases. Curr. Med. Chem. 2004, 11, 3041–3052. [Google Scholar] [CrossRef]
- Haas, B.J.; Papanicolaou, A.; Yassour, M.; Grabherr, M.; Blood, P.D.; Bowden, J.; Couger, M.B.; Eccles, D.; Li, B.; Lieber, M.; et al. De Novo Transcript Sequence Reconstruction from RNA-Seq: Reference Generation and Analysis with Trinity. Nat. Protoc. 2013, 8, 1494–1512. [Google Scholar] [CrossRef]
- Ashwood, L.M.; Mitchell, M.L.; Madio, B.; Hurwood, D.A.; King, G.F.; Undheim, E.A.B.; Norton, R.S.; Prentis, P.J. Tentacle Morphological Variation Coincides with Differential Expression of Toxins in Sea Anemones. Toxins 2021, 13, 452. [Google Scholar] [CrossRef] [PubMed]
- Madio, B.; Undheim, E.A.B.; King, G.F. Revisiting Venom of the Sea Anemone Stichodactyla haddoni: Omics Techniques Reveal the Complete Toxin Arsenal of a Well-Studied Sea Anemone Genus. J. Proteom. 2017, 166, 83–92. [Google Scholar] [CrossRef] [PubMed]
- Macrander, J.; Broe, M.; Daly, M. Tissue-Specific Venom Composition and Differential Gene Expression in Sea Anemones. Genome Biol. Evol. 2016, 8, 2358–2375. [Google Scholar] [CrossRef] [Green Version]
- Fry, B.G.; Roelants, K.; Champagne, D.E.; Scheib, H.; Tyndall, J.D.A.; King, G.F.; Nevalainen, T.J.; Norman, J.A.; Lewis, R.J.; Norton, R.S.; et al. The Toxicogenomic Multiverse: Convergent Recruitment of Proteins Into Animal Venoms. Annu. Rev. Genom. Hum. Genet. 2009, 10, 483–511. [Google Scholar] [CrossRef] [Green Version]
- Huang, C.; Morlighem, J.-É.R.; Zhou, H.; Lima, É.P.; Gomes, P.B.; Cai, J.; Lou, I.; Pérez, C.D.; Lee, S.M.; Rádis-Baptista, G. The Transcriptome of the Zoanthid Protopalythoa variabilis (Cnidaria, Anthozoa) Predicts a Basal Repertoire of Toxin-like and Venom-Auxiliary Polypeptides. Genome Biol. Evol. 2016, 8, 3045–3064. [Google Scholar] [CrossRef] [Green Version]
- Ponce, D.; Brinkman, D.L.; Potriquet, J.; Mulvenna, J. Tentacle Transcriptome and Venom Proteome of the Pacific Sea Nettle, Chrysaora fuscescens (Cnidaria: Scyphozoa). Toxins 2016, 8, 102. [Google Scholar] [CrossRef] [Green Version]
- Liao, Q.; Feng, Y.; Yang, B.; Lee, S.M.-Y. Cnidarian Peptide Neurotoxins: A New Source of Various Ion Channel Modulators or Blockers against Central Nervous Systems Disease. Drug Discov. Today 2019, 24, 189–197. [Google Scholar] [CrossRef]
- Li, R.; Yu, H.; Xue, W.; Yue, Y.; Liu, S.; Xing, R.; Li, P. Jellyfish Venomics and Venom Gland Transcriptomics Analysis of Stomolophus meleagris to Reveal the Toxins Associated with Sting. J. Proteom. 2014, 106, 17–29. [Google Scholar] [CrossRef]
- OmPraba, G.; Chapeaurouge, A.; Doley, R.; Devi, K.R.; Padmanaban, P.; Venkatraman, C.; Velmurugan, D.; Lin, Q.; Kini, R.M. Identification of a Novel Family of Snake Venom Proteins Veficolins from Cerberus rynchops Using a Venom Gland Transcriptomics and Proteomics Approach. J. Proteome Res. 2010, 9, 1882–1893. [Google Scholar] [CrossRef]
- Choo, Y.M.; Lee, K.S.; Yoon, H.J.; Kim, B.Y.; Sohn, M.R.; Roh, J.Y.; Je, Y.H.; Kim, N.J.; Kim, I.; Woo, S.D.; et al. Dual Function of a Bee Venom Serine Protease: Prophenoloxidase-Activating Factor in Arthropods and Fibrin(Ogen)Olytic Enzyme in Mammals. PLoS ONE 2010, 5, e10393. [Google Scholar] [CrossRef]
- Choo, Y.M.; Lee, K.S.; Yoon, H.J.; Qiu, Y.; Wan, H.; Sohn, M.R.; Sohn, H.D.; Jin, B.R. Antifibrinolytic Role of a Bee Venom Serine Protease Inhibitor That Acts as a Plasmin Inhibitor. PLoS ONE 2012, 7, e32269. [Google Scholar] [CrossRef] [Green Version]
- Zhong, S.-R.; Jin, Y.; Wu, J.-B.; Chen, R.-Q.; Jia, Y.-H.; Wang, W.-Y.; Xiong, Y.-L.; Zhang, Y. Characterization and Molecular Cloning of Dabocetin, a Potent Antiplatelet C-Type Lectin-like Protein from Daboia russellii siamensis Venom. Toxicon 2006, 47, 104–112. [Google Scholar] [CrossRef]
- Kita, M.; Nakamura, Y.; Okumura, Y.; Ohdachi, S.D.; Oba, Y.; Yoshikuni, M.; Kido, H.; Uemura, D. Blarina Toxin, a Mammalian Lethal Venom from the Short-Tailed Shrew Blarina brevicauda: Isolation and Characterization. Proc. Natl. Acad. Sci. USA 2004, 101, 7542–7547. [Google Scholar] [CrossRef] [Green Version]
- Rokyta, D.R.; Lemmon, A.R.; Margres, M.J.; Aronow, K. The Venom-Gland Transcriptome of the Eastern Diamondback Rattlesnake (Crotalus adamanteus). BMC Genom. 2012, 13, 312. [Google Scholar] [CrossRef] [Green Version]
- Lopes-Ferreira, M.; Magalhães, G.S.; Fernandez, J.H.; Junqueira-de-Azevedo, I.d.L.M.; Le Ho, P.; Lima, C.; Valente, R.H.; Moura-da-Silva, A.M. Structural and Biological Characterization of Nattectin, a New C-Type Lectin from the Venomous Fish Thalassophryne nattereri. Biochimie 2011, 93, 971–980. [Google Scholar] [CrossRef] [Green Version]
- Junqueira-de-Azevedo, I.d.L.M.; Ho, P.L. A Survey of Gene Expression and Diversity in the Venom Glands of the Pitviper Snake Bothrops insularis through the Generation of Expressed Sequence Tags (ESTs). Gene 2002, 299, 279–291. [Google Scholar] [CrossRef]
- Magalhães, G.S.; Lopes-Ferreira, M.; Junqueira-de-Azevedo, I.d.L.M.; Spencer, P.J.; Araújo, M.S.; Portaro, F.C.V.; Ma, L.; Valente, R.H.; Juliano, L.; Fox, J.W.; et al. Natterins, a New Class of Proteins with Kininogenase Activity Characterized from Thalassophryne nattereri Fish Venom. Biochimie 2005, 87, 687–699. [Google Scholar] [CrossRef]
- Prentis, P.J.; Pavasovic, A.; Norton, R.S. Sea Anemones: Quiet Achievers in the Field of Peptide Toxins. Toxins 2018, 10, 36. [Google Scholar] [CrossRef] [Green Version]
- Bosmans, F.; Tytgat, J. Sea Anemone Venom as a Source of Insecticidal Peptides Acting on Voltage-Gated Na+ Channels. Toxicon 2007, 49, 550–560. [Google Scholar] [CrossRef]
- Upadhyay, S.K.; Eckel-Mahan, K.L.; Mirbolooki, M.R.; Tjong, I.; Griffey, S.M.; Schmunk, G.; Koehne, A.; Halbout, B.; Iadonato, S.; Pedersen, B.; et al. Selective Kv1.3 Channel Blocker as Therapeutic for Obesity and Insulin Resistance. Proc. Natl. Acad. Sci. USA 2013, 110, E2239–E2248. [Google Scholar] [CrossRef] [Green Version]
- Yan, F.; Cheng, X.; Ding, X.; Yao, T.; Chen, H.; Li, W.; Hu, S.; Yu, Z.; Sun, Y.; Zhang, Y.; et al. Improved Insecticidal Toxicity by Fusing Cry1Ac of Bacillus Thuringiensis with Av3 of Anemonia viridis. Curr. Microbiol. 2014, 68, 604–609. [Google Scholar] [CrossRef]
- Chi, V.; Pennington, M.W.; Norton, R.S.; Tarcha, E.J.; Londono, L.M.; Sims-Fahey, B.; Upadhyay, S.K.; Lakey, J.T.; Iadonato, S.; Wulff, H.; et al. Development of a Sea Anemone Toxin as an Immunomodulator for Therapy of Autoimmune Diseases. Toxicon 2012, 59, 529–546. [Google Scholar] [CrossRef] [Green Version]
- Watkins, M.; Hillyard, D.R.; Olivera, B.M. Genes Expressed in a Turrid Venom Duct: Divergence and Similarity to Conotoxins. J. Mol. Evol. 2006, 62, 247–256. [Google Scholar] [CrossRef]
- Ward, M.J.; Rokyta, D.R. Venom-Gland Transcriptomics and Venom Proteomics of the Giant Florida Blue Centipede, Scolopendra viridis. Toxicon 2018, 152, 121–136. [Google Scholar] [CrossRef]
- Undheim, E.A.B.; Sunagar, K.; Herzig, V.; Kely, L.; Low, D.H.W.; Jackson, T.N.W.; Jones, A.; Kurniawan, N.; King, G.F.; Ali, S.A.; et al. A Proteomics and Transcriptomics Investigation of the Venom from the Barychelid Spider Trittame loki (Brush-Foot Trapdoor). Toxins 2013, 5, 2488–2503. [Google Scholar] [CrossRef] [Green Version]
- Moran, Y.; Gurevitz, M. When Positive Selection of Neurotoxin Genes Is Missing. FEBS J. 2006, 273, 3886–3892. [Google Scholar] [CrossRef]
- Baconguis, I.; Gouaux, E. Structural Plasticity and Dynamic Selectivity of Acid-Sensing Ion Channel–Spider Toxin Complexes. Nature 2012, 489, 400–405. [Google Scholar] [CrossRef] [Green Version]
- Laustsen, A.H. Toxin Synergism in Snake Venoms. Toxin Rev. 2016, 35, 165–170. [Google Scholar] [CrossRef] [Green Version]
- Moran, Y.; Praher, D.; Schlesinger, A.; Ayalon, A.; Tal, Y.; Technau, U. Analysis of Soluble Protein Contents from the Nematocysts of a Model Sea Anemone Sheds Light on Venom Evolution. Mar. Biotechnol. 2013, 15, 329–339. [Google Scholar] [CrossRef]
- Baumgarten, S.; Simakov, O.; Esherick, L.Y.; Liew, Y.J.; Lehnert, E.M.; Michell, C.T.; Li, Y.; Hambleton, E.A.; Guse, A.; Oates, M.E.; et al. The Genome of Aiptasia, a Sea Anemone Model for Coral Symbiosis. Proc. Natl. Acad. Sci. USA 2015, 112, 11893–11898. [Google Scholar] [CrossRef] [Green Version]
- Ruiming, Z.; Yibao, M.; Yawen, H.; Zhiyong, D.; Yingliang, W.; Zhijian, C.; Wenxin, L. Comparative Venom Gland Transcriptome Analysis of the Scorpion Lychas mucronatus Reveals Intraspecific Toxic Gene Diversity and New Venomous Components. BMC Genom. 2010, 11, 452. [Google Scholar] [CrossRef] [Green Version]
- Dodds, D.; Schlimgen, A.K.; Lu, S.-Y.; Perin, M.S. Novel Reticular Calcium Binding Protein Is Purified on Taipoxin Columns. J. Neurochem. 1995, 64, 2339–2344. [Google Scholar] [CrossRef]
- Six, D.A.; Dennis, E.A. The Expanding Superfamily of Phospholipase A2 Enzymes: Classification and Characterization. Biochim. Biophys. Acta BBA—Mol. Cell Biol. Lipids 2000, 1488, 1–19. [Google Scholar] [CrossRef]
- Podobnik, M.; Anderluh, G. Pore-Forming Toxins in Cnidaria. Semin. Cell Dev. Biol. 2017, 72, 133–141. [Google Scholar] [CrossRef]
- Parker, M.W.; Feil, S.C. Pore-Forming Protein Toxins: From Structure to Function. Prog. Biophys. Mol. Biol. 2005, 88, 91–142. [Google Scholar] [CrossRef]
- Anderluh, G.; Maček, P. Dissecting the Actinoporin Pore-Forming Mechanism. Structure 2003, 11, 1312–1313. [Google Scholar] [CrossRef] [Green Version]
- Anderluh, G.; Kisovec, M.; Kraševec, N.; Gilbert, R.J.C. Distribution of MACPF/CDC proteins. In MACPF/CDC Proteins—Agents of Defence, Attack and Invasion; Anderluh, G., Gilbert, R., Eds.; Subcellular Biochemistry; Springer: Dordrecht, The Netherlands, 2014; pp. 7–30. ISBN 978-94-017-8881-6. [Google Scholar]
- Tibballs, J.; Yanagihara, A.A.; Turner, H.C.; Winkel, K. Immunological and Toxinological Responses to Jellyfish Stings. Inflamm. Allergy—Drug Targets Former. Curr. Drug Targets—Inflamm. Allergy 2011, 10, 438–446. [Google Scholar] [CrossRef] [Green Version]
- Horiike, T.; Nagai, H.; Kitani, S. Identification of Allergens in the Box Jellyfish Chironex yamaguchii That Cause Sting Dermatitis. Int. Arch. Allergy Immunol. 2015, 167, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Tibballs, J. Australian chirodropid cubozoan jellyfish envenomation. In Clinical Toxinology; Gopalakrishnakone, P., Faiz, S.M.A., Gnanathasan, C.A., Habib, A.G., Fernando, R., Yang, C.-C., Vogel, C.-W., Tambourgi, D.V., Seifert, S.A., Eds.; Toxinology; Springer: Dordrecht, The Netherlands, 2017; pp. 1–19. ISBN 978-94-007-6288-6. [Google Scholar]
- Wang, Y.-M.; Wang, S.-R.; Tsai, I.-H. Serine Protease Isoforms of Deinagkistrodon acutus Venom: Cloning, Sequencing and Phylogenetic Analysis. Biochem. J. 2001, 354, 161–168. [Google Scholar] [CrossRef]
- Weinstock, G.M.; Robinson, G.E.; Gibbs, R.A.; Weinstock, G.M.; Weinstock, G.M.; Robinson, G.E.; Worley, K.C.; Evans, J.D.; Maleszka, R.; Robertson, H.M.; et al. Insights into Social Insects from the Genome of the Honeybee Apis mellifera. Nature 2006, 443, 931–949. [Google Scholar] [CrossRef] [Green Version]
- Bernheimer, A.W.; Weinstein, S.A.; Linder, R. Isoelectric Analysis of Some Australian Elapid Snake Venoms with Special Reference to Phospholipase B and Hemolysis. Toxicon 1986, 24, 841–849. [Google Scholar] [CrossRef]
- Sabourault, C.; Ganot, P.; Deleury, E.; Allemand, D.; Furla, P. Comprehensive EST Analysis of the Symbiotic Sea Anemone, Anemonia Viridis. BMC Genom. 2009, 10, 333. [Google Scholar] [CrossRef] [Green Version]
- Kozlov, S.; Grishin, E. The Mining of Toxin-like Polypeptides from EST Database by Single Residue Distribution Analysis. BMC Genom. 2011, 12, 88. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, J.S.; Fuentes-Silva, D.; King, G.F. Development of a Rational Nomenclature for Naming Peptide and Protein Toxins from Sea Anemones. Toxicon 2012, 60, 539–550. [Google Scholar] [CrossRef]
- Hines, D.E.; Pawlik, J.R. Assessing the Antipredatory Defensive Strategies of Caribbean Non-Scleractinian Zoantharians (Cnidaria): Is the Sting the Only Thing? Mar. Biol. 2012, 159, 389–398. [Google Scholar] [CrossRef]
- Ashwood, L.M.; Norton, R.S.; Undheim, E.A.B.; Hurwood, D.A.; Prentis, P.J. Characterising Functional Venom Profiles of Anthozoans and Medusozoans within Their Ecological Context. Mar. Drugs 2020, 18, 202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jaimes-Becerra, A.; Gacesa, R.; Doonan, L.B.; Hartigan, A.; Marques, A.C.; Okamura, B.; Long, P.F. “Beyond Primary Sequence”—Proteomic Data Reveal Complex Toxins in Cnidarian Venoms. Integr. Comp. Biol. 2019, 59, 777–785. [Google Scholar] [CrossRef]
- Schendel, V.; Rash, L.D.; Jenner, R.A.; Undheim, E.A.B. The Diversity of Venom: The Importance of Behavior and Venom System Morphology in Understanding Its Ecology and Evolution. Toxins 2019, 11, 666. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pennington, M.W.; Czerwinski, A.; Norton, R.S. Peptide Therapeutics from Venom: Current Status and Potential. Bioorg. Med. Chem. 2018, 26, 2738–2758. [Google Scholar] [CrossRef]
- Tarcha, E.J.; Chi, V.; Muñoz-Elías, E.J.; Bailey, D.; Londono, L.M.; Upadhyay, S.K.; Norton, K.; Banks, A.; Tjong, I.; Nguyen, H.; et al. Durable Pharmacological Responses from the Peptide ShK-186, a Specific Kv1.3 Channel Inhibitor That Suppresses T Cell Mediators of Autoimmune Disease. J. Pharmacol. Exp. Ther. 2012, 342, 642–653. [Google Scholar] [CrossRef] [Green Version]
- Tarcha, E.J.; Olsen, C.M.; Probst, P.; Peckham, D.; Muñoz-Elías, E.J.; Kruger, J.G.; Iadonato, S.P. Safety and Pharmacodynamics of Dalazatide, a Kv1.3 Channel Inhibitor, in the Treatment of Plaque Psoriasis: A Randomized Phase 1b Trial. PLoS ONE 2017, 12, e0180762. [Google Scholar] [CrossRef] [PubMed]
- Klompen, A.M.L.; Macrander, J.; Reitzel, A.M.; Stampar, S.N. Transcriptomic Analysis of Four Cerianthid (Cnidaria, Ceriantharia) Venoms. Mar. Drugs 2020, 18, 413. [Google Scholar] [CrossRef]
- Li, R.; Yu, H.; Yue, Y.; Liu, S.; Xing, R.; Chen, X.; Li, P. Combined Proteomics and Transcriptomics Identifies Sting-Related Toxins of Jellyfish Cyanea nozakii. J. Proteom. 2016, 148, 57–64. [Google Scholar] [CrossRef]
- Brinkman, D.L.; Jia, X.; Potriquet, J.; Kumar, D.; Dash, D.; Kvaskoff, D.; Mulvenna, J. Transcriptome and Venom Proteome of the Box Jellyfish Chironex fleckeri. BMC Genom. 2015, 16, 407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, J.; He, Y.; Peng, C.; Tang, T.; Jin, A.; Liao, Y.; Shi, Q.; Gao, B. Transcriptome Sequencing of the Pale Anemones (Exaiptasia diaphana) Revealed Functional Peptide Gene Resources of Sea Anemone. Front. Mar. Sci. 2022, 9, 856501. [Google Scholar] [CrossRef]
- Mitchell, M.L.; Hossain, M.A.; Lin, F.; Pinheiro-Junior, E.L.; Peigneur, S.; Wai, D.C.C.; Delaine, C.; Blyth, A.J.; Forbes, B.E.; Tytgat, J.; et al. Identification, Synthesis, Conformation and Activity of an Insulin-like Peptide from a Sea Anemone. Biomolecules 2021, 11, 1785. [Google Scholar] [CrossRef]
- Krishnarjuna, B.; Sunanda, P.; Villegas–Moreno, J.; Csoti, A.; Morales, R.A.V.; Wai, D.C.C.; Panyi, G.; Prentis, P.; Norton, R.S. A Disulfide-Stabilised Helical Hairpin Fold in Acrorhagin I: An Emerging Structural Motif in Peptide Toxins. J. Struct. Biol. 2021, 213, 107692. [Google Scholar] [CrossRef]
- Honma, T.; Minagawa, S.; Nagai, H.; Ishida, M.; Nagashima, Y.; Shiomi, K. Novel Peptide Toxins from Acrorhagi, Aggressive Organs of the Sea Anemone Actinia equina. Toxicon 2005, 46, 768–774. [Google Scholar] [CrossRef]
- Macrander, J.; Brugler, M.R.; Daly, M. A RNA-Seq Approach to Identify Putative Toxins from Acrorhagi in Aggressive and Non-Aggressive Anthopleura elegantissima Polyps. BMC Genom. 2015, 16, 221. [Google Scholar] [CrossRef]
- Minagawa, S.; Sugiyama, M.; Ishida, M.; Nagashima, Y.; Shiomi, K. Kunitz-Type Protease Inhibitors from Acrorhagi of Three Species of Sea Anemones. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2008, 150, 240–245. [Google Scholar] [CrossRef] [PubMed]
- Columbus-Shenkar, Y.Y.; Sachkova, M.Y.; Macrander, J.; Fridrich, A.; Modepalli, V.; Reitzel, A.M.; Sunagar, K.; Moran, Y. Dynamics of Venom Composition across a Complex Life Cycle. eLife 2018, 7, e35014. [Google Scholar] [CrossRef] [PubMed]
- Leach, W.B.; Reitzel, A.M. Transcriptional Remodelling upon Light Removal in a Model Cnidarian: Losses and Gains in Gene Expression. Mol. Ecol. 2019, 28, 3413–3426. [Google Scholar] [CrossRef]
- Sachkova, M.Y.; Singer, S.A.; Macrander, J.; Reitzel, A.M.; Peigneur, S.; Tytgat, J.; Moran, Y. The Birth and Death of Toxins with Distinct Functions: A Case Study in the Sea Anemone Nematostella. Mol. Biol. Evol. 2019, 36, 2001–2012. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.G.; Surm, J.M.; Macrander, J.; Simhi, A.; Amir, G.; Sachkova, M.Y.; Lewandowska, M.; Reitzel, A.M.; Moran, Y. Dominant Toxin Hypothesis: Unravelling the Venom Phenotype across Micro and Macroevolution. bioRxiv 2022. [Google Scholar] [CrossRef]
- Verdes, A.; Taboada, S.; Hamilton, B.R.; Undheim, E.A.B.; Sonoda, G.G.; Andrade, S.C.S.; Morato, E.; Marina, A.I.; Cárdenas, C.A.; Riesgo, A. Evolution, Expression Patterns, and Distribution of Novel Ribbon Worm Predatory and Defensive Toxins. Mol. Biol. Evol. 2022, 39, msac096. [Google Scholar] [CrossRef]
- Ramírez-Carreto, S.; Vera-Estrella, R.; Portillo-Bobadilla, T.; Licea-Navarro, A.; Bernaldez-Sarabia, J.; Rudiño-Piñera, E.; Verleyen, J.J.; Rodríguez, E.; Rodríguez-Almazán, C. Transcriptomic and Proteomic Analysis of the Tentacles and Mucus of Anthopleura dowii Verrill, 1869. Mar. Drugs 2019, 17, 436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holding, M.L.; Margres, M.J.; Mason, A.J.; Parkinson, C.L.; Rokyta, D.R. Evaluating the Performance of De Novo Assembly Methods for Venom-Gland Transcriptomics. Toxins 2018, 10, 249. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-Length Transcriptome Assembly from RNA-Seq Data without a Reference Genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simão, F.A.; Waterhouse, R.M.; Ioannidis, P.; Kriventseva, E.V.; Zdobnov, E.M. BUSCO: Assessing Genome Assembly and Annotation Completeness with Single-Copy Orthologs. Bioinformatics 2015, 31, 3210–3212. [Google Scholar] [CrossRef]
- Boratyn, G.M.; Camacho, C.; Cooper, P.S.; Coulouris, G.; Fong, A.; Ma, N.; Madden, T.L.; Matten, W.T.; McGinnis, S.D.; Merezhuk, Y.; et al. BLAST: A More Efficient Report with Usability Improvements. Nucleic Acids Res. 2013, 41, W29–W33. [Google Scholar] [CrossRef] [Green Version]
- Finn, R.D.; Clements, J.; Arndt, W.; Miller, B.L.; Wheeler, T.J.; Schreiber, F.; Bateman, A.; Eddy, S.R. HMMER Web Server: 2015 Update. Nucleic Acids Res. 2015, 43, W30–W38. [Google Scholar] [CrossRef]
- Armenteros, J.J.A.; Salvatore, M.; Emanuelsson, O.; Winther, O.; von Heijne, G.; Elofsson, A.; Nielsen, H. Detecting Sequence Signals in Targeting Peptides Using Deep Learning. Life Sci. Alliance 2019, 2, e201900429. [Google Scholar] [CrossRef] [Green Version]
- Fu, L.; Niu, B.; Zhu, Z.; Wu, S.; Li, W. CD-HIT: Accelerated for Clustering the next-Generation Sequencing Data. Bioinformatics 2012, 28, 3150–3152. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple Sequence Alignment with High Accuracy and High Throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Madeira, F.; Pearce, M.; Tivey, A.R.N.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and Sequence Analysis Tools Services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef] [PubMed]
- The UniProt Consortium UniProt: A Worldwide Hub of Protein Knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [CrossRef]


{kind=link}
| Clade | Taxon [Source] | Transcripts | Genes | N50 | BUSCO |
|---|---|---|---|---|---|
| Stichodactylina | C. adhaesivum [45] | 905,882 (628,469) | 484,240 (451,132) | 1308 (609) | 96.2 (93.9) |
| S. haddoni [46] | 196,433 (1,800,000) | 150,054 (269,628) | 1038 (-) | 83.6 (-) | |
| Ha. hemprichii [45] | 600,056 (101,150) | 165,714 (74,496) | 1055 (1370) | 94.5 (92) | |
| Heteractina | Hs. crispa [47] | 655,116 (-) | 581,957 (-) | 545 (-) | 91.8 (-) |
| M. doreensis [45] | 624,291 (-) | 218,128 (-) | 1328 (-) | 91.4 (-) | |
| Entacmaea | E. quadricolor [PRJEB21970] | 1,198,081 (-) | 296,968 (-) | 1088 (-) | 98.3 (-) |
| T | TF | AT | ATF | |
|---|---|---|---|---|
| C. adhaesivum | 256 | 34 | 88 | 13 |
| S. haddoni | 112 | 33 | 46 | 12 |
| Ha. hemprichii | 115 | 32 | 27 | 13 |
| Stychodactylina | 483 | 47 | 161 | 18 |
| M. doreensis | 135 | 25 | 56 | 12 |
| Hs. crispa | 175 | 35 | 59 | 12 |
| Heteractina | 310 | 37 | 115 | 13 |
| Entacmaea quadricolor (Entacmaea) | 328 | 37 | 79 | 13 |
| Total | 1121 | 51 | 355 | 18 |
| Venom Category | Species | Venom ID | CA | SH | HH | MD | HC | EQ |
|---|---|---|---|---|---|---|---|---|
| Auxiliary | sea anemone | Astacin-like metalloprotease toxin M12A | 15 | 5 | 2 | 15 | 13 | 22 |
| Nematocyst expressed protein 6 | 24 | 8 | 4 | 9 | 13 | 21 | ||
| snake | Cystatin-2 | - | 2 | - | 1 | 1 | 1 | |
| scorpion | Venom protein 302 | 1 | - | - | - | - | - | |
| Venom protein 59.1 | - | 1 | - | - | - | - | ||
| Pore Forming | sea anemone | Actinoporin family | 1 | - | 1 | 1 | 5 | 2 |
| MAC/PF | 1 | - | 1 | 7 | 1 | 1 | ||
| Neurotoxin | sea anemone | Cnidaria small cysteine-rich protein (SCRiP) family | - | - | 1 | - | - | 1 |
| Cysteine-rich venom protein | 8 | - | - | 2 | 5 | - | ||
| Neurotoxin ShK-like1 | - | - | 1 | - | - | - | ||
| Sea anemone short toxin (type III) family (Delta-actitoxin-Avd2b1) | - | 1 | 1 | - | - | - | ||
| Sea anemone structural class 9a | 1 | 1 | - | - | - | 3 | ||
| Sea anemone type 1 potassium channel toxin subfamily | 4 | 3 | - | 1 | 1 | 1 | ||
| Sea anemone type 3 potassium channel toxin subfamily (BDS) | 7 | 8 | 6 | 1 | 5 | 4 | ||
| sodium channel toxin Type I | - | 3 | - | 2 | 2 | - | ||
| sodium channel toxin Type II | 3 | 2 | 6 | - | - | - | ||
| sea snail | Conopeptide P-like superfamily | 10 | 2 | 3 | 2 | 5 | 8 | |
| spider | Omega-theraphotoxin-Pm1a, Neurotoxin 10 | - | 1 | - | - | - | - | |
| Psalmotoxin-1 (PcTx1) | - | 1 | - | - | - | - | ||
| centipede | Scoloptoxin | - | 1 | - | - | 2 | - | |
| Mixed Enzyme | sea anemone | Phospholipase A2 | - | 2 | - | - | 5 | 2 |
| Protease Inhibitor | sea anemone | Venom Kunitz-type family | 7 | 3 | 3 | 9 | 2 | 7 |
| Allergen and Innate Immunity | snake | A. superbus venom factor 1 | 2 | - | 2 | - | 1 | 3 |
| snake venom serine protease Dav-PA | 3 | - | - | - | - | - | ||
| Snake venom serine protease salmobin | - | - | - | 2 | 1 | - | ||
| Venom serine carboxypeptidase | 8 | 2 | - | 6 | 7 | 5 | ||
| spider | Techylectin-like protein | - | - | - | - | - | 1 | |
| ant | Venom allergen 5 | 2 | 3 | 1 | 3 | 6 | 5 | |
| moth | Venom serine protease | 7 | 1 | 3 | 2 | 1 | - | |
| bee | Venom serine protease Bi-VSP | 7 | 1 | 3 | 2 | 1 | 3 | |
| Hemostatic and Hemorrhagic Toxin | shrew | Blarina toxin | 13 | 1 | 4 | - | 3 | 9 |
| snake | Coagulation factor V | - | - | 1 | - | 3 | 1 | |
| Coagulation factor X-activating enzyme | 4 | 1 | - | - | 3 | 4 | ||
| Cobra venom factor | - | 1 | 2 | - | - | 1 | ||
| Ryncolin | 14 | 8 | 13 | 3 | 3 | 35 | ||
| Snaclec family | 7 | 7 | 1 | 10 | 13 | 6 | ||
| True venom lectin family | 4 | 3 | - | 5 | 3 | 26 | ||
| Veficolin-1 | 17 | 9 | 14 | 9 | 10 | 45 | ||
| Venom factor (VF) | 3 | - | 4 | - | - | 1 | ||
| Venom phosphodiesterase | - | - | - | 1 | - | |||
| Venom prothrombin activator | 25 | 5 | 15 | 8 | 32 | 48 | ||
| Zinc metalloproteinase/disintegrin-like (M12B) | 7 | 2 | 6 | 6 | 3 | 9 | ||
| fish | Galactose-specific lectin nattectin | 8 | 6 | - | 6 | 1 | 4 | |
| Natterin-4 | - | - | - | - | 4 | 1 | ||
| lizard | Venom protease | 6 | 4 | 5 | 11 | 4 | 12 | |
| spider | Venom peptide isomerase heavy chain | 17 | 5 | 6 | 3 | 7 | 15 | |
| unknown | sea anemone | EGF domain peptide family | 6 | 3 | 1 | 2 | 1 | 5 |
| Sea anemone 8 toxin family | 1 | - | - | 1 | - | 1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delgado, A.; Benedict, C.; Macrander, J.; Daly, M. Never, Ever Make an Enemy… Out of an Anemone: Transcriptomic Comparison of Clownfish Hosting Sea Anemone Venoms. Mar. Drugs 2022, 20, 730. https://doi.org/10.3390/md20120730
Delgado A, Benedict C, Macrander J, Daly M. Never, Ever Make an Enemy… Out of an Anemone: Transcriptomic Comparison of Clownfish Hosting Sea Anemone Venoms. Marine Drugs. 2022; 20(12):730. https://doi.org/10.3390/md20120730
Chicago/Turabian StyleDelgado, Alonso, Charlotte Benedict, Jason Macrander, and Marymegan Daly. 2022. "Never, Ever Make an Enemy… Out of an Anemone: Transcriptomic Comparison of Clownfish Hosting Sea Anemone Venoms" Marine Drugs 20, no. 12: 730. https://doi.org/10.3390/md20120730
APA StyleDelgado, A., Benedict, C., Macrander, J., & Daly, M. (2022). Never, Ever Make an Enemy… Out of an Anemone: Transcriptomic Comparison of Clownfish Hosting Sea Anemone Venoms. Marine Drugs, 20(12), 730. https://doi.org/10.3390/md20120730