
Anti-Larval and Anti-Algal Natural Products from Marine Microorganisms as Sources of Anti-Biofilm Agents

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Anti-Biofilm and Antifouling Natural Products from Marine Bacteria
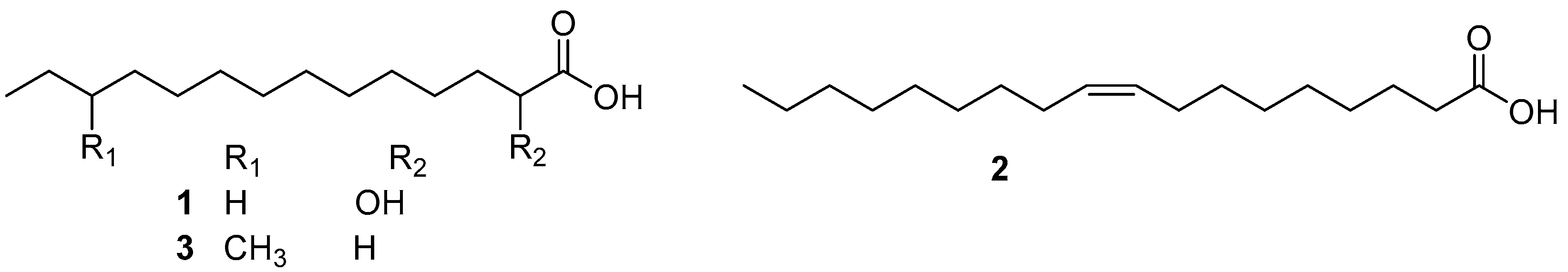
2.1. Fatty Acids
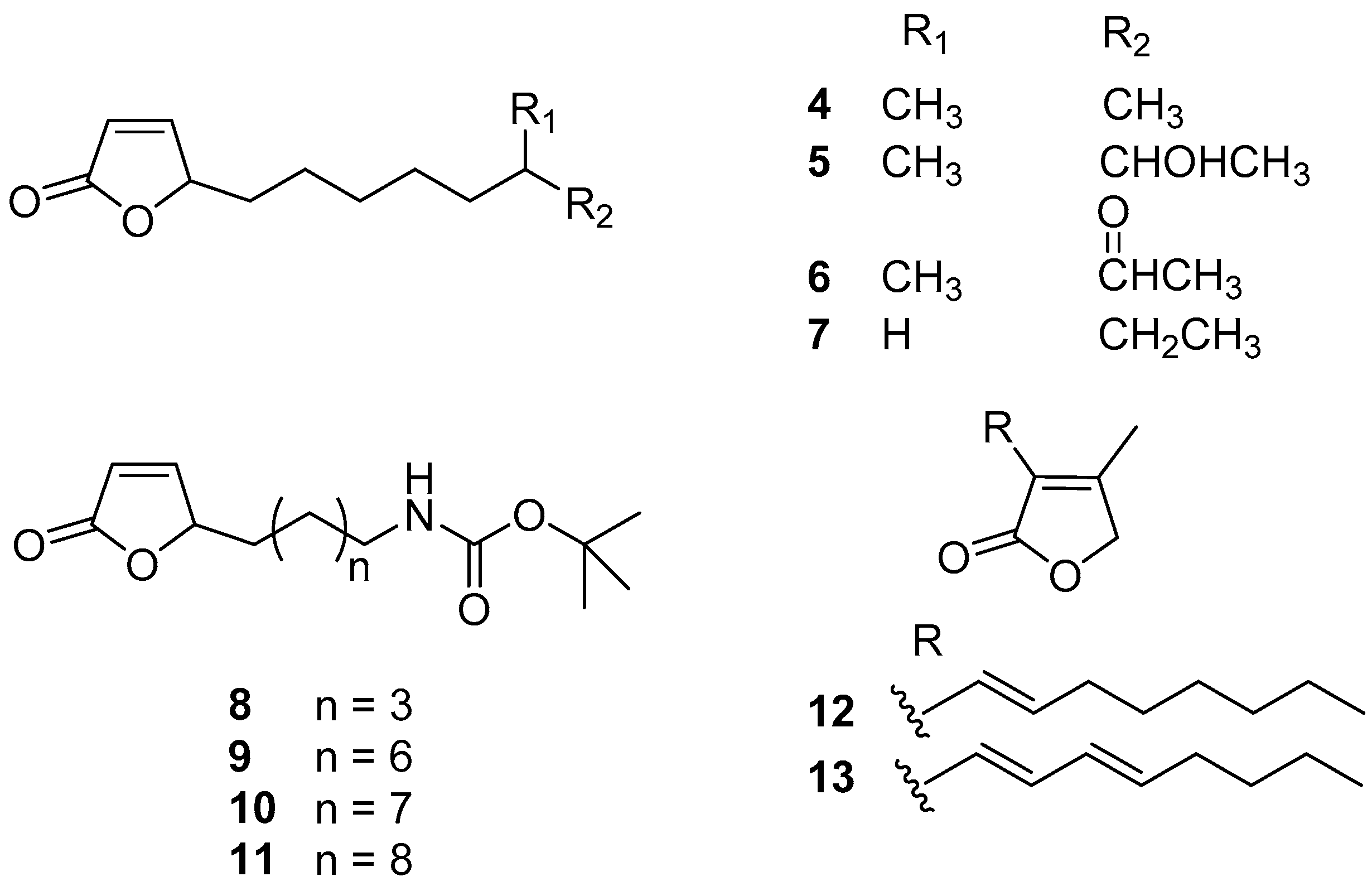
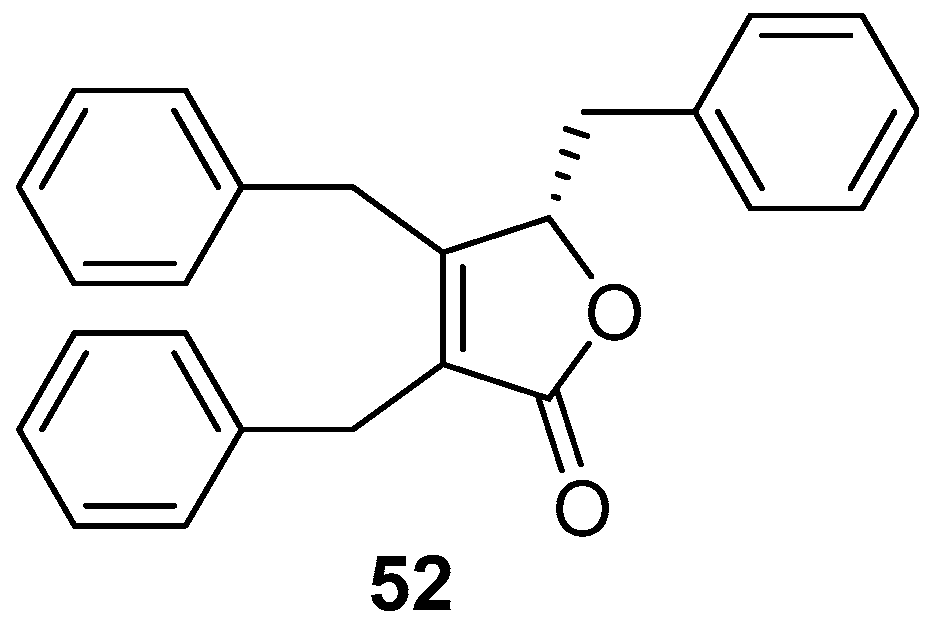
2.2. Butenolides
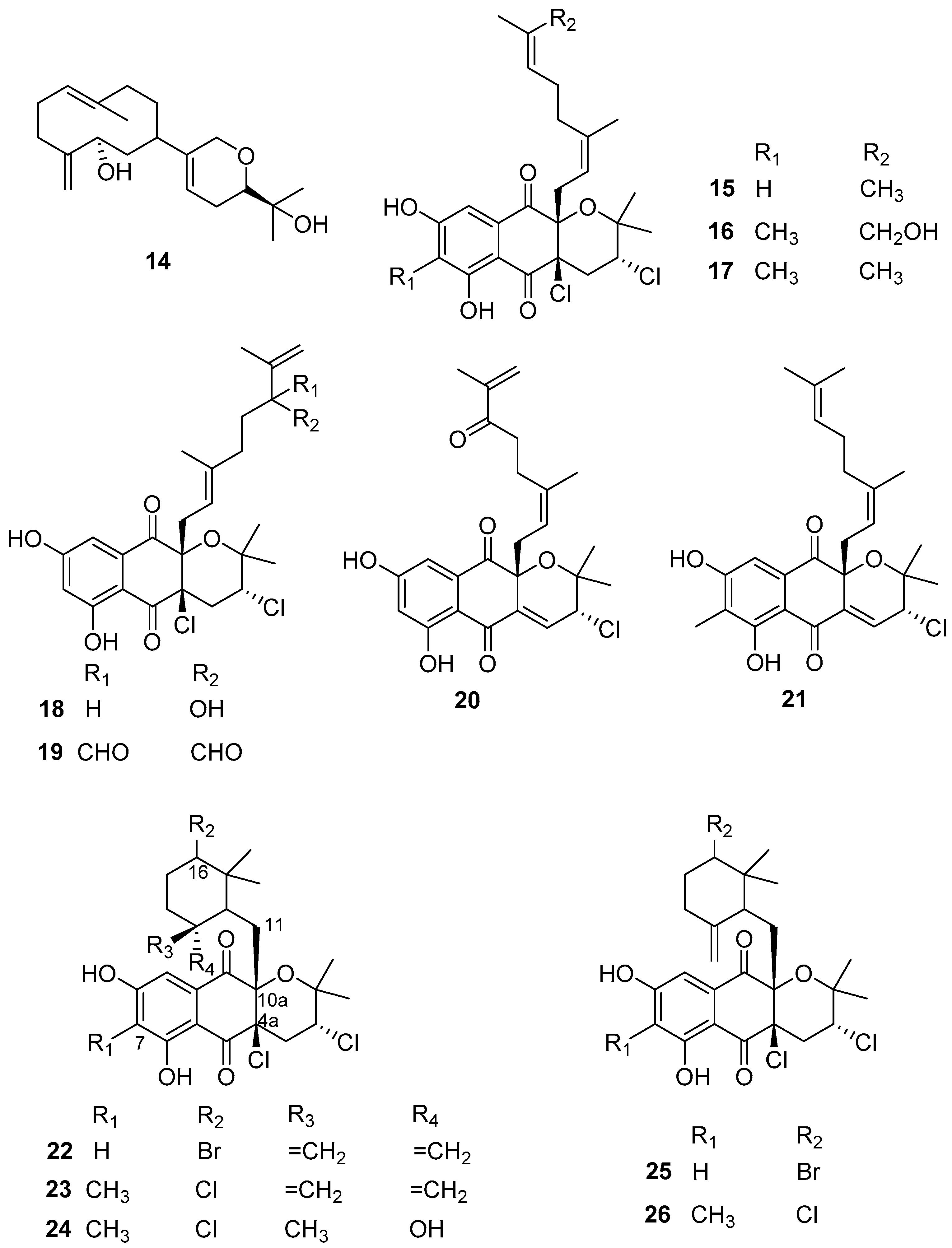
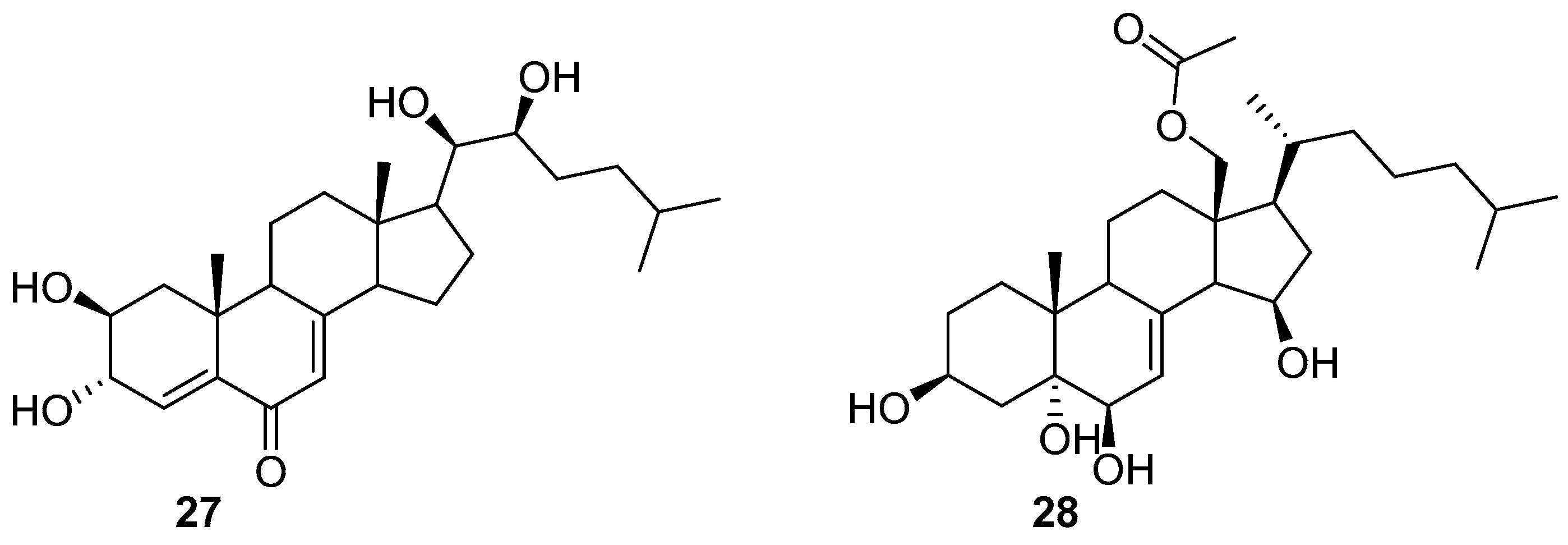
2.3. Terpenoids and Steroids
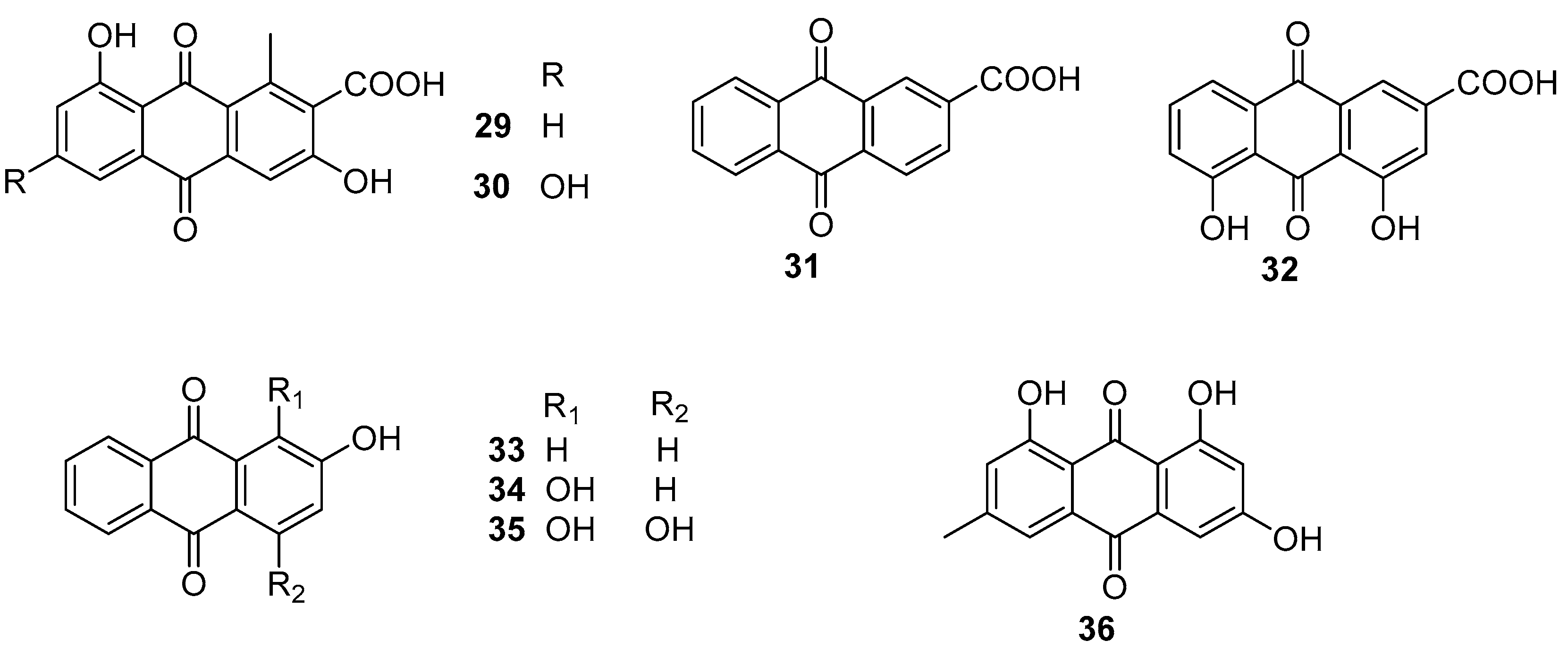
2.4. Anthraquinones
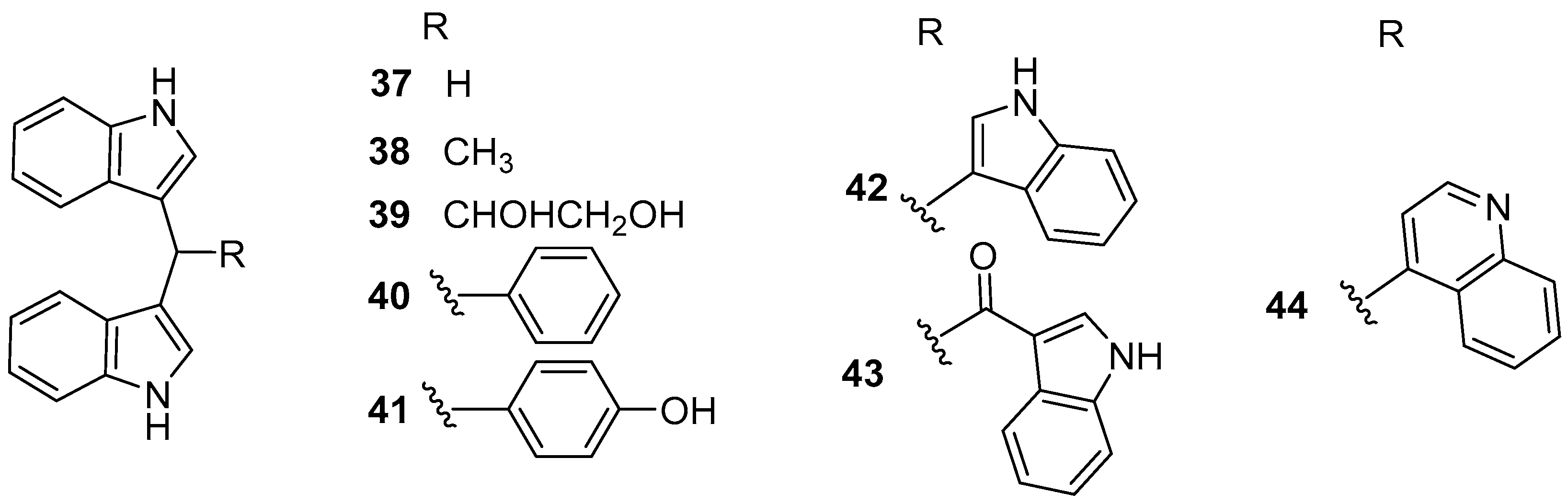
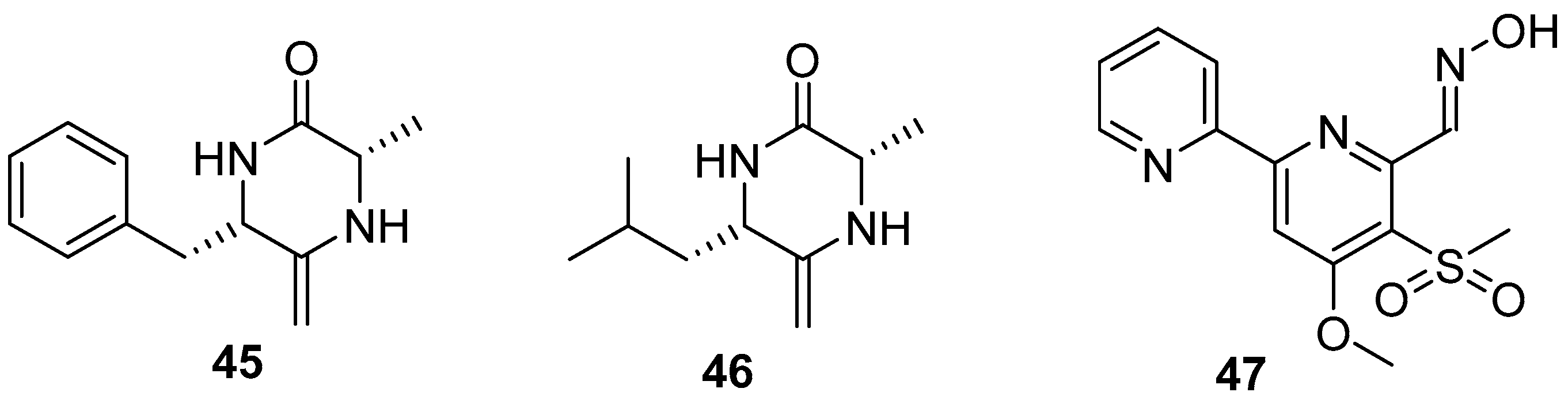
2.5. Alkaloids
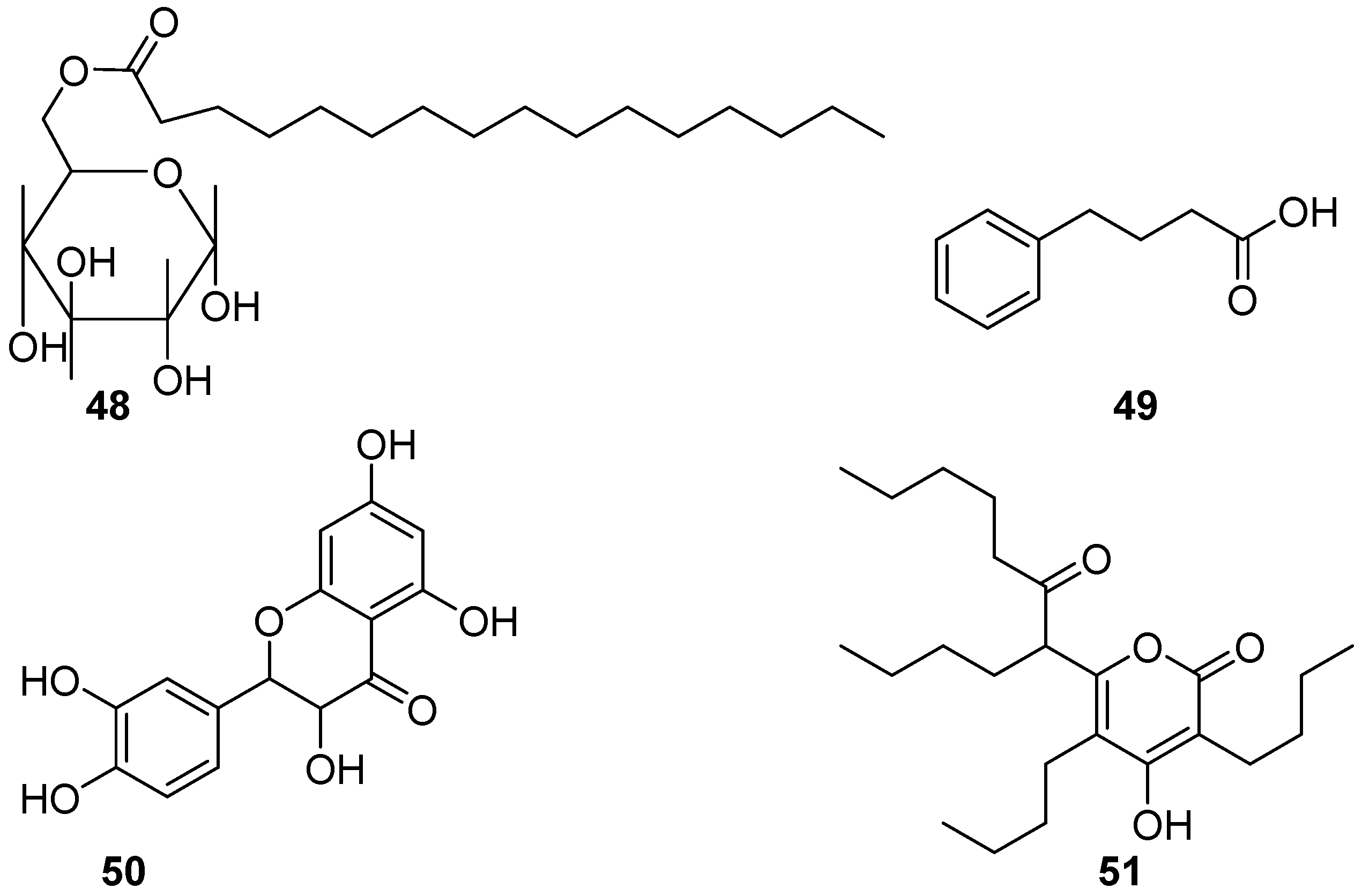
2.6. Glycolipids, Benzenoids, Flavonoids, Polyketides, and Enzymes
3. Antifouling Natural Products from Marine Cyanobacteria
3.1. Butenolides
3.2. Polyketide–Polypeptide Structural Compounds
4. Anti-Biofilm and Antifouling Natural Products from Marine Fungi
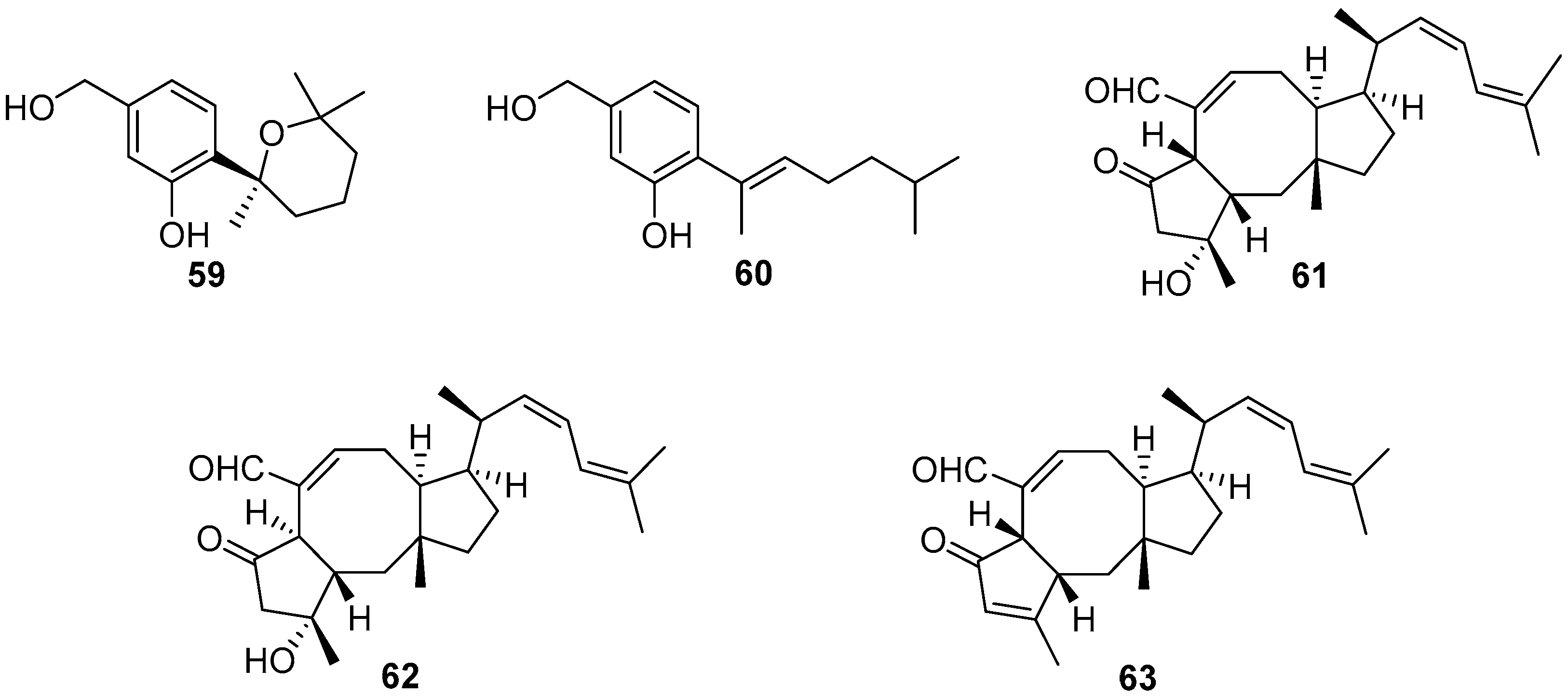
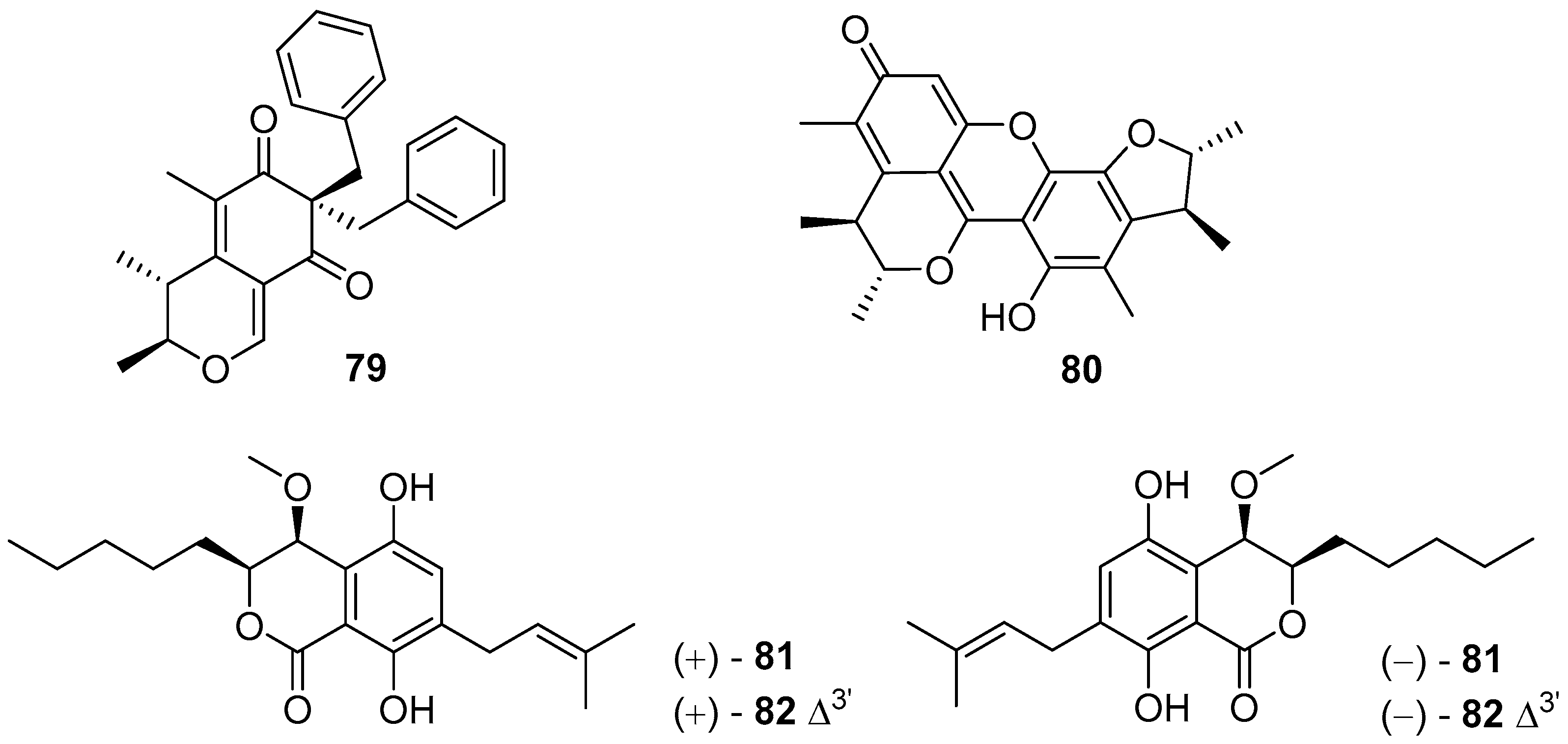
4.1. Terpenoids
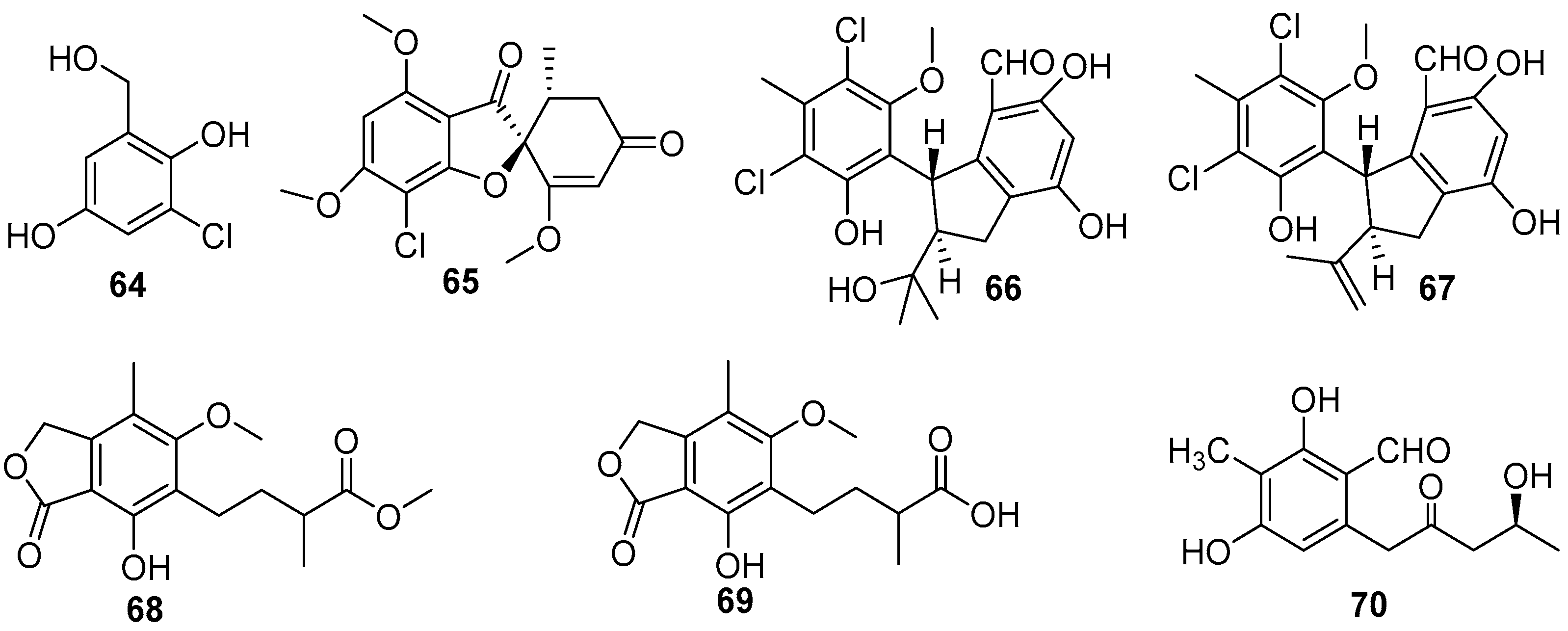
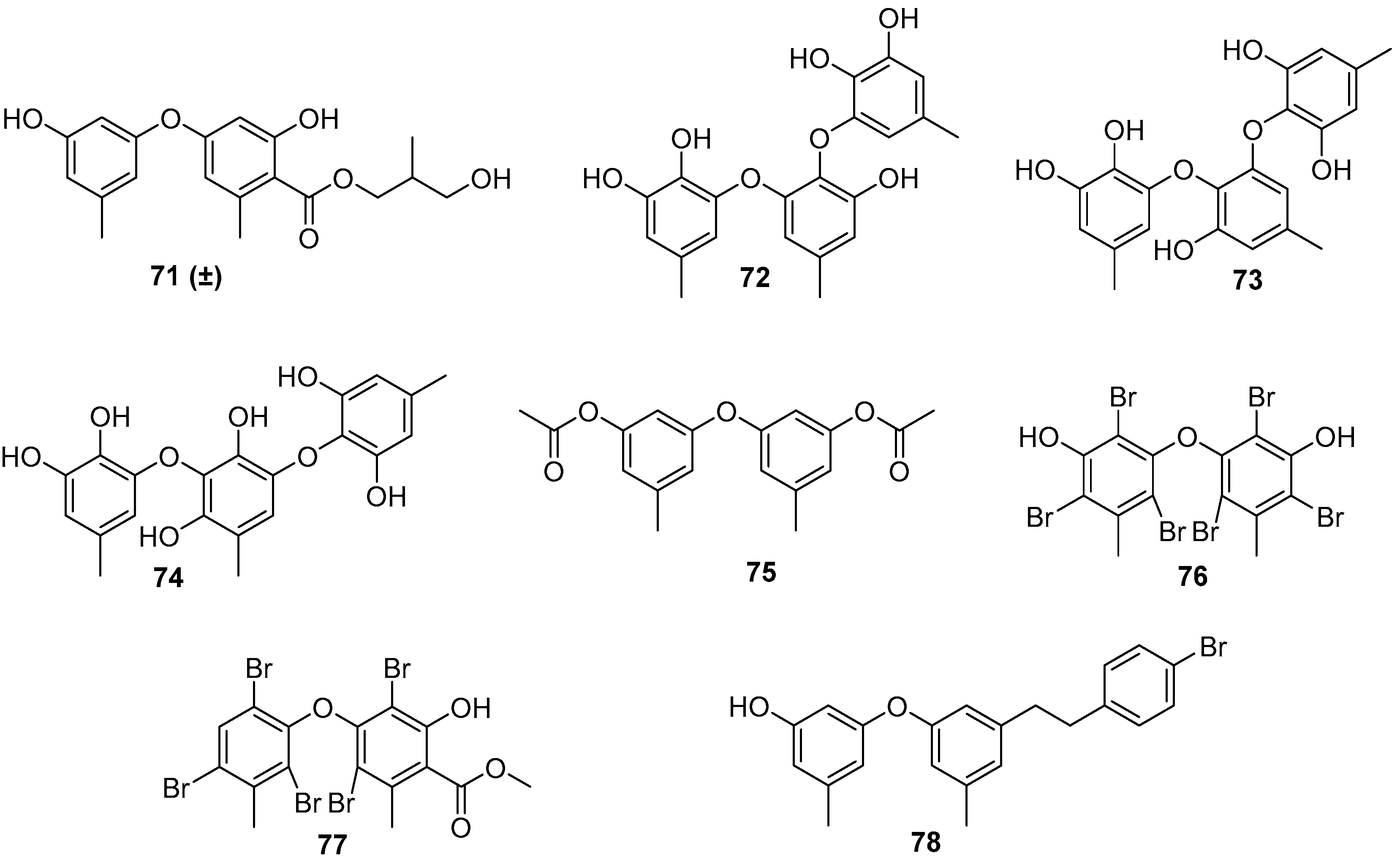
4.2. Phenols and Phenyl Ethers
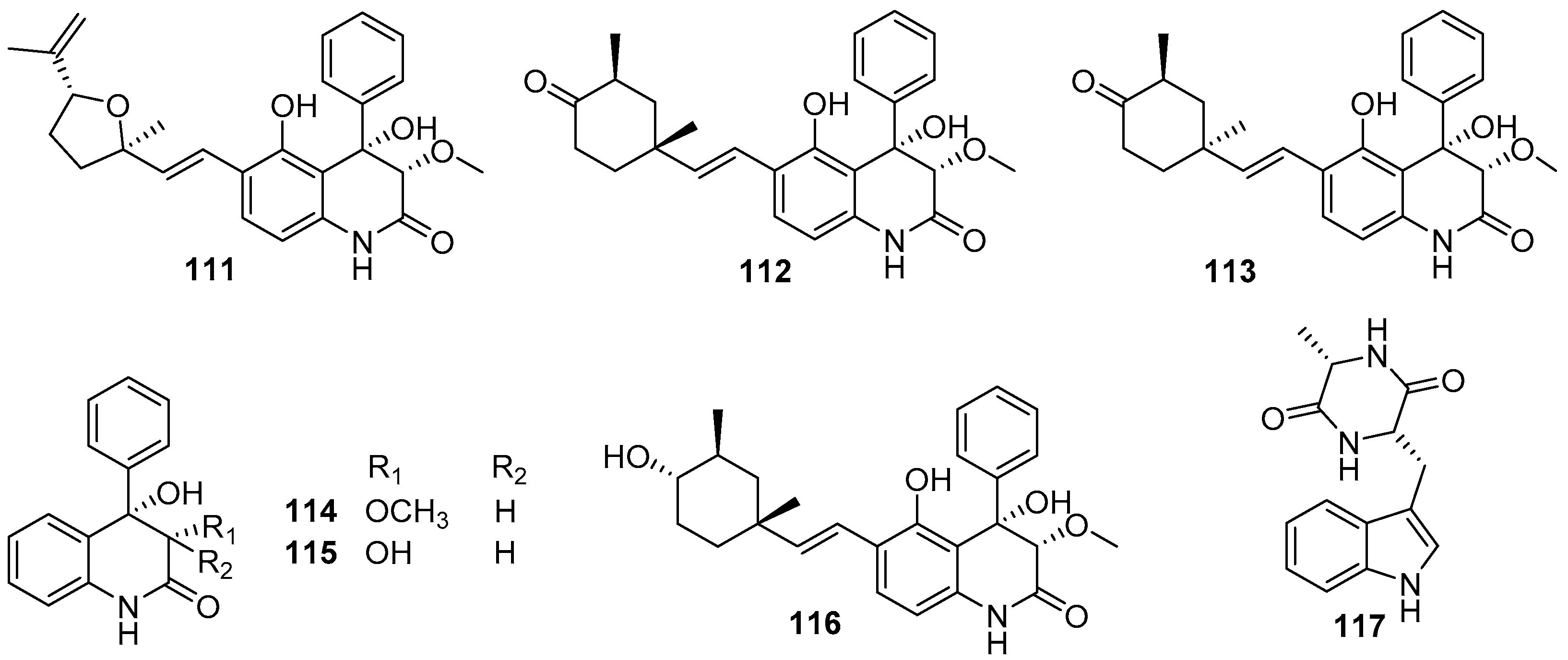
4.3. Polyketides
4.3.1. Benzylazaphilones and Dihydroisocoumarins
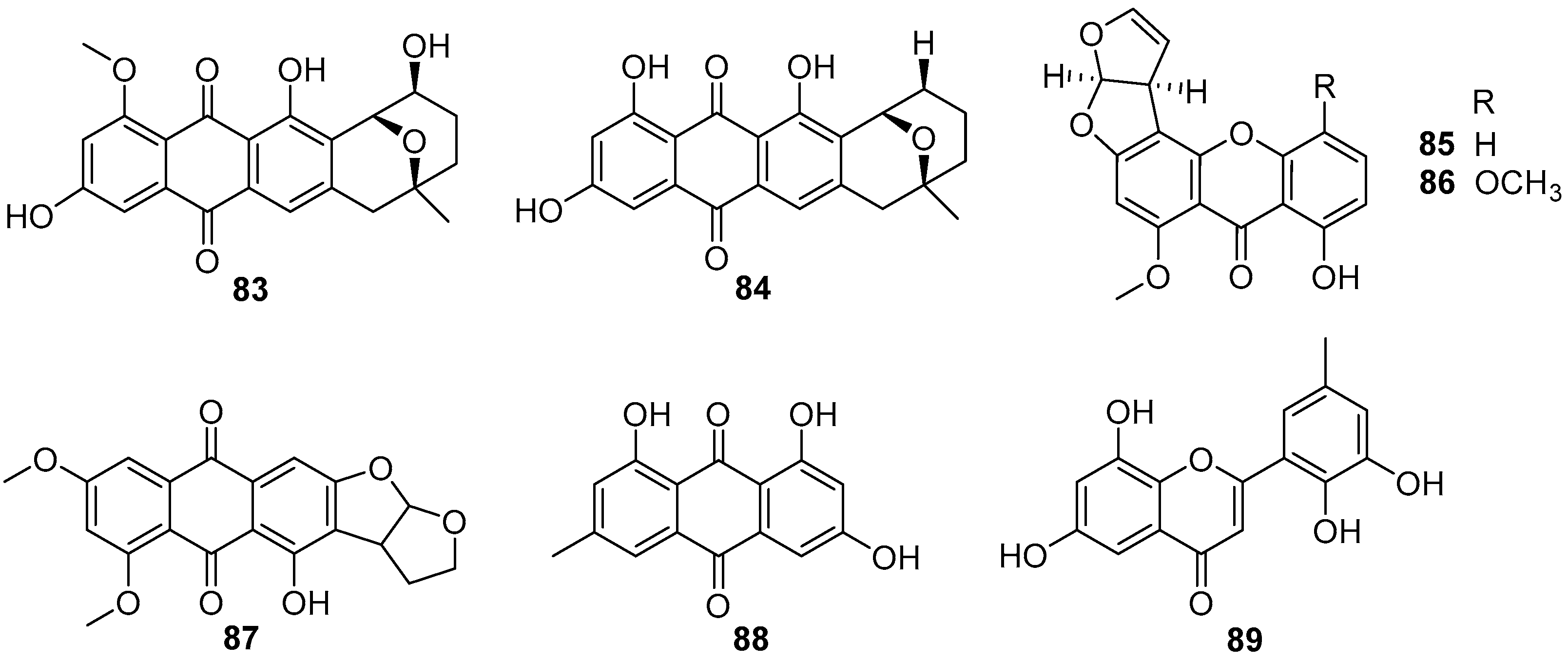
4.3.2. Anthraquinones and Xanthones
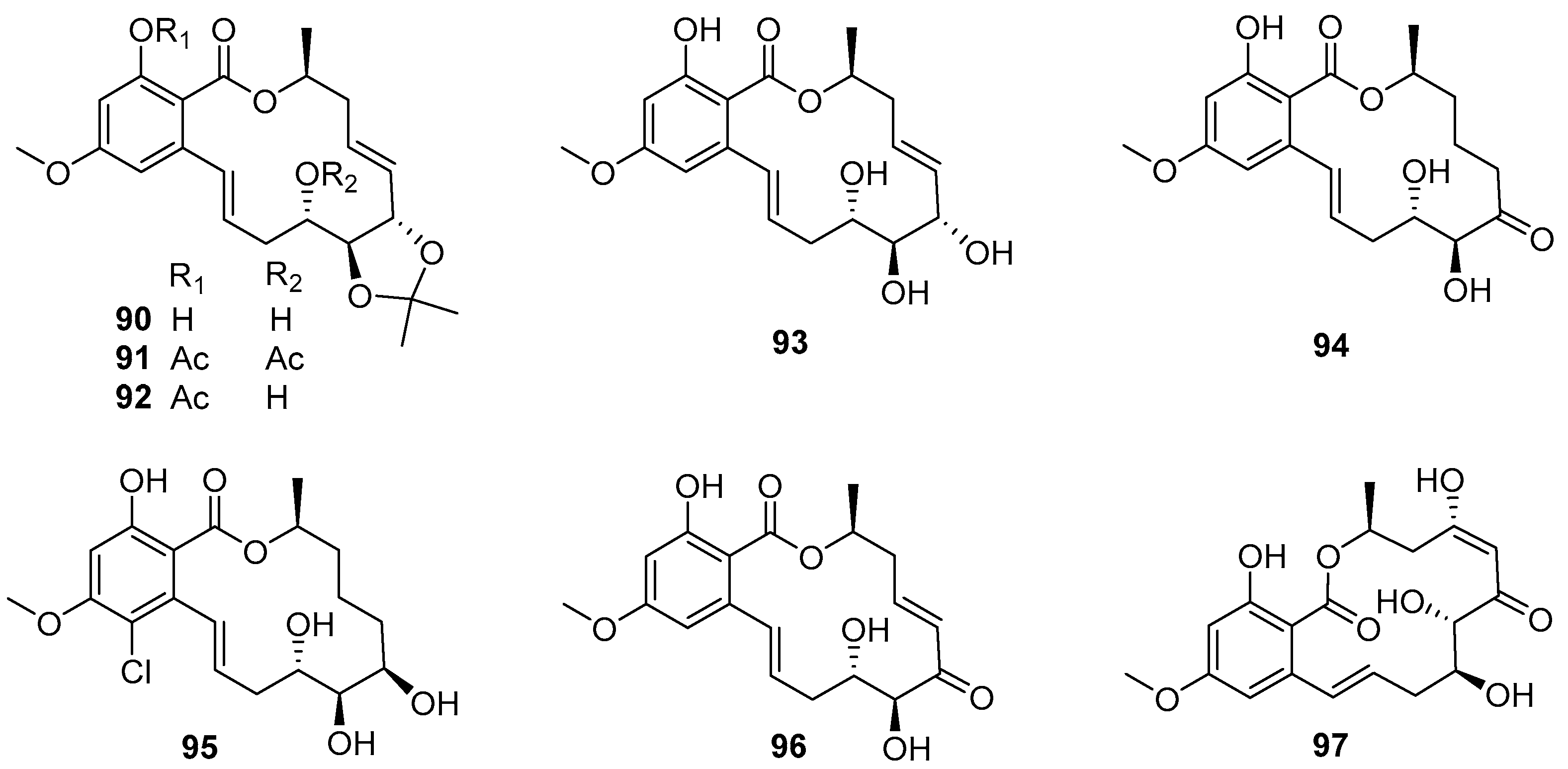
4.3.3. 14-Membered Resorcylic Acid Lactones
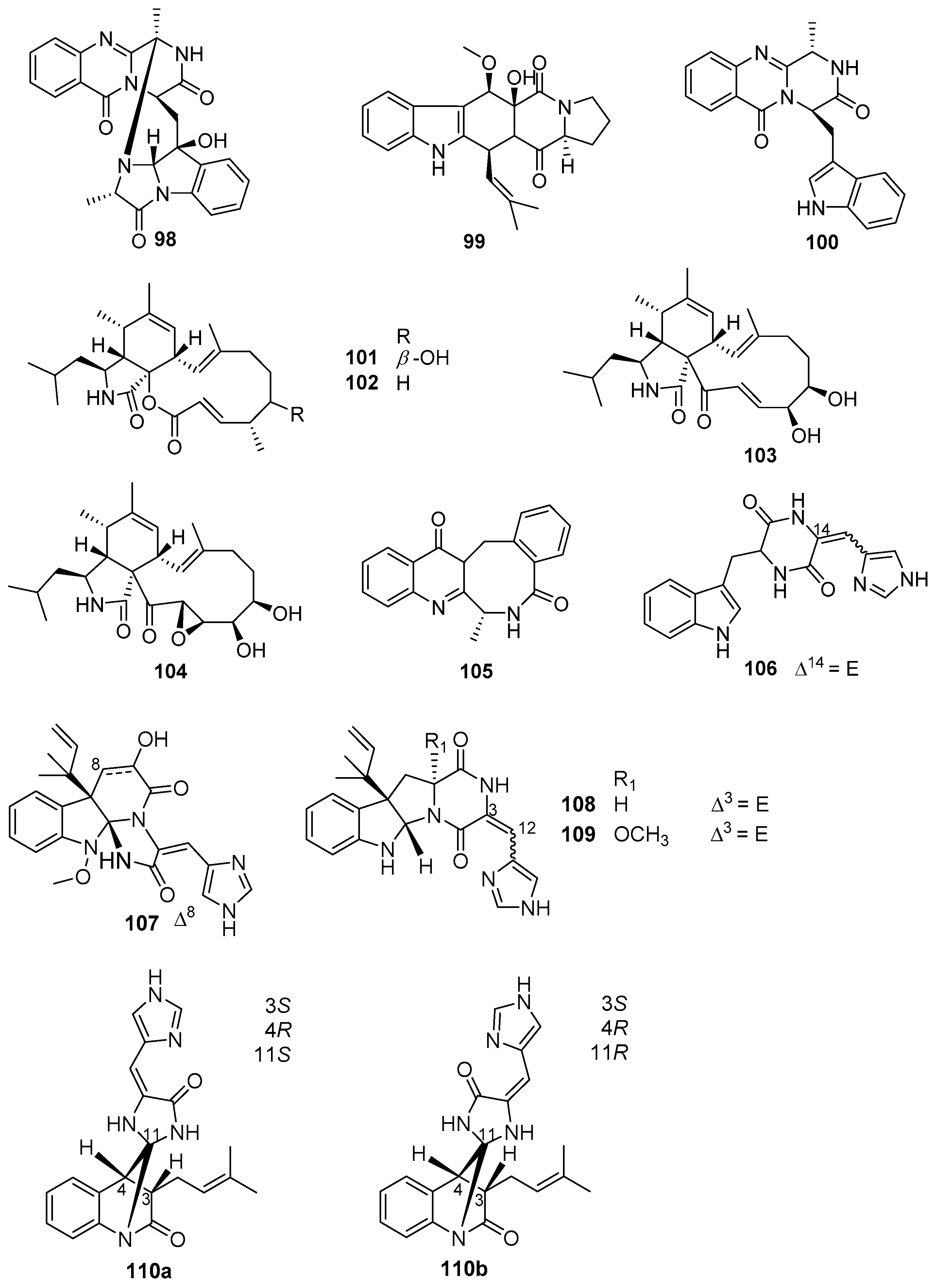
4.4. Alkaloids
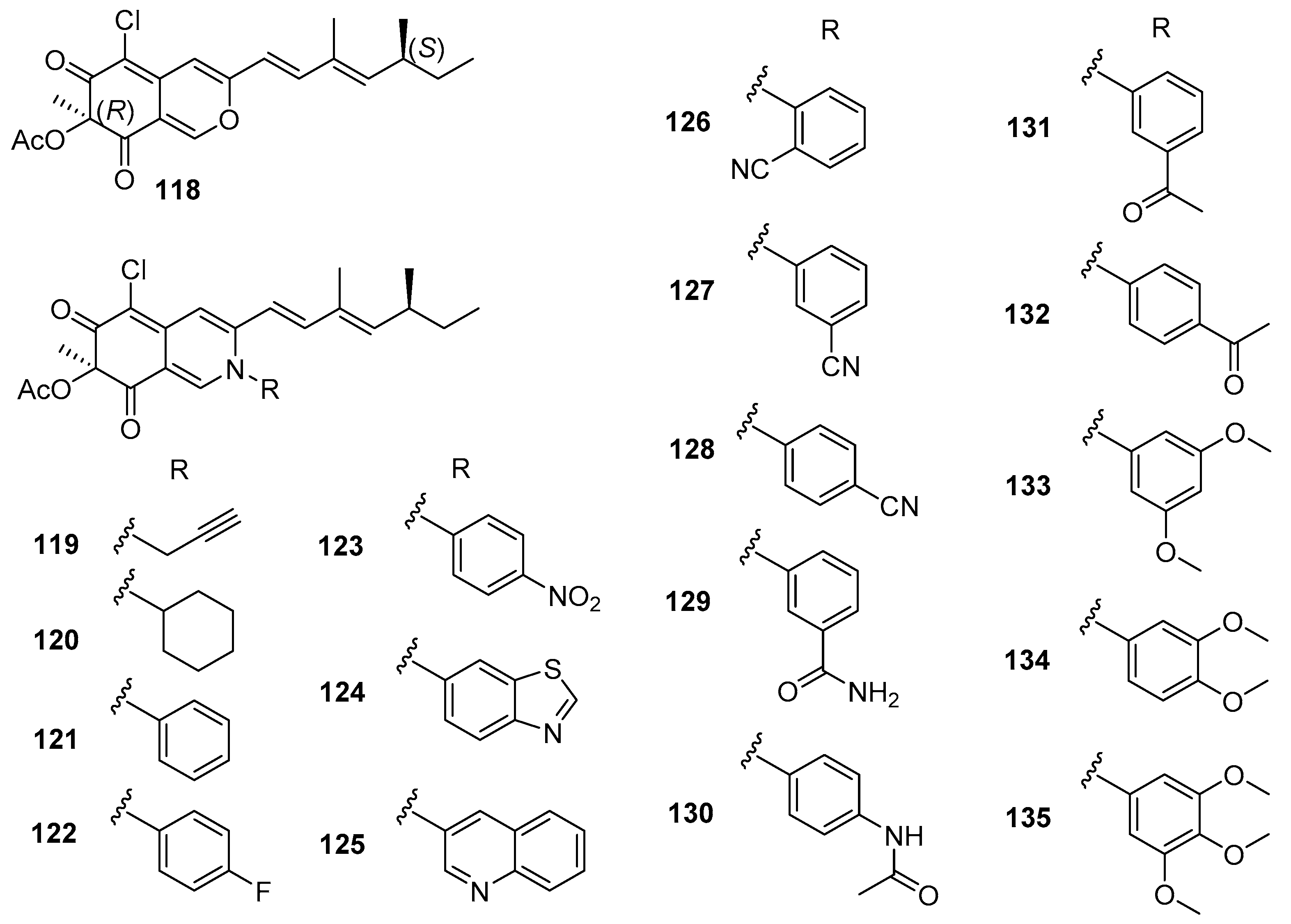
4.5. Amines
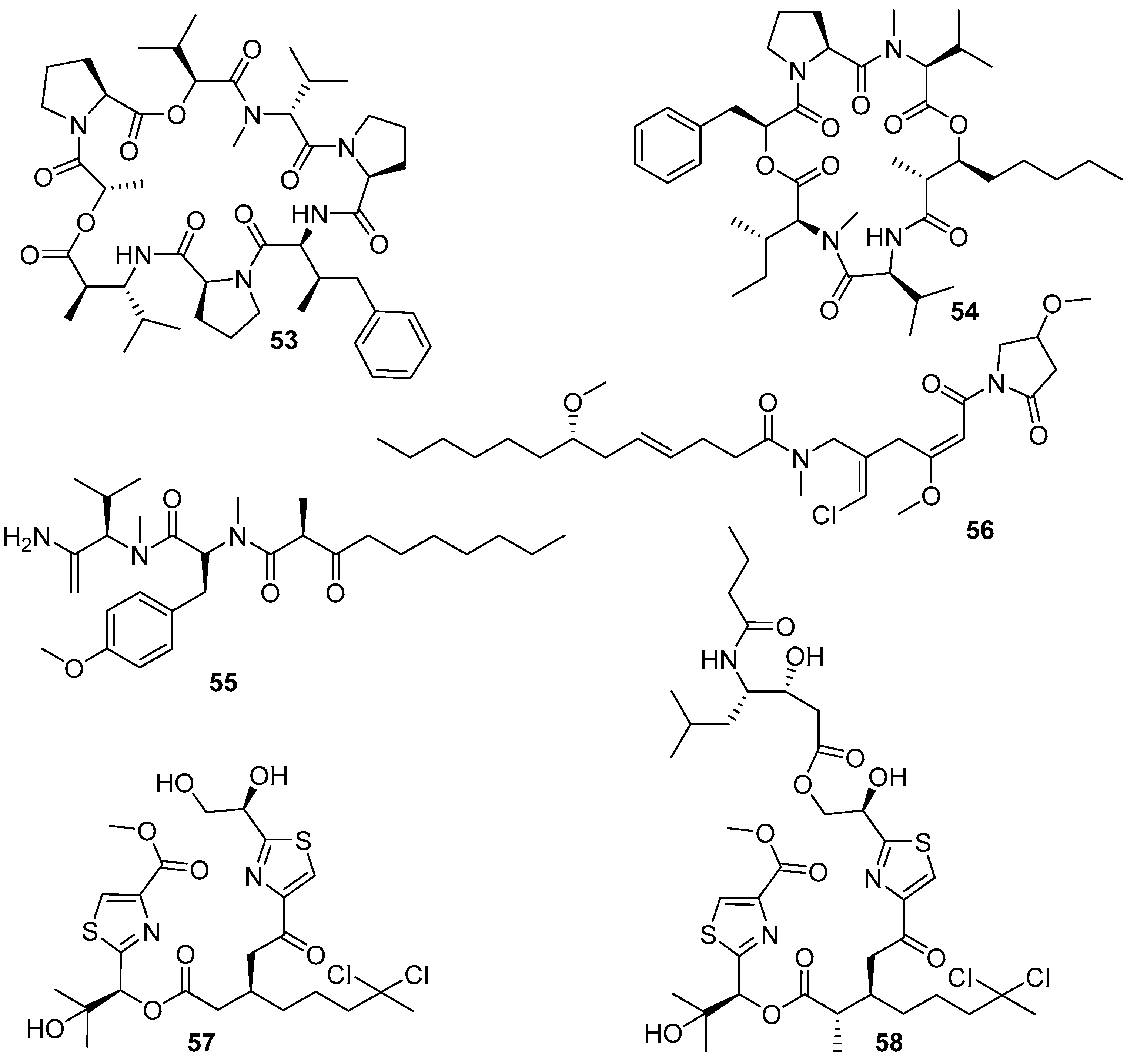
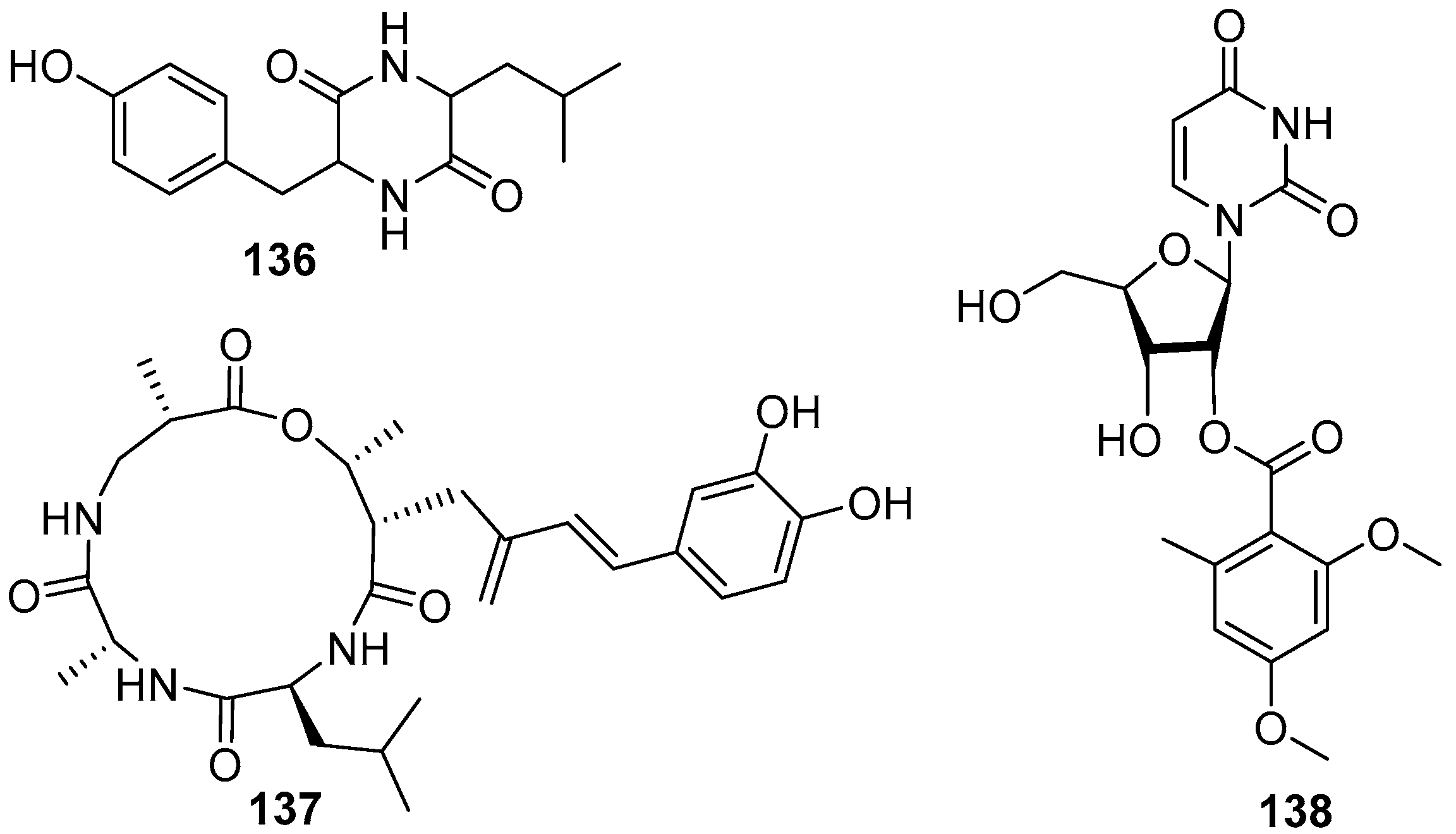
4.6. Peptides and Nucleosides
5. Conclusions and Looking Ahead
Author Contributions
Funding
Conflicts of Interest
References
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Stowe, S.D.; Richards, J.J.; Tucker, A.T.; Thompson, R.; Melander, C.; Cavanagh, J. Anti-biofilm compounds derived from marine sponges. Mar. Drugs 2011, 9, 2010–2035. [Google Scholar] [CrossRef] [Green Version]
- Yebra, D.M.; Kiil, S.; Dam-Johansen, K. Antifouling technology—past, present and future steps towards efficient and environmentally friendly antifouling coatings. Prog. Org. Coat. 2004, 50, 75–104. [Google Scholar] [CrossRef]
- Wahl, M. Marine epibiosis. I. Fouling and antifouling: Some basic aspects. Mar. Ecol. Prog. Ser. 1989, 58, 175–189. [Google Scholar] [CrossRef] [Green Version]
- Dobretsov, S.; Dahms, H.U.; Qian, P.-Y. Inhibition of biofouling by marine microorganisms and their metabolites. Biofouling 2006, 22, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Qian, P.Y.; Lau, S.; Dahms, H.U.; Dobretsov, S.; Harder, T. Marine biofilms as mediators of colonization by marine macroorganisms: Implications for antifouling and aquaculture. Mar. Biotechnol. 2007, 9, 399–410. [Google Scholar] [CrossRef] [PubMed]
- Dobretsov, S.; Abed, R.M.M.; Teplitski, M. Mini-review: Inhibition of biofouling by marine microorganisms. Biofouling 2013, 29, 423–441. [Google Scholar] [CrossRef]
- Satheesh, S.; Ba-akdah, M.A.; Al-Sofyani, A.A. Natural antifouling compound production by microbes associated with marine macroorganisms—A review. Electron. J. Biotechnol. 2016, 21, 26–35. [Google Scholar] [CrossRef] [Green Version]
- Dobretsov, S.; Teplitski, M.; Alagely, A.; Gunasekera, S.P.; Paul, V.J. Malyngolide from the cyanobacterium Lyngbya majuscula interferes with quorum sensing circuitry. Environ. Microbiol. Rep. 2010, 2, 739–744. [Google Scholar] [CrossRef]
- Davies, D. Understanding biofilm resistance to antibacterial agents. Nat. Rev. Drug Discov. 2003, 2, 114–122. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Issues in Healthcare Settings: CDC’s Seven Healthcare Safety Challenges; United States Department of Health and Human Services: Atlanta, GA, USA, 2001.
- Carroll, A.R.; Copp, B.R.; Davis, R.A.; Keyzers, R.A.; Prinsep, M.R. Marine natural products. Nat. Prod. Rep. 2019, 36, 122–173. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, L.; Hu, W.; Tian, Z.; Yuan, D.; Yi, G.; Zhou, Y.; Cheng, Q.; Zhu, J.; Li, M. Developing natural products as potential anti-biofilm agents. Chin. Med. 2019, 14, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Melander, R.J.; Basak, A.K.; Melander, C. Natural products as inspiration for the development of bacterial antibiofilm agents. Nat. Prod. Rep. 2020, 37, 1454–1477. [Google Scholar] [CrossRef]
- Mishra, R.; Panda, A.K.; De Mandal, S.; Shakeel, M.; Bisht, S.S.; Khan, J. Natural anti-biofilm agents: Strategies to control biofilm-forming pathogens. Front. Microbiol. 2020, 11, 2640. [Google Scholar] [CrossRef] [PubMed]
- Fusetani, N. Biofouling and antifouling. Nat. Prod. Rep. 2004, 21, 94–104. [Google Scholar] [CrossRef]
- Qian, P.-Y.; Xu, Y.; Fusetani, N. Natural products as antifouling compounds: Recent progress and future perspectives. Biofouling 2009, 26, 223–234. [Google Scholar] [CrossRef]
- Fusetani, N. Antifouling marine natural products. Nat. Prod. Rep. 2011, 28, 400–410. [Google Scholar] [CrossRef]
- Qian, P.-Y.; Li, Z.; Xu, Y.; Li, Y.; Fusetani, N. Mini-review: Marine natural products and their synthetic analogs as antifouling compounds: 2009–2014. Biofouling 2015, 31, 101–122. [Google Scholar] [CrossRef]
- Wang, K.-L.; Wu, Z.-H.; Wang, Y.; Wang, C.-Y.; Xu, Y. Mini-Review: Antifouling natural products from marine microorganisms and their synthetic analogs. Mar. Drugs 2017, 15, 266. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhattarai, H.D.; Ganti, V.S.; Paudel, B.; Lee, Y.K.; Lee, H.K.; Hong, Y.K.; Shin, H.W. Isolation of antifouling compounds from the marine bacterium, Shewanella oneidensis SCH0402. World J. Microbiol. Biotechnol. 2007, 23, 243–249. [Google Scholar] [CrossRef]
- Xu, Y.; Li, H.; Li, X.; Xiao, X.; Qian, P.-Y. Inhibitory effects of a branched-chain fatty acid on larval settlement of the polychaete Hydroides elegans. Mar. Biotechnol. 2009, 11, 495–504. [Google Scholar] [CrossRef]
- Dickschat, J.S.; Martens, T.; Brinkhoff, T.; Simon, M.; Schulz, S. Volatiles released by a Streptomyces species isolated from the North Sea. Chem. Biodivers. 2005, 2, 837–865. [Google Scholar] [CrossRef]
- Xu, Y.; He, H.; Schulz, S.; Liu, X.; Fusetani, N.; Xiong, H.; Xiao, X.; Qian, P.-Y. Potent antifouling compounds produced by marine Streptomyces. Bioresour. Technol. 2010, 101, 1331–1336. [Google Scholar] [CrossRef] [PubMed]
- Chiang, H.Y.; Cheng, J.; Liu, X.; Ma, C.; Qian, P.Y. Synthetic analogue of butenolide as an antifouling agent. Mar. Drugs 2021, 19, 481. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhang, F.; Xu, Y.; Matsumura, K.; Han, Z.; Liu, L.; Lin, W.; Jia, Y.; Qian, P.-Y. Structural optimization and evaluation of butenolides as potent antifouling agents: Modification of the side chain affects the biological activities of compounds. Biofouling 2012, 28, 857–864. [Google Scholar] [CrossRef]
- Hong, Y.K.; Cho, J.Y. Effect of seaweed epibiotic bacterium Streptomyces violaceoruber SCH-09 on marine fouling organisms. Fish. Sci. 2013, 79, 469–475. [Google Scholar] [CrossRef]
- Zhang, Y.; Xu, Y.; Arellano, S.M.; Xiao, K.; Qian, P.-Y. Comparative proteome and phosphoproteome analyses during cyprid development of the barnacle Balanus (=Amphibalanus) amphitrite. J. Proteome Res. 2010, 9, 3146–3157. [Google Scholar] [CrossRef]
- Qian, P.-Y.; Wong, Y.H.; Zhang, Y. Changes in the proteome and phosphoproteome expression in the bryozoan Bugula neritina larvae in response to the antifouling agent butenolide. Proteomics 2010, 10, 3435–3446. [Google Scholar] [CrossRef] [PubMed]
- Ding, W.; Ma, C.; Zhang, W.; Chiang, H.; Qian, P.-Y. Anti-biofilm effect of a butenolide polymer coating and metatranscriptomic analyses. Biofouling 2018, 34, 111–122. [Google Scholar] [CrossRef] [Green Version]
- Yin, Q.; Liang, J.; Zhang, W.; Zhang, L.; Hu, Z.L.; Zhang, Y.; Ying, X. Butenolide, a marine-derived broad-spectrum antibiofilm agent against both gram-positive and gram-negative pathogenic bacteria. Mar. Biotechnol. 2019, 21, 88–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, J.Y.; Kim, M.S. Induction of antifouling diterpene production by Streptomyces cinnabarinus PK209 in co-culture with marine-derived Alteromonas sp. KNS-16. Biosci. Biotechnol. Biochem. 2012, 76, 1849–1854. [Google Scholar] [CrossRef] [Green Version]
- Bauermeister, A.; Pereira, F.; Grilo, I.R.; Godinho, C.C.; Paulino, M.; Almeida, V.; Gobbo-Neto, L.; Prieto-Davo, A.; Sobral, R.G.; Lopes, N.P.; et al. Intra-clade metabolomic profiling of MAR4 Streptomyces from the Macaronesia Atlantic region reveals a source of anti-biofilm metabolites. Environ. Microbiol. 2019, 21, 1099–1112. [Google Scholar] [CrossRef]
- Pereira, F.; Almeida, J.R.; Paulino, M.; Grilo, I.R.; Macedo, H.; Cunha, I.; Sobral, R.G.; Vasconcelos, V.; Gaudêncio, S.P. Antifouling napyradiomycins from marine-derived actinomycetes Streptomyces aculeolatus. Mar. Drugs 2020, 18, 63. [Google Scholar] [CrossRef] [Green Version]
- Cho, J.Y. Antifouling steroids isolated from red alga epiphyte filamentous bacterium Leucothrix mucor. Fish. Sci. 2012, 78, 683–689. [Google Scholar] [CrossRef]
- Song, Z.-M.; Zhang, J.-L.; Zhou, K.; Yue, L.-M.; Zhang, Y.; Wang, C.-Y.; Wang, K.-L.; Xu, Y. Anthraquinones as potential antibiofilm agents against Methicillin-resistant Staphylococcus aureus. Front. Microbiol. 2021, 12, 709826. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.-L.; Xu, Y.; Lu, L.; Li, Y.; Han, Z.; Zhang, J.; Shao, C.-L.; Wang, C.-Y.; Qian, P.-Y. Low-toxicity diindol-3-ylmethanes as potent antifouling compounds. Mar. Biotechnol. 2015, 17, 624–632. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.Y.; Kang, J.Y.; Hong, Y.K.; Baek, H.H.; Shin, H.W.; Kim, M.S. Isolation and structural determination of the antifouling diketopiperazines from marine-derived Streptomyces praecox 291-11. Biosci. Biotechnol. Biochem. 2012, 76, 1116–1121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.-L.; Liang, X.-Y.; Zhang, S.-L.; Song, Z.-M.; Wang, C.-Y.; Xu, Y. Maipomycin A, a novel natural compound with promising anti-biofilm activity against Gram-negative pathogenic bacteria. Front. Microbiol. 2021, 11, 3480. [Google Scholar] [CrossRef] [PubMed]
- Dusane, D.H.; Pawar, V.S.; Nancharaiah, Y.V.; Venugopalan, V.P.; Kumar, A.R.; Zinjarde, S.S. Anti-biofilm potential of a glycolipid surfactant produced by a tropical marine strain of Serratia marcescens. Biofouling 2011, 27, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Nithya, C.; Devi, M.G.; Pandian, S.K. A novel compound from the marine bacterium Bacillus pumilus S6-15 inhibits biofilm formation in gram-positive and gram-negative species. Biofouling 2011, 27, 519–528. [Google Scholar] [CrossRef] [PubMed]
- Gopikrishnan, V.; Radhakrishnan, M.; Shanmugasundaram, T.; Gopikrishnan, V.; Radhakrishnan, M.; Shanmugasundaram, T.; Ramakodi, M.P.; Balagurunathan, R. Isolation, characterization and identification of antibiofouling metabolite from mangrove derived Streptomyces sampsonii PM33. Sci. Rep. 2019, 9, 12975. [Google Scholar] [CrossRef]
- Long, L.; Wang, R.; Chiang, H.Y.; Ding, W.; Li, Y.X.; Chen, F.; Qian, P.Y. Discovery of antibiofilm activity of elasnin against marine biofilms and its application in the marine antifouling coatings. Mar. Drugs 2021, 19, 19. [Google Scholar] [CrossRef] [PubMed]
- Dobretsov, S.; Xiong, H.; Xu, Y.; Levin, L.A.; Qian, P.-Y. Novel antifoulants: Inhibition of larval attachment by proteases. Mar. Biotechnol. 2007, 9, 388–397. [Google Scholar] [CrossRef] [PubMed]
- Brown, G.D.; Wong, H.F.; Hutchinson, N.; Lee, S.C.; Chan, B.K.K.; Williams, G.A. Chemistry and biology of maculalactone A from the marine cyanobacterium Kyrtuthrix maculans. Phytochem. Rev. 2004, 3, 381–400. [Google Scholar] [CrossRef]
- Bader, S.L.; Luescher, M.U.; Gademann, K. Synthesis of maculalactone A and derivatives for environmental fate tracking studies. Org. Biomol. Chem. 2015, 13, 199–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, L.T.; Goh, B.P.L.; Tripathi, A.; Lim, M.G.; Dickinson, G.H.; Lee, S.S.C.; Teo, S.L.M. Natural antifoulants from the marine cyanobacterium Lyngbya majuscula. Biofouling 2010, 26, 685–695. [Google Scholar] [CrossRef]
- Petitbois, J.G.; Casalme, L.O.; Lopez, J.A.V.; Alarif, W.M.; Abdel-Lateff, A.; Al-Lihaibi, S.S.; Yoshimura, E.; Nogata, Y.; Umezawa, T.; Matsuda, F.; et al. Serinolamides and lyngbyabellins from an Okeania sp. cyanobacterium collected from the Red Sea. J. Nat. Prod. 2017, 80, 2708–2715. [Google Scholar] [CrossRef] [PubMed]
- Antunes, J.; Pereira, S.; Ribeiro, T.; Plowman, J.E.; Thomas, A.; Clerens, S.; Campos, A.; Vasconcelos, V.; Almeida, J.R. A multi-bioassay integrated approach to assess the antifouling potential of the cyanobacterial metabolites portoamides. Mar. Drugs 2019, 17, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Casalme, L.O.; Yamauchi, A.; Sato, A.; Petitbois, J.G.; Nogata, Y.; Yoshimura, E.; Okino, T.; Umezawa, T.; Matsuda, F. Total synthesis and biological activity of dolastatin 16. Org. Biomol. Chem. 2017, 15, 1140–1150. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Xu, Y.; Shao, C.-L.; Yang, R.-Y.; Zheng, C.-J.; Chen, Y.-Y.; Fu, X.-M.; Qian, P.-Y.; She, Z.-G.; Voogd, N.J.D.; et al. Antibacterial bisabolane-type sesquiterpenoids from the sponge-derived fungus Aspergillus sp. Mar. Drugs 2012, 10, 234–241. [Google Scholar] [CrossRef] [Green Version]
- Arai, M.; Niikawa, H.; Kobayashi, M. Marine-derived fungal sesterterpenes, ophiobolins, inhibit biofilm formation of Mycobacterium species. J. Nat. Med. 2013, 67, 271–275. [Google Scholar] [CrossRef] [PubMed]
- Kwong, T.F.N.; Miao, L.; Li, X.; Qian, P.-Y. Novel antifouling and antimicrobial compound from a marine-derived fungus Ampelomyces sp. Mar. Biotechnol. 2006, 8, 634–640. [Google Scholar] [CrossRef]
- Xing, Q.; Gan, L.-S.; Mou, X.-F.; Wang, W.; Wang, C.-Y.; Wei, M.-Y.; Shao, C.-L. Isolation, resolution and biological evaluation of pestalachlorides E and F containing both point and axial chirality. RSC Adv. 2016, 6, 22653–22658. [Google Scholar] [CrossRef]
- Xu, X.; Zhang, X.; Nong, X.; Wang, J.; Qi, S.-H. Brevianamides and mycophenolic acid derivatives from the deep-sea-derived fungus penicillium brevicompactum DFFSCS025. Mar. Drugs 2017, 15, 43. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, Z.; Nong, X.; Ren, Z.; Zhe, R.; Wang, J.; Zhang, X.; Qi, S.-H. Anti-HSV-1, antioxidant and antifouling phenolic compounds from the deep-sea-derived fungus Aspergillus versicolor SCSIO 41502. Bioorg. Med. Chem. Lett. 2017, 27, 787–791. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-Y.; Wang, K.-L.; Qian, P.-Y.; Xu, Y.; Chen, M.; Zheng, J.-J.; Liu, M.; Shao, C.-L.; Wang, C.-Y. Antifouling phenyl ethers and other compounds from the invertebrates and their symbiotic fungi collected from the South China Sea. AMB Express 2016, 6, 102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shao, C.-L.; Wang, C.-Y.; Wei, M.-Y.; Gu, Y.-C.; She, Z.-G.; Qian, P.-Y.; Lin, Y.-C. Aspergilones A and B, two benzylazaphilones with an unprecedented carbon skeleton from the gorgonian-derived fungus Aspergillus sp. Bioorg. Med. Chem. Lett. 2011, 21, 690–693. [Google Scholar] [CrossRef] [PubMed]
- Nong, X.-H.; Zheng, Z.-H.; Zhang, X.-Y.; Lu, X.-H.; Qi, S.-H. Polyketides from a marine-derived fungus Xylariaceae sp. Mar. Drugs 2013, 11, 1718–1727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, M.; Shao, C.-L.; Wang, K.-L.; Xu, Y.; She, Z.-G.; Wang, C.-Y. Dihydroisocoumarin derivatives with antifouling activities from a gorgonian-derived Eurotium sp. fungus. Tetrahedron 2014, 70, 9132–9138. [Google Scholar] [CrossRef]
- Li, Y.X.; Wu, H.X.; Xu, Y.; Shao, C.-L.; Wang, C.-Y.; Qian, P.-Y. Antifouling activity of secondary metabolites isolated from Chinese marine organisms. Mar. Biotechnol. 2013, 15, 552–558. [Google Scholar] [CrossRef]
- Shao, C.-L.; She, Z.-G.; Guo, Z.; Peng, H.; Cai, X.; Zhou, S.; Gu, Y.-C.; Lin, Y.-C. 1H and 13C NMR assignments for two anthraquinones and two xanthones from the mangrove fungus (ZSUH-36). Magn. Reson. Chem. 2007, 45, 434–438. [Google Scholar] [CrossRef]
- Bao, J.; Sun, Y.-L.; Zhang, X.-Y.; Han, Z.; Gao, H.-C.; He, F.; Qian, P.-Y.; Qi, S.-H. Antifouling and antibacterial polyketides from marine gorgonian coral-associated fungus Penicillium sp. SCSGAF 0023. J. Antibiot. 2013, 66, 219–223. [Google Scholar] [CrossRef] [Green Version]
- Shao, C.-L.; Wu, H.-X.; Wang, C.-Y.; Liu, Q.-A.; Xu, Y.; Wei, M.-Y.; Qian, P.-Y.; Gu, Y.-C.; Zheng, C.-J.; She, Z.-G.; et al. Potent antifouling resorcylic acid lactones from the gorgonian-derived fungus Cochliobolus lunatus. J. Nat. Prod. 2011, 74, 629–633. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.-L.; Zhang, G.; Sun, J.; Xu, Y.; Han, Z.; Liu, L.-L.; Shao, C.-L.; Liu, Q.-A.; Wang, C.-Y.; Qian, P.-Y. Cochliomycin A inhibits the larval settlement of Amphibalanus amphitrite by activating the NO/cGMP pathway. Biofouling 2016, 32, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.A.; Shao, C.-L.; Gu, Y.-C.; Blum, M.; Gan, L.S.; Wang, K.-L.; Chen, M.; Wang, C.-Y. Antifouling and fungicidal resorcylic acid lactones from the Sea Anemone-derived fungus Cochliobolus lunatus. J. Agric. Food Chem. 2014, 62, 3183–3191. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.-F.; Xue, X.-J.; Qi, Y.-X.; Wu, N.-N.; Shao, C.-L. Cochliomycin G, a 14-membered resorcylic acid lactone from a marine-derived fungus Cochliobolus lunatus. Nat. Prod. Res. 2019, 35, 490–493. [Google Scholar] [CrossRef] [PubMed]
- He, F.; Han, Z.; Peng, J.; Qian, P.-Y.; Qi, S.-H. Antifouling indole alkaloids from two marine derived Fungi. Nat. Prod. Commun. 2013, 8, 329–332. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, C.-J.; Shao, C.-L.; Wu, L.Y.; Chen, M.; Wang, K.-L.; Zhao, D.-L.; Sun, X.-P.; Chen, G.-Y.; Wang, C.-Y. Bioactive phenylalanine derivatives and cytochalasins from the soft coral-derived fungus, Aspergillus elegans. Mar. Drugs 2013, 11, 2054–2068. [Google Scholar] [CrossRef] [Green Version]
- Fredimoses, M.; Zhou, X.; Ai, W.; Tian, X.; Yang, B.; Lin, X.; Xian, J.Y.; Liu, Y. Westerdijkin A, a new hydroxyphenylacetic acid derivative from deep sea fungus Aspergillus westerdijkiae SCSIO 05233. Nat. Prod. Res. 2015, 29, 158–162. [Google Scholar] [CrossRef]
- He, F.; Liu, Z.; Yang, J.; Fu, P.; Peng, J.; Zhu, W.-M.; Qi, S.-H. A novel antifouling alkaloid from halotolerant fungus Penicillium sp. OUCMDZ-776. Tetrahedron Lett. 2012, 53, 2280–2283. [Google Scholar] [CrossRef]
- Shao, C.-L.; Xu, R.-F.; Wang, C.-Y.; Qian, P.-Y.; Wang, K.-L.; Wei, M.-Y. Potent antifouling marine dihydroquinolin-2(1H)-one-containing alkaloids from the gorgonian coral-derived fungus Scopulariopsis sp. Mar. Biotechnol. 2015, 17, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Mou, X.-F.; Liu, X.; Xu, R.-F.; Wei, M.-Y.; Fang, Y.-W.; Shao, C.-L. Scopuquinolone B, a new monoterpenoid dihydroquinolin-2(1H)-one isolated from the coral-derived scopulariopsis sp. fungus. Nat. Prod. Res. 2018, 32, 773–776. [Google Scholar] [CrossRef] [PubMed]
- Bovio, E.; Fauchon, M.; Toueix, Y.; Mehiri, M.; Varese, G.C.; Hellio, C. The sponge-associated fungus Eurotium chevalieri MUT 2316 and its bioactive molecules: Potential applications in the field of antifouling. Mar. Biotechnol. 2019, 21, 743–752. [Google Scholar] [CrossRef]
- Wei, M.-Y.; Wang, C.-F.; Wang, K.-L.; Qian, P.-Y.; Wang, C.-Y.; Shao, C.-L. Preparation, structure, and potent antifouling activity of sclerotioramine derivatives. Mar. Biotechnol. 2017, 19, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Scopel, M.; Abraham, W.R.; Henriques, A.T.; Macedo, A.J. Dipeptide cis-cyclo(Leucyl-Tyrosyl) produced by sponge associated Penicillium sp. F37 inhibits biofilm formation of the pathogenic Staphylococcus epidermidis. Bioorg. Med. Chem. Lett. 2013, 23, 624–626. [Google Scholar] [CrossRef] [PubMed]
- Bao, J.; Zhang, X.-Y.; Xu, X.-Y.; He, F.; Nong, X.-H.; Qi, S.-H. New cyclic tetrapeptides and asteltoxins from gorgonian-derived fungus Aspergillus sp. SCSGAF 0076. Tetrahedron 2013, 69, 2113–2117. [Google Scholar] [CrossRef]
- Chen, M.; Fu, X.M.; Kong, C.-J.; Wang, C.-Y. Nucleoside derivatives from the marine-derived fungus Aspergillus versicolor. Nat. Prod. Res. 2014, 28, 895–900. [Google Scholar] [CrossRef]
- Zhang, H.; Wong, Y.H.; Wang, H.; Chen, Z.; Arellano, S.M.; Ravasi, T.; Qian, P.Y. Quantitative proteomics identify molecular targets that are crucial in larval settlement and metamorphosis of Bugula neritina. J. Proteome Res. 2011, 10, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; He, L.S.; Zhang, G.; Xu, Y.; Lee, O.O.; Matsumura, K.; Qian, P.Y. The regulatory role of the NO/cGMP signal transduction cascade during larval attachment and metamorphosis of the barnacle Balanus (=Amphibalanus) amphitrite. J. Exp. Biol. 2012, 215, 3813–3822. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.F.; Zhang, H.; He, L.; Liu, C.; Xu, Y.; Qian, P.Y. Butenolide inhibits marine fouling by altering the primary metabolism of three target organisms. ACS Chem. Biol. 2012, 7, 1049–1058. [Google Scholar] [CrossRef]
- Han, Z.; Sun, J.; Zhang, Y.; He, F.; Xu, Y.; Matsumura, K.; He, L.S.; Qiu, J.W.; Qi, S.H.; Qian, P.Y. iTRAQ-based proteomic profiling of the barnacle Balanus amphitrite in response to the antifouling compound meleagrin. J. Proteome. Res. 2013, 12, 2090–2100. [Google Scholar] [CrossRef]
- Artal-Sanz, M.; de Jong, L.; Tavernarakis, N. Caenorhabditis elegans: A versatile platform for drug discovery. Biotechnol. J. 2006, 1, 1405–1418. [Google Scholar] [CrossRef] [PubMed]
- Boyd, W.A.; McBride, S.J.; Rice, J.R.; Snyder, D.W.; Freedman, J.H. A high-throughput method for assessing chemical toxicity using a Caenorhabditis elegans reproduction assay. Toxicol. Appl. Pharmacol. 2010, 245, 153–159. [Google Scholar] [CrossRef] [Green Version]
- Burns, A.R.; Kwok, T.C.Y.; Howard, A.; Houston, E.; Johanson, K.; Chan, A.; Cutler, S.R.; McCourt, P.; Roy, P.J. High-throughput screening of small molecules for bioactivity and target identification in Caenorhabditis elegans. Nat. Protoc. 2006, 1, 1906–1914. [Google Scholar] [CrossRef]
- Kaletta, T.; Hengartner, M.O. Finding function in novel targets: C. elegans as a model organism. Nat. Rev. Drug Discov. 2006, 5, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Leung, M.C.K.; Williams, P.L.; Benedetto, A.; Au, C.; Helmcke, K.J.; Aschner, M.; Meyer, J.N. Caenorhabditis elegans: An emerging model in biomedical and environmental toxicology. Toxicol. Sci. 2008, 106, 5–28. [Google Scholar] [CrossRef] [PubMed]
- National Research Council. Scientific Frontiers in Developmental Toxicology and Risk Assessment; National Academy Press: Washington, DC, USA, 2000. [Google Scholar]
- National Toxicology Program. A National Toxicology Program for the 21st Century: Roadmap to Achieve the NTP Vision; National Institute of Environmental Health Sciences: Research Triangle, NC, USA, 2004.
- Bischof, L.J.; Huffman, D.L.; Aroian, R.V. Assays for Toxicity Studies in C. elegans with Bt Crystal Proteins. In C. elegans: Methods and Applications; Strange, K., Ed.; Humana Press: Totowa, NJ, USA, 2006; Volume 351, pp. 139–154. [Google Scholar]
- Breger, J.; Fuchs, B.B.; Aperis, G.; Moy, T.I.; Ausubel, F.M.; Mylonakis, E. Antifungal chemical compounds identified using a C. elegans pathogenicity assay. PLoS Pathog. 2007, 3, e18. [Google Scholar] [CrossRef]
- Heidarian, S.; Mohammadipanah, F.; Maghsoudlou, A.; Dashti, Y.; Challis, G. Anti-microfouling activity of Glycomyces sediminimaris UTMC 2460 on dominant fouling bacteria of Iran marine habitats. Front. Microbiol. 2019, 9, 3148. [Google Scholar] [CrossRef] [PubMed]
- Viju, N.; Satheesh, S.; Punitha, S.M.J. Antifouling activities of antagonistic marine bacterium Pseudomonas putida associated with an Octopus. Proc. Natl. Acad. Sci. India Sect. B Biol. Sci. 2015, 87, 1113–1124. [Google Scholar] [CrossRef]




















Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, K.-L.; Dou, Z.-R.; Gong, G.-F.; Li, H.-F.; Jiang, B.; Xu, Y. Anti-Larval and Anti-Algal Natural Products from Marine Microorganisms as Sources of Anti-Biofilm Agents. Mar. Drugs 2022, 20, 90. https://doi.org/10.3390/md20020090
Wang K-L, Dou Z-R, Gong G-F, Li H-F, Jiang B, Xu Y. Anti-Larval and Anti-Algal Natural Products from Marine Microorganisms as Sources of Anti-Biofilm Agents. Marine Drugs. 2022; 20(2):90. https://doi.org/10.3390/md20020090
Chicago/Turabian StyleWang, Kai-Ling, Zheng-Rong Dou, Gao-Fen Gong, Hai-Feng Li, Bei Jiang, and Ying Xu. 2022. "Anti-Larval and Anti-Algal Natural Products from Marine Microorganisms as Sources of Anti-Biofilm Agents" Marine Drugs 20, no. 2: 90. https://doi.org/10.3390/md20020090
APA StyleWang, K. -L., Dou, Z. -R., Gong, G. -F., Li, H. -F., Jiang, B., & Xu, Y. (2022). Anti-Larval and Anti-Algal Natural Products from Marine Microorganisms as Sources of Anti-Biofilm Agents. Marine Drugs, 20(2), 90. https://doi.org/10.3390/md20020090