
First Identification of 12β-Deoxygonyautoxin 5 (12α-Gonyautoxinol 5) in the Cyanobacterium Dolichospermum circinale (TA04) and 12β-Deoxysaxitoxin (12α-Saxitoxinol) in D. circinale (TA04) and the Dinoflagellate Alexandrium pacificum (Group IV) (120518KureAC)

 , ,
, ,
Abstract
:1. Introduction

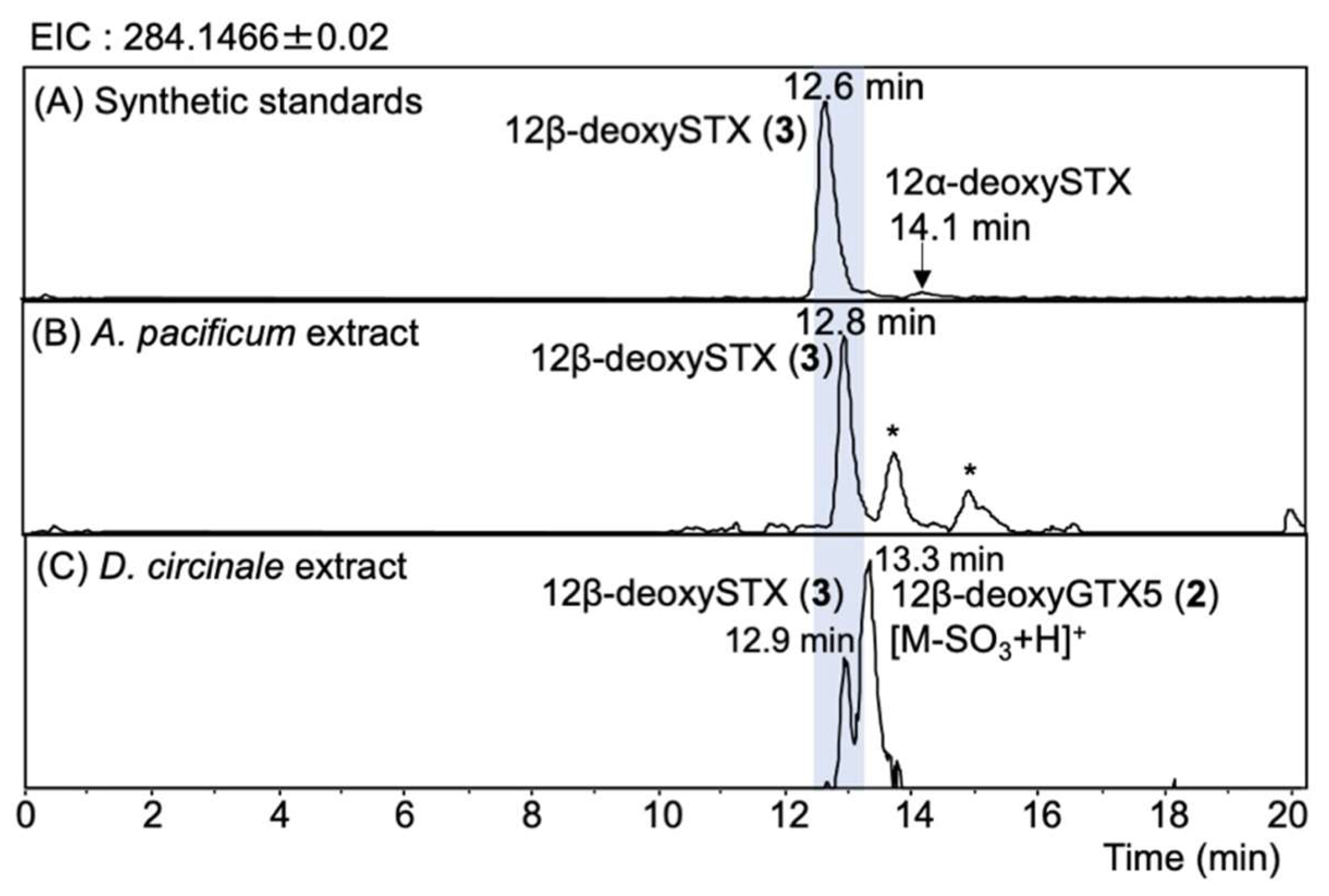
2. Results
2.1. Screening for New PSTs in the PST-Producing Cyanobacterium D. circinale (TA04) and the Dinoflagellate A. pacificum (Group IV) (120518KureAC)
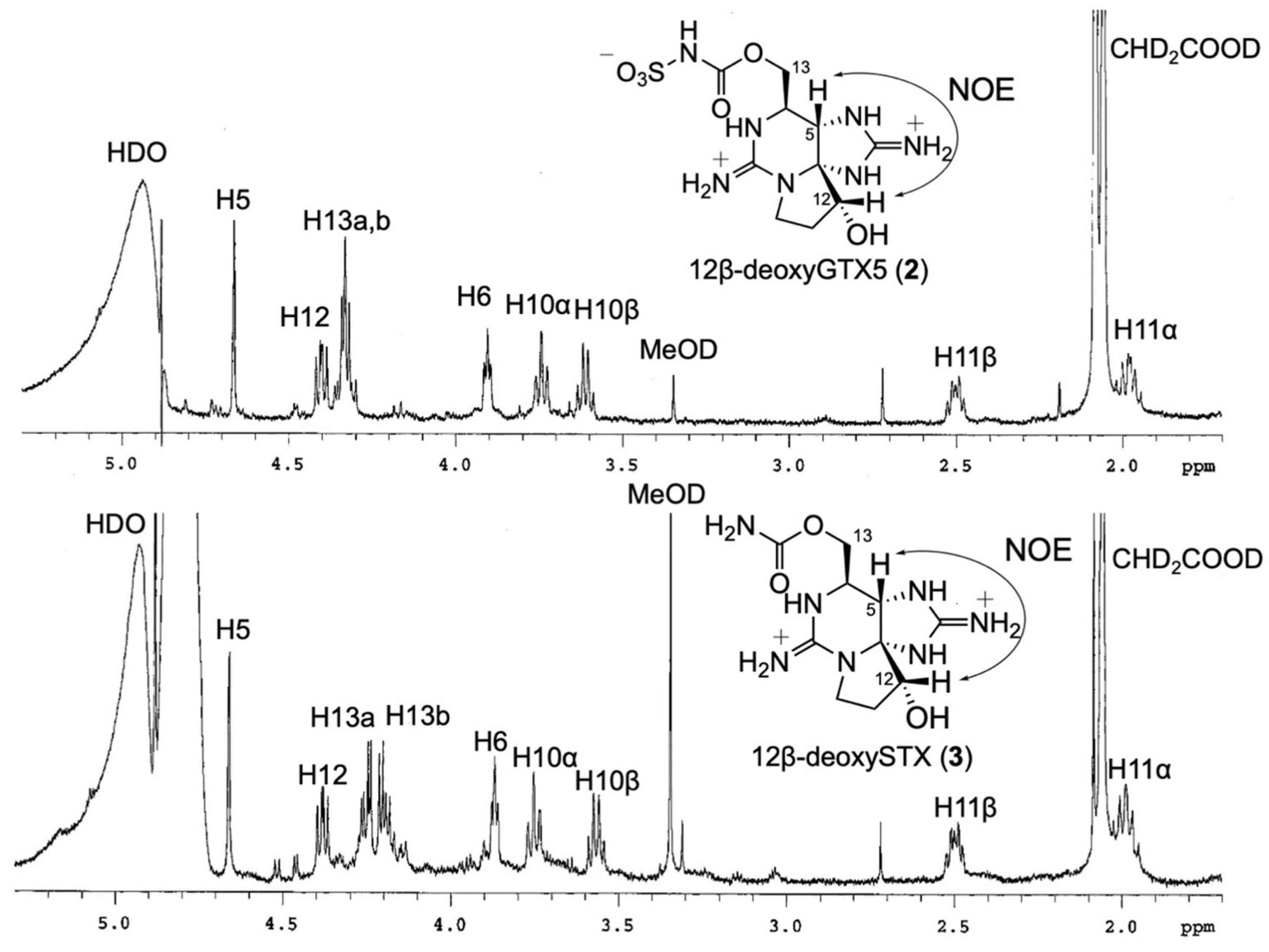
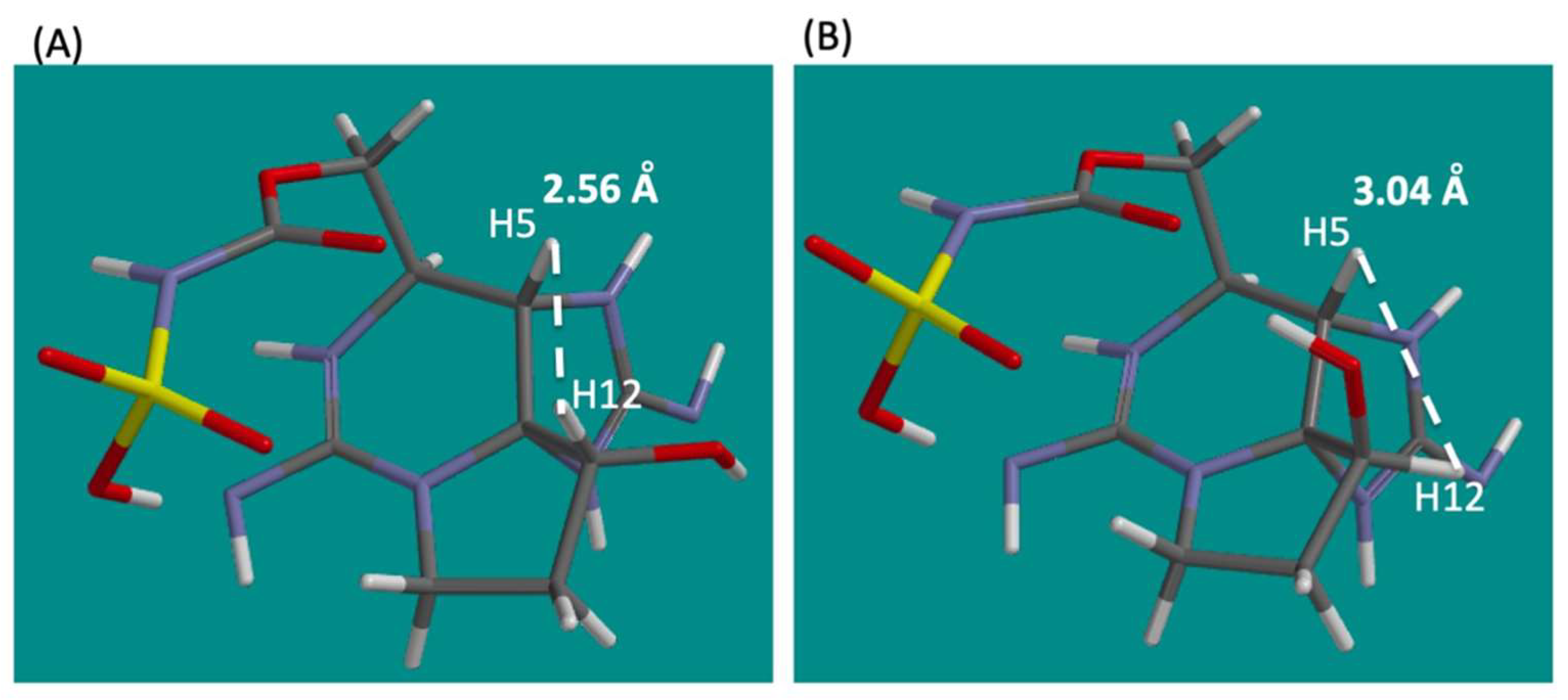
2.2. Chemical Preparation of Authentic 12β-DeoxyGTX5 (2), 12β-DeoxySTX (3), and 12α-DeoxyGTX5 (5)
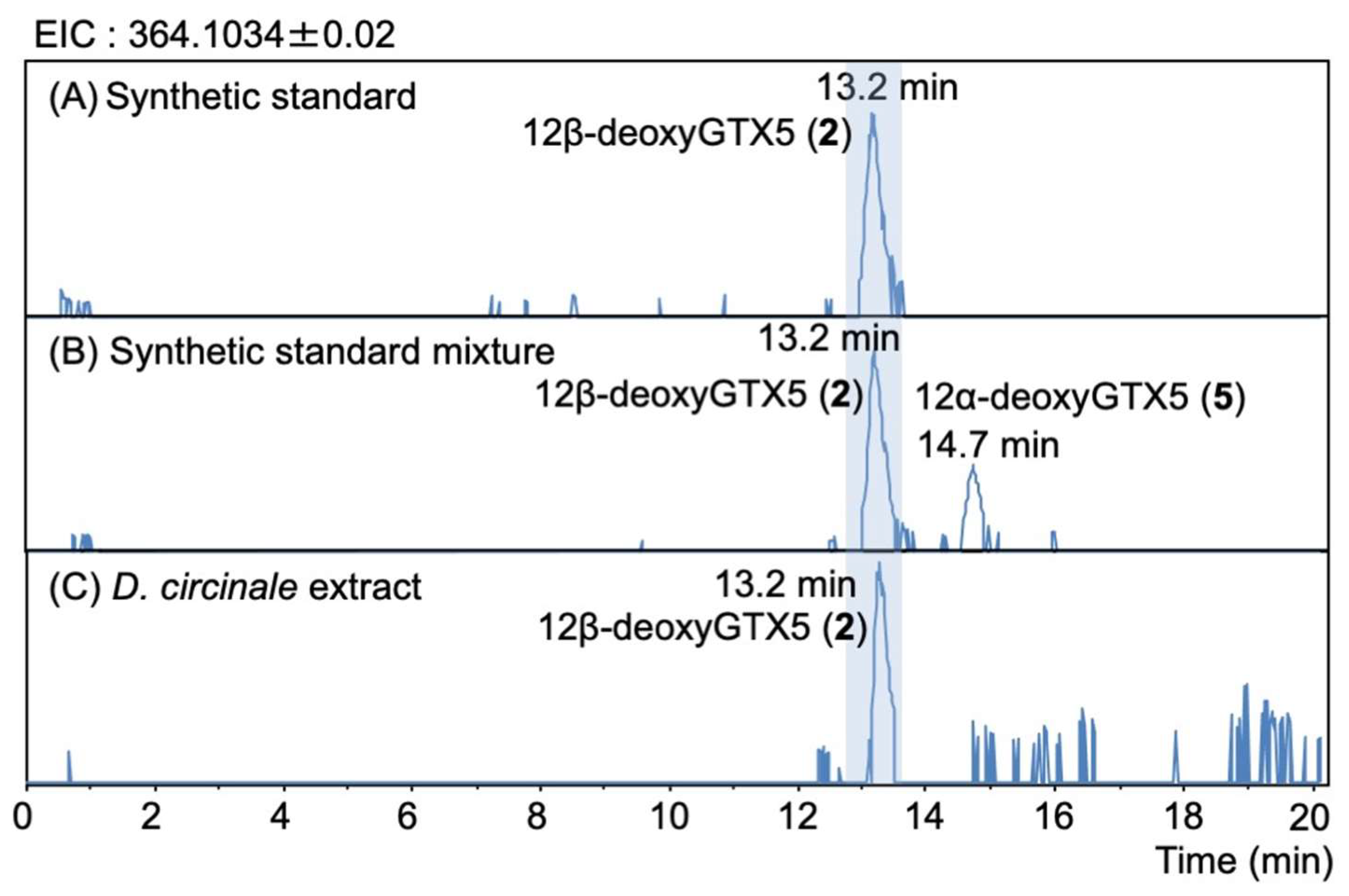
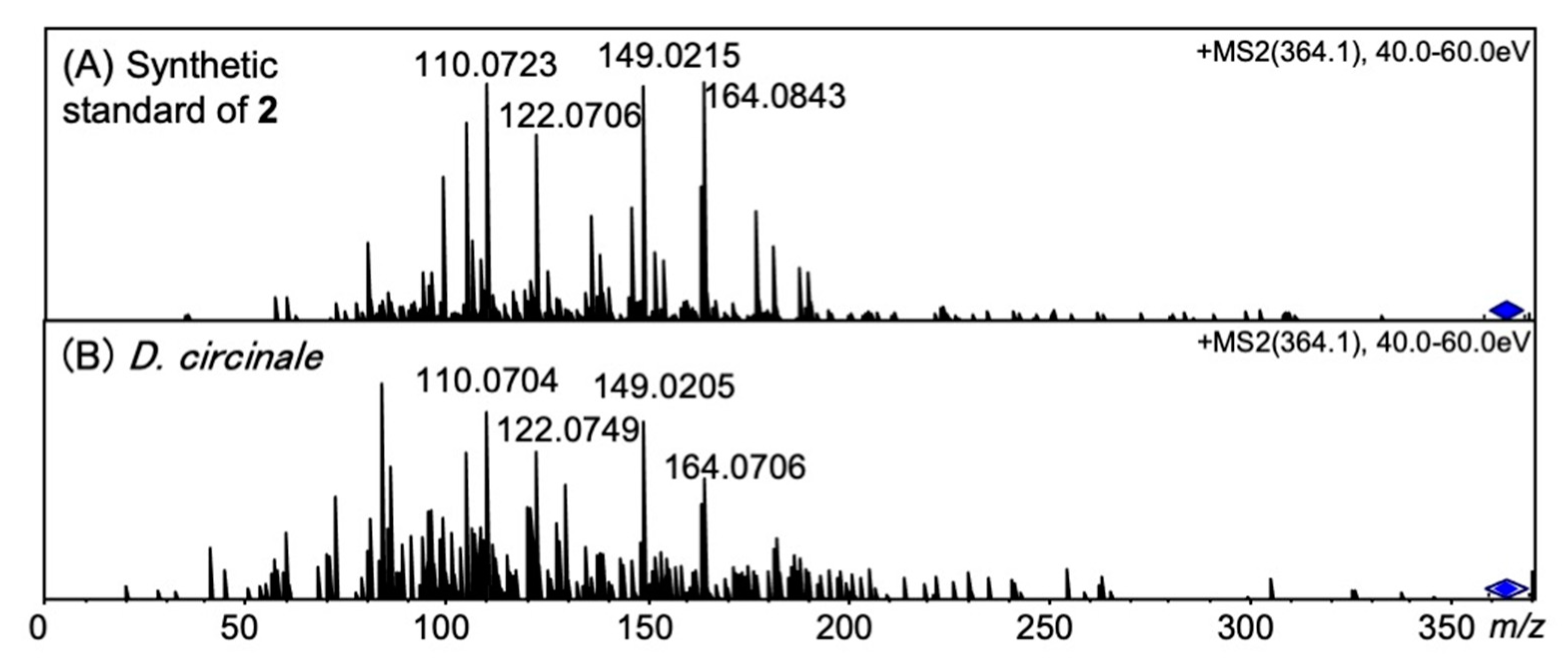
2.3. Identification of 12β-DeoxyGTX5 (2) in D. circinale (TA04)
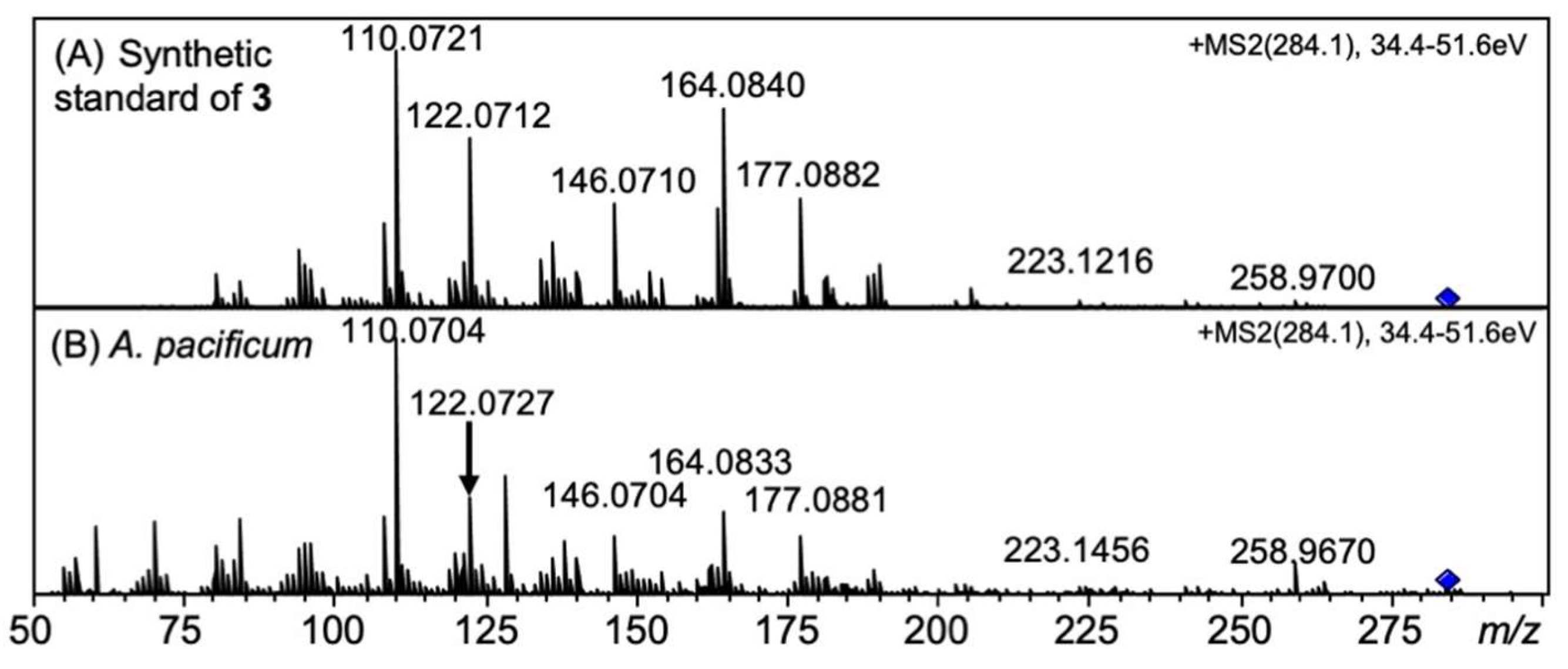
2.4. Identification of 12β-DeoxySTX (3) (12α-STXol) in A. pacificum (Group IV) (120518KureAC) and D. circinale (TA04)
3. Discussion
4. Methods and Materials
4.1. General Information
4.2. Preparation of 12β-DeoxyGTX5 (2) and 12β-DeoxySTX (3) from C1/C2
4.3. Preparation of the Mixture of 12α/β-DeoxyGTX5 (2, 5) from GTX5 for LCMS Analysis
4.4. Preparation of 12α-DeoxyGTX5 (5) for NMR Analysis
4.5. Harvest and Preparation of D. circinale (TA04) Cell Extract
4.6. Harvest and Preparation of Alexandrium pacificum (Group IV) (120518KureAC) Cell Extract
4.7. HR-RP-LCMS and HR-RP-LC-MS/MS Conditions for PSTs Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kao, C.Y.; Nishiyama, A. Actions of saxitoxin on peripheral neuromuscular systems. J. Physiol. 1965, 180, 50–66. [Google Scholar] [PubMed]
- Llewellyn, L.E. Saxitoxin, a toxic marine natural product that targets a multitude of receptors. Nat. Prod. Rep. 2006, 23, 200–222. [Google Scholar] [CrossRef] [PubMed]
- Thottumkara, A.P.; Parsons, W.H.; Du Bois, J. Saxitoxin. Angew. Chem. Int. Ed. 2014, 53, 5760–5784. [Google Scholar] [CrossRef] [PubMed]
- Wiese, M.; D’Agostino, P.M.; Mihali, T.K.; Moffitt, M.C.; Neilan, B.A. Neurotoxic alkaloids: Saxitoxin and its analogs. Mar. Drugs 2010, 8, 2185–2211. [Google Scholar] [CrossRef] [Green Version]
- Carmichael, W.W.; Evans, W.R.; Yin, Q.Q.; Bell, P.; Moczydlowski, E. Evidence for paralytic shellfish poisons in the freshwater cyanobacterium Lyngbya wollei (Farlow ex Gomont) comb. nov. Appl. Environ. Microbiol. 1997, 63, 3104–3110. [Google Scholar] [CrossRef] [Green Version]
- Anderson, D.M.; Alpermann, T.J.; Cembella, A.D.; Collos, Y.; Masseret, E.; Montresor, M. The globally distributed genus Alexandrium: Multifaceted roles in marine ecosystems and impacts on human health. Harmful Algae 2012, 14, 10–35. [Google Scholar] [CrossRef] [Green Version]
- Adachi, K.; Yamada, T.; Ishizuka, H.; Oki, M.; Tsunogae, S.; Shimada, N.; Chiba, O.; Orihara, T.; Hidaka, M.; Hirokawa, T.; et al. Synthesis of C12-keto saxitoxin derivatives with unusual inhibitory activity against voltage-gated sodium channels. Chem.—A Eur. J. 2019, 26, 2025–2033. [Google Scholar] [CrossRef]
- Shen, H.; Liu, D.; Wu, K.; Lei, J.; Yan, N. Structures of human Nav1.7 channel in complex with auxiliary subunits and animal toxins. Science 2019, 363, 1303–1308. [Google Scholar] [CrossRef]
- Shimizu, Y.; Norte, M.; Hori, A.; Genenah, A.; Kobayashi, M. Biosynthesis of saxitoxin analogues: The unexpected pathway. J. Am. Chem. Soc. 1984, 106, 6433–6434. [Google Scholar] [CrossRef]
- Kellmann, R.; Mihali, T.K.; Young, J.J.; Pickford, R.; Pomati, F.; Neilan, B.A. Biosynthetic intermediate analysis and functional homology reveal a saxitoxin gene cluster in cyanobacteria. Appl. Environ. Microbiol. 2008, 74, 4044–4053. [Google Scholar] [CrossRef] [Green Version]
- Stüken, A.; Orr, R.J.S.; Kellmann, R.; Murray, S.A.; Neilan, B.A.; Jakobsen, K.S. Discovery of nuclear-encoded genes for the neurotoxin saxitoxin in dinoflagellates. PLoS ONE 2011, 6, e20096. [Google Scholar] [CrossRef] [Green Version]
- Tsuchiya, S.; Cho, Y.; Konoki, K.; Nagasawa, K.; Oshima, Y.; Yotsu-Yamashita, M. Synthesis and identification of proposed biosynthetic intermediates of saxitoxin in the cyanobacterium Anabaena circinalis (TA04) and the dinoflagellate Alexandrium tamarense (Axat-2). Org. Biomol. Chem. 2014, 12, 3016–3020. [Google Scholar] [CrossRef]
- Tsuchiya, S.; Cho, Y.; Konoki, K.; Nagasawa, K.; Oshima, Y.; Yotsu-Yamashita, M. Synthesis of a tricyclic bisguanidine compound structurally related to saxitoxin and its identification in paralytic shellfish toxin-producing microorganisms. Chem.—A Eur. J. 2015, 21, 7835–7840. [Google Scholar] [CrossRef]
- Tsuchiya, S.; Cho, Y.; Konoki, K.; Nagasawa, K.; Oshima, Y.; Yotsu-Yamashita, M. Biosynthetic route towards saxitoxin and shunt pathway. Sci. Rep. 2016, 6, 1–9. [Google Scholar]
- Tsuchiya, S.; Cho, Y.; Yoshioka, R.; Konoki, K.; Nagasawa, K.; Oshima, Y.; Yotsu-Yamashita, M. Synthesis and Identification of Key Biosynthetic Intermediates for the Formation of the Tricyclic Skeleton of Saxitoxin. Angew. Chem. Int. Ed. 2017, 56, 5327–5331. [Google Scholar] [CrossRef]
- Cho, Y.; Hidema, S.; Omura, T.; Koike, K.; Koike, K.; Oikawa, H.; Konoki, K.; Oshima, Y.; Yotsu-Yamashita, M. SxtA localizes to chloroplasts and changes to its 3′UTR may reduce toxin biosynthesis in non-toxic Alexandrium catenella (Group I). Harmful Algae 2021, 101, 101972. [Google Scholar] [CrossRef]
- Lukowski, A.L.; Ellinwood, D.C.; Hinze, M.E.; Deluca, R.J.; Du Bois, J.; Hall, S.; Narayan, A.R.H. C-H Hydroxylation in Paralytic Shellfish Toxin Biosynthesis. J. Am. Chem. Soc. 2018, 140, 11863–11869. [Google Scholar] [CrossRef]
- Lukowski, A.L.; Denomme, N.; Hinze, M.E.; Hall, S.; Isom, L.L.; Narayan, A.R.H. Biocatalytic detoxification of paralytic shellfish toxins. ACS Chem. Biol. 2019, 14, 941–948. [Google Scholar] [CrossRef] [Green Version]
- Lukowski, A.L.; Mallik, L.; Hinze, M.E.; Carlson, B.M.; Ellinwood, D.C.; Pyser, J.B.; Koutmos, M.; Narayan, A.R.H. Substrate promiscuity of a paralytic shellfish toxin amidinotransferase. ACS Chem. Biol. 2020, 15, 626–631. [Google Scholar] [CrossRef]
- Lukowski, A.L.; Liu, J.; Bridwell-rabb, J.; Narayan, A.R.H. Structural basis for divergent C—H hydroxylation selectivity in two Rieske oxygenases. Nat. Commun. 2020, 11, 2991. [Google Scholar] [CrossRef]
- Chun, S.W.; Hinze, M.E.; Skiba, M.A.; Narayan, A.R.H. Chemistry of a unique polyketide-like synthase. J. Am. Chem. Soc. 2018, 140, 2430–2433. [Google Scholar] [CrossRef] [PubMed]
- Soeriyadi, A.H.; Mazmouz, R.; Pickford, R.; Al-Sinawi, B.; Kellmann, R.; Pearson, L.A.; Neilan, B.A. Heterologous expression of an unusual ketosynthase, SxtA, leads to production of saxitoxin intermediates in Escherichia coli. ChemBioChem 2021, 22, 845–849. [Google Scholar] [CrossRef] [PubMed]
- Harada, T.; Oshima, Y.; Yasumoto, T. Structures of two paralytic shellfish toxins, gonyautoxins V and VI, isolated from a tropical dinoflagellate, Pyrodinium bahamense var. compressa. Agric. Biol. Chem. 1982, 46, 1861–1864. [Google Scholar]
- Dell’ Aversano, C.D.; Walter, J.A.; Burton, I.W.; Stirling, D.J.; Fattorusso, E.; Quilliam, M.A. Isolation and structure elucidation of new and unusual saxitoxin analogues from mussels. J. Nat. Prod. 2008, 71, 1518–1523. [Google Scholar] [CrossRef]
- Numano, S.; Kudo, Y.; Cho, Y.; Konoki, K.; Yotsu-Yamashita, M. Temporal variation of the profile and concentrations of paralytic shellfish toxins and tetrodotoxin in the scallop, Patinopecten yessoensis, cultured in a bay of East Japan. Mar. Drugs 2019, 17, 653. [Google Scholar] [CrossRef] [Green Version]
- Onodera, H.; Satake, M.; Oshima, Y.; Yasumoto, T.; Carmichael, W.W. New saxitoxin analogues from the freshwater filamentous cyanobacterium Lyngbya wollei. Nat. Toxins 1998, 5, 146–151. [Google Scholar] [CrossRef]
- Cho, Y.; Tsuchiya, S.; Yoshioka, R.; Omura, T.; Konoki, K.; Oshima, Y.; Yotsu-Yamashita, M. Column switching combined with hydrophilic interaction chromatography-tandem mass spectrometry for the analysis of saxitoxin analogues, and their biosynthetic intermediates in dinoflagellates. J. Chromatogr. A 2016, 1474, 109–120. [Google Scholar] [CrossRef]
- Lim, P.T.; Sato, S.; Van Thuoc, C.; Tu, P.T.; Huyen, N.T.M.; Takata, Y.; Yoshida, M.; Kobiyama, A.; Koike, K.; Ogata, T. Toxic Alexandrium minutum (Dinophyceae) from Vietnam with new gonyautoxin analogue. Harmful Algae 2007, 6, 321–331. [Google Scholar] [CrossRef]
- Minowa, T.; Cho, Y.; Oshima, Y.; Konoki, K.; Yotsu-Yamashita, M. Identification of a novel saxitoxin analogue, 12β-deoxygonyautoxin 3, in the cyanobacterium, Anabaena circinalis (TA04). Toxins 2019, 11, 539. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, R.; Suzuki, T.; Oshima, Y. Preparation of calibration standards of N1-H paralytic shellfish toxin analogues by large-scale culture of cyanobacterium Anabaena circinalis (TA04). Mar. Drugs 2011, 9, 466–477. [Google Scholar] [CrossRef]
- Koehn, F.E.; Ghazarossian, V.E.; Schantz, E.J.; Schnoes, H.K.; Strong, F.M. Derivatives of saxitoxin. Bioorg. Chem. 1981, 10, 412–428. [Google Scholar] [CrossRef]
- Negri, A.P.; Jones, G.; Blackburn, S.I.; Oshima, Y.; Onodera, H. Effect of culture and bloom development and sample storage on paralytic shellfish poisons in the cyanobacterum Anabaena circinalis. J. Phycol. 1997, 33, 26–35. [Google Scholar] [CrossRef]
- Cho, Y.; Tsuchiya, S.; Omura, T.; Koike, K.; Oikawa, H.; Konoki, K.; Oshima, Y.; Yotsu-Yamashita, M. Metabolomic study of saxitoxin analogues and biosynthetic intermediates in dinoflagellates using 15N-labelled sodium nitrate as a nitrogen source. Sci. Rep. 2019, 9, 2–12. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 12β-deoxyGTX5 (12α-GTXol 5) (2) * | 12β-deoxySTX (12α-STXol) (3) * | 12α-deoxyGTX5 (12β-GTXol 5) (5) * | ||||
|---|---|---|---|---|---|---|
| No. | δH | Multiplicity | δH | Multiplicity | δH | Multiplicity |
| (J in Hz) | (J in Hz) | (J in Hz) | ||||
| 5 | 4.67 | s | 4.66 | s | 4.81 | s |
| 6 | 3.91 | t 5.3 | 3.87 | t 5.2 | 3.88 | dd 9.6, 5.6 |
| 10α | 3.74 | t 9.8 | 3.75 | t 9.7 | 3.77 | t 10.0 |
| 10β | 3.61 | q 9.5 | 3.56 | q 9.3 | 3.72 | t 9.1 |
| 11α | 1.98 | m | 1.99 | m | 2.25 ** | dd 6.5, 12.1 |
| 11β | 2.50 | quint. 9.7 | 2.50 | quint. 10.9 | 2.41 ** | m |
| 12 | 4.40 | dd 7.6, 11.8 | 4.38 | dd 7.6, 11.8 | 4.34 | d 4.7 |
| 13a | 4.33 | m | 4.25 | dd 4.6, 11.8 | 4.39 | t 9.1 |
| 13b | 4.33 | m | 4.20 | dd 6.4, 11.8 | 4.13 | dd 5.9, 9.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akamatsu, M.; Hirozumi, R.; Cho, Y.; Kudo, Y.; Konoki, K.; Oshima, Y.; Yotsu-Yamashita, M. First Identification of 12β-Deoxygonyautoxin 5 (12α-Gonyautoxinol 5) in the Cyanobacterium Dolichospermum circinale (TA04) and 12β-Deoxysaxitoxin (12α-Saxitoxinol) in D. circinale (TA04) and the Dinoflagellate Alexandrium pacificum (Group IV) (120518KureAC). Mar. Drugs 2022, 20, 166. https://doi.org/10.3390/md20030166
Akamatsu M, Hirozumi R, Cho Y, Kudo Y, Konoki K, Oshima Y, Yotsu-Yamashita M. First Identification of 12β-Deoxygonyautoxin 5 (12α-Gonyautoxinol 5) in the Cyanobacterium Dolichospermum circinale (TA04) and 12β-Deoxysaxitoxin (12α-Saxitoxinol) in D. circinale (TA04) and the Dinoflagellate Alexandrium pacificum (Group IV) (120518KureAC). Marine Drugs. 2022; 20(3):166. https://doi.org/10.3390/md20030166
Chicago/Turabian StyleAkamatsu, Michiru, Ryosuke Hirozumi, Yuko Cho, Yuta Kudo, Keiichi Konoki, Yasukatsu Oshima, and Mari Yotsu-Yamashita. 2022. "First Identification of 12β-Deoxygonyautoxin 5 (12α-Gonyautoxinol 5) in the Cyanobacterium Dolichospermum circinale (TA04) and 12β-Deoxysaxitoxin (12α-Saxitoxinol) in D. circinale (TA04) and the Dinoflagellate Alexandrium pacificum (Group IV) (120518KureAC)" Marine Drugs 20, no. 3: 166. https://doi.org/10.3390/md20030166
APA StyleAkamatsu, M., Hirozumi, R., Cho, Y., Kudo, Y., Konoki, K., Oshima, Y., & Yotsu-Yamashita, M. (2022). First Identification of 12β-Deoxygonyautoxin 5 (12α-Gonyautoxinol 5) in the Cyanobacterium Dolichospermum circinale (TA04) and 12β-Deoxysaxitoxin (12α-Saxitoxinol) in D. circinale (TA04) and the Dinoflagellate Alexandrium pacificum (Group IV) (120518KureAC). Marine Drugs, 20(3), 166. https://doi.org/10.3390/md20030166