
Degradation of Alginate by a Newly Isolated Marine Bacterium Agarivorans sp. B2Z047

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
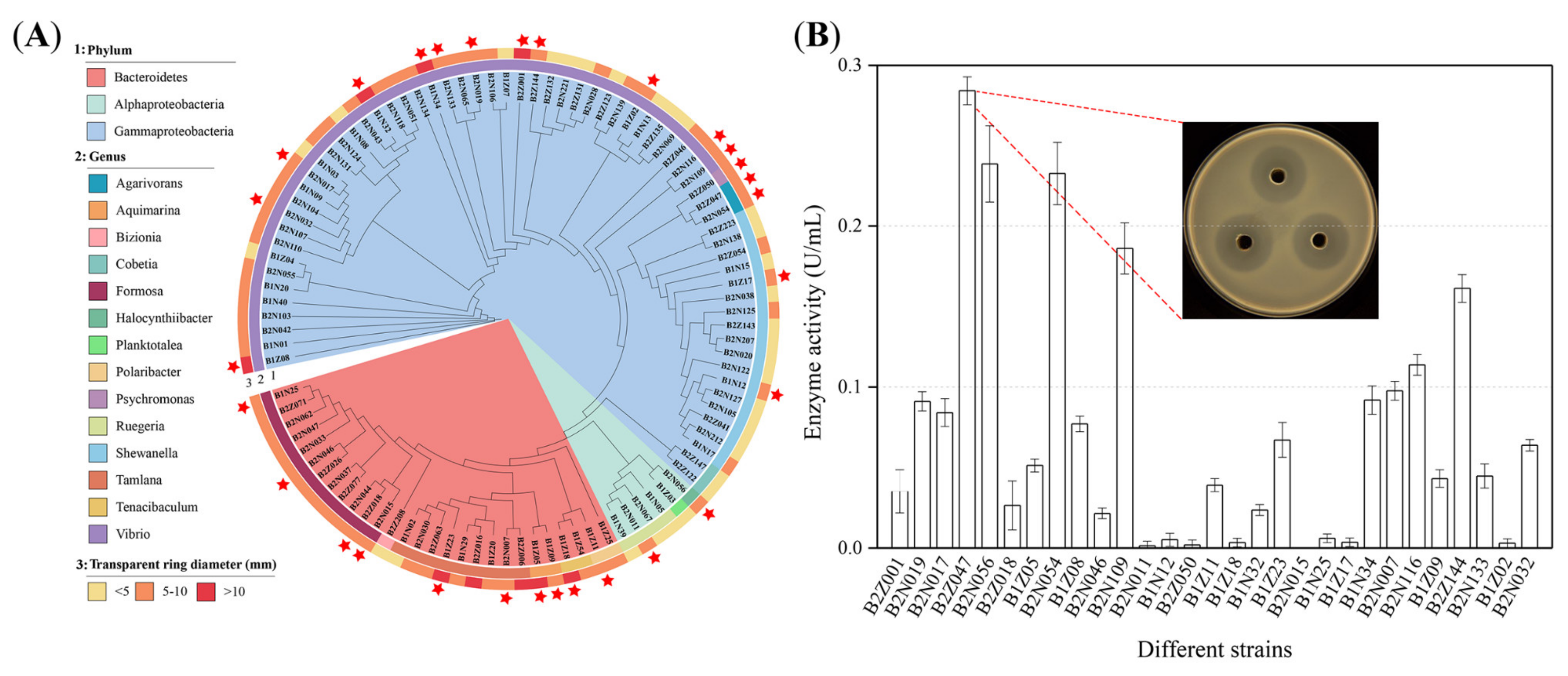
2.1. Isolation and Cultivation of Alginate-Degrading Strains
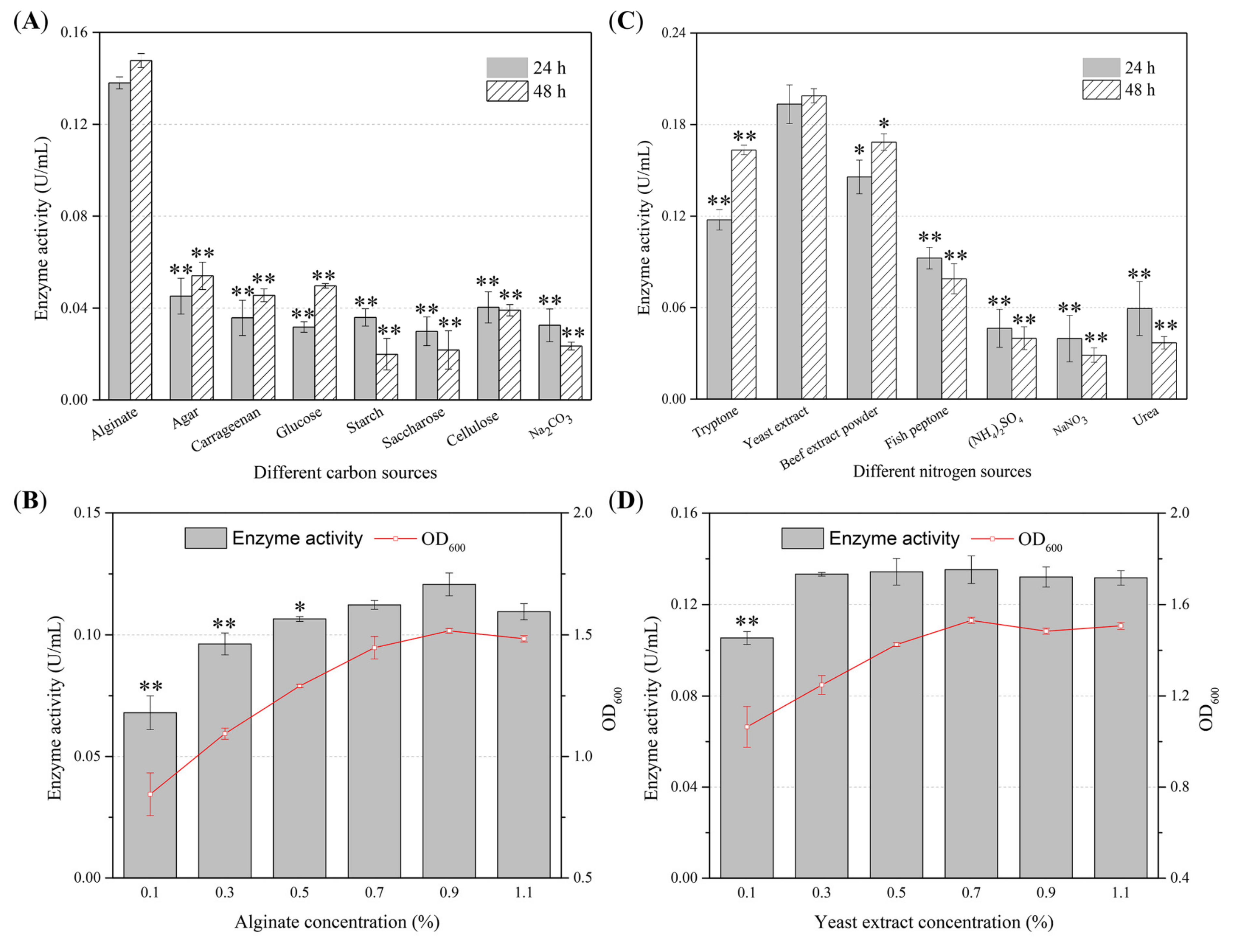
2.2. Optimization of the Production Conditions of the Alginate Lyase by Agarivorans sp. B2Z047
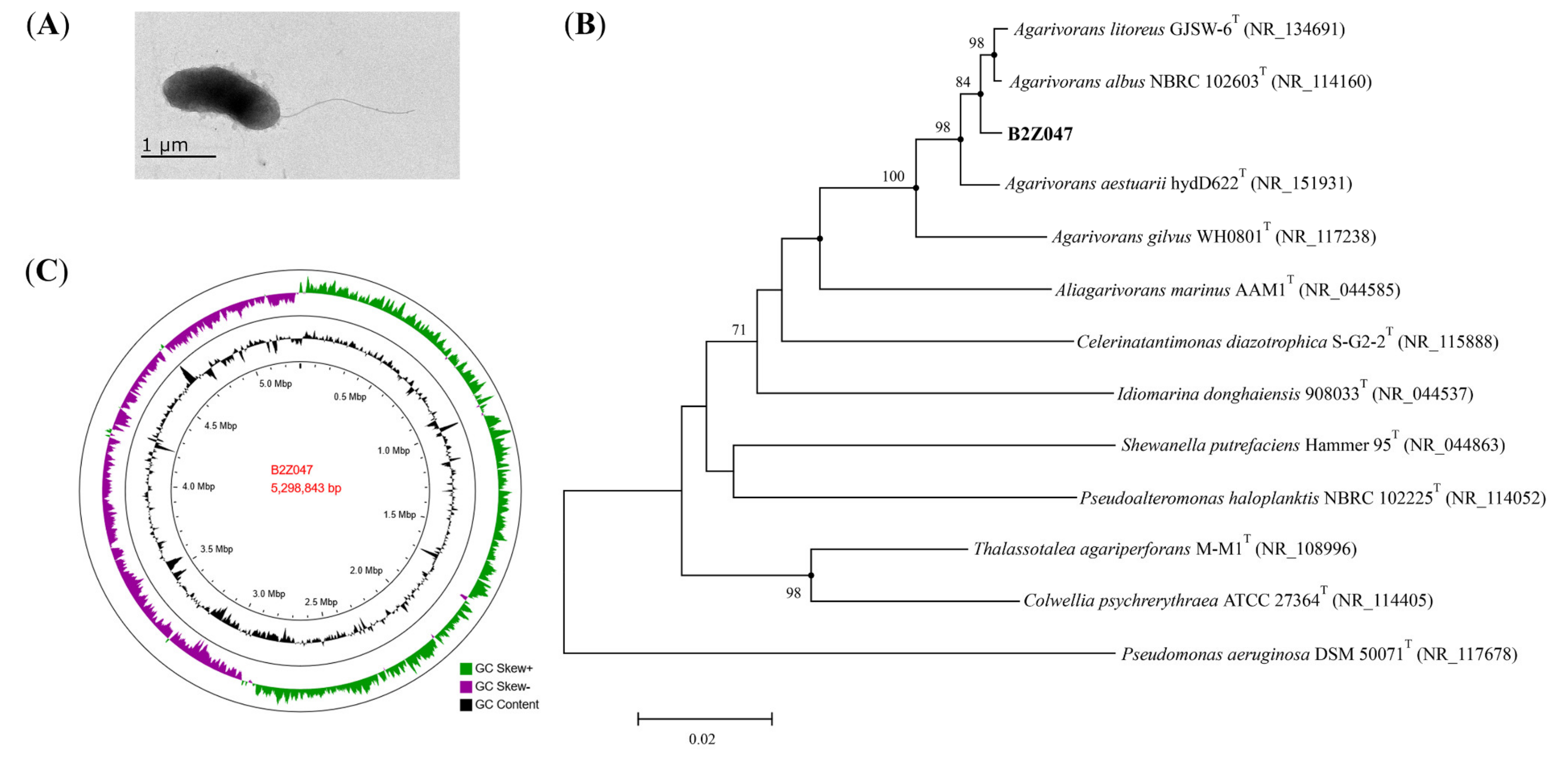
2.3. Characterization of Strain B2Z047
2.4. Sequence Analyses of Alginate Lyases
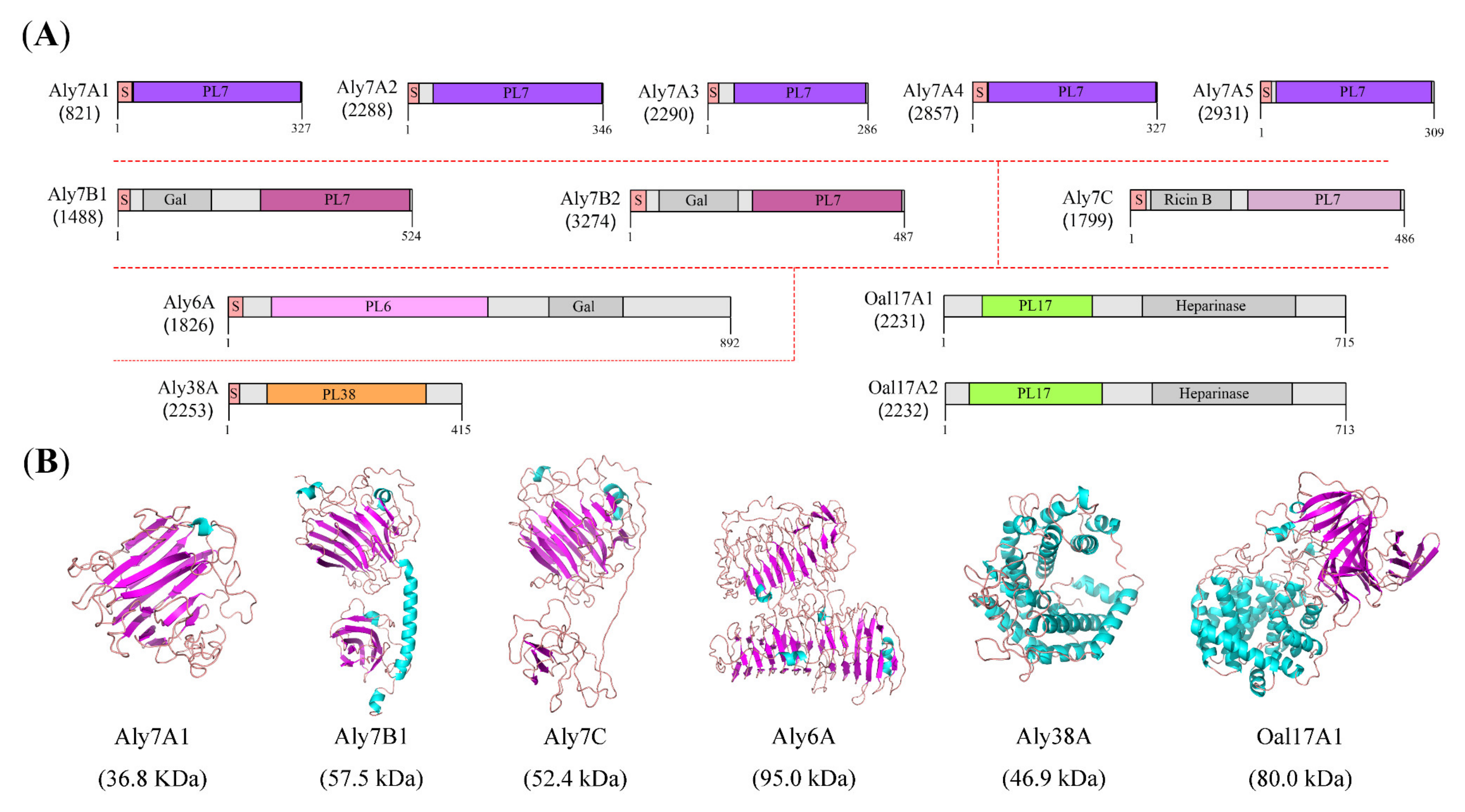
2.5. Conserved Domain and Protein Structure Prediction of Alginate Lyases
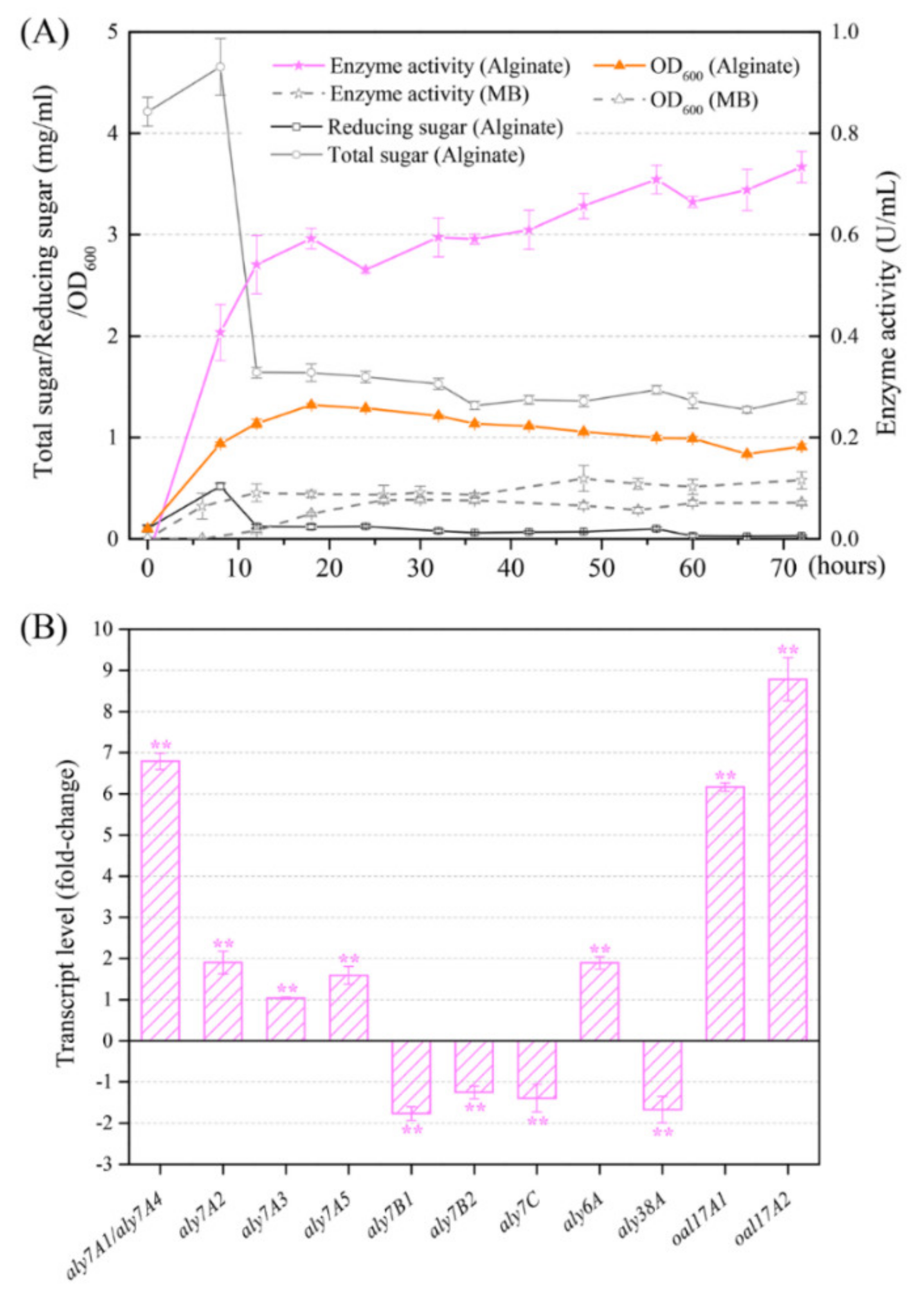
2.6. Enzymatic Activity of Alginate Lyases from Strain B2Z047
2.7. Identification of Key Alginate Lyases
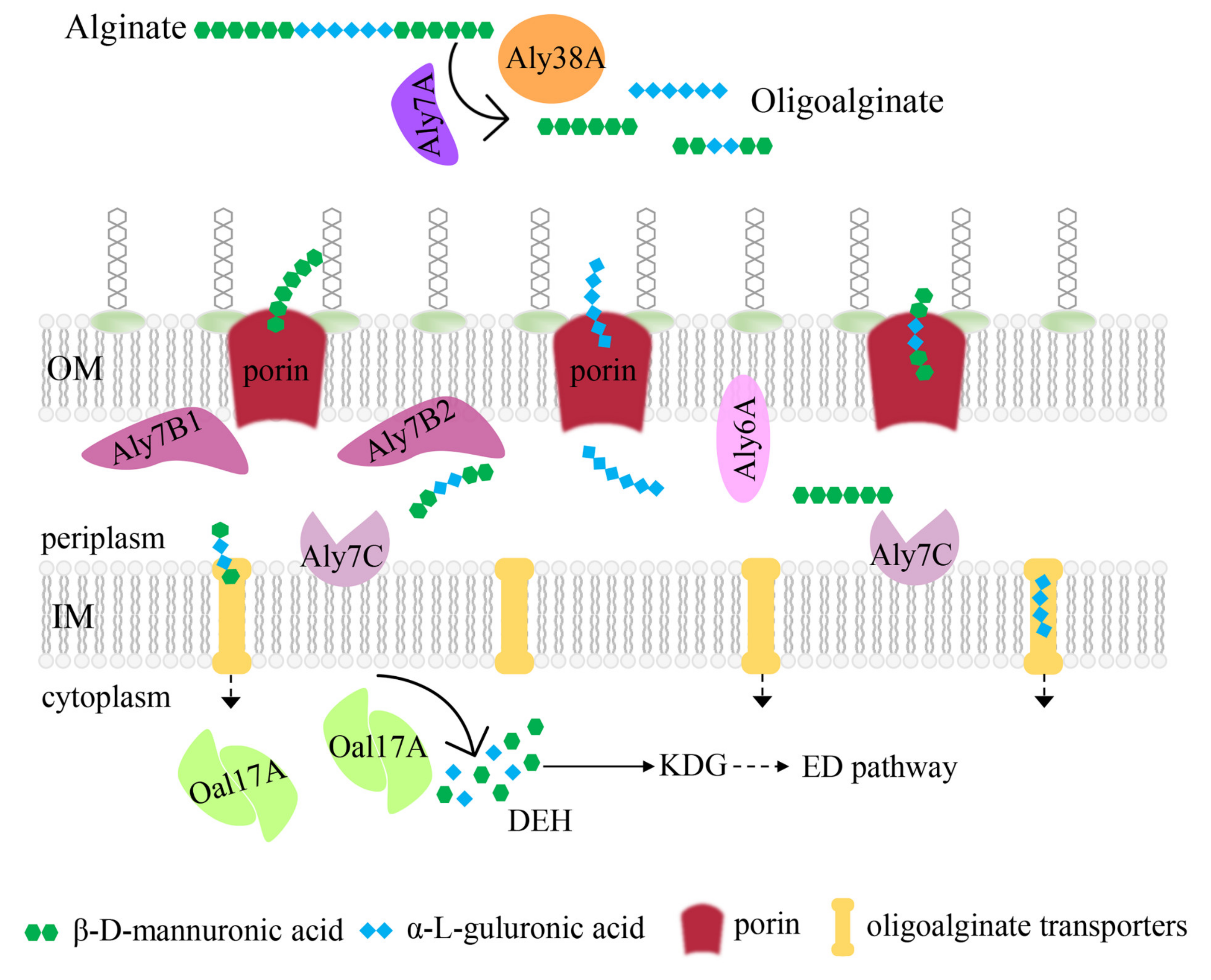
2.8. Potential Alginate-Degrading Pathway of Strain B2Z047
3. Materials and Methods
3.1. Alginate-Degrading Strains Isolated
3.2. Enzyme Activity Assay
3.3. Optimization of Alginate Lyase Production Conditions
3.4. Whole Genome Sequencing, Assembly, and Annotation
3.5. Phenotypic Characteristics
3.6. 16S rRNA Gene Sequence Analyses
3.7. Alginate Lyase Sequence Analyses
3.8. Cloning and Expression of Alginate Lyase and Its Degradation Products
3.9. Total Sugar Assay
3.10. q-PCR Assay Based on SYBR Green
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Field, C.B.; Behrenfeld, M.J.; Randerson, J.T.; Falkowski, P. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science 1998, 281, 237–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haug, A.; Larsen, B.; Smidsrod, O. Studies on sequence of uronic acid residues in alginic acid. Acta Chem. Scand. 1967, 21, 691–704. [Google Scholar] [CrossRef] [Green Version]
- Strand, B.L.; Morch, Y.A.; Skjak-Braek, G. Alginate as immobilization matrix for cells. Minerva Biotecnol. 2000, 12, 223–233. [Google Scholar]
- Liu, J.; Yang, S.; Li, X.; Yan, Q.; Reaney, M.J.T.; Jiang, Z. Alginate oligosaccharides: Production, biological activities, and potential applications. Compr. Rev. Food Sci. Food Saf. 2019, 18, 1859–1881. [Google Scholar] [CrossRef] [Green Version]
- Zhu, B.; Yin, H. Alginate lyase: Review of major sources and classification, properties, structure-function analysis and applications. Bioengineered 2015, 6, 125–131. [Google Scholar] [CrossRef] [Green Version]
- Cheng, D.; Jiang, C.; Xu, J.; Liu, Z.; Mao, X. Characteristics and applications of alginate lyases: A review. Int. J. Biol. Macromol. 2020, 164, 1304–1320. [Google Scholar] [CrossRef]
- Yu, Z.C.; Zhu, B.W.; Wang, W.X.; Tan, H.D.; Yin, H. Characterization of a new oligoalginate lyase from marine bacterium Vibrio sp. Int. J. Biol. Macromol. 2018, 112, 937–942. [Google Scholar] [CrossRef]
- Li, S.Y.; Wang, L.N.; Chen, X.H.; Zhao, W.W.; Sun, M.; Han, Y.T. Cloning, expression, and biochemical characterization of two new oligoalginate lyases with synergistic degradation capability. Mar. Biotechnol. 2018, 20, 75–86. [Google Scholar] [CrossRef]
- Park, D.; Jagtap, S.; Nair, S.K. Structure of a PL17 family alginate lyase demonstrates functional similarities among exotype depolymerases. J. Biol. Chem. 2014, 289, 8645–8655. [Google Scholar] [CrossRef] [Green Version]
- Ochiai, A.; Yamasaki, M.; Mikami, B.; Hashimoto, W.; Murata, K. Crystal structure of exotype alginate lyase Atu3025 from Agrobacterium tumefaciens. J. Biol. Chem. 2010, 285, 24519–24528. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Li, X.; Zhang, X.; Li, Y.; Wang, L. Bacterial alginate metabolism: An important pathway for bioconversion of brown algae. Biotechnol. Biofuels 2021, 14, 1–18. [Google Scholar] [CrossRef]
- Wang, B.; Dong, S.; Li, F.L.; Ma, X.Q. Structural basis for the exolytic activity of polysaccharide lyase family 6 alginate lyase BcAlyPL6 from human gut microbe Bacteroides clarus. Biochem. Biophys. Res. Commun. 2021, 547, 111–117. [Google Scholar] [CrossRef]
- Lyu, Q.Q.; Zhang, K.K.; Zhu, Q.Y.; Li, Z.J.; Liu, Y.J.; Fitzek, E.; Yohe, T.; Zhao, L.M.; Li, W.H.; Liu, T.; et al. Structural and biochemical characterization of a multidomain alginate lyase reveals a novel role of CBM32 in CAZymes. Biochim. Biophys. Acta-Gen. Subj. 2018, 1862, 1862–1869. [Google Scholar] [CrossRef]
- Jouanneau, D.; Klau, L.J.; Larocque, R.; Jaffrennou, A.; Duval, G.; Le Duff, N.; Roret, T.; Jeudy, A.; Aachmann, F.L.; Czjzek, M.; et al. Structure-function analysis of a new PL17 oligoalginate lyase from the marine bacterium Zobellia galactanivorans DsijT. Glycobiology 2021, 31, 1364–1377. [Google Scholar] [CrossRef]
- Kikuchi, M.; Konno, N.; Suzuki, T.; Fujii, Y.; Kodama, Y.; Isogai, A.; Habu, N. A bacterial endo-β-1,4-glucuronan lyase, CUL-I from Brevundimonas sp. SH203, belonging to a novel polysaccharide lyase family. Protein Expr. Purif. 2020, 166, 105502. [Google Scholar] [CrossRef]
- Pilgaard, B.; Vuillemin, M.; Munk, L.; Holck, J.; Meier, S.; Wilkens, C.; Meyer, A.S. Discovery of a novel glucuronan lyase system in Trichoderma parareesei. Appl. Environ. Microbiol. 2022, 88, e01819-21. [Google Scholar] [CrossRef]
- Li, M.M.; Shang, Q.S.; Li, G.S.; Wang, X.; Yu, G.L. Degradation of marine algae-derived carbohydrates by Bacteroidetes isolated from human gut microbiota. Mar. Drugs 2017, 15, 92. [Google Scholar] [CrossRef]
- Tang, J.C.; Taniguchi, H.; Chu, H.; Zhou, Q.; Nagata, S. Isolation and characterization of alginate-degrading bacteria for disposal of seaweed wastes. Lett. Appl. Microbiol. 2009, 48, 38–43. [Google Scholar] [CrossRef]
- Wang, M.P.; Chen, L.; Liu, Z.Y.; Zhang, Z.J.; Qin, S.; Yan, P.S. Isolation of a novel alginate lyase-producing Bacillus litoralis strain and its potential to ferment Sargassum horneri for biofertilizer. Microbiologyopen 2016, 5, 1038–1049. [Google Scholar] [CrossRef] [Green Version]
- An, Q.D.; Zhang, G.L.; Wu, H.T.; Zhang, Z.C.; Zheng, G.S.; Luan, L.; Murata, Y.; Li, X. Alginate-deriving oligosaccharide production by alginase from newly isolated Flavobacterium sp. LXA and its potential application in protection against pathogens. J. Appl. Microbiol. 2009, 106, 161–170. [Google Scholar] [CrossRef]
- Ito, M.; Watanabe, K.; Maruyama, T.; Mori, T.; Niwa, K.; Chow, S.; Takeyama, H. Enrichment of bacteria and alginate lyase genes potentially involved in brown alga degradation in the gut of marine gastropods. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Erasmus, J.H.; Cook, P.A.; Coyne, V.E. The role of bacteria in the digestion of seaweed by the abalone Haliotis midae. Aquaculture 1997, 155, 377–386. [Google Scholar] [CrossRef]
- Bansemer, M.S.; Qin, J.G.; Harris, J.O.; Howarth, G.S.; Stone, D.A.J. Nutritional requirements and use of macroalgae as ingredients in abalone feed. Rev. Aquac. 2016, 8, 121–135. [Google Scholar] [CrossRef]
- Tanaka, R.; Shibata, T.; Miyake, H.; Mori, T.; Tamaru, Y.; Ueda, M.; Bossier, P. Temporal fluctuation in the abundance of alginate-degrading bacteria in the gut of abalone Haliotis gigantea over 1 year. Aquac. Res. 2016, 47, 2899–2908. [Google Scholar] [CrossRef]
- Kurahashi, M.; Yokota, A. Agarivorans albus gen. nov., sp. nov., a gamma-proteobacterium isolated from marine animals. Int. J. Syst. Evol. Microbiol. 2004, 54, 693–697. [Google Scholar] [CrossRef]
- Long, M.X.; Yu, Z.N.; Xu, X. A novel beta-agarase with high ph stability from marine Agarivorans sp. LQ48. Mar. Biotechnol. 2010, 12, 62–69. [Google Scholar] [CrossRef]
- Du, Z.J.; Wang, J.; Yang, L.J.; Chen, G.J. Identification of a marine agarolytic bacterium Agarivorans albus QM38 and cloning and sequencing its beta-agarase genes. Acta Oceanol. Sin. 2011, 30, 118–124. [Google Scholar] [CrossRef]
- Lee, D.G.; Jeon, M.J.; Lee, S.H. Cloning, expression, and characterization of a glycoside hydrolase family 118 beta-agarase from Agarivorans sp. JA-1. J. Microbiol. Biotechnol. 2012, 22, 1692–1697. [Google Scholar] [CrossRef]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Li, S.Y.; Yang, X.M.; Bao, M.M.; Wu, Y.; Yu, W.G.; Han, F. Family 13 carbohydrate-binding module of alginate lyase from Agarivorans sp. L11 enhances its catalytic efficiency and thermostability, and alters its substrate preference and product distribution. Fems Microbiol. Lett. 2015, 362, fnv054. [Google Scholar] [CrossRef] [Green Version]
- Li, S.Y.; Yang, X.M.; Zhang, L.; Yu, W.G.; Han, F. Cloning, expression, and characterization of a cold-adapted and surfactant-stable alginate lyase from marine bacterium Agarivorans sp. L11. J. Microbiol. Biotechnol. 2015, 25, 681–686. [Google Scholar] [CrossRef] [PubMed]
- Uchimura, K.; Miyazaki, M.; Nogi, Y.; Kobayashi, T.; Horikoshi, K. Cloning and sequencing of alginate lyase genes from deep-sea strains of Vibrio and Agarivorans and characterization of a new Vibrio enzyme. Mar. Biotechnol. 2010, 12, 526–533. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, T.; Uchimura, K.; Miyazaki, M.; Nogi, Y.; Horikoshi, K. A new high-alkaline alginate lyase from a deep-sea bacterium Agarivorans sp. Extremophiles 2009, 13, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Hisano, T.; Nishimura, M.; Yamashita, T.; Imanaka, T.; Muramatsu, T.; Kimura, A.; Murata, K. A simple method for determination of substrate-specificity of alginate lyases. J. Ferment. Bioeng. 1994, 78, 182–184. [Google Scholar] [CrossRef]
- Al-Saari, N.; Gao, F.; Rohul, A.; Sato, K.; Sato, K.; Mino, S.; Suda, W.; Oshima, K.; Hattori, M.; Ohkuma, M.; et al. Advanced microbial taxonomy combined with genome-based-approaches reveals that Vibrio astriarenae sp. nov., an agarolytic marine bacterium, forms a new clade in Vibrionaceae. PLoS ONE 2015, 10, e0136279. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.P.; Jin, X.K.; Wu, C.; Zhu, X.Y.; Liu, M.; Call, D.R.; Zhao, Z. Genome-wide identification and functional characterization of β-agarases in Vibrio astriarenae strain HN897. Front. Microbiol. 2020, 11, 1404. [Google Scholar] [CrossRef]
- El Rayes, J.; Rodriguez-Alonso, R.; Collet, J.F. Lipoproteins in Gram-negative bacteria: New insights into their biogenesis, subcellular targeting and functional roles. Curr. Opin. Microbiol. 2021, 61, 25–34. [Google Scholar] [CrossRef]
- Srisodsuk, M.; Reinikainen, T.; Penttila, M.; Teeri, T.T. Role of the interdomain linker peptide of trichoderma-reesei cellobiohydrolase-i in its interaction with crystalline cellulose. J. Biol. Chem. 1993, 268, 20756–20761. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, T.R.; Long, L.K.; Ding, S.J. Altering the linker in processive GH5 endoglucanase 1 modulates lignin binding and catalytic properties. Biotechnol. Biofuels 2018, 11, 1–11. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.M.; Lim, J.; Kwon, S.; Chun, J. A large-scale evaluation of algorithms to calculate average nucleotide identity. Antonie Van Leeuwenhoek 2017, 110, 1281–1286. [Google Scholar] [CrossRef]
- Meier-Kolthoff, J.P.; Carbasse, J.S.; Peinado-Olarte, R.L.; Goker, M. TYGS and LPSN: A database tandem for fast and reliable genome-based classification and nomenclature of prokaryotes. Nucleic Acids Res. 2022, 50, D801–D807. [Google Scholar] [CrossRef]
- Smibert, R.; Krieg, N. Phenotypic characterization. In Methods for General and Molecular Bacteriology; Gerhardt, P., Murray, R.G.E., Wood, W.A., Krieg, N.R., Eds.; American Society For Microbiology: Washington, DC, USA, 1994; pp. 611–651. [Google Scholar]
- Bowman, J.P. Description of Cellulophaga algicola sp. nov., isolated from the surfaces of Antarctic algae, and reclassification of Cytophaga uliginosa (ZoBell and Upham 1944) Reichenbach 1989 as Cellulophaga uliginosa comb. nov. Int. J. Syst. Evol. Microbiol. 2000, 50, 1861–1868. [Google Scholar] [CrossRef] [Green Version]
- Dong, X.-Z.; Cai, M.-Y. Determinative Manual for Routine Bacteriology; Scientific Press: Beijing, China, 2001. [Google Scholar]
- Edgar, R.C.; Soc, I.C. MUSCLE: Multiple sequence alignment with improved accuracy and speed. In Proceedings of the IEEE Computational Systems Bioinformatics Conference (CSB 2004), Stanford, CA, USA, 16 19 August 2004; pp. 728–729. [Google Scholar]
- Capella-Gutierrez, S.; Silla-Martinez, J.M.; Gabaldon, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Trifinopoulos, J.; Nguyen, L.T.; von Haeseler, A.; Minh, B.Q. W-IQ-TREE: A fast online phylogenetic tool for maximum likelihood analysis. Nucleic Acids Res. 2016, 44, W232–W235. [Google Scholar] [CrossRef] [Green Version]
- Letunic, I.; Bork, P. Interactive Tree Of Life v2: Online annotation and display of phylogenetic trees made easy. Nucleic Acids Res. 2011, 39, W475–W478. [Google Scholar] [CrossRef]
- Yoon, S.H.; Ha, S.M.; Kwon, S.; Lim, J.; Kim, Y.; Seo, H.; Chun, J. Introducing EzBioCloud: A taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 2017, 67, 1613–1617. [Google Scholar] [CrossRef]
- Thompson, J.D.; Gibson, T.J.; Plewniak, F.; Jeanmougin, F.; Higgins, D.G. The CLUSTAL_X windows interface: Flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res. 1997, 25, 4876–4882. [Google Scholar] [CrossRef] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Fitch, W.M. Toward defining the course of evolution-minimum change for a specific tree topology. Syst. Zool. 1971, 20, 406–416. [Google Scholar] [CrossRef]
- Felsenstein, J. Evolutionary trees from DNA sequences: A maximum likelihood approach. J. Mol. Evol. 1981, 17, 368–376. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- Kimura, M. A simple method for estimating evolutionary rates of base substitutions through comparative studies of nucleotide-sequences. J. Mol. Evol. 1980, 16, 111–120. [Google Scholar] [CrossRef]
- Zhang, H.; Yohe, T.; Huang, L.; Entwistle, S.; Wu, P.Z.; Yang, Z.L.; Busk, P.K.; Xu, Y.; Yin, Y.B. dbCAN2: A meta server for automated carbohydrate-active enzyme annotation. Nucleic Acids Res. 2018, 46, W95–W101. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.B.; Mao, X.Z.; Yang, J.C.; Chen, X.; Mao, F.L.; Xu, Y. dbCAN: A web resource for automated carbohydrate-active enzyme annotation. Nucleic Acids Res. 2012, 40, W445–W451. [Google Scholar] [CrossRef]
- Yang, J.Y.; Yan, R.X.; Roy, A.; Xu, D.; Poisson, J.; Zhang, Y. The I-TASSER Suite: Protein structure and function prediction. Nat. Methods 2015, 12, 7–8. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Skolnick, J. TM-align: A protein structure alignment algorithm based on the TM-score. Nucleic Acids Res. 2005, 33, 2302–2309. [Google Scholar] [CrossRef]
- Juncker, A.S.; Willenbrock, H.; Von Heijne, G.; Brunak, S.; Nielsen, H.; Krogh, A. Prediction of lipoprotein signal peptides in Gram-negative bacteria. Protein Sci. 2003, 12, 1652–1662. [Google Scholar] [CrossRef]
- Savojardo, C.; Martelli, P.L.; Fariselli, P.; Profiti, G.; Casadio, R. BUSCA: An integrative web server to predict subcellular localization of proteins. Nucleic Acids Res. 2018, 46, W459–W466. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Locus | Protein Name | Length (aa) | Mw (kDa) | pI | Instability Index | GRAVY | Most Similar Sequence | PL Family | |
|---|---|---|---|---|---|---|---|---|---|
| Sequence ID (%) | Organism | ||||||||
| 821 | Aly7A1 | 327 | 36.7 | 4.66 | 30.51 | −0.420 | WP_164648089 (55.38) | Vibrio astriarenae | PL7 |
| 2288 | Aly7A2 | 346 | 38.3 | 5.93 | 19.83 | −0.522 | WP_221076907 (98.55) | Agarivorans aestuarii | PL7 |
| 2290 | Aly7A3 | 286 | 32.2 | 6.53 | 25.97 | −0.535 | WP_040307007 (98.60) | Agarivorans albus | PL7 |
| 2857 | Aly7A4 | 327 | 36.8 | 4.63 | 29.92 | −0.422 | WP_164648089 (54.75) | Vibrio astriarenae | PL7 |
| 2931 | Aly7A5 | 309 | 34.4 | 5.93 | 23.80 | −0.570 | WP_220718128 (98.71) | Agarivorans litoreus | PL7 |
| 1488 | Aly7B1 | 524 | 57.5 | 4.99 | 35.73 | −0.469 | WP_016400966 (99.81) | Agarivorans albus | PL7 |
| 3274 | Aly7B2 | 487 | 53.1 | 4.26 | 37.40 | −0.560 | WP_221074174 (97.95) | Agarivorans aestuarii | PL7 |
| 1799 | Aly7C | 486 | 52.4 | 5.26 | 33.12 | −0.573 | WP_221077217 (99.43) | Agarivorans aestuarii | PL7 |
| 1826 | Aly6A | 892 | 95.1 | 4.16 | 31.75 | −0.291 | WP_221076585 (98.43) | Agarivorans aestuarii | PL6 |
| 2253 | Aly38A | 415 | 46.9 | 6.99 | 40.25 | −0.578 | WP_016404079 (99.04) | Agarivorans albus | PL38 |
| 2231 | Oal17A1 | 715 | 80.0 | 5.45 | 41.05 | −0.302 | WP_016403996 (99.58) | Agarivorans albus | PL17 |
| 2232 | Oal17A2 | 712 | 80.1 | 5.40 | 35.63 | −0.388 | WP_016403995 (99.72) | Agarivorans albus | PL17 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, X.-K.; Gong, Y.; Shang, D.-D.; Liu, B.-T.; Du, Z.-J.; Chen, G.-J. Degradation of Alginate by a Newly Isolated Marine Bacterium Agarivorans sp. B2Z047. Mar. Drugs 2022, 20, 254. https://doi.org/10.3390/md20040254
Sun X-K, Gong Y, Shang D-D, Liu B-T, Du Z-J, Chen G-J. Degradation of Alginate by a Newly Isolated Marine Bacterium Agarivorans sp. B2Z047. Marine Drugs. 2022; 20(4):254. https://doi.org/10.3390/md20040254
Chicago/Turabian StyleSun, Xun-Ke, Ya Gong, Dan-Dan Shang, Bang-Tao Liu, Zong-Jun Du, and Guan-Jun Chen. 2022. "Degradation of Alginate by a Newly Isolated Marine Bacterium Agarivorans sp. B2Z047" Marine Drugs 20, no. 4: 254. https://doi.org/10.3390/md20040254
APA StyleSun, X. -K., Gong, Y., Shang, D. -D., Liu, B. -T., Du, Z. -J., & Chen, G. -J. (2022). Degradation of Alginate by a Newly Isolated Marine Bacterium Agarivorans sp. B2Z047. Marine Drugs, 20(4), 254. https://doi.org/10.3390/md20040254