
Antioxidative Effect of Chlorella Pyrenoidosa Protein Hydrolysates and Their Application in Krill Oil-in-Water Emulsions

 , and
, and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
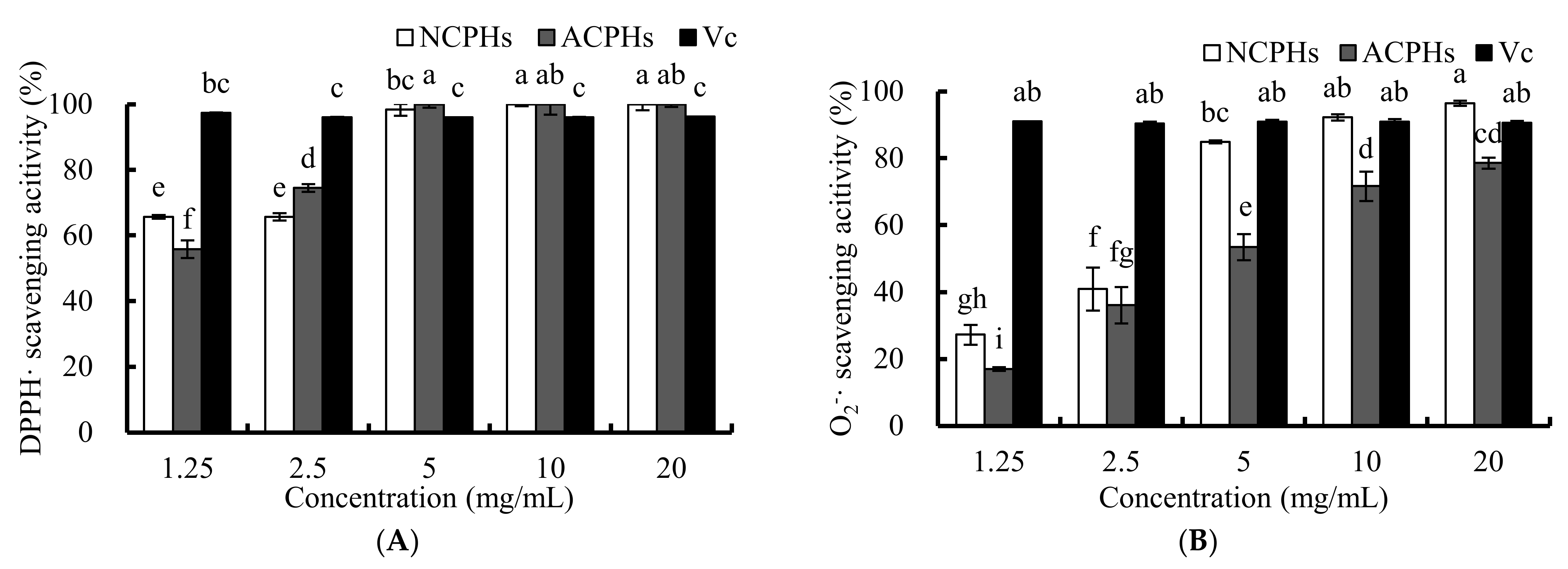
2.1. Antioxidant Activities of CPHs
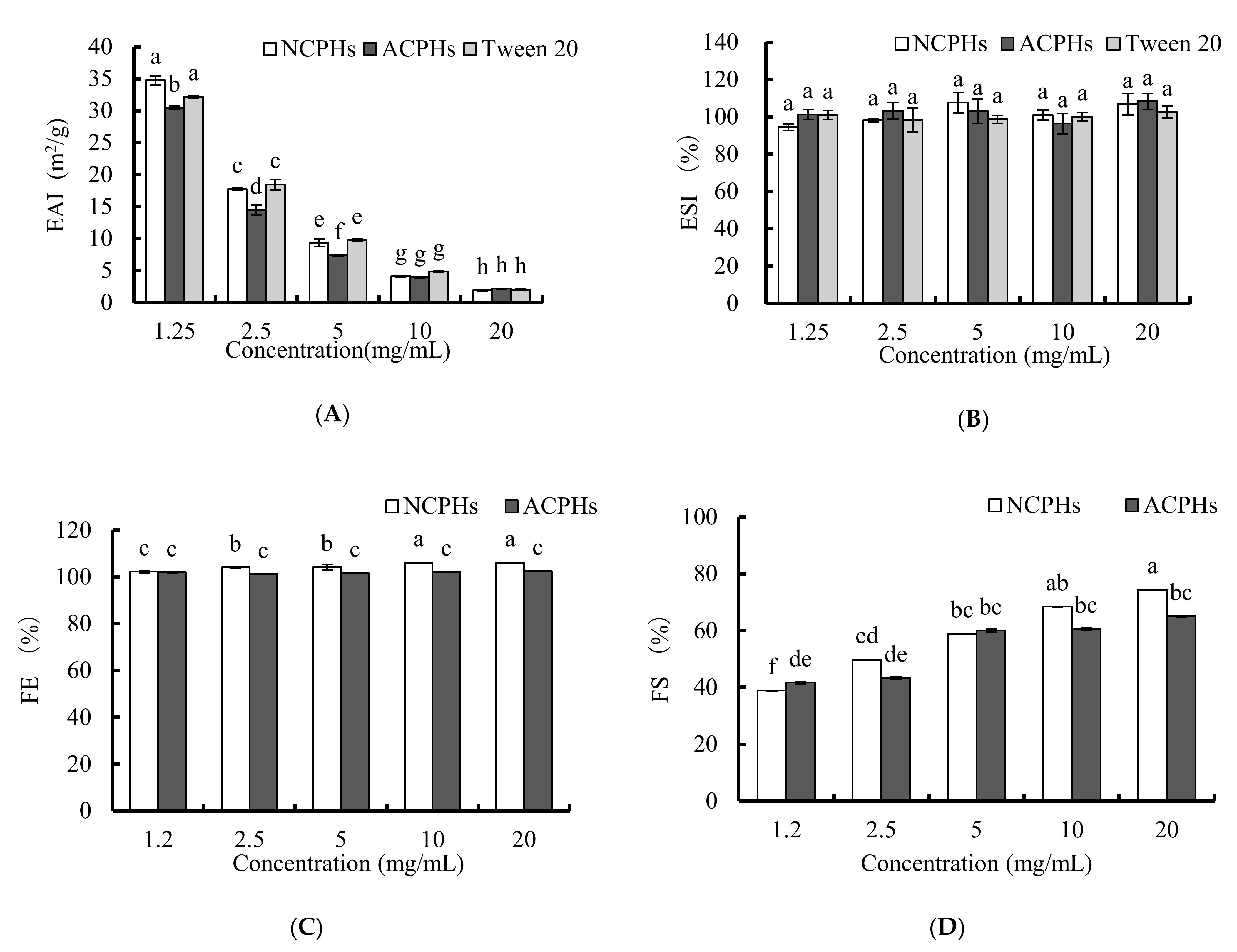
2.2. Emulsifying and Foaming Activities of CPHs
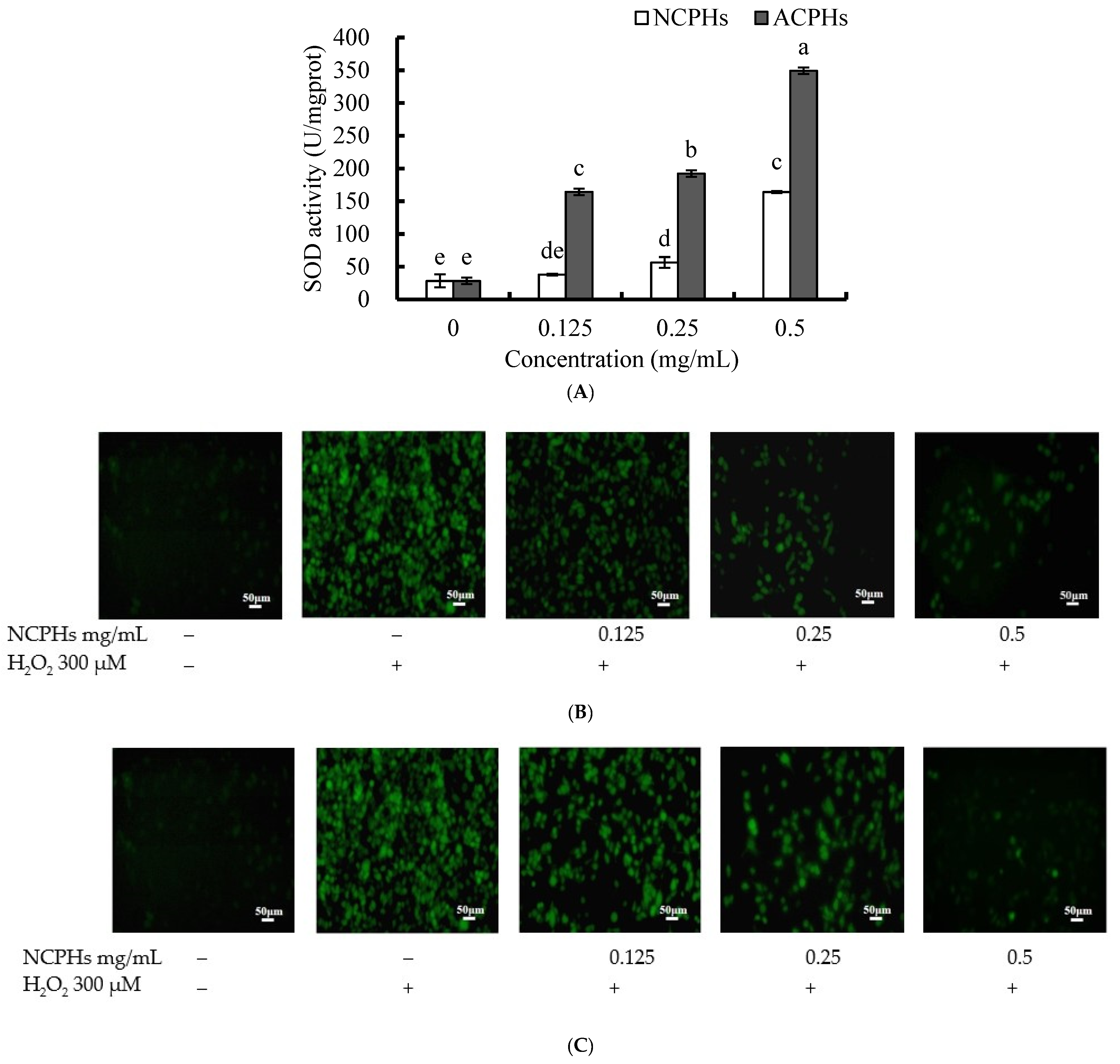
2.3. Intracellular Antioxidant Activity of CPHs In Vitro
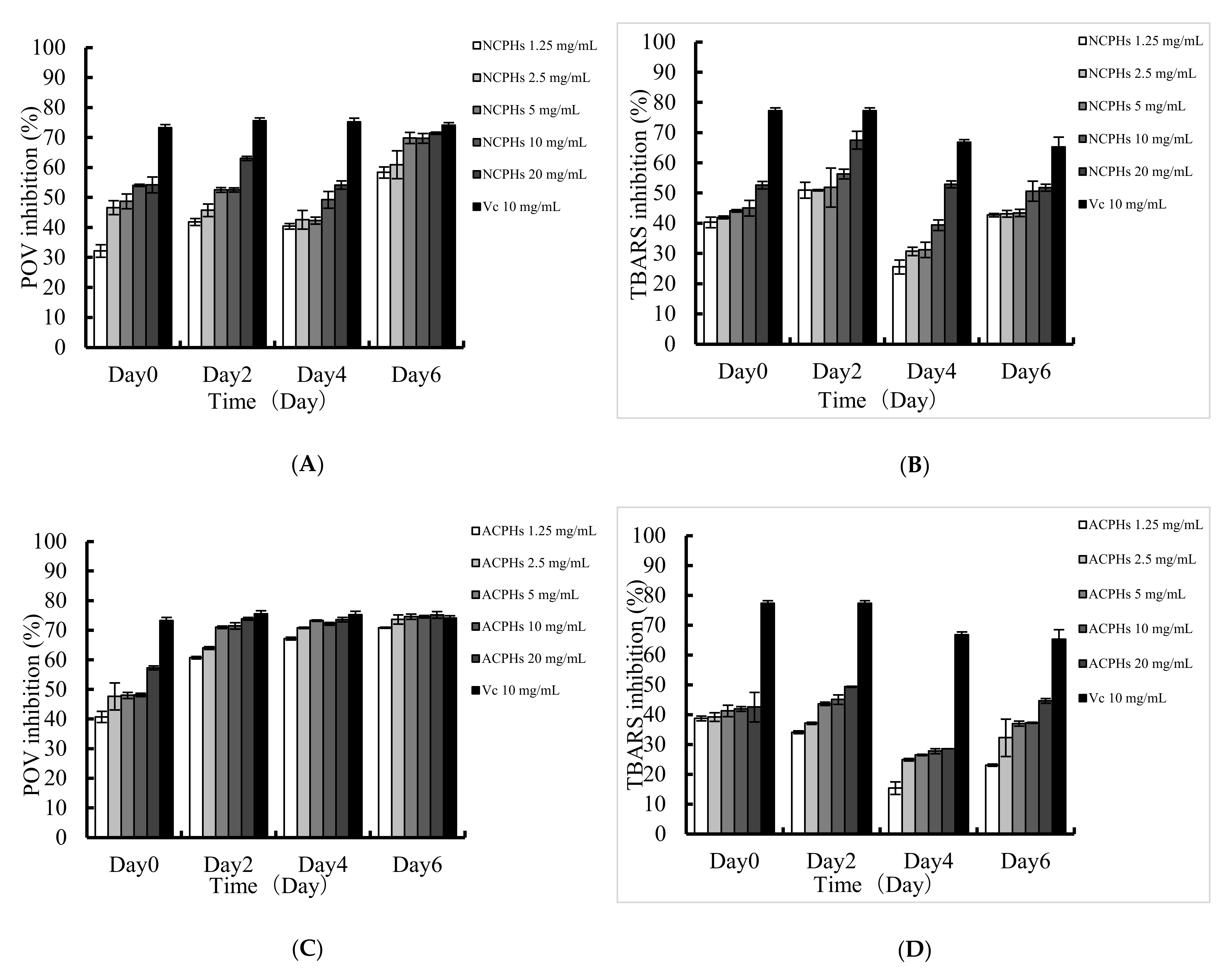
2.4. Lipid Peroxidation Inhibition Assay
2.5. CPH-Loaded Antarctic Krill Oil Emulsion
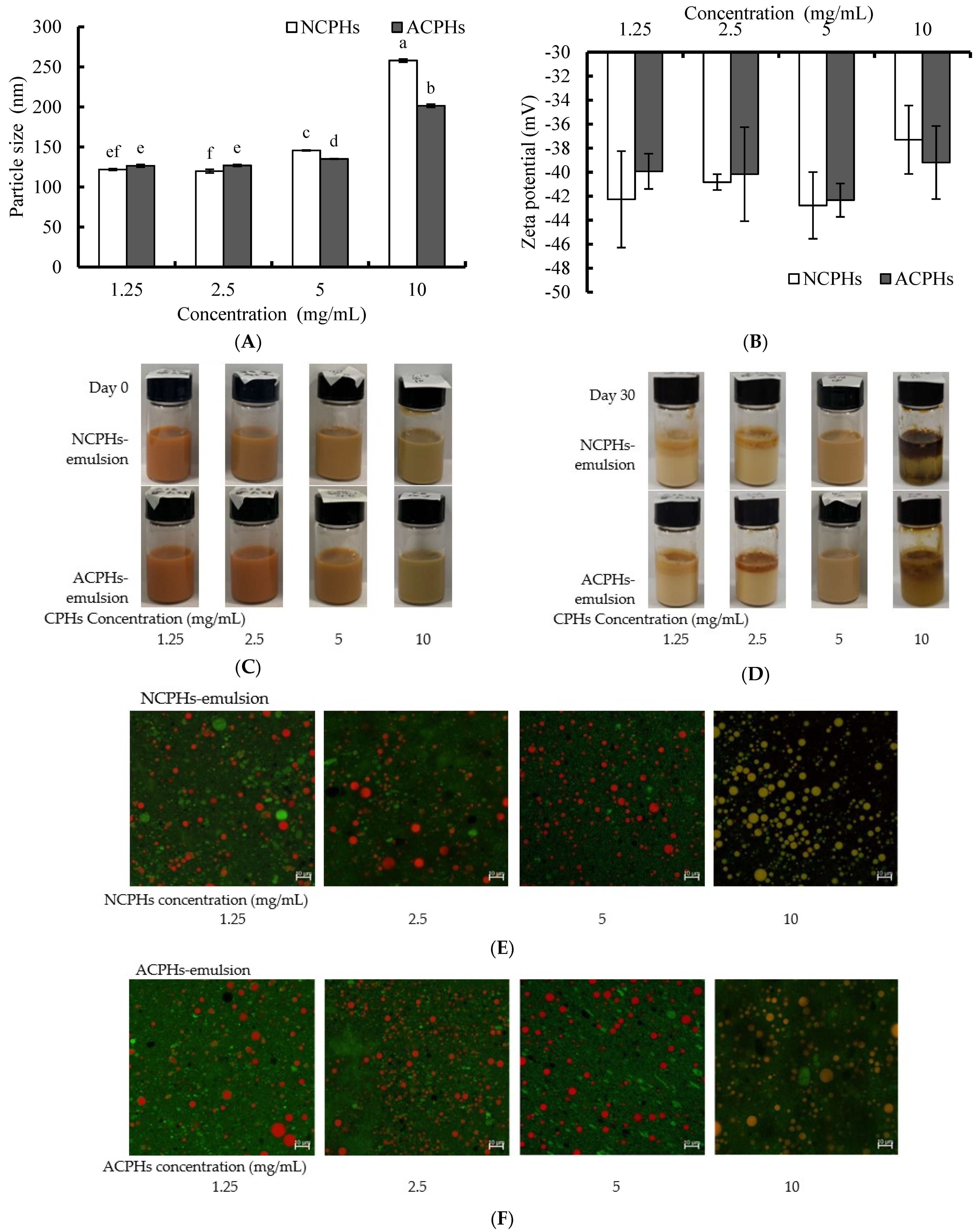
2.5.1. Particle Size and Zeta Potential of CPH-Loaded Antarctic Krill Oil Emulsions
2.5.2. Morphology of CPH-Loaded KO Emulsions
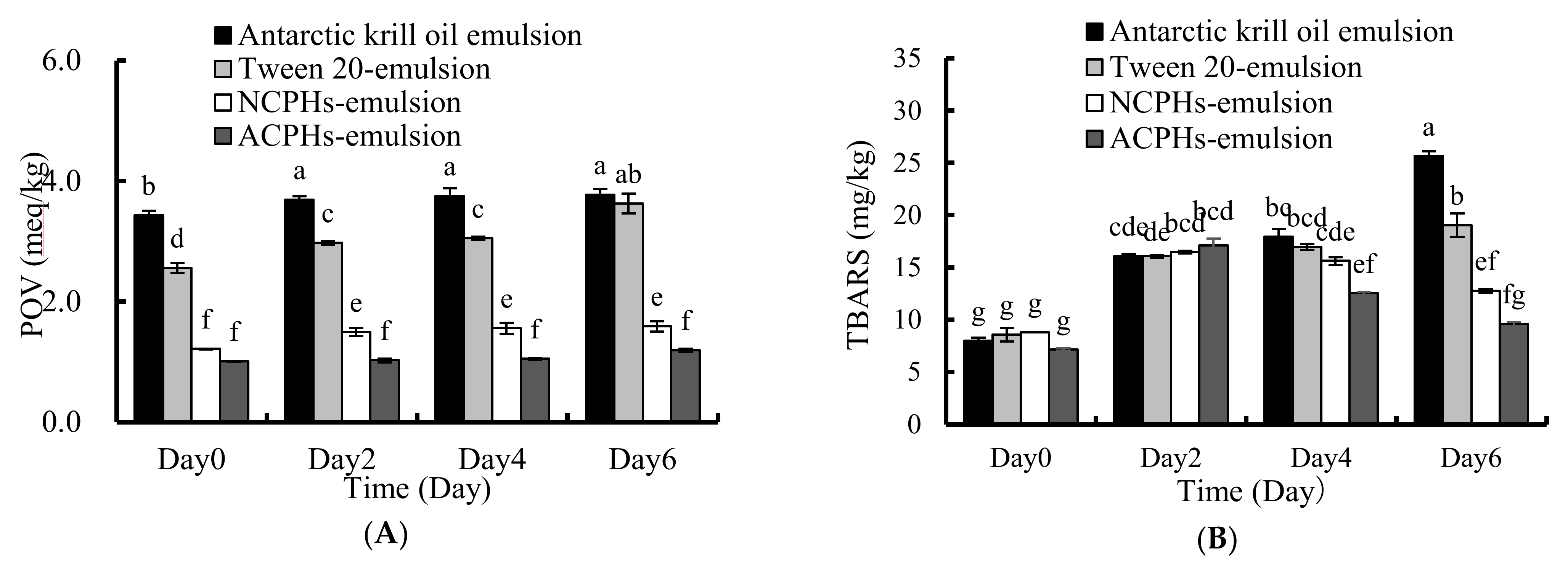
2.6. Oxidative Stability of the Emulsion
3. Materials and Methods
3.1. Chemicals
3.2. Hydrolysates from Chlorella Protein
3.3. Determination of the Antioxidant Activity of Chlorella Protein Hydrolysates (CPHs)
3.3.1. DPPH Radical Scavenging Activity Assay
3.3.2. Hydroxyl Radical (HO) Scavenging Activity
3.3.3. Superoxide Anion Radical (O2−) Scavenging Activity
3.3.4. Determination of ABTS Scavenging Activity
3.4. Emulsifying Properties
3.5. Foaming Properties
3.6. Activity of SOD
3.7. Reactive Oxygen Species (ROS) Level Determination
3.8. Lipid Peroxidation Inhibitory Activity Assay of CPHs
3.9. Emulsion Preparation
3.10. Particle Size and Zeta Potential Measurements
3.11. Confocal Laser Scanning Microscopy (CLSM)
3.12. Oxidative Stability of CPH Emulsions
3.13. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Antarctic krill oil | KO |
| 2,2′-azinobis (3-ethylbenzothiazoline-6-sulphonic acid) | ABTS |
| Chlorella pyrenoidosa | C. pyrenoidosa |
| Chlorella protein hydrolysate | CPHs |
| oil-in-water emulsions | O/W |
| 1,1-diphenyl-2-picrylhydrazyl | DPPH |
| peroxide value | POV |
| eicosapentaenoic acid | EPA |
| docosahexaenoic acid | DHA |
| Maillard reaction products | MRP |
| thiobarbituric acid | TBA |
| reactive oxygen species | ROS |
| superoxide dismutase | SOD |
References
- Kohler, A.; Sarkkinen, E.; Tapola, N.; Niskanen, T.; Bruheim, I. Bioavailability of fatty acids from krill oil, krill meal and fish oil in healthy subjects--a randomized, single-dose, cross-over trial. Lipids Health Dis. 2015, 14, 19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tou, J.C.; Jaczynski, J.; Chen, Y.-C. Krill for human consumption: Nutritional value and potential health benefits. Nutr. Rev. 2007, 65, 63–77. [Google Scholar] [CrossRef] [PubMed]
- Wijendran, V.; Huang, M.-C.; Diau, G.-Y.; Boehm, G.; Nathanielsz, P.W.; Brenna, J.T. Efficacy of dietary arachidonic acid provided as triglyceride or phospholipid as substrates for brain arachidonic acid accretion in baboon neonates. Pediatric Res. 2002, 51, 265–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, Z.; Bhail, S.; Sanguansri, L.; Augustin, M.A. Improving the Oxidative Stability of Krill Oil-in-Water Emulsions. J. Am. Oil Chem. Soc. 2014, 91, 1347–1354. [Google Scholar] [CrossRef]
- Schröder, A.; Berton-Carabin, C.; Venema, P.; Cornacchia, L. Interfacial properties of whey protein and whey protein hydrolysates and their influence on O/W emulsion stability. Food Hydrocoll. 2017, 73, 129–140. [Google Scholar] [CrossRef]
- Liu, H.; Li, Y.; Diao, X.; Kong, B.; Liu, Q. Effect of porcine bone protein hydrolysates on the emulsifying and oxidative stability of oil-in-water emulsions. Colloids Surf. A Physicochem. Eng. Asp. 2018, 538, 757–764. [Google Scholar] [CrossRef]
- Zhao, J.; Xiong, Y.L. Interfacial peptide partitioning and undiminished antioxidative and emulsifying activity of oxidatively stressed soy protein hydrolysate in an O/W emulsion. LWT-Food Sci. Technol. 2015, 61, 322–329. [Google Scholar] [CrossRef]
- Zhao, Q.; Selomulya, C.; Wang, S.; Xiong, H.; Chen, X.D.; Li, W.; Peng, H.; Xie, J.; Sun, W.; Zhou, Q. Enhancing the oxidative stability of food emulsions with rice dreg protein hydrolysate. Food Res. Int. 2012, 48, 876–884. [Google Scholar] [CrossRef]
- Shastik, E.; Li, L.; Liu, J. New methods for hydrogen production by marine microalga Chlorella pyrenoidosa in natural seawater. Int. J. Hydrogen Energy 2019, 44, 14707–14714. [Google Scholar] [CrossRef]
- Singhal, R.; Basu, H.; Pimple, M.; Manisha, V.; Basan, M.; Reddy, A.V.R. Spectroscopic determination of U (VI) species sorbed by the Chlorella (Chlorella pyrenoidosa) fresh water algae. J. Radioanal. Nucl. Chem. 2013, 298, 587–592. [Google Scholar] [CrossRef]
- He, M.; Li, L.; Liu, J.; Zhang, L. Improvement of H2 photoproduction in Chlorella pyrenoidosa in artificial and natural seawater by addition of acetic acid and control of nutrients. Algal Res. 2015, 10, 104–109. [Google Scholar] [CrossRef]
- Waghmare, A.G.; Salve, M.K.; LeBlanc, J.G.; Arya, S.S. Concentration and characterization of microalgae proteins from Chlorella pyrenoidosa. Bioresour. Bioprocess. 2016, 3, 16. [Google Scholar] [CrossRef] [Green Version]
- Cherng, J.; Liu, C.; Shen, C.; Lin, H.; Shih, M. Beneficial effects of Chlorella-11 peptide on blocking LPS-induced macrophage activation and alleviating thermal injury-induced inflammation in rats. Int. J. Immunopathol. Pharmacol. 2010, 23, 811–820. [Google Scholar] [CrossRef]
- Yamaguchi, N. Studies on Antioxidative Activities of Amino Compounds on Fats and Oils Part, I. Oxidation of methionine during course of autoxidation of linoleic acid. J. Jpn. Soc. Food Sci. Technol. 1971, 18, 313–318. [Google Scholar] [CrossRef]
- Chen, Y.; Huang, F.; Xie, B.; Sun, Z.; McClements, D.J.; Deng, Q. Fabrication and characterization of whey protein isolates- lotus seedpod proanthocyanin conjugate: Its potential application in oxidizable emulsions. Food Chem. 2021, 346, 128680. [Google Scholar] [CrossRef]
- Linke, A.; Hinrichs, J.; Kohlus, R. Impact of the powder particle size on the oxidative stability of microencapsulated oil. Powder Technol. 2020, 364, 115–122. [Google Scholar] [CrossRef]
- Sivaraman, B.; Shakila, R.J.; Jeyasekaran, G.; Sukumar, D.; Manimaran, U.; Sumathi, G. Antioxidant activities of squid protein hydrolysates prepared with papain using response surface methodology. Food Sci. Biotechnol. 2016, 25, 665–672. [Google Scholar] [CrossRef]
- Nalinanon, S.; Benjakul, S.; Kishimura, H.; Shahidi, F. Functionalities and antioxidant properties of protein hydrolysates from the muscle of ornate threadfin bream treated with pepsin from skipjack tuna. Food Chem. 2011, 124, 1354–1362. [Google Scholar] [CrossRef]
- Wang, Z.; Liu, X.; Xie, H.; Liu, Z.; Rakariyatham, K.; Yu, C.; Shahidi, F.; Zhou, D. Antioxidant activity and functional properties of Alcalase-hydrolyzed scallop protein hydrolysate and its role in the inhibition of cytotoxicity in vitro. Food Chem. 2021, 344, 128566. [Google Scholar] [CrossRef]
- Lawal, O.S. Functionality of African locust bean (Parkia biglobossa) protein isolate: Effects of pH, ionic strength and various protein concentrations. Food Chem. 2004, 86, 345–355. [Google Scholar] [CrossRef]
- Wouters, A.G.B.; Rombouts, I.; Fierens, E.; Brijs, K.; Delcour, J.A. Relevance of the Functional Properties of Enzymatic Plant Protein Hydrolysates in Food Systems. Compr. Rev. Food Sci. Food Saf. 2016, 15, 786–800. [Google Scholar] [CrossRef] [Green Version]
- Bowler, C.; Montagu, M.V.; Inze, D. Superoxide dismutase and stress tolerance. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1992, 43, 83–116. [Google Scholar] [CrossRef]
- Hu, X.M.; Wang, Y.M.; Zhao, Y.Q.; Chi, C.F.; Wang, B. Antioxidant Peptides from the Protein Hydrolysate of Monkfish (Lophius litulon) Muscle: Purification, Identification, and Cytoprotective Function on HepG2 Cells Damage by H2O2. Mar. Drugs 2020, 18, 153. [Google Scholar] [CrossRef] [Green Version]
- Wu, R.; Wu, C.; Liu, D.; Yang, X.; Huang, J.; Zhang, J.; Liao, B.; He, H. Antioxidant and anti-freezing peptides from salmon collagen hydrolysate prepared by bacterial extracellular protease. Food Chem. 2018, 248, 346–352. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.-C.; Kim, D.; Jeon, Y.-J. Protective effect of a novel antioxidative peptide purified from a marine Chlorella ellipsoidea protein against free radical-induced oxidative stress. Food Chem. Toxicol. 2012, 50, 2294–2302. [Google Scholar] [CrossRef] [PubMed]
- Sheih, I.C.; Fang, T.J.; Wu, T.K.; Lin, P.H. Anticancer and antioxidant activities of the peptide fraction from algae protein waste. J. Agric. Food Chem. 2010, 58, 1202–1207. [Google Scholar] [CrossRef] [PubMed]
- Kang, K.-H.; Qian, Z.-J.; Ryu, B.; Karadeniz, F.; Kim, D.; Kim, S.-K. Antioxidant Peptides from Protein Hydrolysate of Microalgae Navicula incerta and their Protective Effects in Hepg2/CYP2E1 Cells Induced by Ethanol. Phytother. Res. 2012, 26, 1555–1563. [Google Scholar] [CrossRef] [PubMed]
- Ejike, C.E.C.C.; Collins, S.A.; Balasuriya, N.; Swanson, A.K.; Mason, B.; Udenigwe, C.C. Prospects of microalgae proteins in producing peptide-based functional foods for promoting cardiovascular health. Trends Food Sci. Technol. 2017, 59, 30–36. [Google Scholar] [CrossRef]
- Meshginfar, N.; Sadeghi Mahoonak, A.; Ghorbani, M.; Aalami, M. Effects of Protein Hydrolysate from Sheep Visceral on Oxidative Stability of Soybean Oil and Chicken Sausage. J. Food Process. Preserv. 2017, 41, e12875. [Google Scholar] [CrossRef]
- Lam, R.S.; Nickerson, M.T. Food proteins: A review on their emulsifying properties using a structure-function approach. Food Chem. 2013, 141, 975–984. [Google Scholar] [CrossRef]
- Padial-Dominguez, M.; Espejo-Carpio, F.J.; Perez-Galvez, R.; Guadix, A.; Guadix, E.M. Optimization of the Emulsifying Properties of Food Protein Hydrolysates for the Production of Fish Oil-in-Water Emulsions. Foods 2020, 9, 636. [Google Scholar] [CrossRef]
- Faraji, H.; McClements, D.J.; Decker, E.A. Role of continuous phase protein on the oxidative stability of fish oil-in-water emulsions. J. Agric. Food Chem. 2004, 52, 4558–4564. [Google Scholar] [CrossRef]
- Wong, B.T.; Zhai, J.; Hoffmann, S.V.; Aguilar, M.-I.; Augustin, M.; Wooster, T.J.; Day, L. Conformational changes to deamidated wheat gliadins and β-casein upon adsorption to oil–water emulsion interfaces. Food Hydrocoll. 2012, 27, 91–101. [Google Scholar] [CrossRef]
- Shimada, K.; Fujikawa, K.; Yahara, K.; Nakamura, T. Antioxidative properties of xanthan on the autoxidation of soybean oil in cyclodextrin emulsion. J. Agric. Food Chem. 1992, 40, 945–948. [Google Scholar] [CrossRef]
- Liu, J.; Wen, X.Y.; Zhang, X.Q.; Pu, H.M.; Kan, J.; Jin, C.H. Extraction, characterization and in vitro antioxidant activity of polysaccharides from black soybean. Int. J. Biol. Macromol. 2015, 72, 1182–1190. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.; Aruoma, O.I. The deoxyribose method: A simple “test-tube” assay for determination of rate constants for reactions of hydroxyl radicals. Anal. Biochem. 1987, 165, 215–219. [Google Scholar] [CrossRef]
- Pearce, K.N.; Kinsella, J.E. Emulsifying properties of proteins: Evaluation of a turbidimetric technique. J. Agric. Food Chem. 1978, 26, 716–723. [Google Scholar] [CrossRef]
- Shahidi, F.; Han, X.-Q.; Synowiecki, J. Production and characteristics of protein hydrolysates from capelin (Mallotus villosus). Food Chem. 1995, 53, 285–293. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- GB 5009.227-2016; The Chinese Food Safety Standards, Standards for Determination of Peroxide Value in Food. 2016.
- Kavosi, M.; Mohammadi, A.; Shojaee-Aliabadi, S.; Khaksar, R.; Hosseini, S.M. Agriculture, Characterization and oxidative stability of purslane seed oil microencapsulated in yeast cells biocapsules. J. Sci. Food Agric. 2018, 98, 2490–2497. [Google Scholar] [CrossRef]
- Undeland, I.; Hultin, H.O.; Richards, M.P. Added triacylglycerols do not hasten hemoglobin-mediated lipid oxidation in washed minced cod muscle. J. Agric. Food Chem. 2002, 50, 6847. [Google Scholar] [CrossRef]
- GB 5009.181-2016; The Chinese Food Safety Standards, Standards for Determination of Malondialdehyde in Food. 2016.






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Qi, Y.; Wang, Q.; Yin, F.; Zhan, H.; Wang, H.; Liu, B.; Nakamura, Y.; Wang, J. Antioxidative Effect of Chlorella Pyrenoidosa Protein Hydrolysates and Their Application in Krill Oil-in-Water Emulsions. Mar. Drugs 2022, 20, 345. https://doi.org/10.3390/md20060345
Liu Y, Qi Y, Wang Q, Yin F, Zhan H, Wang H, Liu B, Nakamura Y, Wang J. Antioxidative Effect of Chlorella Pyrenoidosa Protein Hydrolysates and Their Application in Krill Oil-in-Water Emulsions. Marine Drugs. 2022; 20(6):345. https://doi.org/10.3390/md20060345
Chicago/Turabian StyleLiu, Yujia, Yuli Qi, Qi Wang, Fawen Yin, Honglei Zhan, Han Wang, Bingnan Liu, Yoshimasa Nakamura, and Jihui Wang. 2022. "Antioxidative Effect of Chlorella Pyrenoidosa Protein Hydrolysates and Their Application in Krill Oil-in-Water Emulsions" Marine Drugs 20, no. 6: 345. https://doi.org/10.3390/md20060345
APA StyleLiu, Y., Qi, Y., Wang, Q., Yin, F., Zhan, H., Wang, H., Liu, B., Nakamura, Y., & Wang, J. (2022). Antioxidative Effect of Chlorella Pyrenoidosa Protein Hydrolysates and Their Application in Krill Oil-in-Water Emulsions. Marine Drugs, 20(6), 345. https://doi.org/10.3390/md20060345