
Seaweed-Derived Polysaccharides Attenuate Heat Stress-Induced Splenic Oxidative Stress and Inflammatory Response via Regulating Nrf2 and NF-κB Signaling Pathways



 and
and
Abstract
:1. Introduction
2. Results
2.1. Composition Characterization of SDP
2.2. Histology, Apoptosis Analysis and Relative Weight of Spleen
2.3. Antioxidant Capacity
2.4. Cytokine Levels and Cytokine Gene Expression
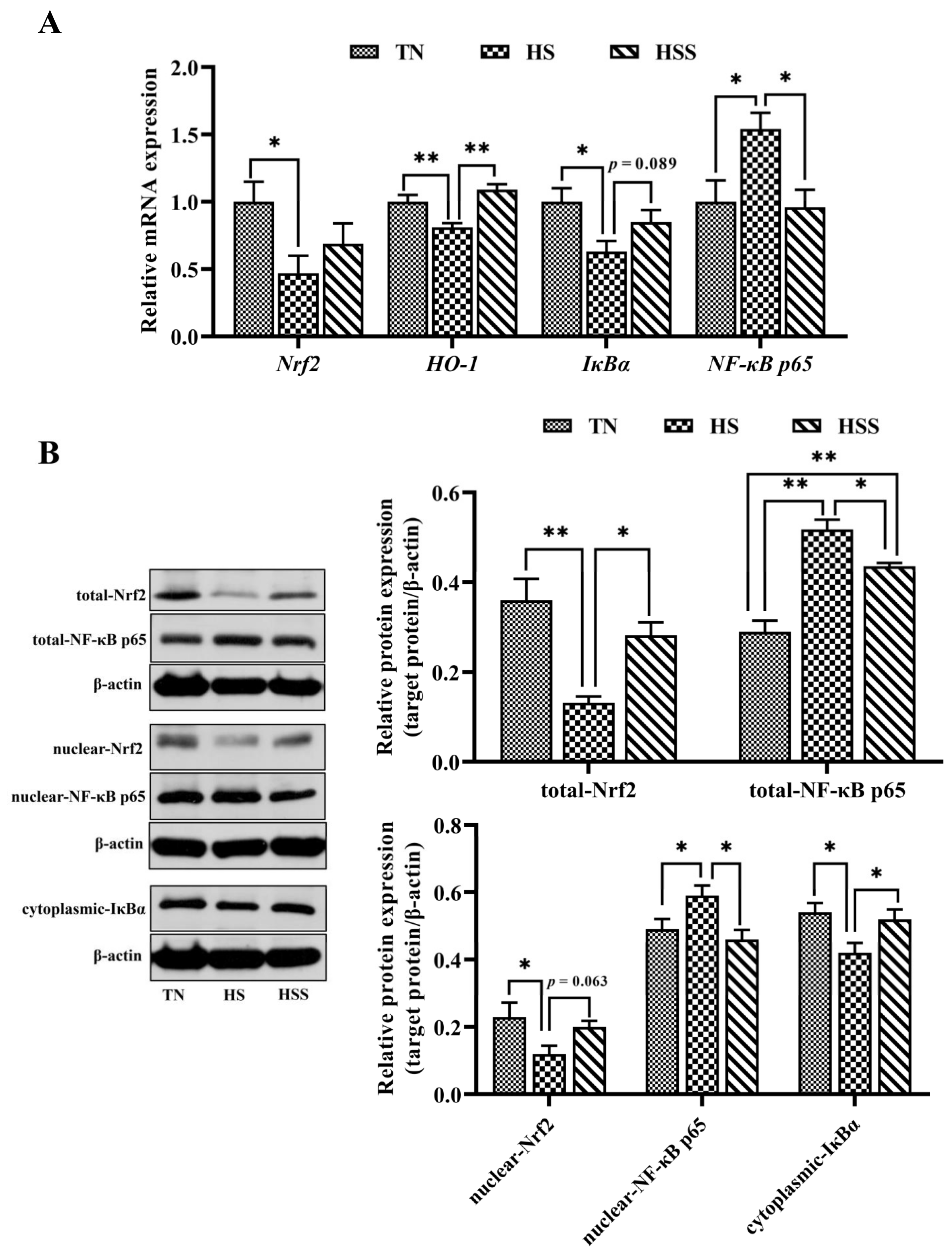
2.5. Expression of Nrf2 and NF-κB p65 Signaling Pathway–Related Molecules
3. Discussion
4. Materials and Methods
4.1. Preparation, Chemical Composition, and Structure Analysis of SDP
4.1.1. Preparation of SDP
4.1.2. Assay to Determine the Chemical Composition of SDP
4.1.3. Determination of the Molecular Weight of SDP
4.1.4. Assay to Determine the Monosaccharide Composition of SDP
4.1.5. Fourier-Transform Infrared (FT-IR) Spectroscopy
4.2. Animals, Diet, and Experimental Design
4.3. Sampling
4.4. Histology, Apoptosis and Relative Weight of Spleen
4.5. Determination of Antioxidant Parameters and Cytokine Concentrations
4.6. Quantitative Real-Time Polymerase Chain Reaction (qPCR) to Analyze Gene Expression
4.7. Western Blotting
4.8. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chauhan, S.S.; Rashamol, V.; Bagath, M.; Sejian, V.; Dunshea, F.R. Impacts of heat stress on immune responses and oxidative stress in farm animals and nutritional strategies for amelioration. Int. J. Biometeorol. 2021, 65, 1231–1244. [Google Scholar] [CrossRef]
- Liu, W.C.; Ou, B.H.; Liang, Z.L.; Zhang, R.; Zhao, Z.H. Algae-derived polysaccharides supplementation ameliorates heat stress-induced impairment of bursa of Fabricius via modulating NF-κB signaling pathway in broilers. Poult. Sci. 2021, 100, 101139. [Google Scholar] [CrossRef]
- Rhoads, R.P.; Baumgard, L.H.; Suagee, J.K.; Sanders, S.R. Nutritional interventions to alleviate the negative consequences of heat stress. Adv. Nutr. 2013, 4, 267–276. [Google Scholar] [CrossRef] [Green Version]
- Emami, N.K.; Jung, U.; Voy, B.; Dridi, S. Radical Response: Effects of heat stress-induced oxidative stress on lipid metabolism in the avian liver. Antioxidants 2021, 10, 35. [Google Scholar] [CrossRef]
- Hirakawa, R.; Nurjanah, S.; Furukawa, K.; Murai, A.; Kikusato, M.; Nochi, T.; Toyomizu, M. Heat stress causes immune abnormalities via massive damage to effect proliferation and differentiation of lymphocytes in broiler chickens. Front. Vet. Sci. 2020, 7, 46. [Google Scholar] [CrossRef]
- Guo, Y.; Balasubramanian, B.; Zhao, Z.H.; Liu, W.C. Heat stress alters serum lipid metabolism of Chinese indigenous broiler chickens-a lipidomics study. Environ. Sci. Pollut. Res. 2021, 28, 10707–10717. [Google Scholar] [CrossRef]
- Quinteiro-Filho, W.M.; Ribeiro, A.; Ferraz-de-Paula, V.; Pinheiro, M.; Sakai, M.; Sá, L.; Ferreira, A.; Palermo-Neto, J. Heat stress impairs performance parameters, induces intestinal injury, and decreases macrophage activity in broiler chickens. Poult. Sci. 2010, 89, 1905–1914. [Google Scholar] [CrossRef]
- Chegini, S.; Kiani, A.; Rokni, H. Alleviation of thermal and overcrowding stress in finishing broilers by dietary propolis supplementation. Ital. J. Anim. Sci. 2018, 17, 377–385. [Google Scholar] [CrossRef]
- Ohtsu, H.; Yamazaki, M.; Abe, H.; Murakam, H.; Toyomizu, M. Heat stress modulates cytokine gene expression in the spleen of broiler chickens. J. Poult. Sci. 2015, 52, 282–287. [Google Scholar] [CrossRef] [Green Version]
- He, S.; Yu, Q.; He, Y.; Hu, R.; Xia, S.; He, J. Dietary resveratrol supplementation inhibits heat stress-induced high-activated innate immunity and inflammatory response in spleen of yellow-feather broilers. Poult. Sci. 2019, 98, 6378–6387. [Google Scholar] [CrossRef] [PubMed]
- Sahin, K.; Orhan, C.; Tuzcu, M.; Sahin, N.; Hayirli, A.; Bilgili, S.; Kucuk, O. Lycopene activates antioxidant enzymes and nuclear transcription factor systems in heat-stressed broilers. Poult. Sci. 2016, 95, 1088–1095. [Google Scholar] [CrossRef]
- Awais, M.M.; Akhtar, M.; Anwar, M.I.; Khaliq, K. Evaluation of Saccharum officinarum L. bagasse-derived polysaccharides as native immunomodulatory and anticoccidial agents in broilers. Vet. Parasitol. 2018, 249, 74–81. [Google Scholar] [CrossRef]
- Wassie, T.; Duan, X.; Xie, C.; Wang, R.; Wu, X. Dietary Enteromorpha polysaccharide-Zn supplementation regulates amino acid and fatty acid metabolism by improving the antioxidant activity in chicken. J. Anim. Sci. Biotechnol. 2022, 13, 18. [Google Scholar] [CrossRef]
- Wassie, T.; Lu, Z.; Duan, X.; Xie, C.; Gebeyew, K.; Yumei, Z.; Wu, X. Dietary Enteromorpha polysaccharide enhances intestinal immune response, integrity, and caecal microbial activity of broiler chickens. Front. Nutr. 2021, 8, 783819. [Google Scholar] [CrossRef]
- Zhong, R.; Wan, X.; Wang, D.; Zhao, C.; Liu, D.; Gao, L.; Wang, M.; Wu, C.; Nabavid, S.M.; Daglia, M. Polysaccharides from marine Enteromorpha: Structure and function. Trends Food Sci. Technol. 2020, 99, 11–20. [Google Scholar] [CrossRef]
- Shi, M.J.; Wang, F.; Jiang, H.; Qian, W.W.; Xie, Y.Y.; Wei, X.Y.; Zhou, T. Effect of enzymatic degraded polysaccharides from Enteromorpha prolifera on the physical and oxidative stability of fish oil-in-water emulsions. Food Chem. 2020, 322, 126774. [Google Scholar] [CrossRef]
- Shi, M.J.; Wei, X.; Xu, J.; Chen, B.J.; Zhao, D.Y.; Cui, S.; Zhou, T. Carboxymethylated degraded polysaccharides from Enteromorpha prolifera: Preparation and in vitro antioxidant activity. Food Chem. 2017, 215, 76–83. [Google Scholar] [CrossRef]
- Wei, J.; Wang, S.; Liu, G.; Pei, D.; Liu, Y.; Liu, Y.; Di, D. Polysaccharides from Enteromorpha prolifera enhance the immunity of normal mice. Int. J. Biol. Macromol. 2014, 64, 1–5. [Google Scholar] [CrossRef]
- Zhao, Y.; Balasubramanian, B.; Guo, Y.; Qiu, S.J.; Jha, R.; Liu, W.C. Dietary Enteromorpha polysaccharides supplementation improves breast muscle yield and is associated with modification of mRNA transcriptome in broiler chickens. Front. Vet Sci. 2021, 8, 663988. [Google Scholar] [CrossRef]
- Zhou, Z.; Pan, S.; Wu, S. Modulation of the growth performance, body composition and nonspecific immunity of crucian carp Carassius auratus upon Enteromorpha prolifera polysaccharide. Int. J. Biol. Macromol. 2020, 147, 29–33. [Google Scholar] [CrossRef]
- Guo, F.; Zhuang, X.; Han, M.; Lin, W. Polysaccharides from Enteromorpha prolifera protect against carbon tetrachloride-induced acute liver injury in mice via activation of Nrf2/HO-1 signaling, and suppression of oxidative stress, inflammation and apoptosis. Food Funct. 2020, 11, 4485–4498. [Google Scholar] [CrossRef]
- Guo, Y.; Zhao, Z.H.; Pan, Z.Y.; An, L.L.; Balasubramanian, B.; Liu, W.C. New insights into the role of dietary marine-derived polysaccharides on productive performance, egg quality, antioxidant capacity, and jejunal morphology in late-phase laying hens. Poult. Sci. 2020, 99, 2100–2107. [Google Scholar] [CrossRef]
- Liu, W.C.; Guo, Y.; Zhao, Z.H.; Jha, R.; Balasubramanian, B. Algae-derived polysaccharides promote growth performance by improving antioxidant capacity and intestinal barrier function in broiler chickens. Front. Vet. Sci. 2020, 7, 601336. [Google Scholar] [CrossRef]
- Guo, Y.; Balasubramanian, B.; Zhao, Z.H.; Liu, W.C. Marine algal polysaccharides alleviate aflatoxin B1-induced bursa of Fabricius injury by regulating redox and apoptotic signaling pathway in broilers. Poult. Sci. 2021, 100, 844–857. [Google Scholar] [CrossRef]
- Harley, C.D.; Anderson, K.M.; Demes, K.W.; Jorve, J.P.; Kordas, R.L.; Coyle, T.A.; Graham, M.H. Effects of climate change on global seaweed communities. J. Phycol. 2012, 48, 1064–1078. [Google Scholar] [CrossRef]
- Wang, Z.; Fu, M.; Xiao, J.; Zhang, X.; Song, W. Progress on the study of the Yellow Sea green tides caused by Enteromorpha prolifera. Acta Oceanol. Sin. 2018, 40, 1–13. [Google Scholar] [CrossRef]
- Lin, Y. Studies on Extraction, Purification and Bioactivities of Polysaccharides from Enteromorpha Prolifera. Master’s Thesis, Fujian Agriculture and Forestry University, Fuzhou, China, 2009. [Google Scholar]
- Hu, W. Polymer Physics: A molecular Approach; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012. [Google Scholar]
- Liu, L.L.; He, J.H.; Xie, H.B.; Yang, Y.S.; Li, J.C.; Zou, Y. Resveratrol induces antioxidant and heat shock protein mRNA expression in response to heat stress in black-boned chickens. Poult. Sci. 2014, 93, 54–62. [Google Scholar] [CrossRef]
- Sohail, M.; Ijaz, A.; Younus, M.; Shabbir, M.; Kamran, Z.; Ahmad, S.; Anwar, H.; Yousaf, M.; Ashraf, K.; Shahzad, A. Effect of supplementation of mannan oligosaccharide and probiotic on growth performance, relative weights of viscera, and population of selected intestinal bacteria in cyclic heat-stressed broilers. J. Appl. Poult. Res. 2013, 22, 485–491. [Google Scholar] [CrossRef]
- Hosseini-Vashan, S.J.; Raei-Moghadam, M.S. Antioxidant and immune system status, plasma lipid, abdominal fat, and growth performance of broilers exposed to heat stress and fed diets supplemented with pomegranate pulp (Punica granatum L.). J. Appl. Anim. Res. 2019, 47, 521–531. [Google Scholar] [CrossRef] [Green Version]
- Xu, D.N.; Li, W.Y.; Li, B.X.; Tian, Y.B.; Huang, Y. The effect of selenium and polysaccharide of Atractylodes macrocephala Koidz. (PAMK) on endoplasmic reticulum stress and apoptosis in chicken spleen induced by heat stress. RSC Adv. 2017, 7, 7519–7525. [Google Scholar] [CrossRef] [Green Version]
- Vandana, G.D.; Sejian, V.; Lees, A.M.; Pragna, P.; Silpa, M.V.; Maloney, S.K. Heat stress and poultry production: Impact and amelioration. Int. J. Biometeorol. 2021, 65, 163–179. [Google Scholar] [CrossRef] [PubMed]
- Awad, E.; Zulkifli, I.; Ramiah, S.; Khalil, E.; Abdallh, M. Prebiotics supplementation: An effective approach to mitigate the detrimental effects of heat stress in broiler chickens. World’s Poult. Sci. J. 2020, 7, 7519–7525. [Google Scholar] [CrossRef]
- Liu, W.C.; Zhou, S.H.; Balasubramanian, B.; Zeng, F.Y.; Sun, C.B.; Pang, H.Y. Dietary seaweed (Enteromorpha) polysaccharides improves growth performance involved in regulation of immune responses, intestinal morphology and microbial community in banana shrimp Fenneropenaeus merguiensis. Fish Shellfish Immun. 2020, 104, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Cui, H.; He, Y.; Cui, W.; Fang, J.; Zuo, Z.; Lai, W. Excess dietary sodium selenite alters apoptotic population and oxidative stress markers of spleens in broilers. Biol. Trace Elem. Res. 2012, 145, 47–51. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.S.; Li, Y.H.; Lin, M.F. Chronic exposure to the Fusarium mycotoxin deoxynivalenol: Impact on performance, immune organ, and intestinal integrity of slow-growing chickens. Toxins 2017, 9, 334. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Han, S.; Wang, Y.; Li, D.; Zhao, X.; Zhu, Q.; Yin, H. Oxidative stress and apoptotic changes in broiler chicken splenocytes exposed to T-2 toxin. BioMed Res. Inter. 2019, 2019, 5493870. [Google Scholar] [CrossRef]
- Zhao, H.; He, Y.; Li, S.; Sun, X.; Wang, Y.; Shao, Y.; Hou, Z.; Xing, M. Subchronic arsenism-induced oxidative stress and inflammation contribute to apoptosis through mitochondrial and death receptor dependent pathways in chicken immune organs. Oncotarget 2017, 8, 40327. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.C.; Zhu, Y.R.; Zhao, Z.H.; Jiang, P.; Yin, F.Q. Effects of dietary supplementation of algae-derived polysaccharides on morphology, tight junctions, antioxidant capacity and immune response of duodenum in broilers under heat stress. Animals 2021, 11, 2279. [Google Scholar] [CrossRef]
- Li, X.; Ding, X.; Peng, X.; Chi, X.; Cui, H.; Zuo, Z.; Fang, J. Effect of chitosan oligosaccharides on antioxidant function, lymphocyte cycle and apoptosis in ileum mucosa of broiler. Kafkas Univ. Vet. Fak. Derg. 2017, 23, 571–577. [Google Scholar]
- Banerjee, B.D.; Seth, V.; Bhattacharya, A.; Pasha, S.T.; Chakraborty, A.K. Biochemical effects of some pesticides on lipid peroxidation and free-radical scavengers. Toxicol. Lett. 1999, 107, 33–47. [Google Scholar] [CrossRef]
- Itoh, K.; Chiba, T.; Takahashi, S.; Ishii, T.; Igarashi, K.; Katoh, Y.; Oyake, T.; Hayashi, N.; Satoh, K.; Hatayama, I. An Nrf2/small Maf heterodimer mediates the induction of phase II detoxifying enzyme genes through antioxidant response elements. Biochem. Biophys. Res. Commun. 1997, 236, 313–322. [Google Scholar] [CrossRef]
- Arain, M.A.; Mei, Z.; Hassan, F.; Saeed, M.; Alagawany, M.; Shar, A.; Rajput, I. Lycopene: A natural antioxidant for prevention of heat-induced oxidative stress in poultry. World’s Poult. Sci. J. 2018, 74, 89–100. [Google Scholar] [CrossRef]
- Sahin, K.; Orhan, C.; Tuzcu, M.; Borawska, M.H.; Jabłonski, J.; Guler, O.; Sahin, N.; Hayirli, A. Berberis vulgaris root extract alleviates the adverse effects of heat stress via modulating hepatic nuclear transcription factors in quails. Brit. J. Nutr. 2013, 110, 609–616. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, T.; Suzuki, T.; Kobayashi, A.; Wakabayashi, J.; Maher, J.; Motohashi, H.; Yamamoto, M. Physiological significance of reactive cysteine residues of Keap1 in determining Nrf2 activity. Mol. Cell. Biol. 2008, 28, 2758–2770. [Google Scholar] [CrossRef] [Green Version]
- Itoh, K.; Wakabayashi, N.; Katoh, Y.; Ishii, T.; Igarashi, K.; Engel, J.D.; Yamamoto, M. Keap1 represses nuclear activation of antioxidant responsive elements by Nrf2 through binding to the amino-terminal Neh2 domain. Gene. Dev. 1999, 13, 76–86. [Google Scholar] [CrossRef] [Green Version]
- Sahin, K. Modulation of NF-κB and Nrf2 pathways by lycopene supplementation in heat-stressed poultry. World’s Poult. Sci. J. 2015, 71, 271–284. [Google Scholar] [CrossRef]
- Cheng, Y.; Chen, Y.; Chen, R.; Su, Y.; Zhang, R.; He, Q.; Wang, K.; Wen, C.; Zhou, Y. Dietary mannan oligosaccharide ameliorates cyclic heat stress-induced damages on intestinal oxidative status and barrier integrity of broilers. Poult. Sci. 2019, 98, 4767–4776. [Google Scholar] [CrossRef]
- Sandner, G.; Mueller, A.S.; Zhou, X.; Stadlbauer, V.; Schwarzinger, B.; Schwarzinger, C.; Wenzel, U.; Maenner, K.; van der Klis, J.D.; Hirtenlehner, S. Ginseng extract ameliorates the negative physiological effects of heat stress by supporting heat shock response and improving intestinal barrier integrity: Evidence from studies with heat-stressed Caco-2 cells, C. elegans and growing broilers. Molecules 2020, 25, 835. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Bai, K.; Su, W.; Wang, A.; Zhang, L.; Huang, K.; Wang, T. Curcumin attenuates heat-stress-induced oxidant damage by simultaneous activation of GSH-related antioxidant enzymes and Nrf2-mediated phase II detoxifying enzyme systems in broiler chickens. Poult. Sci. 2018, 97, 1209–1219. [Google Scholar] [CrossRef]
- Hu, X.; Zhang, R.; Xie, Y.; Wang, H.; Ge, M. The protective effects of polysaccharides from Agaricus blazei Murill against cadmium-induced oxidant stress and inflammatory damage in chicken livers. Biol. Trace Elem. Res. 2017, 178, 117–126. [Google Scholar] [CrossRef]
- Long, L.; Kang, B.; Jiang, Q.; Chen, J. Effects of dietary Lycium barbarum polysaccharides on growth performance, digestive enzyme activities, antioxidant status, and immunity of broiler chickens. Poult. Sci. 2020, 99, 744–751. [Google Scholar] [CrossRef]
- Zhang, C.; Li, C.; Shao, Q.; Chen, W.; Ma, L.; Xu, W.; Li, Y.; Huang, S.; Ma, Y. Effects of Glycyrrhiza polysaccharide in diet on growth performance, serum antioxidant capacity, and biochemistry of broilers. Poult. Sci. 2021, 100, 100927. [Google Scholar] [CrossRef]
- Zhang, S.; Wang, C.; Sun, Y.; Wang, G.; Chen, H.; Li, D.; Yu, X.; Chen, G. Xylanase and fermented Polysaccharide of Hericium caputmedusae reduce pathogenic infection of broilers by improving antioxidant and anti-inflammatory properties. Oxid. Med. Cell. Longev. 2018, 2018, 4296985. [Google Scholar] [CrossRef] [Green Version]
- Zhou, M.; Tao, Y.; Lai, C.; Huang, C.; Zhou, Y.; Yong, Q. Effects of mannanoligosaccharide supplementation on the growth performance, immunity, and oxidative status of partridge shank chickens. Animals 2019, 9, 817. [Google Scholar] [CrossRef] [Green Version]
- Xing, Y.Y.; Zheng, Y.K.; Yang, S.; Zhang, L.H.; Guo, S.W.; Shi, L.L.; Xu, Y.Q.; Jin, X.; Yan, S.M.; Shi, B.L. Artemisia ordosica polysaccharide alleviated lipopolysaccharide-induced oxidative stress of broilers via Nrf2/Keap1 and TLR4/NF-κB pathway. Ecotox. Environ. Saf. 2021, 223, 112566. [Google Scholar] [CrossRef]
- Xing, Y.Y.; Xu, Y.Q.; Jin, X.; Shi, L.L.; Guo, S.W.; Yan, S.M.; Shi, B.L. Optimization extraction and characterization of Artemisia ordosica polysaccharide and its beneficial effects on antioxidant function and gut microbiota in rats. RSC Adv. 2020, 10, 26151–26164. [Google Scholar] [CrossRef]
- Huang, C.; Cao, X.; Chen, X.; Fu, Y.; Zhu, Y.; Chen, Z.; Luo, Q.; Li, L.; Song, X.; Jia, R.; et al. A pectic polysaccharide from Ligusticum chuanxiong promotes intestine antioxidant defense in aged mice. Carbohyd. Polym. 2017, 174, 915–922. [Google Scholar] [CrossRef]
- Goel, A.; Ncho, C.M.; Choi, Y.H. Regulation of gene expression in chickens by heat stress. J. Anim. Sci. Biotechnol. 2021, 12, 11. [Google Scholar] [CrossRef] [PubMed]
- Lan, X.; Hsieh, J.C.; Schmidt, C.J.; Zhu, Q.; Lamont, S.J. Heat stress alters immune pathways in liver of divergent chicken lines. Anim. Indust. Rep. 2017, 663, 49. [Google Scholar]
- Baldwin, A.S., Jr. The NF-κB and IκB proteins: New discoveries and insights. Annu. Rev. Immunol. 1996, 14, 649–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chi, Q.; Wang, D.; Hu, X.; Li, S.; Li, S. Hydrogen sulfide gas exposure induces necroptosis and promotes inflammation through the MAPK/NF-κB pathway in broiler spleen. Oxid. Med. Cell. Longev. 2019, 2019, 8061823. [Google Scholar] [CrossRef] [Green Version]
- Romagnani, S. T-cell subsets (Th1 versus Th2). Ann. Allergy Asthma Immunol. 2000, 85, 9–21. [Google Scholar] [CrossRef]
- Han, Q.; Tong, J.; Sun, Q.; Teng, X.; Zhang, H.; Teng, X. The involvement of miR-6615-5p/Smad7 axis and immune imbalance in ammonia-caused inflammatory injury via NF-κB pathway in broiler kidneys. Poult. Sci. 2020, 99, 5378–5388. [Google Scholar] [CrossRef]
- Hu, X.; Chi, Q.; Wang, D.; Chi, X.; Teng, X.; Li, S. Hydrogen sulfide inhalation-induced immune damage is involved in oxidative stress, inflammation, apoptosis and the Th1/Th2 imbalance in broiler bursa of Fabricius. Ecotox. Environ. Saf. 2018, 164, 201–209. [Google Scholar] [CrossRef]
- Zhao, F.; Qu, J.; Wang, W.; Li, S.; Xu, S. The imbalance of Th1/Th2 triggers an inflammatory response in chicken spleens after ammonia exposure. Poult. Sci. 2020, 99, 3817–3822. [Google Scholar] [CrossRef]
- Liu, L.; Shen, J.; Zhao, C.; Wang, X.; Yao, J.; Gong, Y.; Yang, X. Dietary Astragalus polysaccharide alleviated immunological stress in broilers exposed to lipopolysaccharide. Int. J. Biol. Macromol. 2015, 72, 624–632. [Google Scholar] [CrossRef]
- Dong, N.; Li, X.; Xue, C.; Wang, C.; Xu, X.; Bi, C.; Shan, A.; Li, D. Astragalus polysaccharides attenuated inflammation and balanced the gut microflora in mice challenged with Salmonella typhimurium. Int. Immunopharmacol. 2019, 74, 105681. [Google Scholar] [CrossRef]
- Farag, M.R.; Elhady, W.M.; Ahmed, S.Y.; Taha, H.S.; Alagawany, M. Astragalus polysaccharides alleviate tilmicosin-induced toxicity in rats by inhibiting oxidative damage and modulating the expressions of HSP70, NF-kB and Nrf2/HO-1 pathway. Res. Vet. Sci. 2019, 124, 137–148. [Google Scholar] [CrossRef]
- Lambrou, G.I.; Hatziagapiou, K.; Vlahopoulos, S. Inflammation and tissue homeostasis: The NF-κB system in physiology and malignant progression. Mol. Biol. Rep. 2020, 47, 4047–4063. [Google Scholar] [CrossRef]
- Hoffmann, A.; Baltimore, D. Circuitry of nuclear factor κB signaling. Immunol. Rev. 2006, 210, 171–186. [Google Scholar] [CrossRef]
- Zhang, L.; Xiao, X.; Arnold, P.R.; Li, X.C. Transcriptional and epigenetic regulation of immune tolerance: Roles of the NF-κB family members. Cell Mol. Immunol. 2019, 16, 315–323. [Google Scholar] [CrossRef] [Green Version]
- Chawla, M.; Roy, P.; Basak, S. Role of the NF-κB system in context-specific tuning of the inflammatory gene response. Curr. Opin. Immunol. 2020, 68, 21–27. [Google Scholar] [CrossRef]
- Surai, P.F.; Kochish, I.I.; Kidd, M.T. Redox homeostasis in poultry: Regulatory roles of NF-κB. Antioxidants 2021, 10, 186. [Google Scholar] [CrossRef]
- Cheng, P.; Wang, T.; Li, W.; Muhammad, I.; Wang, H.; Sun, X.; Yang, Y.; Li, J.; Xiao, T.; Zhang, X. Baicalin alleviates lipopolysaccharide-induced liver inflammation in chicken by suppressing TLR4-mediated NF-κB pathway. Front. Pharmacol. 2017, 8, 547. [Google Scholar] [CrossRef] [Green Version]
- Tong, Y.; Yu, C.; Xie, Z.; Ziang, X.; Yang, Z.; Wang, T. Trans-anethole ameliorates lipopolysaccharide-induced acute liver inflammation in broilers via inhibiting NF-κB signaling pathway. Poult. Sci. 2022, 101, 101962. [Google Scholar] [CrossRef]
- Zhen, W.; Shao, Y.; Wu, Y. Dietary yeast β-glucan supplementation improves eggshell color and fertile eggs hatchability as well as enhances immune functions in breeder laying hens. Int. J. Biol. Macromol. 2020, 159, 607–621. [Google Scholar] [CrossRef]
- AOAC. Official methods of analysis of the Association of Official Analytical Chemists. 2012. Available online: https://www.aoac.org/ (accessed on 1 January 2012).
- Ishida, H. Quantitative surface FT-IR spectroscopic analysis of polymer. Rubber Chem. Technol. 1987, 60, 497–554. [Google Scholar] [CrossRef]
- Saeed, M.; Abbas, G.; Alagawany, M.; Kamboh, A.A.; Abd El-Hack, M.E.; Khafaga, A.F.; Chao, S. Heat stress management in poultry farms: A comprehensive overview. J. Therm. Biol. 2019, 84, 414–425. [Google Scholar] [CrossRef]
- NY/T33-2004; Chicken Feeding Standard. China Standard Press: Beijing, China, 2004.
- Gao, X.; Xiao, Z.H.; Liu, M.; Zhang, N.Y.; Khalil, M.M.; Gu, C.Q.; Qi, D.S.; Sun, L.H. Dietary silymarin supplementation alleviates zearalenone-induced hepatotoxicity and reproductive toxicity in rats. J. Nutr. 2018, 148, 1209–1216. [Google Scholar] [CrossRef] [Green Version]
- Luo, J.; Liu, H.; Wang, J.; Li, L.; Han, C.; Gan, X.; Li, Y.; Bai, L.; Mustafa, A. Transcriptome reveals B lymphocyte apoptosis in duck embryonic bursa of Fabricius mediated by mitochondrial and Fas signaling pathways. Mol. Immunol. 2018, 101, 120–129. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Contents, % |
|---|---|
| Total polysaccharides | 53.32 |
| Sulfate | 19.87 |
| Uronic acid | 12.66 |
| Moisture | 3.58 |
| Protein | 1.65 |
| Unspecified compounds (lipid, salt, etc.) | 8.92 |
| Items | Contents, % |
|---|---|
| Glucosamine (GlcN) | 50.81 |
| Glucose (Glc) | 27.70 |
| Galacturonic acid (GalA) | 11.75 |
| Mannose (Man) | 2.78 |
| Xylose (Xyl) | 2.63 |
| Galactose (Gal) | 1.95 |
| Arabinose (Ara) | 0.93 |
| Glucuronic acid (GlcA) | 0.76 |
| Fucose (Fuc) | 0.27 |
| Galactosamine (GalN) | 0.25 |
| Ribose (Rib) | 0.17 |
| Items | Units | TN | HS | HSS | p-Value | ||
|---|---|---|---|---|---|---|---|
| TN vs. HS | HS vs. HSS | TN vs. HSS | |||||
| T-AOC | mmol/mg protein | 36.36 ± 1.39 | 23.91 ± 2.37 | 32.10 ± 1.70 | <0.001 | 0.010 | 0.132 |
| T-SOD | U/mg protein | 63.12 ± 2.79 | 46.21 ± 4.14 | 54.90 ± 4.28 | 0.029 | 0.218 | 0.242 |
| GSH-Px | U/mg protein | 126.39 ± 8.02 | 130.06 ± 7.86 | 125.28 ± 4.02 | 0.709 | 0.627 | 0.908 |
| CAT | U/mg protein | 3.62 ± 0.46 | 3.38 ± 0.39 | 3.15 ± 0.37 | 0.698 | 0.746 | 0.481 |
| GST | U/mg protein | 42.00 ± 1.96 | 26.42 ± 1.41 | 38.39 ± 2.00 | <0.001 | 0.002 | 0.223 |
| MDA | nmol/mg protein | 2.28 ± 0.15 | 4.12 ± 0.30 | 2.95 ± 0.43 | 0.019 | 0.045 | 0.211 |
| ROS | fluorescence/mg protein | 92.98 ± 3.04 | 131.75 ± 10.91 | 114.50 ± 6.18 | 0.016 | 0.225 | 0.138 |
| Items | Units | TN | HS | HSS | p-Value | ||
|---|---|---|---|---|---|---|---|
| TN vs. HS | HS vs. HSS | TN vs. HSS | |||||
| TNF-α | pg/mg protein | 203.68 ± 5.91 | 252.65 ± 10.38 | 218.10 ± 5.10 | 0.001 | 0.011 | 0.224 |
| IFN-γ | pg/mg protein | 421.19 ± 9.87 | 377.94 ± 7.88 | 407.04 ± 10.96 | 0.024 | 0.103 | 0.404 |
| IL-1β | pg/mg protein | 310.97 ± 9.60 | 338.40 ± 9.48 | 320.76 ± 10.08 | 0.141 | 0.329 | 0.581 |
| IL-2 | ng/mg protein | 10.38 ± 0.41 | 8.25 ± 0.27 | 10.16 ± 0.33 | <0.001 | 0.001 | 0.648 |
| IL-4 | ng/mg protein | 72.69 ± 6.09 | 113.72 ± 7.45 | 81.12 ± 1.76 | <0.001 | <0.001 | 0.853 |
| IL-6 | pg/mg protein | 186.48 ± 6.81 | 207.26 ± 9.46 | 198.05 ± 2.80 | 0.126 | 0.475 | 0.376 |
| IL-10 | ng/mg protein | 181.22 ± 7.83 | 224.09 ± 12.62 | 190.46 ± 6.71 | 0.008 | 0.026 | 0.489 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, W.-C.; Zhuang, D.-P.; Zhao, Y.; Balasubramanian, B.; Zhao, Z.-H. Seaweed-Derived Polysaccharides Attenuate Heat Stress-Induced Splenic Oxidative Stress and Inflammatory Response via Regulating Nrf2 and NF-κB Signaling Pathways. Mar. Drugs 2022, 20, 358. https://doi.org/10.3390/md20060358
Liu W-C, Zhuang D-P, Zhao Y, Balasubramanian B, Zhao Z-H. Seaweed-Derived Polysaccharides Attenuate Heat Stress-Induced Splenic Oxidative Stress and Inflammatory Response via Regulating Nrf2 and NF-κB Signaling Pathways. Marine Drugs. 2022; 20(6):358. https://doi.org/10.3390/md20060358
Chicago/Turabian StyleLiu, Wen-Chao, Di-Pai Zhuang, Yue Zhao, Balamuralikrishnan Balasubramanian, and Zhi-Hui Zhao. 2022. "Seaweed-Derived Polysaccharides Attenuate Heat Stress-Induced Splenic Oxidative Stress and Inflammatory Response via Regulating Nrf2 and NF-κB Signaling Pathways" Marine Drugs 20, no. 6: 358. https://doi.org/10.3390/md20060358
APA StyleLiu, W. -C., Zhuang, D. -P., Zhao, Y., Balasubramanian, B., & Zhao, Z. -H. (2022). Seaweed-Derived Polysaccharides Attenuate Heat Stress-Induced Splenic Oxidative Stress and Inflammatory Response via Regulating Nrf2 and NF-κB Signaling Pathways. Marine Drugs, 20(6), 358. https://doi.org/10.3390/md20060358