
Antitumor Potential of Immunomodulatory Natural Products

 ,
, 
 , and
, and 
Abstract
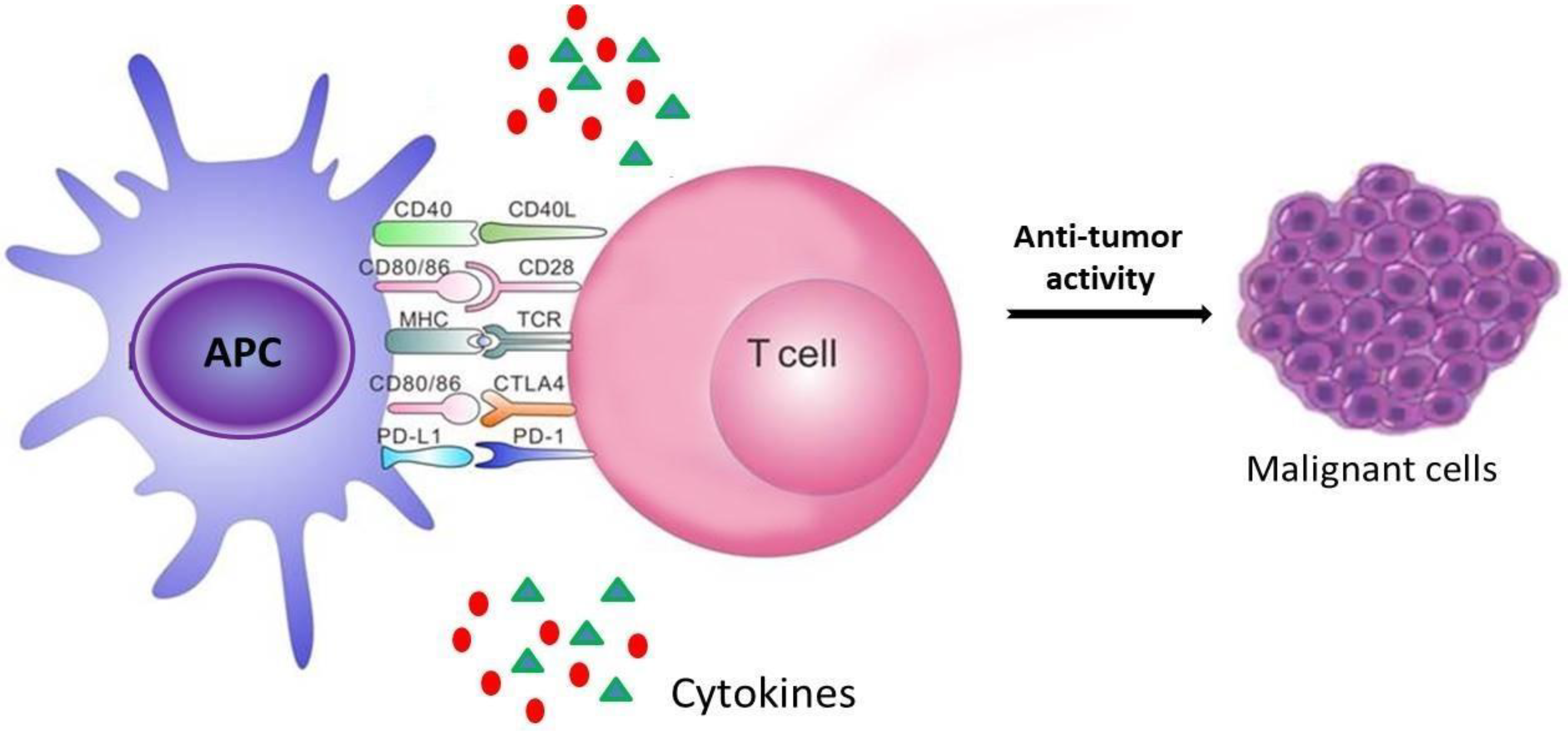
:1. Introduction
2. Terrestrial Environment
2.1. Plant Compounds
2.1.1. Terpenes
2.1.2. Phenolic Compounds
2.2. Therapeutic Antitumor Activity and Natural Compounds from Spices
2.3. Macromycetes
2.4. Other Sources

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Molecule | Type of Compound | Source | Tumor | Immuno System’s Role | References |
|---|---|---|---|---|---|
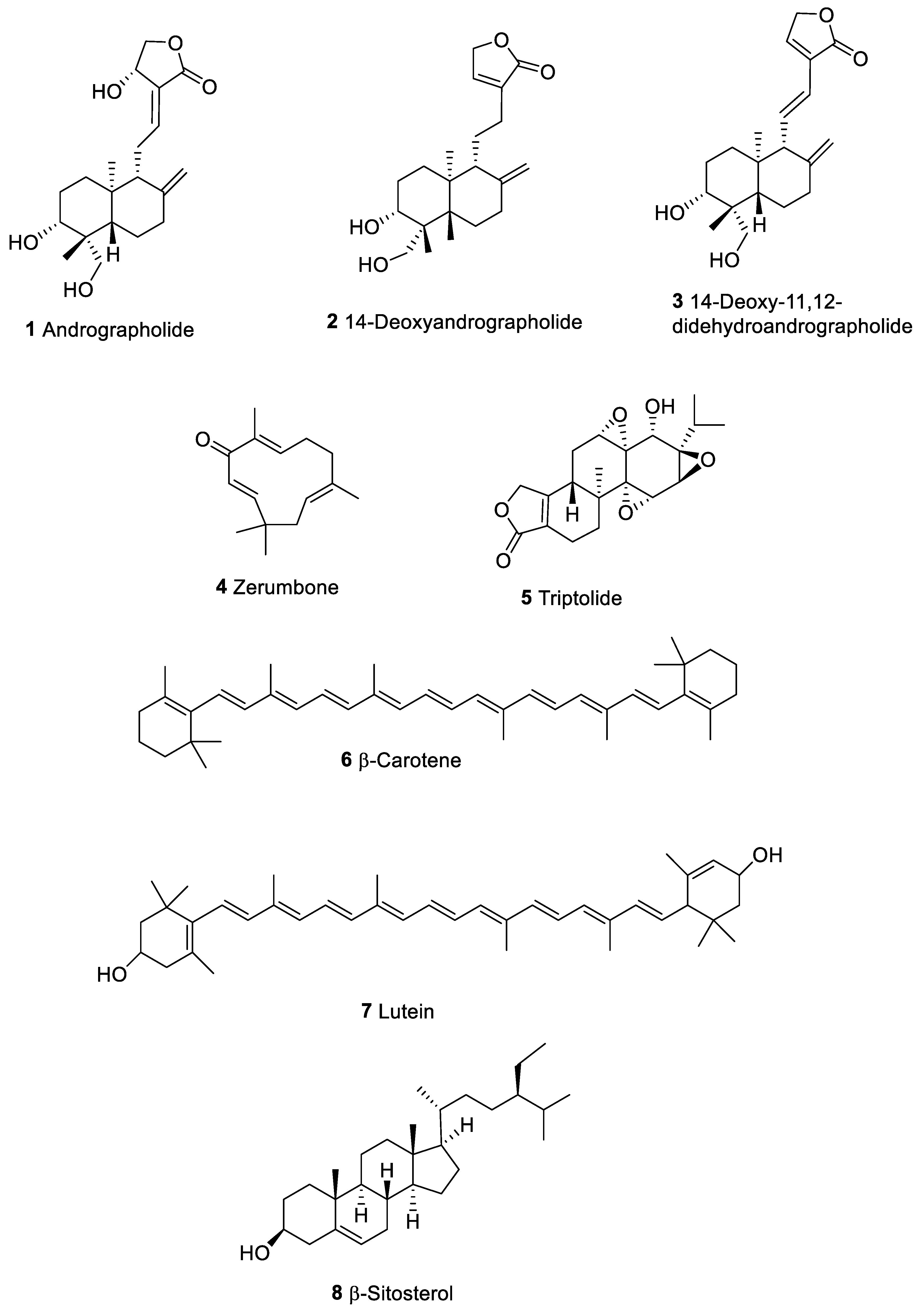
| Andrographolide (1) | Terpene | Andrographis paniculata | Human epidermal carcinoma (KB, ED50 1.5 µg/mL); lymphocytic leukemia (P388, ED50 1.0 µg/mL) | Stimulate antigen specific and non-specific immune responses in mice | [29] |
| Triptolide (5) | Terpene | Chinese Tripterygium Hook F (TWHF) | Solid tumor cells | Apoptosis induced by TNFα, inhibition of NF-kB | [44] |
| Zerumbone (4) | Terpene | Zingiberaceae | Human cancer cell lines of the ovary (Coav-3) breast (MCF-7) promyelocytic leukemia (HL-60) and colon adenocarcinoma HCT116 | Immunosuppressive effects via inhibition of AP-1 and NF-kB | [36,37,38,39] |
| β-Carotene (6); Lutein (7) | Carotenoids | Plant | Lung human cancer, mammary tumor bearing mice model | Stimulate NK cell activities, increase the number of leukocyte immune cells, CD4/CD8 ratio, and surface expression of MHC I molecules Stimulation effect on IFN-γ mRNA expression; suppression of IL-10 in splenocytes | [47,48,49] |
| β-Sitosterol (8) | Terpene | Plant | Human cancer cell line of the colon (HT-29) and prostate (LNCaP) | stimulated blood lymphocyte proliferation in vitro; enhanced lytic and cytotoxic activities of NK cells | [53,161] |
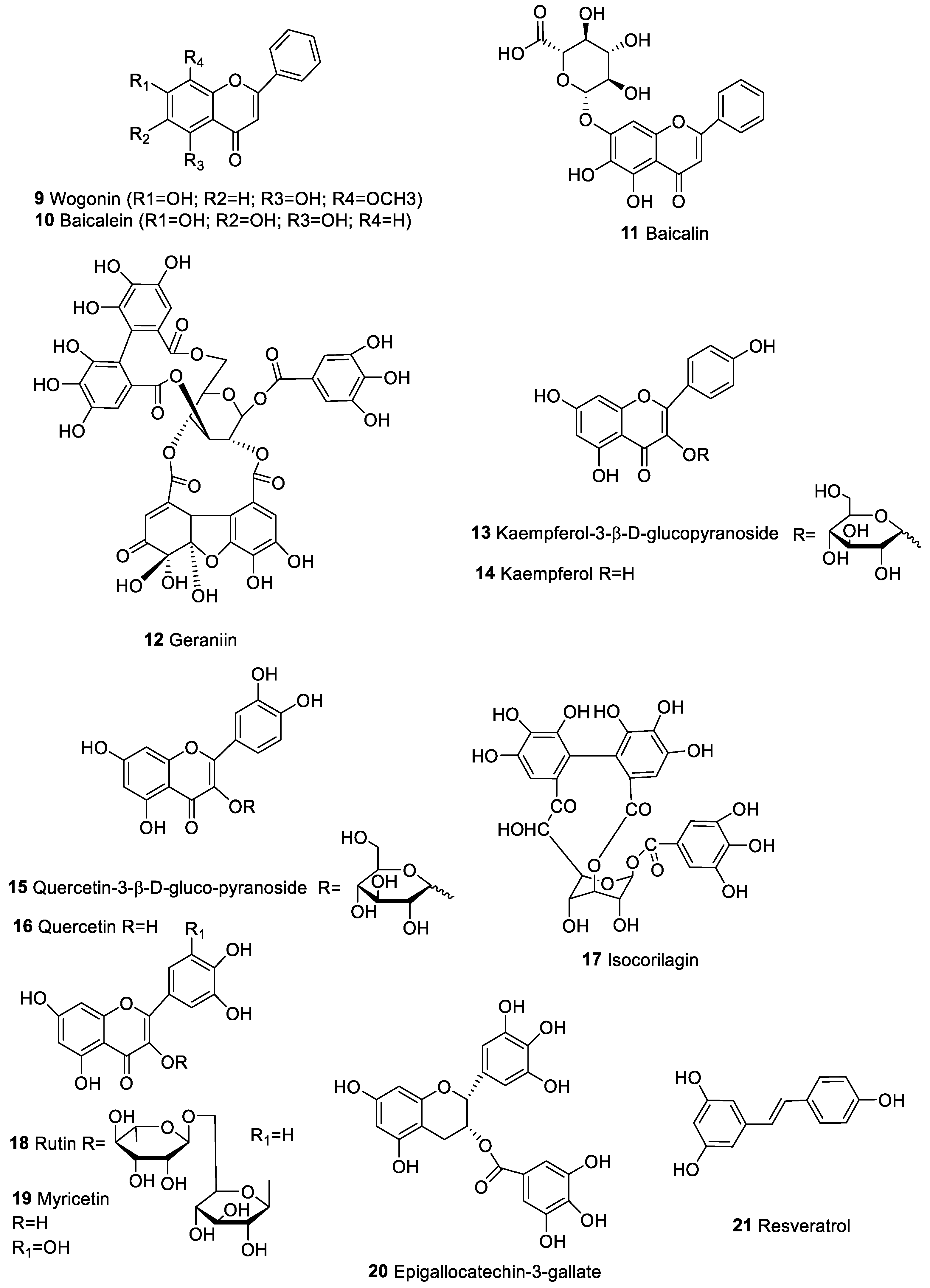
| Wogonin (9), Baicalein (10), Baicalin (11) | Flavones | Scutellaria baicalenis | Breast, prostate, and lung human cancer | Activation of NF-kB factor; cell cycle regulation | [60] |
| Geraniin (12), | Phenolic compounds | Phyllanthus emblica Linnaeus | Human cancer cell line of the breast (MCF-7) and embryonic fibroblast (HELF) | Promoted the level of serum IL-18 and NK cell cytotoxicity, suggesting stimulation of macrophages, thereby upregulating the NK cell-mediated antitumor immune response | [79] |
| Kaempferol 3-β−d-glucopyranoside (13) | |||||
| Kaempferol (14) | |||||
| Quercetin 3-β-d-glucopyranoside (15) | |||||
| Quercetin (16) | |||||
| Isocorilagin (17) | |||||
| Ruitin (18) | Phenolic compounds | Diospyros kaki L. | Leukemia, colorectal, neuroblastoma, melanotic melanoma and prostate human cancer. | Increase of IL-18; Upregulation of NK cells | [83,84,85] |
| Myricetin (19) | |||||
| Epigallocatechin-3-gallate (20) | Phenolic compound | Camelia sinesis | Human lung cancer cell lines | Induction of apoptosis and suppression of NF-kB pathway Inhibition of PD-L1 | [89,90,91,97] |
| Resveratrol (21) | Phennolic compound | Grapes and red wine | Brest, oral, liver, prostate and colon human cancer | Inhibition of citokyne production, (IFN-γ, IL-2, TNF-α and IL-12); block the activation of transcription factor NF-kB | [104,105,106,110] |
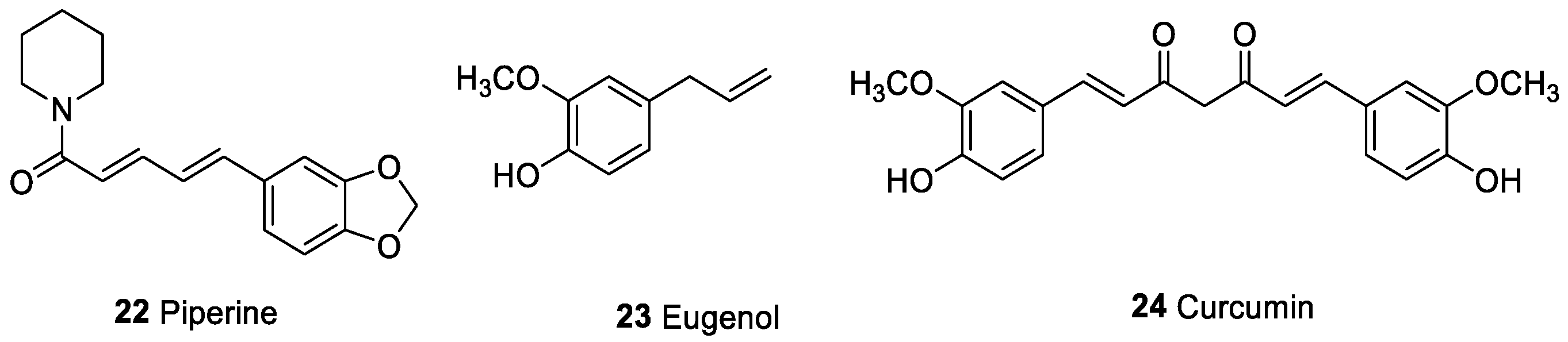
| Piperine (22) | Alkaloid | Piper nigrum | Human colon cancer cell lines | Cytotoxic activity of NK cells; suppression of the relase of Th2 cytokines IL-4 and IL-10; enhance murine splenocyte proliferation | [112,115,116,117,121] |
| Eugenol (23) | Phenolic compound | Cardamom | Inhibition of tumor formation in vivo | Cytotoxic activity of NK cells; suppression of the relase of Th2 cytokines IL-4 and IL-10 | [112,123,124,125,126] |
| Curcumin (24) | Phenolic compound | Curcuma longa | Human breast cancer | Modulation of NF-kB; reduction of IL-6; inhibit inflammation-mediated PD-L1 expression | [129,130,131,133,134] |
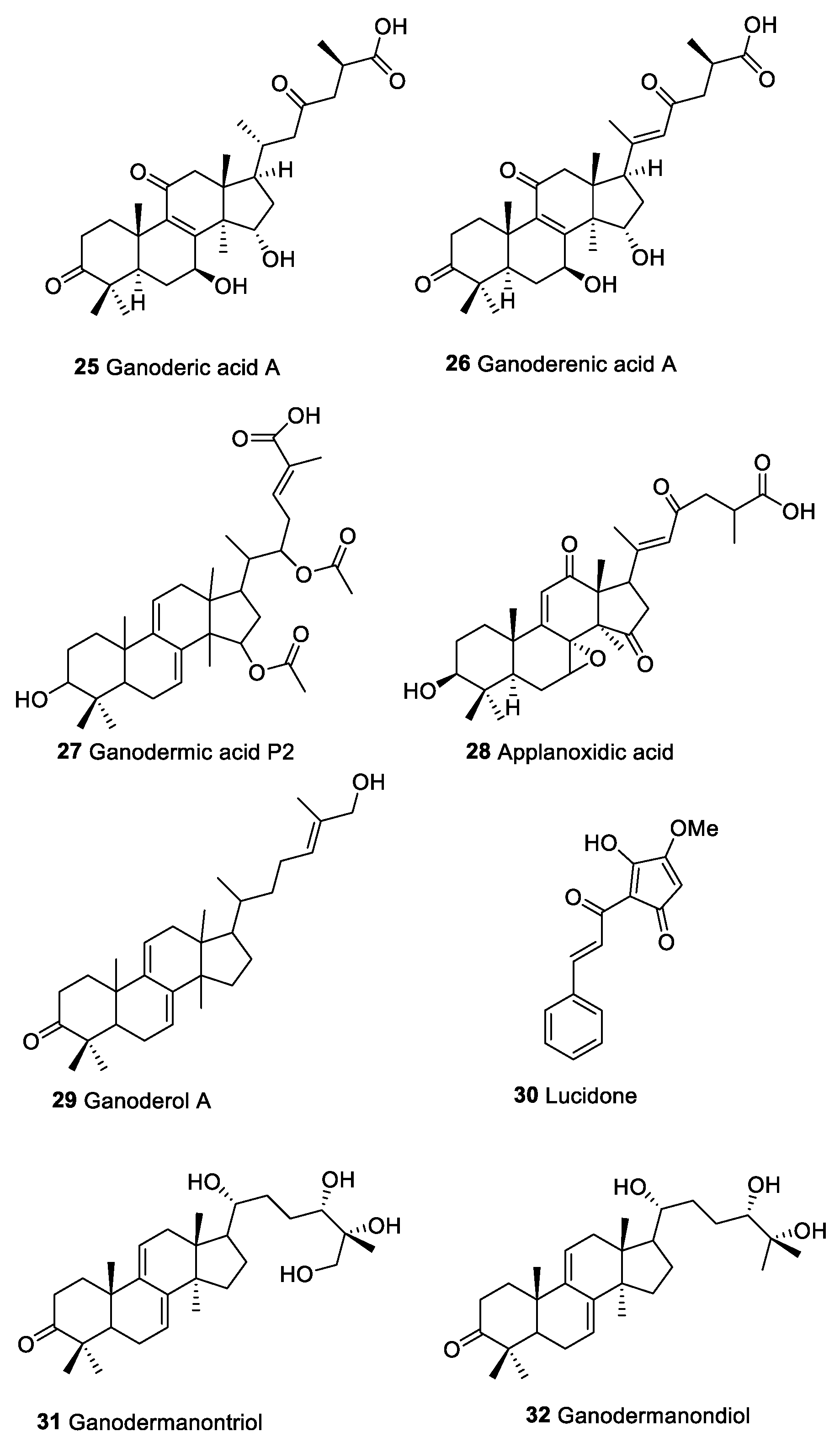
| Ganoderic acids (25) | Triterpenoid compounds | Macromycetes | Act on immune effecter cells such as hematpoietic stem cells, lymphocytes, macrophages, T cells, DCs, and NK cells Activation NF-kB pathway and modulate Ras/Erk, c-myc, CREB protein and MAPK | [139] | |
| Ganoderenic acids (26) | |||||
| Ganodermic acids P2 (27) | |||||
| Applanoxidic acid (28) | |||||
| Ganoderol A (29) | |||||
| Lucidone (30) | |||||
| Ganodermanontriol (31) | |||||
| Ganodermanondiol (32) | |||||
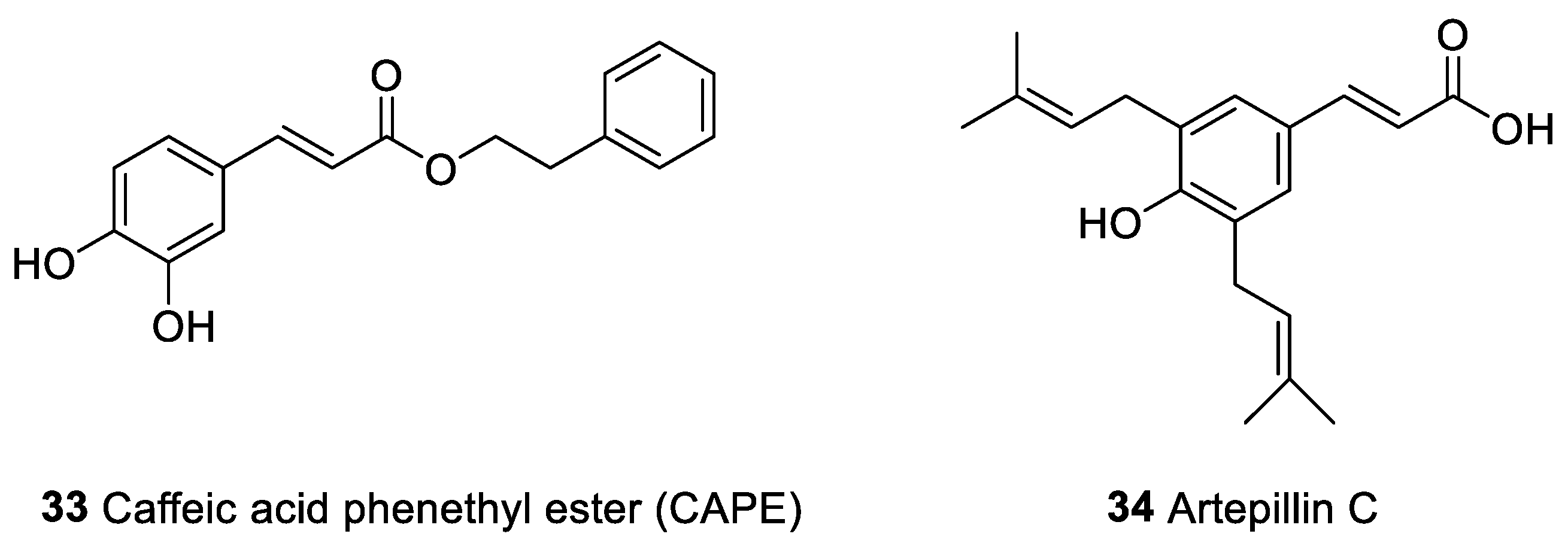
| Caffeic acid phenethyl ester (CAPE, 33) | Phenolic compounds | Propolis | Oral human cancer and human cancer cell lines of the promyelocytic leukemia (HL-60) | Inhibition of T cell receptor-mediated T cell proliferation | [150,151,154,158,159] |
| Artepilin C (34) |
3. Compounds from Marine Environment
| Molecule | Source | Tumor | Immuno System’s Role | References |
|---|---|---|---|---|
| Glycopeptide | Alexandrium minutum | A549 Lung adenocarcinoma cell line | Mitophagy and ICD inducer | [202] |
| Polyunsaturated aldehydes | diatoms | Programmed cell death in lung and colon adenocarcinoma | Induce the release of ATP and others immune signals which are known as ICD inducers | [21,163,202,203,204,205] |
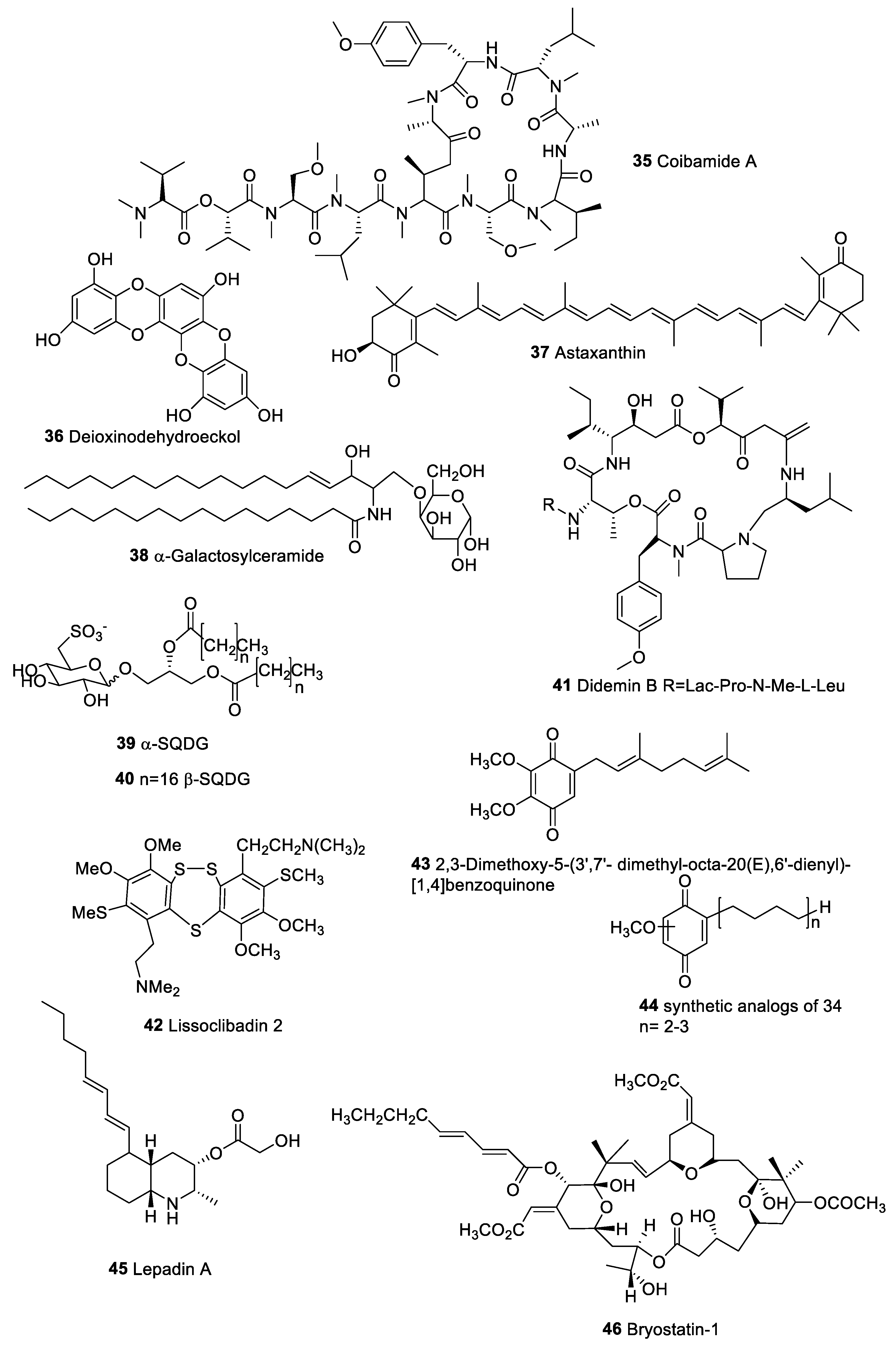
| Coibamida A (35) | Leptolyngbya sp. | Breast camcer | Caspase-independent cell death and ICD inducer | [168] |
| Dioxinodehydroeckol (36) | Ecklonia cava | Human cancer cell line of the breast (MCF-7) | Induction of apoptosis through NF-kB family and NF-kB-dependent pathway | [169] |
| Astaxanthin (37) | Seaweeds | Antitumoral activity in the post-initiation phase of carcinogen-induced colon and oral cancer models | Improves antitumor immune responses by inhibiting lipid peroxidation induced by stress | [170,171,172,173] |
| α Galactosylceramide (38) | Sponge | Antitumor effects in mice | Stimulation of NKT cells to produce both Th1 and Th2 cytokines | [174] |
| α-Sulfoquinovosides (39) | Marine microalgae | Synthetic β-sulfoquinovosides derivative as adjuvant in vaccine against a murine B16F10 melanoma cell line | Maturation of human DCs. | [176,177] |
| Didemin B (41) | Trididemnum solidum | Inhibition of lymphocyte activation | [181,182,183] | |
| Lissoclibadin 2 (42) | Lissoclinum cf. badium | Human colon cancer lines (DLD-1) and (HCT116), breast cancer lines MDA-MB-231, renal cancer line ACHN; non-small-cell lung cancer line NCI-H460 | Increase of IL-8 production | [187] |
| 2,3-Dimethoxy-5-(3′,7′-dimethyl-octa-20(E),6′-dienyl)-[1,4] benzoquinone (43) | Aplidium glabrum | JB6 CI41 cancer cell | Inhibition of p53; Increase transcription of AP-1 and NF-kB | [186,188] |
| Lepadin A (45) | Clavelina lepadiformis sp. B | Human lung carcinoma, melanoma, and multiple myeloma | Mouse DCs | [189] |
| Bryostatin 1 (46) | Bugula neritina | Antitumor activity against leukemia, lymphoma ovarian cancer, and melanoma | Activation of PKC family; Stimulation of cytokine production | [193,194] |
4. Conclusions: Future Prospects of Natural Compounds as Potential Anti-Cancer Agents
Author Contributions
Funding
Conflicts of Interest
Abbreviations
References
- Thomford, N.E.; Senthebane, D.A.; Rowe, A.; Munro, D.; Seele, P.; Maroyi, A.; Dzobo, K. Natural Products for Drug Discovery in the 21st Century: Innovations for Novel Drug Discovery. Int. J. Mol. Sci. 2018, 19, 1578. [Google Scholar] [CrossRef] [Green Version]
- Reid, R.G.; Sarker, S.D. Isolation of Natural Products by Low-Pressure Column Chromatography. Methods Mol. Biol. 2012, 864, 155–187. [Google Scholar] [CrossRef]
- Sticher, O. Natural Product Isolation. Nat. Prod. Rep. 2008, 25, 517–554. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs over the Nearly Four Decades from 01/1981 to 09/2019. J. Nat. Prod. 2020, 83, 770–803. [Google Scholar] [CrossRef]
- Rayan, A.; Raiyn, J.; Falah, M. Nature Is the Best Source of Anticancer Drugs: Indexing Natural Products for Their Anticancer Bioactivity. PLoS ONE 2017, 12, e0187925. [Google Scholar] [CrossRef]
- Heinig, U.; Jennewein, S. Taxol: A complex diterpenoid natural product with an evolutionarily obscure origin. Afr. J. Biotechnol. 2009, 8, 1370–1385. [Google Scholar]
- Cuevas, C.; Francesch, A. Development of Yondelis® (Trabectedin, ET-743). A Semisynthetic Process Solves the Supply Problem. Nat. Prod. Rep. 2009, 26, 322–337. [Google Scholar] [CrossRef]
- Pommier, Y.; Kohlhagen, G.; Bailly, C.; Waring, M.; Mazumder, A.; Kohn, K.W. DNA Sequence-and Structure-Selective Alkylation of Guanine N2 in the DNA Minor Groove by Ecteinascidin 743, a Potent Antitumor Compound from the Caribbean Tunicate Ecteinascidia Turbinata. Biochemistry 1996, 35, 13303–13309. [Google Scholar] [CrossRef]
- Bracci, L.; Schiavoni, G.; Sistigu, A.; Belardelli, F. Immune-Based Mechanisms of Cytotoxic Chemotherapy: Implications for the Design of Novel and Rationale-Based Combined Treatments against Cancer. Cell Death Differ. 2014, 21, 15–25. [Google Scholar] [CrossRef] [Green Version]
- Norling, L.V.; Serhan, C.N. Profiling in Resolving Inflammatory Exudates Identifies Novel Anti-Inflammatory and pro-Resolving Mediators and Signals for Termination. J. Intern. Med. 2010, 268, 15–24. [Google Scholar] [CrossRef]
- Zhu, H.F.; Li, Y. Small-Molecule Targets in Tumor Immunotherapy. Nat. Prod. Bioprospec. 2018, 8, 297–301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ngamkham, S.; Holden, J.E.; Smith, E.L. A Systematic Review: Mindfulness Intervention for Cancer-Related Pain. Asia-Pac. J. Oncol. Nurs. 2019, 6, 161–169. [Google Scholar] [CrossRef] [PubMed]
- Yoest, J. Clinical Features, Predictive Correlates, and Pathophysiology of Immune-Related Adverse Events in Immune Checkpoint Inhibitor Treatments in Cancer: A Short Review. Immuno. Targets Ther. 2017, 6, 73–82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhong, Z.; Vong, C.T.; Chen, F.; Tan, H.; Zhang, C.; Wang, N.; Cui, L.; Wang, Y.; Feng, Y. Immunomodulatory Potential of Natural Products from Herbal Medicines as Immune Checkpoints Inhibitors: Helping to Fight against Cancer via Multiple Targets. Med. Res. Rev. 2022, 42, 1246–1279. [Google Scholar] [CrossRef] [PubMed]
- Zitvogel, L.; Apetoh, L.; Ghiringhelli, F.; Kroemer, G. Immunological Aspects of Cancer Chemotherapy. Nat. Rev. Immunol. 2008, 8, 59–73. [Google Scholar] [CrossRef]
- Galluzzi, L.; Senovilla, L.; Zitvogel, L.; Kroemer, G. The Secret Ally: Immunostimulation by Anticancer Drugs. Nat. Rev. Drug Discov. 2012, 11, 215–233. [Google Scholar] [CrossRef]
- Kawano, M.; Tanaka, K.; Itonaga, I.; Iwasaki, T.; Miyazaki, M.; Ikeda, S.; Tsumura, H. Dendritic Cells Combined with Doxorubicin Induces Immunogenic Cell Death and Exhibits Antitumor Effects for Osteosarcoma. Oncol. Lett. 2016, 11, 2169–2175. [Google Scholar] [CrossRef] [Green Version]
- Pan, P.; Huang, Y.W.; Oshima, K.; Yearsley, M.; Zhang, J.; Arnold, M.; Yu, J.; Wang, L.S. The Immunomodulatory Potential of Natural Compounds in Tumor-Bearing Mice and Humans. Crit. Rev. Food Sci. Nutr. 2019, 59, 992–1007. [Google Scholar] [CrossRef]
- Mohamed, S.I.A.; Jantan, I.; Haque, M.A. Naturally Occurring Immunomodulators with Antitumor Activity: An Insight on Their Mechanisms of Action. Int. Immunopharmacol. 2017, 50, 291–304. [Google Scholar] [CrossRef]
- Moody, R.; Wilson, K.; Jaworowski, A.; Plebanski, M. Natural Compounds with Potential to Modulate Cancer Therapies and Self-Reactive Immune Cells. Cancers 2020, 12, 673. [Google Scholar] [CrossRef] [Green Version]
- Sansone, C.; Bruno, A.; Piscitelli, C.; Baci, D.; Fontana, A.; Brunet, C.; Noonan, D.M.; Albini, A. Natural Compounds of Marine Origin as Inducers of Immunogenic Cell Death (ICD): Potential Role for Cancer Interception and Therapy. Cells 2021, 10, 1–20. [Google Scholar] [CrossRef]
- Katanaev, V.L.; di Falco, S.; Khotimchenko, Y. The Anticancer Drug Discovery Potential of Marine Invertebrates from Russian Pacific. Marine Drugs 2019, 17, 474. [Google Scholar] [CrossRef] [Green Version]
- Riccio, G.; Lauritano, C. Microalgae with Immunomodulatory Activities. Marine Drugs 2020, 18, 2. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, B.B.; Kunnumakkara, A.B.; Harlkumar, K.B.; Tharakan, S.T.; Sung, B.; Anand, P. Potential of Spice-Derived Phytochemicals for Cancer Prevention. Planta Med. 2008, 74, 1560–1569. [Google Scholar] [CrossRef] [Green Version]
- Shen, Y.-C.; Chen, C.-F.; Chiou, W.-F. Andrographolide Prevents Oxygen Radical Production by Human Neutrophils: Possible Mechanism(s) Involved in Its Anti-Intinflammatory Effect. Br. J. Pharmacol. 2002, 135, 399–406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shihman Chang, R.; Ding, L.; Gai-qing, C.; Ze-lin, Z.; SMITHt, K.M. Dehydroandrographolide Succinic Acid Monoester as an Inhibitor against the Human Immunodeficiency Virus (43225). Proc. Soc. Exp. Biol. Med. 1991, 197, 59–66. [Google Scholar] [CrossRef]
- Hossain, S.; Urbi, Z.; Karuniawati, H.; Mohiuddin, R.B.; Moh Qrimida, A.; Allzrag, A.M.M.; Ming, L.C.; Pagano, E.; Capasso, R. Andrographis paniculata (Burm. f.) Wall. ex Nees: An Updated Review of Phytochemistry, Antimicrobial Pharmacology, and Clinical Safety and Efficacy. Life 2021, 11, 348. [Google Scholar] [CrossRef]
- Puri, A.; Saxena, R.; Saxena, R.P.; Saxena, K.C.; Srivastava, V.; Tandón, J.S. Immunostimulant Agents from Andrographis Paniculata1. J. Nat. Prod. 1993, 56, 995–999. [Google Scholar] [CrossRef]
- Ajaya Kumar, R.; Sridevi, K.; Vijaya Kumar, N.; Nanduri, S.; Rajagopal, S. Anticancer and Immunostimulatory Compounds from Andrographis Paniculata. J. Ethnopharmacol. 2004, 92, 291–295. [Google Scholar] [CrossRef]
- Ghazalee, N.S.; Jantan, I.; Arshad, L.; Haque, M.A. Immunosuppressive Effects of the Standardized Extract of Zingiber Zerumbet on Innate Immune Responses in Wistar Rats. Phytother. Res. 2019, 33, 929–938. [Google Scholar] [CrossRef]
- Girisa, S.; Shabnam, B.; Monisha, J.; Fan, L.; Halim, C.E.; Arfuso, F.; Ahn, K.S.; Sethi, G.; Kunnumakkara, A.B. Potential of Zerumbone as an Anti-Cancer Agent. Molecules 2019, 24, 734. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arshad, L.; Jantan, I.; Bukhari, S.N.A.; Haque, M.A. Immunosuppressive Effects of Natural α,β-Unsaturated Carbonyl-Based Compounds, and Their Analogs and Derivatives, on Immune Cells: A Review. Front. Pharmacol. 2017, 8, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prasannan, R.; Kalesh, K.A.; Shanmugam, M.K.; Nachiyappan, A.; Ramachandran, L.; Nguyen, A.H.; Kumar, A.P.; Lakshmanan, M.; Ahn, K.S.; Sethi, G. Key Cell Signaling Pathways Modulated by Zerumbone: Role in the Prevention and Treatment of Cancer. Biochem. Pharmacol. 2012, 84, 1268–1276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haque, M.A.; Jantan, I.; Arshad, L.; Bukhari, S.N.A. Exploring the Immunomodulatory and Anticancer Properties of Zerumbone. Food Funct. 2017, 8, 3410–3431. [Google Scholar] [CrossRef] [PubMed]
- Murakami, A.; Shigemori, T.; Ohigashi, H. International Conference on Diet, Nutrition, and Cancer Zingiberaceous and Citrus Constituents, 1-Acetoxychavicol Acetate, Zerumbone, Auraptene, and Nobiletin, Suppress Lipopolysaccharide-Induced Cyclooxygenase-2 Expression in RAW264.7 Murine Macrophages through Different Modes of Action. J. Nutr. 2005, 135, 2987S–2992S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eguchi, A.; Kaneko, Y.; Murakami, A.; Ohigashi, H. Zerumbone Suppresses Phorbol Ester-Induced Expression of Multiple Scavenger Receptor Genes in THP-1 Human Monocytic Cells. Biosci. Biotechnol. Biochem. 2007, 71, 935–945. [Google Scholar] [CrossRef] [Green Version]
- Murakami, A.; Takahashi, D.; Kinoshita, T.; Koshimizu, K.; Kim, H.W.; Yoshihiro, A.; Nakamura, Y.; Jiwajinda, S.; Terao, J.; Ohigashi, H. Zerumbone, a Southeast Asian Ginger Sesquiterpene, Markedly Suppresses Free Radical Generation, Proinflammatory Protein Production, and Cancer Cell Proliferation Accompanied by Apoptosis: The α,β-Unsaturated Carbonyl Group Is a Prerequisite. Carcinogenesis 2002, 23, 795–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdel Wahab, S.I.; Abdul, A.B.; Alzubairi, A.S.; Mohamed Elhassan, M.; Mohan, S. In Vitro Ultramorphological Assessment of Apoptosis Induced by Zerumbone on (HeLa). J. Biomed. Biotechnol. 2009, 2009. [Google Scholar] [CrossRef]
- Yodkeeree, S.; Sung, B.; Limtrakul, P.; Aggarwal, B.B. Zerumbone Enhances TRAIL-Induced Apoptosis through the Induction of Death Receptors in Human Colon Cancer Cells: Evidence for an Essential Role of Reactive Oxygen Species. Cancer Res. 2009, 69, 6581–6589. [Google Scholar] [CrossRef] [Green Version]
- Kupchan, S.M.; Court, W.A.; Dailey, R.G., Jr.; Gilmore, C.J.; Bryan, R.F. Triptolide and Tripdiolide, Novel Antileukemic Diterpenoid Triepoxides from Tripterygium Wilfordii. J. Am. Chem. Soc. 1972, 94, 7194–7195. [Google Scholar] [CrossRef]
- Tao, X.; Cai, J.J.; Lipsky, P.E. The Identity of Immunosuppressive Components of the Ethyl Acetate Extract and Chloroform Methanol Extract. J. Pharmacol. Exp. Ther. 1995, 272, 1305–1312. [Google Scholar] [PubMed]
- Tao, X.; Davis, L.S.; Hashhoto, K.; Lipsky, R.E. The Chinese Herbal Remedy, T2. Inhibits Mitogen-Induced Cytokine Gene Transcription by T Cells, but Not Initial Signal Transduction. J. Pharmacol. Exp.Ther. 1996, 276, 316–325. [Google Scholar]
- Qiu, D.; Zhao, G.; Aoki, Y.; Shi, L.; Uyei, A.; Nazarian, S.; Ng, J.C.H.; Kao, P.N. Immunosuppressant PG490 (Triptolide) Inhibits T-Cell Interleukin-2 Expression at the Level of Purine-Box/Nuclear Factor of Activated T-Cells and NF-ΚB Transcriptional Activation. J. Biol. Chem. 1999, 274, 13443–13450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.Y.; Chang, W.T.; Qiu, D.; Kao, P.N.; Rosen, G.D. PG490 (Triptolide) Cooperates with Tumor Necrosis Factor-α to Induce Apoptosis in Tumor Cells. J. Biol. Chem. 1999, 274, 13451–13455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, W.T.; Kang, J.J.; Lee, K.Y.; Wei, K.; Anderson, E.; Gotmare, S.; Ross, J.A.; Rosen, G.D. Triptolide and Chemotherapy Cooperate in Tumor Cell Apoptosis. A Role for the P53 Pathway. J. Biol. Chem. 2001, 276, 2221–2227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rongzhen, X.; Hongjian, J. C14-Hydroxyl Esterified Amino Acid Derivatives of Triptolide, and Preparation Method and Use Thereof. CN Patent. EP3248981A1. Available online: https://patents.google.com/patent/EP3248981A1/en (accessed on 13 April 2022).
- Namin, M.H.; Ebrahimzadeh, H.; Ghareyazie, B.; Radjabian, T.; Gharavi, S.; Tafreshi, N.; Lee, H. In Vitro Expression of Apocarotenoid Genes in Crocus Sativus L. Afr. J. Biotechnol. 2009, 8, 5378–5382. [Google Scholar]
- Wyss, A. Carotene Oxygenases: A New Family of Double Bond Cleavage Enzymes. J. Nutr. 2004, 134, 246S–250S. [Google Scholar] [CrossRef] [Green Version]
- Bolhassani, A.; Khavari, A.; Bathaie, S.Z. Saffron and Natural Carotenoids: Biochemical Activities and Anti-Tumor Effects. Biochim. Et Biophys. Acta-Rev. Cancer 2014, 1845, 20–30. [Google Scholar] [CrossRef]
- Abar, L.; Vieira, A.R.; Aune, D.; Stevens, C.; Vingeliene, S.; Navarro Rosenblatt, D.A.; Chan, D.; Greenwood, D.C.; Norat, T. Blood Concentrations of Carotenoids and Retinol and Lung Cancer Risk: An Update of the WCRF–AICR Systematic Review of Published Prospective Studies. Cancer Med. 2016, 5, 2069–2083. [Google Scholar] [CrossRef] [Green Version]
- Rakic, J.M.; Liu, C.; Veeramachaneni, S.; Wu, D.; Paul, L.; Chen, C.Y.O.; Ausman, L.M.; Wang, X.D. Lycopene Inhibits Smoke-Induced Chronic Obstructive Pulmonary Disease and Lung Carcinogenesis by Modulating Reverse Cholesterol Transport in Ferrets. Cancer Prev. Res. 2019, 12, 421–432. [Google Scholar] [CrossRef]
- Eid, S.Y.; El-Readi, M.Z.; Wink, M. Carotenoids Reverse Multidrug Resistance in Cancer Cells by Interfering with ABC-Transporters. Phytomedicine 2012, 19, 977–987. [Google Scholar] [CrossRef] [PubMed]
- Awad, A.B.; Chen, Y.C.; Fink, C.S.; Hennessey, T. Beta-Sitosterol Inhibits HT-29 Human Colon Cancer Cell Growth and Alters Membrane Lipids. Anticancer Res. 1996, 16, 2797–2804. [Google Scholar] [PubMed]
- Bouic, P.J.D. The Role of Phytosterols and Phytosterolins in Immune Modulation: A Review of the Past 10 Years. Curr. Opin. Clin. Nutr. Metab. Care 2001, 4, 471–475. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.J. Effect of Olive Oil Minor Components on Oxidative Stress and Arachidonic Acid Mobilization and Metabolism by Macrophages RAW 264.7. Free. Radic. Biol. Med. 2003, 35, 1073–1081. [Google Scholar] [CrossRef]
- Awad, A.B.; Toczek, J.; Fink, C.S. Phytosterols Decrease Prostaglandin Release in Cultured P388D 1/MAB Macrophages. Prostaglandins Leukot. Essent. Fat. Acids 2004, 70, 511–520. [Google Scholar] [CrossRef]
- Ikemoto, S.; Sugimura, K.; Yoshida, N.; Yasumoto, R.; Wada, S.; Yamamoto, K.; Kishimoto, T. Antitumor Effects of Scutellariae Radix and Its Components Baicalein, Baicalin, and Wogonin on Bladder Cancer Cell Lines. Urology 2000, 55, 951–955. [Google Scholar] [CrossRef]
- Kaplya, O.A.; Sherstoboev, E.Y.; Zueva, E.P.; Razina, T.G.; Amosova, E.N.; Krylova, S.G. Effect of Baikal Skullcap Extract Administered Alone or in Combination with Cyclophosphamide on Natural Cytotoxicity System in Mice with Lewis Lung Carcinoma. Bull. Exp. Biol. Med. 2004, 137, 471–474. [Google Scholar] [CrossRef]
- Li-Weber, M. New Therapeutic Aspects of Flavones: The Anticancer Properties of Scutellaria and Its Main Active Constituents Wogonin, Baicalein and Baicalin. Cancer Treat. Rev. 2009, 35, 57–68. [Google Scholar] [CrossRef]
- Lamer-Zarawska, E.; Wiśniewska, A.; Błach-Olszewska, Z. Anticancer Properties of Scutellaria Baicalensis Root in Aspect of Innate Immunity Regulation Przeciwnowotworowa Aktywność Tarczycy Bajkalskiej w Świetle Regulacji Wrodzonej Odporności. Adv. Clin. Exp. Med. 2010, 19, 419–428. [Google Scholar]
- Ma, Z.; Otsuyama, K.-I.; Liu, S.; Abroun, S.; Ishikawa, H.; Tsuyama, N.; Obata, M.; Li, F.-J.; Zheng, X.; Maki, Y.; et al. Baicalein, a Component of Scutellaria Radix from Huang-Lian-Jie-Du-Tang (HLJDT), Leads to Suppression of Proliferation and Induction of Apoptosis in Human Myeloma Cells. Blood 2005, 105, 3312–3318. [Google Scholar] [CrossRef] [Green Version]
- Bonham, M.; Posakony, J.; Coleman, I.; Montgomery, B.; Simon, J.; Nelson, P.S. Characterization of Chemical Constituents in Scutellaria Baicalensis with Antiandrogenic and Growth-Inhibitory Activities toward Prostate Carcinoma. Clin. Cancer Res. 2005, 11, 3905–3914. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumagai, T.; Müller, C.I.; Desmond, J.C.; Imai, Y.; Heber, D.; Koeffler, H.P. Scutellaria Baicalensis, a Herbal Medicine: Anti-Proliferative and Apoptotic Activity against Acute Lymphocytic Leukemia, Lymphoma and Myeloma Cell Lines. Leuk. Res. 2007, 31, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Chi, Y.S.; Lim, H.; Park, H.; Kim, H.P. Effects of Wogonin, a Plant Flavone from Scutellaria Radix, on Skin Inflammation: In Vivo Regulation of Inflammation-Associated Gene Expression. Biochem. Pharmacol. 2003, 66, 1271–1278. [Google Scholar] [CrossRef]
- Huang, W.H.; Lee, A.R.; Yang, C.H. Antioxidative and Anti-Inflammatory Activities of Polyhydroxyflavonoids of Scutellaria Baicalensis GEORGI. Biosci. Biotechnol. Biochem. 2006, 70, 2371–2380. [Google Scholar] [CrossRef] [Green Version]
- Lim, H.; Park, H.; Kim, H.P. Inhibition of Contact Dermatitis in Animal Models and Suppression of Proinflammatory Gene Expression by Topically Applied Flavonoid, Wogonin. Arch. Pharm. Res. 2004, 27, 442. [Google Scholar] [CrossRef]
- Błach-Olszewska, Z.; Jatczak, B.; Rak, A.; Lorenc, M.; Gulanowski, B.; Drobna, A.; Lamer-Zarawska, E. Production of Cytokines and Stimulation of Resistance to Viral Infection in Human Leukocytes by Scutellaria Baicalensis Flavones. J. Interferon Cytokine Res. 2008, 28, 571–581. [Google Scholar] [CrossRef]
- Chen, Y.C.; Shen, S.C.; Chen, L.G.; Lee, T.J.; Yang, L.L. Wogonin, Baicalin, and Baicalein Inhibition of Inducible Nitric Oxide Synthase and Cyclooxygenase-2 Gene Expressions Induced by Nitric Oxide Synthase Inhibitors and Lipopolysaccharide. Biochem. Pharmacol. 2001, 61, 1417–1427. [Google Scholar] [CrossRef]
- Chen, L.G.; Hung, L.Y.; Tsai, K.W.; Pan, Y.S.; da Tsai, Y.; Li, Y.Z.; Liu, Y.W. Wogonin, a Bioactive Flavonoid in Herbal Tea, Inhibits Inflammatory Cyclooxygenase-2 Gene Expression in Human Lung Epithelial Cancer Cells. Mol. Nutr. Food Res. 2008, 52, 1349–1357. [Google Scholar] [CrossRef]
- Kim, H.; Kim, Y.S.; Kim, S.Y.; Suk, K. The Plant flavonoid Wogonin Suppresses Death of Activated C6 Rat Glial Cells by Inhibiting Nitric Oxide Production. Neurosci. Lett. 2001, 309, 67–71. [Google Scholar] [CrossRef]
- Fukumura, D.; Kashiwagi, S.; Jain, R.K. The Role of Nitric Oxide in Tumour Progression. Nat. Rev. Cancer 2006, 6, 521–534. [Google Scholar] [CrossRef]
- Ke, M.; Zhang, Z.; Xu, B.; Zhao, S.; Ding, Y.; Wu, X.; Wu, R.; Lv, Y.; Dong, J. Baicalein and Baicalin Promote Antitumor Immunity by Suppressing PD-L1 Expression in Hepatocellular Carcinoma Cells. Int. Immunopharmacol. 2019, 75, 105824. [Google Scholar] [CrossRef]
- Anila, L.; Vijayalakshmi, N.R. Antioxidant Action of Flavonoids from Mangifera Indica and Emblica Officinalis in Hypercholesterolemic Rats. Food Chem. 2003, 83, 569–574. [Google Scholar] [CrossRef]
- Abesundara, K.J.M.; Matsui, T.; Matsumoto, K. α-Glucosidase Inhibitory Activity of Some Sri Lanka Plant Extracts, One of Which, Cassia Auriculata, Exerts a Strong Antihyperglycemic Effect in Rats Comparable to the Therapeutic Drug Acarbose. J. Agric. Food Chem. 2004, 52, 2541–2545. [Google Scholar] [CrossRef] [PubMed]
- Rani, P.; Khullar, N. Antimicrobial Evaluation of Some Medicinal Plants for Their Anti-Enteric Potential against Multi-Drug Resistant Salmonella Typhi. Phytother. Res. 2004, 18, 670–673. [Google Scholar] [CrossRef] [PubMed]
- Ram, M.S.; Neetu, D.; Yogesh, B.; Anju, B.; Dipti, P.; Pauline, T.; Sharma, S.K.; Sarada, S.K.S.; Ilavazhagan, G.; Kumar, D.; et al. Cyto-Protective and Immunomodulating Properties of Amla (Emblica Officinalis) on Lymphocytes: An in-Vitro Study. J. Ethnopharmacol. 2002, 81, 5–10. [Google Scholar] [CrossRef]
- Jose, J.K.; Kuttan, G.; Kuttan, R. Antitumour Activity of Emblica Officinalis. J. Ethnopharmacol. 2001, 75, 65–69. [Google Scholar] [CrossRef]
- Rajeshkumar, N.V.; Joy, K.L.; Kuttan, G.; Ramsewak, R.S.; Nair, M.G.; Kuttan, R. Antitumour and Anticarcinogenic Activity of Phyllanthus Amarus Extract. J. Ethnopharmacol. 2002, 81, 17–22. [Google Scholar] [CrossRef]
- Liu, X.; Zhao, M.; Wu, K.; Chai, X.; Yu, H.; Tao, Z.; Wang, J. Immunomodulatory and Anticancer Activities of Phenolics from Emblica Fruit (Phyllanthus Emblica L.). Food Chem. 2012, 131, 685–690. [Google Scholar] [CrossRef]
- Kawakami, K.; Nishida, H.; Tatewaki, N.; Eguchi-Kasai, K.; Anzai, K.; Eitsuka, T.; Konishi, T.; Hirayama, M. Persimmon Leaf Flavonols Enhance the Anti-Cancer Effect of Heavy Ion Radiotherapy on Murine Xenograft Tumors. J. Cancer Ther. 2013, 4, 1150–1157. [Google Scholar] [CrossRef] [Green Version]
- Ling, W.D.; Du, G. Effect of Flavonoid from Diospyros Kaki Leaves on TGF- β 1 and MMP-9 in Blood Glucose Levels and Kidney Tissues of Rats with Diabetic Nephropathy. Chin. J. Exp. Tradit. Med. Form 2016, 22, 139–143. [Google Scholar]
- Sun, L.; Zhang, J.; Lu, X.; Zhang, L.; Zhang, Y. Evaluation to the Antioxidant Activity of Total Flavonoids Extract from Persimmon (Diospyros Kaki L.) Leaves. Food Chem. Toxicol. 2011, 49, 2689–2696. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Wei, Y.; Zhao, S.; Zhang, M.; Yan, X.; Gao, X.; Li, J.; Gao, Y.; Zhang, A.; Gao, Y. Antitumor and Immunomodulatory Activities of Total Flavonoids Extract from Persimmon Leaves in H22 Liver Tumor-Bearing Mice. Sci. Rep. 2018, 8, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ganeshpurkar, A.; Saluja, A.K. The Pharmacological Potential of Rutin. Saudi Pharm. J. 2017, 25, 149–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, R.; Zhang, Y.; Ye, X.; Xue, S.; Shi, J.; Pan, J.; Chen, Q. Inhibition Effects and Induction of Apoptosis of Flavonoids on the Prostate Cancer Cell Line PC-3 in Vitro. Food Chem. 2013, 138, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Hayat, K.; Iqbal, H.; Malik, U.; Bilal, U.; Mushtaq, S. Tea and Its Consumption: Benefits and Risks. Crit. Rev. Food Sci. Nutr. 2015, 55, 939–954. [Google Scholar] [CrossRef]
- Crespy, V.; Williamson, G. A Review of the Health Effects of Green Tea Catechins in In Vivo Animal Models. J. Nutr. 2004, 134, 3431S–3440S. [Google Scholar] [CrossRef]
- Katiyar, S.; Elmets, C.A.; Katiyar, S.K. Green Tea and Skin Cancer: Photoimmunology, Angiogenesis and DNA Repair. J. Nutr. Biochem. 2007, 18, 287–296. [Google Scholar] [CrossRef]
- Shirakami, Y.; Shimizu, M.; Moriwaki, H. Cancer Chemoprevention with Green Tea Catechins: From Bench to Bed. Current Drug Targets 2012, 13, 1842–1857. [Google Scholar] [CrossRef]
- Butt, M.S.; Sultan, M.T. Green Tea: Nature’s Defense against Malignancies. Crit. Rev. Food Sci. Nutr. 2009, 49, 463–473. [Google Scholar] [CrossRef]
- Fujiki, H.; Watanabe, T.; Sueoka, E.; Rawangkan, A.; Suganuma, M. Cancer Prevention with Green Tea and Its Principal Constituent, EGCG: From Early Investigations to Current Focus on Human Cancer Stem Cells. Mol. Cells 2018, 41, 73–82. [Google Scholar]
- Negri, A.; Naponelli, V.; Rizzi, F.; Bettuzzi, S. Molecular Targets of Epigallocatechin—Gallate (EGCG): A Special Focus on Signal Transduction and Cancer. Nutrients 2018, 10, 1936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santilli, G.; Piotrowska, I.; Cantilena, S.; Chayka, O.; D’Alicarnasso, M.; Morgenstern, D.A.; Himoudi, N.; Pearson, K.; Anderson, J.; Thrasher, A.J.; et al. Polyphenol e Enhances the Antitumor Immune Response in Neuroblastoma by Inactivating Myeloid Suppressor Cells. Clin. Cancer Res. 2013, 19, 1116–1125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tae, H.K.; Jin, H.L.; Chung, K.S.; Hee, D.H.; Byung, C.S.; Pai, S.I.; Hung, C.F.; Trimble, C.; Lim, J.S.; Tae, W.K.; et al. Epigallocatechin-3-Gallate Enhances CD8+ T Cell-Mediated Antitumor Immunity Induced by DNA Vaccination. Cancer Res. 2007, 67, 802–811. [Google Scholar] [CrossRef] [Green Version]
- Mantena, S.K.; Roy, A.M.; Katiyar, S.K. Epigallocatechin-3-Gallate Inhibits Photocarcinogenesis Through Inhibition of Angiogenic Factors and Activation of CD8+ T Cells in Tumors. Photochem. Photobiol. 2005, 81, 1174. [Google Scholar] [CrossRef]
- Hsieh, D.S.; Wang, H.; Tan, S.W.; Huang, Y.H.; Tsai, C.Y.; Yeh, M.K.; Wu, C.J. The Treatment of Bladder Cancer in a Mouse Model by Epigallocatechin-3-Gallate-Gold Nanoparticles. Biomaterials 2011, 32, 7633–7640. [Google Scholar] [CrossRef]
- Rawangkan, A.; Wongsirisin, P.; Namiki, K.; Iida, K.; Kobayashi, Y.; Shimizu, Y.; Fujiki, H.; Suganuma, M. Green Tea Catechin Is an Alternative Immune Checkpoint Inhibitor That Inhibits PD-L1 Expression and Lung Tumor Growth. Molecules 2018, 23, 2071. [Google Scholar] [CrossRef] [Green Version]
- Belguendouz, L.; Frémont, L.; Gozzelino, M.-T. Interaction of Transresveratrol with Plasma Lipoproteins. Biochem. Pharmacol. 1998, 55, 811–816. [Google Scholar] [CrossRef]
- Kisková, T.; Kassayová, M. Resveratrol Action on Lipid Metabolism in Cancer. Int. J. Mol. Sci. 2019, 20, 2704. [Google Scholar] [CrossRef] [Green Version]
- Pace-Asciak, C.R.; Hahn, S.; Diamandis, E.P.; Soleas, G.; Goldberg, D.M. The Red Wine Phenolics Trans-Resveratrol and Quercetin Block Human Platelet Aggregation and Eicosanoid Synthesis: Implications for Protection against Coronary Heart Disease. Clin. Chim. Acta 1995, 235, 207–219. [Google Scholar] [CrossRef]
- Rotondo, S.; Rajtar, G.; Manarini, S.; Celardo, A.; Rotilio, D.; de Gaetano, G.; Evangelista, V.; Cerletti, C. Effect of Trans-Resveratrol, a Natural Polyphenolic Compound, on Human Polymorphonuclear Leukocyte Function. Br. J. Pharmacol. 1998, 123, 1691–1699. [Google Scholar] [CrossRef] [Green Version]
- Kimura, Y.; Okuda, H.; Kubo, M. Effects of Stilbenes Isolated from Medicinal Plants on Arachidonate Metabolism and Degranulation in Human Polymorphonuclear Leukocytes. J. Ethnopharmacol. 1995, 45, 131–139. [Google Scholar] [CrossRef]
- Belguendouz, L.; Fremont, L.; Hard, A. Resveratrol Inhibits Metal Ion-Dependent and Independent Peroxidation of Porcine Low-Density Lipoproteins. Biochem Pharmacol. 1997, 53, 1347–1355. [Google Scholar] [CrossRef]
- Man-Ying Chan, M.; Mattiacci, J.A.; Hwang, H.S.; Shah, A.; Fong, D. Synergy between Ethanol and Grape Polyphenols, Quercetin, and Resveratrol, in the Inhibition of the Inducible Nitric Oxide Synthase Pathway. Biochem. Pharmacol. 2000, 60, 1539–1548. [Google Scholar] [CrossRef]
- Jang, M.; Cai, L.; Udeani, G.O.; Slowing, K.V.; Thomas, C.F.; Beecher, C.W.W.; Fong, H.H.S.; Farnsworth, N.R.; Kinghorn, A.D.; Mehta, R.G.; et al. Cancer Chemopreventive Activity of Resveratrol, a Natural Product Derived from Grapes. Science 1997, 275, 218–220. [Google Scholar] [CrossRef] [Green Version]
- Mgbonyebi, O.P.; Russo, J.; Russo, I.H. Antiproliferative Effect of Synthetic Resveratrol on Human Breast Epithelial Cells. Int. J. Oncol. 1998, 12, 865–874. [Google Scholar] [CrossRef]
- Lu, R.; Serrero, G. Resveratrol, a Natural Product Derived from Grape, Exhibits Antiestrogenic Activity and Inhibits the Growth of Human Breast Cancer Cells. J. Cell Physiol. 1999, 179, 297–304. [Google Scholar] [CrossRef]
- Hsieh, T.-C.; Wu, J.M. Differential Effects on Growth, Cell Cycle Arrest, and Induction of Apoptosis by Resveratrol in Human Prostate Cancer Cell Lines. Exp Cell Res. 1999, 249, 109–115. [Google Scholar] [CrossRef]
- Schneider, Y.; Vincent, F.; Duranton, B.Ã.; Badolo, L.; Gosse, F.; Bergmann, C.; Seiler, N.; Raul, F. Anti-Proliferative Effect of Resveratrol, a Natural Component of Grapes and Wine, on Human Colonic Cancer Cells. Cancer Lett. 2000, 158, 85–91. [Google Scholar] [CrossRef]
- Gao, X.; Xu, Y.X.; Janakiraman, N.; Chapman, R.A.; Gautam, S.C. Immunomodulatory Activity of Resveratrol: Suppression of Lymphocyte Proliferation, Development of Cell-Mediated Cytotoxicity, and Cytokine Production. Biochem. Pharmacol. 2001, 62, 1299–1308. [Google Scholar] [CrossRef]
- Bergman, M.; Levin, G.S.; Bessler, H.; Djaldetti, M.; Salman, H. Resveratrol Affects the Cross Talk between Immune and Colon Cancer Cells. Biomed. Pharmacother. 2013, 67, 43–47. [Google Scholar] [CrossRef]
- Majdalawieh, A.F.; Carr, R.I. In Vitro Investigation of the Potential Immunomodulatory and Anti-Cancer Activities of Black Pepper (Piper Nigrum) and Cardamom (Elettaria Cardamomum). J. Med. Food 2010, 13, 371–381. [Google Scholar] [CrossRef] [PubMed]
- Surh, Y.-J. Anti-Tumor Promoting Potential of Selected Spice Ingredients with Antioxidative and Anti-Inflammatory Activities: A Short Review. Food Chem. Toxicol. 2002, 40, 1091–1097. [Google Scholar] [CrossRef]
- Kaefer, C.M.; Milner, J.A. The Role of Herbs and Spices in Cancer Prevention. J. Nutr. Biochem. 2008, 19, 347–361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pathak, N.; Khandelwal, S. Cytoprotective and Immunomodulating Properties of Piperine on Murine Splenocytes: An in Vitro Study. Eur. J. Pharmacol. 2007, 576, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Duessel, S.; Heuertz, R.M.; Ezekiel, U.R. Growth Inhibition of Human Colon Cancer Cells by Plant Compounds. Am. Soc. Clin. Lab. Sci. 2008, 21, 151–157. [Google Scholar]
- Menon, L.G.; Kuttan, R.; Kuttan, G. Effect of Rasayanas in the Inhibition of Lung Metastasis Induced by B16F-10 Melanoma Cells. J. Exp. Clin. Cancer Res. 1997, 16, 365–368. [Google Scholar]
- Selvendiran, K.; Mumtaz Banu, S.; Sakthisekaran, D. Oral Supplementation of Piperine Leads to Altered Phase II Enzymes and Reduced DNA Damage and DNA-Protein Cross Links in Benzo(a)Pyrene Induced Experimental Lung Carcinogenesis. Mol Cell Biochem. 2005, 268, 141–147. [Google Scholar] [CrossRef]
- Selvendiran, K.; Thirunavukkarasu, C.; Prince, J.; Singh, V.; Padmavathi, R.; Sakthisekaran, D. Chemopreventive Effect of Piperine on Mitochondrial TCA Cycle and Phase-I and Glutathione-Metabolizing Enzymes in Benzo(a)Pyrene Induced Lung Carcinogenesis in Swiss Albino Mice. Mol. Cell Biochem. 2005, 271, 101–106. [Google Scholar] [CrossRef]
- Krishnakumar, N.; Manoharan, S.; Palaniappan, P.R.; Venkatachalam, P.; Manohar, M.G.A. Chemopreventive Efficacy of Piperine in 7,12-Dimethyl Benz [a] Anthracene (DMBA)-Induced Hamster Buccal Pouch Carcinogenesis: An FT-IR Study. Food Chem. Toxicol. 2009, 47, 2813–2820. [Google Scholar] [CrossRef]
- Manoharan, S.; Balakrishnan, S.; Menon, V.P.; Alias, L.M.; Reena, A.R. Chemopreventive Efficacy of Curcumin and Piperine during 7,12-Dimethylbenz [a]Anthracene-Induced Hamster Buccal Pouch Carcinogenesis. Singapore Med. J. 2009, 50, 139. [Google Scholar]
- Block, R.M.; Lewis, R.D.; Shea@, J.B.; Fawley, J.; Richmond, V. Cell-Mediated Immune Response to Dog Pulp Tissue Altered by Eugenol within the Root Canal. Oral Surg. Oral Med. Oral Pathol. 1978, 45, 452–463. [Google Scholar] [CrossRef]
- Van Duuren, B.L.; Sivak, A.; Segal, A.; Orris, L.; Langseth, L. The Tumor-Promoting Agents of Tobacco Leaf and Tobacco Smoke Condensate. J Natl Cancer Inst. 1966, 37, 519–526. [Google Scholar] [PubMed]
- Van Duuren, B.L.; Goldschmidt, B.M. Cocarcinogenic and Tumor-Promoting Agents in Tobacco Carcinogenesis. J. Natl. Cancer Inst. 1976, 56, 1237–1242. [Google Scholar] [CrossRef] [PubMed]
- Sukumaran, K.; Unnikrishnan, M.C.; Kuttan, R. Inhjbition of Tumour Promotion in Mice by Eugenol. Indian J. Physiol. Pharmacol. 1994, 38, 306–308. [Google Scholar]
- Ghosh, R.; Nadiminty, N.; Fitzpatrick, J.E.; Alworth, W.L.; Slaga, T.J.; Kumar, A.P. Eugenol Causes Melanoma Growth Suppression through Inhibition of E2F1 Transcriptional Activity. J. Biol. Chem. 2005, 280, 5812–5819. [Google Scholar] [CrossRef] [Green Version]
- Edwards, R.L.; Luis, P.B.; Varuzza, P.V.; Joseph, A.I.; Presley, S.H.; Chaturvedi, R.; Schneider, C. The Anti-Inflammatory Activity of Curcumin Is Mediated by Its Oxidative Metabolites. J. Biol. Chem. 2017, 292, 21243–21252. [Google Scholar] [CrossRef] [Green Version]
- Galet, C.; Gollapudi, K.; Stepanian, S.; Byrd, J.B.; Henning, S.M.; Grogan, T.; Elashoff, D.; Heber, D.; Said, J.; Cohen, P.; et al. Effect of a Low-Fat Fish Oil Diet on Proinflammatory Eicosanoids and Cell-Cycle Progression Score in Men Undergoing Radical Prostatectomy. Cancer Prev. Res. 2014, 7, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Panahi, Y.; Darvishi, B.; Ghanei, M.; Jowzi, N.; Beiraghdar, F.; Varnamkhasti, B.S. Molecular Mechanisms of Curcumins Suppressing Effects on Tumorigenesis, Angiogenesis and Metastasis, Focusing on NF-ΚB Pathway. Cytokine Growth Factor Rev. 2016, 28, 21–29. [Google Scholar] [CrossRef]
- Wilken, R.; Veena, M.S.; Wang, M.B.; Srivatsan, E.S. Curcumin: A Review of Anti-Cancer Properties and Therapeutic Activity in Head and Neck Squamous Cell Carcinoma. Mol. Cancer 2011, 10, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Chandra, D.; Jahangir, A.; Cornelis, F.; Rombauts, K.; Meheus, L.; Jorcyk, C.L.; Gravekamp, C. Cryoablation and Meriva Have Strong Therapeutic Effect on Triple-Negative Breast Cancer. OncoImmunology 2016, 5, e1049802. [Google Scholar] [CrossRef] [Green Version]
- Nahar, P.P.; Slitt, A.L.; Seeram, N.P. Anti-Inflammatory Effects of Novel Standardized Solid Lipid Curcumin Formulations. J. Med. Food 2015, 18, 786–792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, Y.I.; Kim, S.W.; Jung, I.D.; Lee, J.S.; Chang, J.H.; Lee, C.M.; Chun, S.H.; Yoon, M.S.; Kim, G.T.; Ryu, S.W.; et al. Curcumin Suppresses the Induction of Indoleamine 2,3-Dioxygenase by Blocking the Janus-Activated Kinase-Protein Kinase Cδ-STAT1 Signaling Pathway in Interferon-γ-Stimulated Murine Dendritic Cells. J. Biol. Chem. 2009, 284, 3700–3708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lim, S.O.; Li, C.W.; Xia, W.; Cha, J.H.; Chan, L.C.; Wu, Y.; Chang, S.S.; Lin, W.C.; Hsu, J.M.; Hsu, Y.H.; et al. Deubiquitination and Stabilization of PD-L1 by CSN5. Cancer Cell 2016, 30, 925–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ireson, C.R.; Jones, D.J.L.; Orr, S.; Coughtrie, M.W.H.; Boocock, D.J.; Williams, M.L.; Farmer, P.B.; Steward, W.P.; Gescher, A.J. Metabolism of the Cancer Chemopreventive Agent Curcumin in Human and Rat Intestine. Cancer Epidemiol. Prev. Biomark. 2002, 11, 105–111. [Google Scholar]
- Moradali, M.F.; Mostafavi, H.; Ghods, S.; Hedjaroude, G.A. Immunomodulating and Anticancer Agents in the Realm of Macromycetes Fungi (Macrofungi). Int. Immunopharmacol. 2007, 7, 701–724. [Google Scholar] [CrossRef]
- Wasser, S. Medicinal Mushrooms as a Source of Antitumor and Immunomodulating Polysaccharides. Appl. Microbiol. Biotechnol. 2002, 60, 258–274. [Google Scholar]
- Wachtel-Galor, S.; Yuen, J.; Buswell, J.A.; Benzie, I.F.F. Ganoderma lucidum (Lingzhi or Reishi): A Medicinal Mushroom. In Herbal Medicine: Biomolecular and Clinical Aspects, 2nd ed.; Benzie, I.F.F., Wachtel-Galor, S., Eds.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2011. Available online: https://www.ncbi.nlm.nih.gov/books/NBK92757/ (accessed on 13 April 2022)Chapter 9.
- Gao, Y.; Zhou, S.H.; Chen, G.; Dai, X.; Ye, J. A Phase III Study of a Ganoderma Lucidum (Curt. Fr.) P. Karst. Extract (Ganopoly) in Patients with Advanced Cancer. Int. J. Med. Mushroom 2002, 4, 207–214. [Google Scholar]
- Burdock, G.A. Review of the Biological Properties and Toxicity of Bee Propolis (Propolis). Food Chem. Toxicol. 1998, 36, 347–363. [Google Scholar] [CrossRef]
- Castaldo, S.; Capasso, F. Propolis, an Old Remedy Used in Modern Medicine. Fitoterapia 2002, 73 (Suppl. 1), S1–S6. [Google Scholar] [CrossRef]
- Borrelli, F.; Maffia, P.; Pinto, L.; Ianaro, A.; Russo, A.; Capasso, F.; Ialenti, A. Phytochemical Compounds Involved in the Anti-Inflammatory Effect of Propolis Extract. Fitoterapia 2002, 73 (Suppl. 1), S53–S63. [Google Scholar] [CrossRef]
- Oršolić, N.; Bašić, I. Immunomodulation by Water-Soluble Derivative of Propolis: A Factor of Antitumor Reactivity. J. Ethnopharmacol. 2003, 84, 265–273. [Google Scholar] [CrossRef]
- Marcucci, M.C.; Ferreres, F.; Custódio, A.R.; Ferreira, M.M.C.; Bankovad, V.S.; García-Viguera, C.; Bretz, W.A. Evalution of Phenolic Compounds in Brazilian Propolis from Different Geographic Regions. Z Naturforsch C J. Biosci. 2000, 55, 76–81. [Google Scholar] [CrossRef] [PubMed]
- Cole, N.; Sou, P.W.; Ngo, A.; Tsang, K.H.; Severino, J.A.J.; Arun, S.J.; Duke, C.C.; Reeve, V.E. Topical “sydney” Propolis Protects against UV-Radiation-Induced Inflammation, Lipid Peroxidation and Immune Suppression in Mouse Skin. Int. Arch. Allergy Immunol. 2010, 152, 87–97. [Google Scholar] [CrossRef]
- Bankova, V. Recent Trends and Important Developments in Propolis Research. Evid. -Based Complementary Altern. Med. 2005, 2, 29–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, W.L.; Liang, W.H.; Lee, Y.J.; Chuang, S.K.; Tseng, T.H. Antitumor Progression Potential of Caffeic Acid Phenethyl Ester Involving P75NTR in C6 Glioma Cells. Chem.-Biol. Interact. 2010, 188, 607–615. [Google Scholar] [CrossRef]
- Ang, E.S.M.; Pavlos, N.J.; Chai, L.Y.; Qi, M.; Cheng, T.S.; Steer, J.H.; Joyce, D.A.; Zheng, M.H.; Xu, J. Caffeic Acid Phenethyl Ester, an Active Component of Honeybee Propolis Attenuates Osteoclastogenesis and Bone Resorption via the Suppression of RANKL-Induced NF-ΚB and NFAT Activity. J. Cell. Physiol. 2009, 221, 642–649. [Google Scholar] [CrossRef]
- Park, J.H.; Lee, J.K.; Kim, H.S.; Chung, S.T.; Eom, J.H.; Kim, K.A.; Chung, S.J.; Paik, S.Y.; Oh, H.Y. Immunomodulatory Effect of Caffeic Acid Phenethyl Ester in Balb/c Mice. Int. Immunopharmacol. 2004, 4, 429–436. [Google Scholar] [CrossRef]
- Huang, M.-T.; Ma, W.; Yen, P.; Xie, J.-G.; Han, J.; Frenkel, K.; Grunberger, D.; Conney, H.-A. Inhibitory Effects of Caffeic Acid Phenethyl Ester (CAPE) on 12-0-Tetradecanoylphorbol-13-Acetate-Induced Tumor Promotion in Mouse Skin and the Synthesis of DNA, RNA and Protein in HeLa Cells. Carcinogenesis 1996, 17, 761–765. [Google Scholar] [CrossRef] [Green Version]
- Natarajan, K.; Singh, S.; Burke, T.R.; GRUNBERGERt, D.; Aggarwal, B.B. Caffeic Acid Phenethyl Ester Is a Potent and Specific Inhibitor of Activation of Nuclear Transcription Factor NF-KB (Tumor Necrosis Factor/Okadaic Acid/Ceramide/Phorbol Ester/Hydrogen Peroxide). Proc. Natl. Acad. Sci. USA 1996, 93, 9090–9095. [Google Scholar] [CrossRef] [Green Version]
- Roos, T.U.; Heiss, E.H.; Schwaiberger, A.V.; Schachner, D.; Sroka, I.M.; Oberan, T.; Vollmar, A.M.; Dirsch, V.M. Caffeic Acid Phenethyl Ester Inhibits PDGF-Induced Proliferation of Vascular Smooth Muscle Cells via Activation of P38 MAPK, HIF-1α, and Heme Oxygenase-1. J. Nat. Prod. 2011, 74, 352–356. [Google Scholar] [CrossRef]
- Basini, G.; Baioni, L.; Bussolati, S.; Grasselli, F.; Daquino, C.; Spatafora, C.; Tringali, C. Antiangiogenic Properties of an Unusual Benzo[k,l]Xanthene Lignan Derived from CAPE (Caffeic Acid Phenethyl Ester). Investig. New Drugs 2012, 30, 186–190. [Google Scholar] [CrossRef] [PubMed]
- Márquez, N.; Sancho, R.; Macho, A.; Calzado, M.A.; Fiebich, B.L.; Muñoz, E. Caffeic Acid Phenethyl Ester Inhibits T-Cell Activation by Targeting Both Nuclear Factor of Activated T-Cells and NF-ΚB Transcription Factors. J. Pharmacol. Exp. Ther. 2004, 308, 993–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, L.C.; Lin, Y.L.; Liang, Y.C.; Yang, Y.H.; Lee, J.H.; Yu, H.H.; Wu, W.M.; Chiang, B.L. The Effect of Caffeic Acid Phenethyl Ester on the Functions of Human Monocyte-Derived Dendritic Cells. BMC Immunol. 2009, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.C.; Chu, K.H.; Liang, Y.C.; Lin, Y.L.; Chiang, B.L. Caffeic Acid Phenethyl Ester Inhibits Nuclear Factor-ΚB and Protein Kinase B Signalling Pathways and Induces Caspase-3 Expression in Primary Human CD4+ T Cells. Clin. Exp. Immunol. 2010, 160, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Su, Z.-Z.; Lin, J.; Grunberger, D.; Fisher2, P.B. Growth Suppression and Toxicity Induced by Caffeic Acid Phenethyl Ester (CAPE) in Type 5 Adenovirus-Transformed Rat Embryo Cells Correlate Directly with Transformation Progression. Cancer Res 1994, 54, 1865–1870. [Google Scholar] [PubMed]
- Chena, J.-H.; Shaoa, Y.; Huang, M.; Chinb, C.-K.; Hoa, C.-T. Inhibitory Effect of Caffeic Acid Phenethyl Ester on Human Leukemia HL-60 Cells. Cancer Lett. 1996, 108, 211–214. [Google Scholar] [CrossRef]
- Lee, Y.-J.; Liao, P.-H.; Chen, W.-K.; Yang, C.-C. Preferential Cytotoxicity of Caffeic Acid Phenethyl Ester Analogues on Oral Cancer Cells. Cancer Lett. 2000, 153, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Paulino, N.; Abreu, S.R.L.; Uto, Y.; Koyama, D.; Nagasawa, H.; Hori, H.; Dirsch, V.M.; Vollmar, A.M.; Scremin, A.; Bretz, W.A. Anti-Inflammatory Effects of a Bioavailable Compound, Artepillin C, in Brazilian Propolis. Eur. J. Pharmacol. 2008, 587, 296–301. [Google Scholar] [CrossRef]
- von Holtz, R.L.; Fink, C.S.; Awad, A.B. β-Sitosterol Activates the Sphingomyelin Cycle and Induces Apoptosis in LNCaP Human Prostate Cancer Cells. Nutr. Cancer 1998, 32, 8–12. [Google Scholar] [CrossRef]
- Galasso, C.; Nuzzo, G.; Brunet, C.; Ianora, A.; Sardo, A.; Fontana, A.; Sansone, C. The Marine Dinoflagellate Alexandrium Minutum Activates a Mitophagic Pathway in Human Lung Cancer Cells. Mar. Drugs 2018, 16, 502. [Google Scholar] [CrossRef] [Green Version]
- D’Ippolito, G.; Cutignano, A.; Tucci, S.; Romano, G.; Cimino, G.; Fontana, A. Biosynthetic intermediates and stereochemical aspects of aldehyde biosynthesis in the marine diatom Thalassiosira rotula. Phytochemistry 2006, 67, 314–322. [Google Scholar] [CrossRef]
- Kim, S.K.; Thomas, N.V.; Li, X. Anticancer Compounds from Marine Macroalgae and Their Application as Medicinal Foods. In Advances in Food and Nutrition Research; Academic Press Inc.: Cambridge, MA, USA, 2011; Volume 64, pp. 213–224. [Google Scholar]
- Maruyama, H.; Tamauchi, H.; Iizuka, M.; Nakano, T.; Maruyamá, H. The Role of NK Cells in Antitumor Activity of Dietary Fucoidan from Undaria Pinnatifida Sporophylls (Mekabu). Planta Med. 2006, 72, 1415–1417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, G.; Sun, Y.P.; Xin, H.; Zhang, Y.; Li, Z.; Xu, Z. In Vivo Antitumor and Immunomodulation Activities of Different Molecular Weight Lambda-Carrageenans from Chondrus Ocellatus. Pharmacol. Res. 2004, 50, 47–53. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.; Wang, L.; Zhou, Y. Immunomodulation and Antitumor Activities of Different-Molecular-Weight Polysaccharides from Porphyridium Cruentum. Carbohydr. Polym. 2012, 87, 1206–1210. [Google Scholar] [CrossRef]
- Shi, W.; Lu, D.; Wu, C.; Li, M.; Ding, Z.; Li, Y.; Chen, B.; Lin, X.; Su, W.; Shao, X.; et al. Coibamide A Kills Cancer Cells through Inhibiting Autophagy. Biochem. Biophys. Res. Commun. 2021, 547, 52–58. [Google Scholar] [CrossRef]
- Kong, C.S.; Kim, J.A.; Yoon, N.Y.; Kim, S.K. Induction of Apoptosis by Phloroglucinol Derivative from Ecklonia Cava in MCF-7 Human Breast Cancer Cells. Food Chem. Toxicol. 2009, 47, 1653–1658. [Google Scholar] [CrossRef]
- Kurihara, H.; Koda, H.; Asami, S.; Kiso, Y.; Tanaka, T. Contribution of the Antioxidative Property of Astaxanthin to Its Protective Effect on the Promotion of Cancer Metastasis in Mice Treated with Restraint Stress. Life Sci. 2002, 70, 2509–2520. [Google Scholar] [CrossRef]
- Tanaka, T.; Morishita, Y.; Suzui, M.; Kojima, T.; Okumura, A.; Mori, H. Chemoprevention of Mouse Urinary Bladder Carcinogenesis by the Naturally Occurring Carotenoid Astaxanthin. Carcinogenesis 1994, 15, 15–19. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Shnimizu, M.; Moriwaki, H. Cancer Chemoprevention by Carotenoids. Molecules 2012, 17, 3202–3242. [Google Scholar] [CrossRef]
- Tanaka, T.; Makita, H.; Ohnishi, M.; Mori, H.; Satoh, K.; Hara, A. Chemoprevention of rat oral carcinogenesis by naturally occurring xanthophylls, astaxanthin and canthaxanthin. Cancer Res. 1995, 55, 4059–4064. [Google Scholar]
- Chang, Y.-J.; Huang, J.-R.; Tsai, Y.-C.; Hung, J.-T.; Wu, D.; Fujio, M.; Wong, C.-H.; Yu, A.L. Potent Immune-Modulating and Anticancer Effects of NKT Cell Stimulatory Glycolipids. Proc. Natl. Acad. Sci. USA 2007, 104, 10299–10304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bae, E.A.; Seo, H.; Kim, I.K.; Jeon, I.; Kang, C.Y. Roles of NKT Cells in Cancer Immunotherapy. Arch. Pharmacol. Res. 2019, 42, 543–548. [Google Scholar] [CrossRef] [PubMed]
- Manzo, E.; Cutignano, A.; Pagano, D.; Gallo, C.; Barra, G.; Nuzzo, G.; Sansone, C.; Ianora, A.; Urbanek, K.; Fenoglio, D.; et al. A New Marine-Derived Sulfoglycolipid Triggers Dendritic Cell Activation and Immune Adjuvant Response. Sci. Rep. 2017, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Gallo, C.; Ferrera, F.; Castiglia, D.; Fontana, A. Identication of Sulfavant A as the First Synthetic TREM2 Ligand Discloses a Homeostatic Response of Dendritic Cells After Receptor Engagement. Res. Sq. 2022. [Google Scholar] [CrossRef]
- Rinehart, K.L.; Gloer, J.B.; Cook, J.C.; Mizsak, S.A.; Scahill, T.A. Structures of the Didemnins, Antiviral and Cytotoxic Depsipeptides from a Caribbean Tunicate. J. Am. Chem. Soc. 1981, 103, 1857–1859. [Google Scholar] [CrossRef]
- Thell, K.; Hellinger, R.; Schabbauer, G.; Gruber, C.W. Immunosuppressive Peptides and Their Therapeutic Applications. Drug Discov. Today 2014, 19, 645–653. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.; Currano, J.N.; Carroll, P.J.; Joullié, M.M. Didemnins, Tamandarins and Related Natural Products. Nat. Prod. Rep. 2012, 29, 404–424. [Google Scholar] [CrossRef]
- Tsukimoto, M.; Nagaoka, M.; Shishido, Y.; Fujimoto, J.; Nishisaka, F.; Matsumoto, S.; Harunari, E.; Imada, C.; Matsuzaki, T. Bacterial Production of the Tunicate-Derived Antitumor Cyclic Depsipeptide Didemnin B. J. Nat. Prod. 2011, 74, 11–2329. [Google Scholar] [CrossRef]
- Xu, Y.; Kersten, R.D.; Nam, S.J.; Lu, L.; Al-Suwailem, A.M.; Zheng, H.; Fenical, W.; Dorrestein, P.C.; Moore, B.S.; Qian, P.Y. Bacterial biosynthesis and maturation of the didemnin anti-cancer agents. J. Am. Chem. Soc. 2012, 134, 8625–8632. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Q.T.; Liu, Z.D.; Wang, Z.; Wang, T.; Wang, N.; Wang, N.; Zhang, B.; Zhao, Y.F. Recent Advances in Small Peptides of Marine Origin in Cancer Therapy. Mar. Drugs 2021, 19, 115. [Google Scholar] [CrossRef]
- Delgado-Calle, J.; Kurihara, N.; Atkinson, E.G.; Nelson, J.; Miyagawa, K.; Galmarini, C.M.; Roodman, G.D.; Bellido, T. Aplidin (plitidepsin) is a novel anti-myeloma agent with potent anti-resorptive activity mediated by direct effects on osteoclasts. Oncotarget 2019, 10, 2709–2721. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- White, K.M.; Rosales, R.; Yildiz, S.; Kehrer, T.; Miorin, L.; Moreno, E.; Jangra, S.; Uccellini, M.B.; Rathnasinghe, R.; Coughlan, L.; et al. Plitidepsin has potent preclinical efficacy against SARS-CoV-2 by targeting the host protein eEF1A. Science 2021, 371, 926–931. [Google Scholar] [CrossRef] [PubMed]
- Cooper, E.L.; Yao, D. Diving for Drugs: Tunicate Anticancer Compounds. Drug Discov. Today 2012, 17, 636–648. [Google Scholar] [CrossRef] [PubMed]
- Oda, T.; Kamoshita, K.; Maruyama, S.; Masuda, K.; Nishimoto, M.; Xu, J.; Ukai, K.; Mangindaan, R.E.P.; Namikoshi, M. Cytotoxicity of Lissoclibadins and Lissoclinotoxins, Isolated from a Tropical Ascidian Lissoclinum Cf. Badium, against Human Solid-Tumor-Derived Cell Lines. Biol. Pharm. Bull. 2007, 30, 385–387. [Google Scholar] [CrossRef] [Green Version]
- Fedorov, S.N.; Radchenko, O.S.; Shubina, L.K.; Balaneva, N.N.; Bode, A.M.; Stonik, V.A.; Dong, Z. Evaluation of cancer-preventive activity and structure-activity relationships of 3-demethylubiquinone Q2, isolated from the ascidian Aplidium glabrum, and its synthetic analogs. Pharm. Res. 2006, 23, 70–81. [Google Scholar] [CrossRef] [Green Version]
- Nuzzo, G.; Gallo, C.; Crocetta, F.; Romano, L.; Barra, G.; Senese, G.; dell’Isola, M.; Carbone, D.; Tanduo, V.; Albiani, F.; et al. Identification of the Marine Alkaloid Lepadin A as Potential Inducer of Immunogenic Cell Death. Biomolecules 2022, 12, 246. [Google Scholar] [CrossRef]
- Journal, E.; Meenakshi, V.K.; Senthamarai, S.; Paripooranaselvi, M.; Gomathy, S.; Sankaravadivu, S.; Chamundeswari, K.P. Scholars Research Library In Vitro and in Vivo Antitumor and Immunomodulatory Studies of Microcosmus Exasperatus against DLA Bearing Mice. Eur. J. Appl. Eng. Sci. Res. 2013, 23, 18–25. Available online: http://scholarsresearchlibrary.com/archive.html (accessed on 13 April 2022).
- Pps APC, P. Antitumor and Immunomodulatory Activity of Phallusia Nigra Savigny, 1816 Against Ehrlich Ascites Carcinoma. Res. J. Pharm. Sci. 2012, 1, 7–12. [Google Scholar] [CrossRef]
- Janakiram, N.B.; Mohammed, A.; Rao, C.V. Sea Cucumbers Metabolites as Potent Anti-Cancer Agents. Mar. Drugs 2015, 13, 2909–2923. [Google Scholar] [CrossRef] [Green Version]
- Pettit, G.R.; Herald, C.L.; Doubek, D.L.; Herald, D.L.; Arnold, E.; Clardy, J. Isolation and structure of bryostatin-1. Am. Chem. Soc. 1982, 104, 24–6846. [Google Scholar] [CrossRef]
- Raghuvanshi, R.; Bharate, S.B. Preclinical and Clinical Studies on Bryostatins, A Class of Marine-Derived Protein Kinase C Modulators: A Mini-Review. Curr Top Med Chem. 2020, 20, 1124–1135. [Google Scholar] [CrossRef] [PubMed]
- Trenn, G.; Pettit, G.R.; Takayama, H.; Hu-Li, J.; Sitkovsky, M.V. Immunomodulating Properties of a Novel Series of Protein Kinase C Activators. The Bryostatins. J. Immunol. 1988, 140, 433–439. [Google Scholar] [PubMed]
- Hornung, R.L.; Pearson, J.W.; Beckwith, M.; Longo, D.L. Preclinical Evaluation of Bryostatin as an Anticancer Agent against Several Murine Tumor Cell Lines in Vitro versus in Vivo Activity. Cancer Res. 1992, 52, 101–107. [Google Scholar] [PubMed]
- Mary Varterasian, B.L.; Mohammad, R.M.; Eilender, D.S.; Hulburd, K.; Rodriguez, D.H.; Pemberton, P.A.; Pluda, J.M.; Dan, M.D.; Chen, B.D.; Al-Katib, A.M. Phase I Study of Bryostatin 1 in Patients with Relapsed. Non-Hodgkin’s Lymphoma and Chronic Lymphocytic Leukemia. J. Clin. Oncol. 1998, 16, 56–62. [Google Scholar] [CrossRef]
- Rinehart, K.L.; Holt, T.G.; Fregeau, N.L.; Stroh, J.G.; Keifer, P.A.; Sun, F.; Li, L.H.; Martin, D.G. Ecteinascidins 729, 743, 745, 759A, 759B, and 770: Potent antitumor agents from the Caribbean tunicate Ecteinascidia turbinata. J. Org. Chem. 1990, 55, 4512–4515. [Google Scholar] [CrossRef]
- Larsen, A.K.; Galmarini, C.M.; D’Incalci, M. Unique features of Trabectedin mechanism of action. Cancer Chemother. Pharmacol. 2016, 77, 663–671. [Google Scholar] [CrossRef] [PubMed]
- Belgiovine, C.; Frapolli, R.; Liguori, M.; Digifico, E.; Colombo, F.S.; Meroni, M.; Allavena, P.; D’Incalci, M. Inhibition of tumor-associated macrophages by trabectedin improves the antitumor adaptive immunity in response to anti-PD-1 therapy. Eur. J. Immunol. 2021, 51, 2677–2686. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, P.; Zhang, R.; Ivan, C.; Galletti, G.; Clise-Dwyer, K.; Barbaglio, F.; Scarfò, L.; Aracil, M.; Klein, C.; Wierda, W.; et al. Trabectedin Reveals a Strategy of Immunomodulation in Chronic Lymphocytic Leukemia. Cancer Immunol. Res. 2019, 7, 2036–2051. [Google Scholar] [CrossRef]
- D’Ippolito, G.; Iadicicco, O.; Romano, G. Fontana A Detection of short-chain aldehydes in marine organisms: The diatom Thalassiosira rotula. Tetrahedron. Lett. 2002, 43, 6137–6140. [Google Scholar] [CrossRef]
- Cutignano, A.; D’Ippolito, G.; Romano, G.; Lamari, N.; Cimino, G.; Febbraio, F.; Nucci, R.; Fontana, A. Chloroplastic Glycolipids Fuel Aldehyde Biosynthesis in the Marine Diatom Thalassiosira rotula. ChemBioChem 2006, 7, 450–456. [Google Scholar] [CrossRef]
- D’Ippolito, G.; Romano, G.; Caruso, T.; Spinella, A.; Cimino, G.; Fontana, A. Production of octadienal in the marine diatom Skeletonema costatum. Org. Lett. 2003, 5, 885–887. [Google Scholar] [CrossRef] [PubMed]
- D’Ippolito, G.; Tucci, S.; Cutignano, A.; Giovanna, R.; Cimino, G.; Miralto, A.; Fontana, A. The role of complex lipids in the synthesis of bioactive aldehydes of the marine diatom Skeletonema costatum. Biochim. Biophys Acta–Mol. Cell Biol. Lipids 2004, 1686, 100–107. [Google Scholar] [CrossRef] [PubMed]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nuzzo, G.; Senese, G.; Gallo, C.; Albiani, F.; Romano, L.; d’Ippolito, G.; Manzo, E.; Fontana, A. Antitumor Potential of Immunomodulatory Natural Products. Mar. Drugs 2022, 20, 386. https://doi.org/10.3390/md20060386
Nuzzo G, Senese G, Gallo C, Albiani F, Romano L, d’Ippolito G, Manzo E, Fontana A. Antitumor Potential of Immunomodulatory Natural Products. Marine Drugs. 2022; 20(6):386. https://doi.org/10.3390/md20060386
Chicago/Turabian StyleNuzzo, Genoveffa, Giuseppina Senese, Carmela Gallo, Federica Albiani, Lucia Romano, Giuliana d’Ippolito, Emiliano Manzo, and Angelo Fontana. 2022. "Antitumor Potential of Immunomodulatory Natural Products" Marine Drugs 20, no. 6: 386. https://doi.org/10.3390/md20060386
APA StyleNuzzo, G., Senese, G., Gallo, C., Albiani, F., Romano, L., d’Ippolito, G., Manzo, E., & Fontana, A. (2022). Antitumor Potential of Immunomodulatory Natural Products. Marine Drugs, 20(6), 386. https://doi.org/10.3390/md20060386