
Characterization of Plocamium telfairiae Extract-Functionalized Au Nanostructures and Their Anti-Adipogenic Activity through PLD1

 ,
,  , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
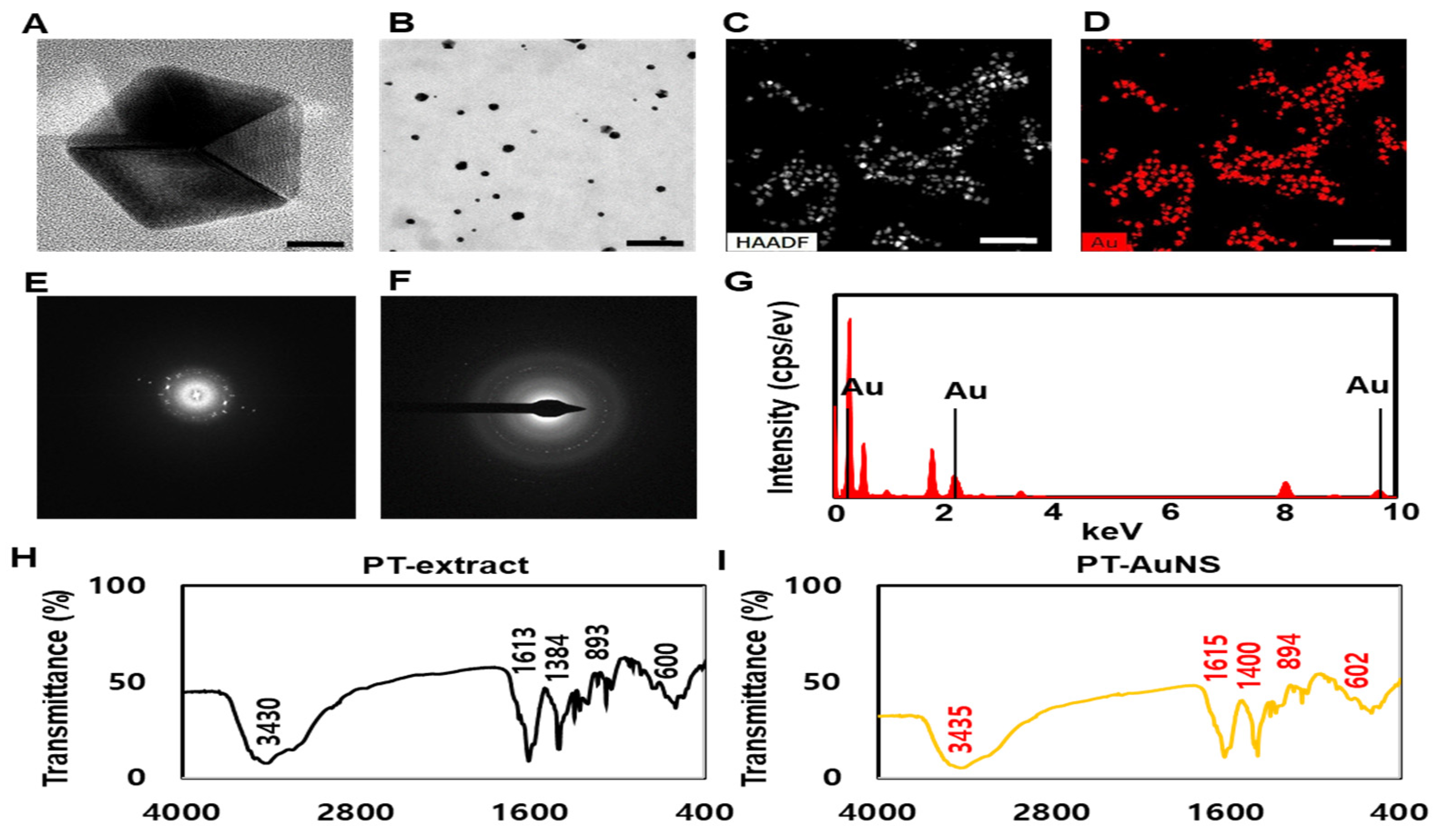
2.1. Physicochemical Characterization of PT-AuNSs
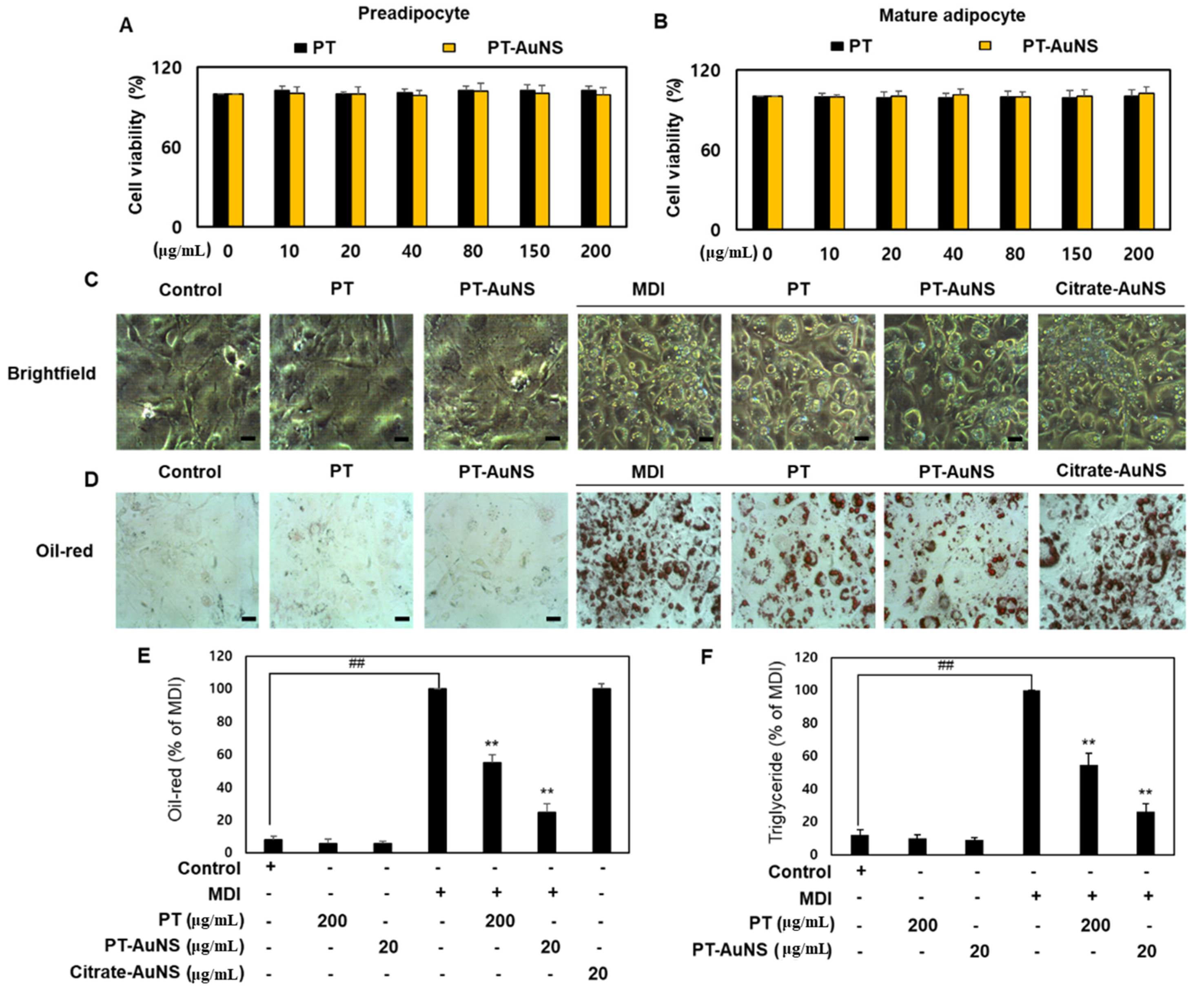
2.2. Effects of PT-AuNS on Oil Red O Staining Activity and TG Accumulation
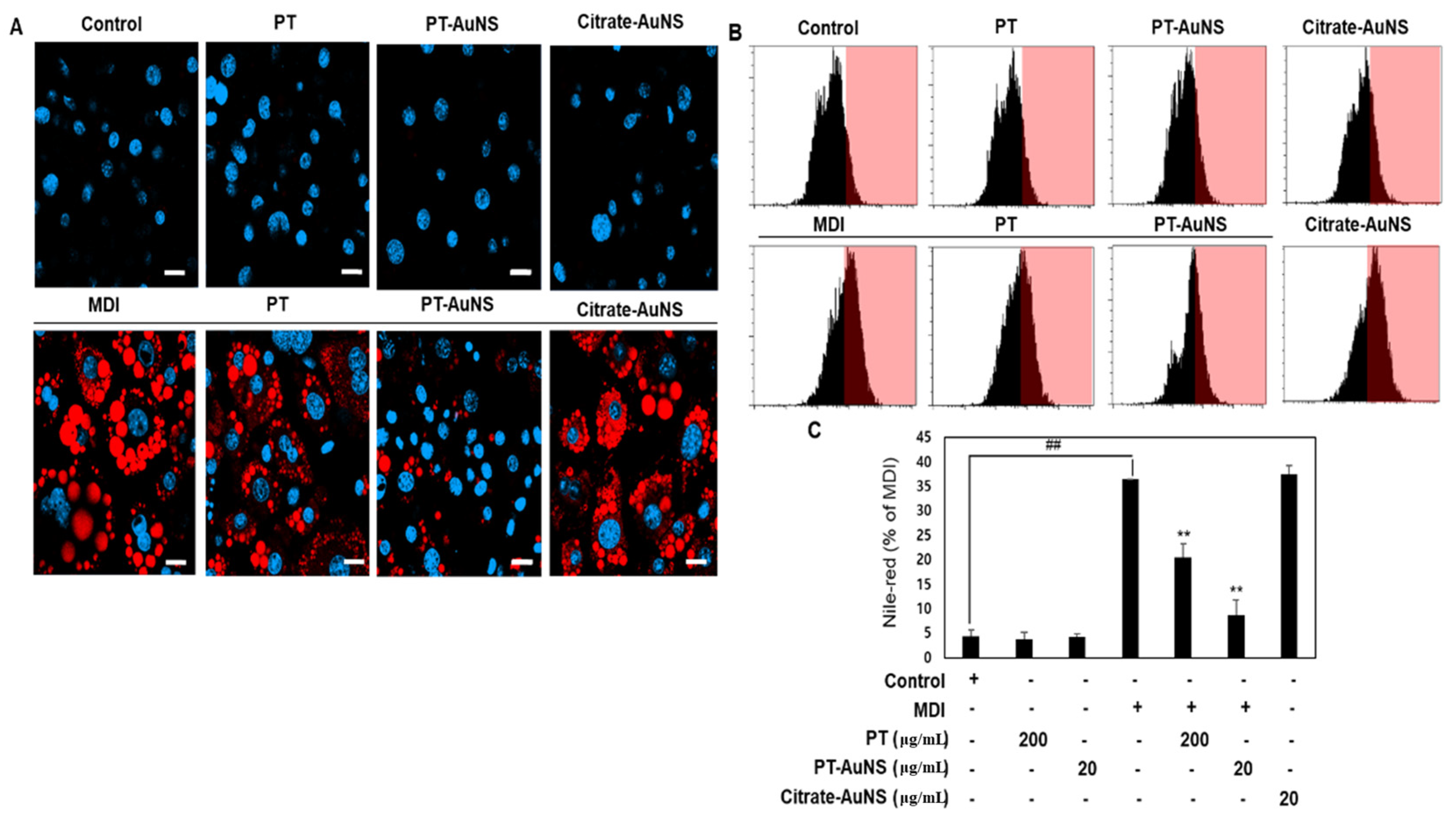
2.3. Effect of PT Extract and PT-AuNS on Nile Red Staining Activity
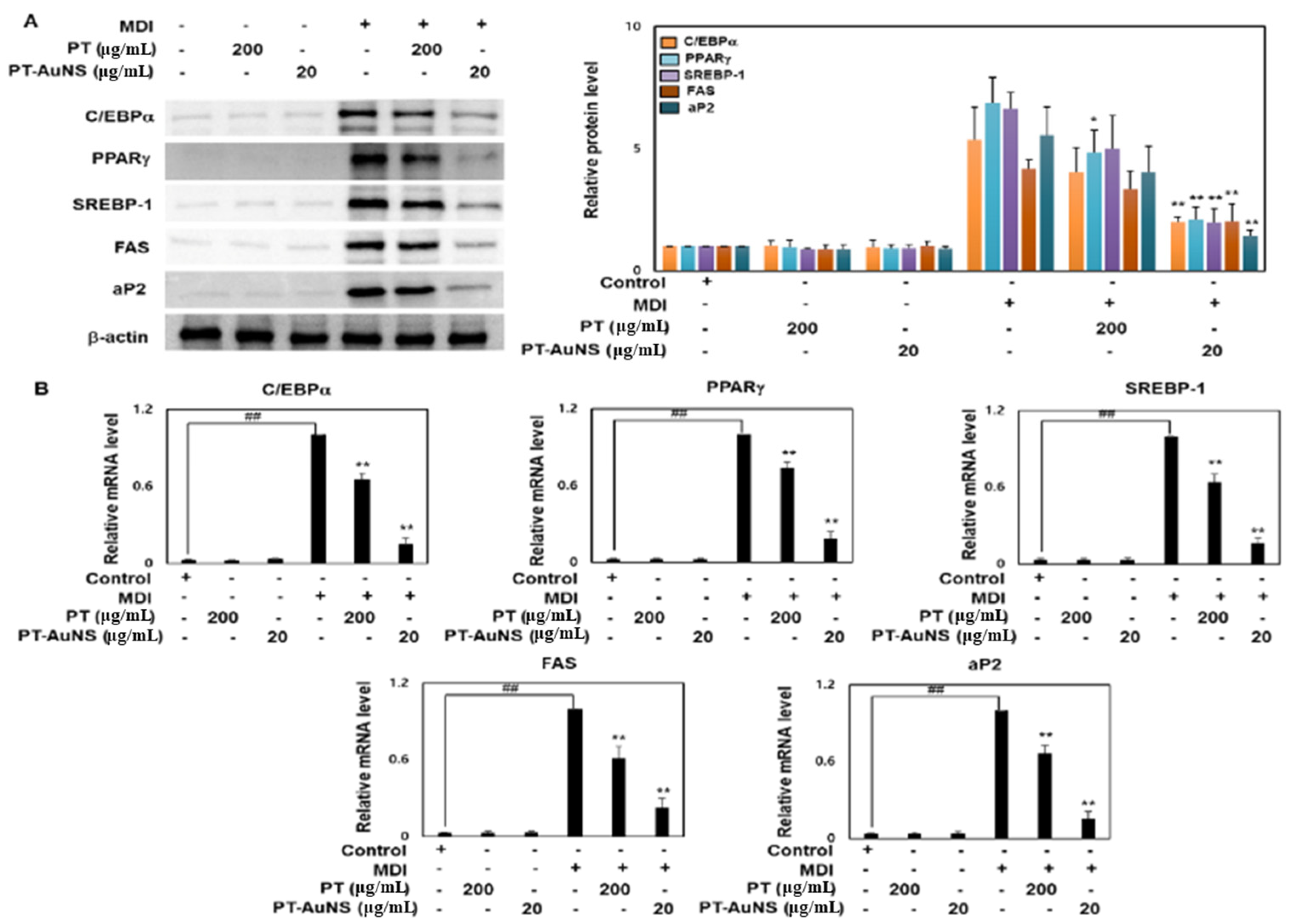
2.4. Effects of PT Extract and PT-AuNS on the Transcript and Protein Expression of Adipogenic Marker
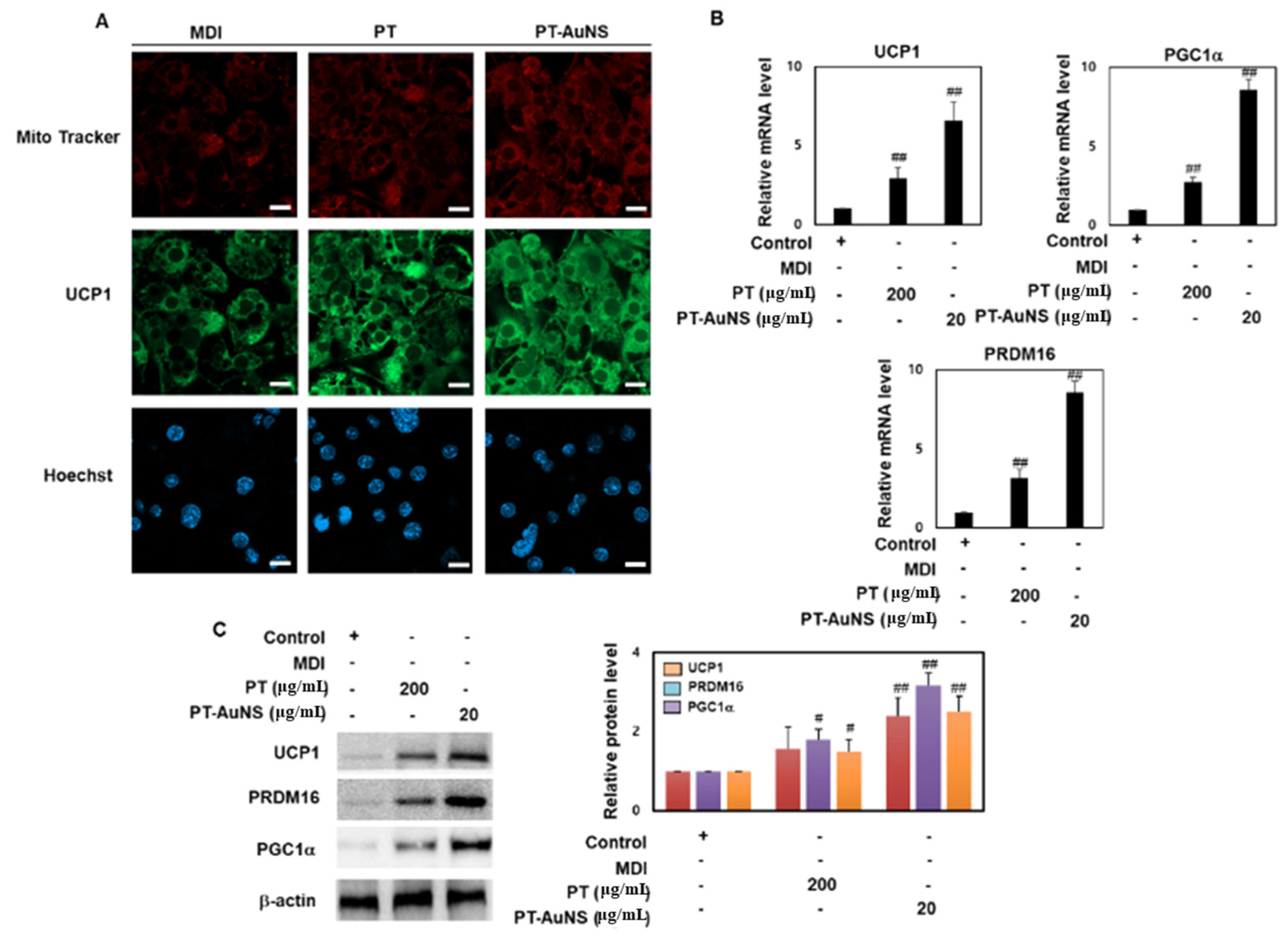
2.5. Effects of PT Extract and PT-AuNS on the Mitochondrial Thermogenesis in Mature Adipocyte
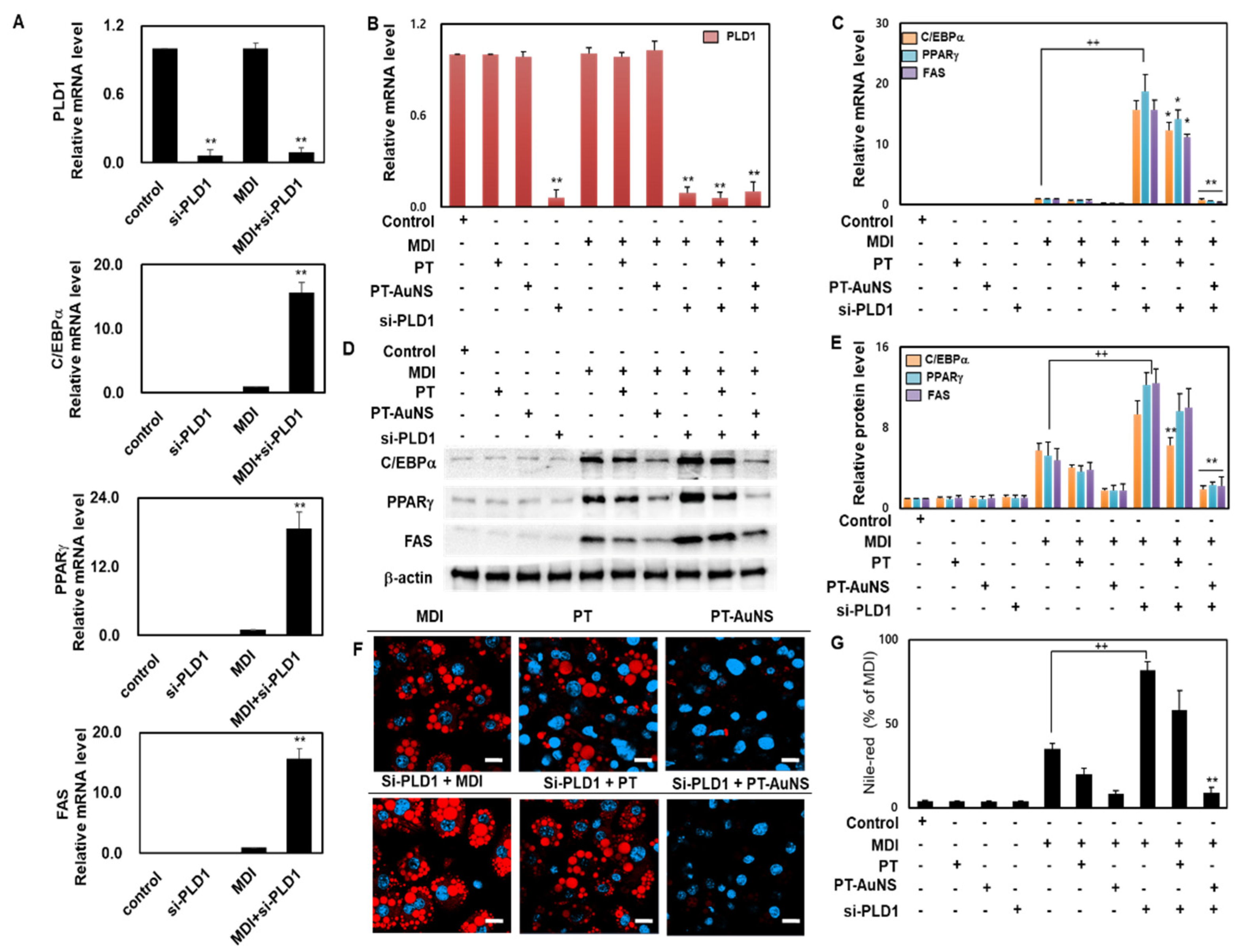
2.6. Anti-Adipogenesis Effects of PT Extract and PT-AuNS via PLD1 Regulation
3. Materials and Methods
3.1. Reagents
3.2. Preparation of PT Extract
3.3. PT-AuNS Synthesis
3.4. Characterization of PT-AuNS
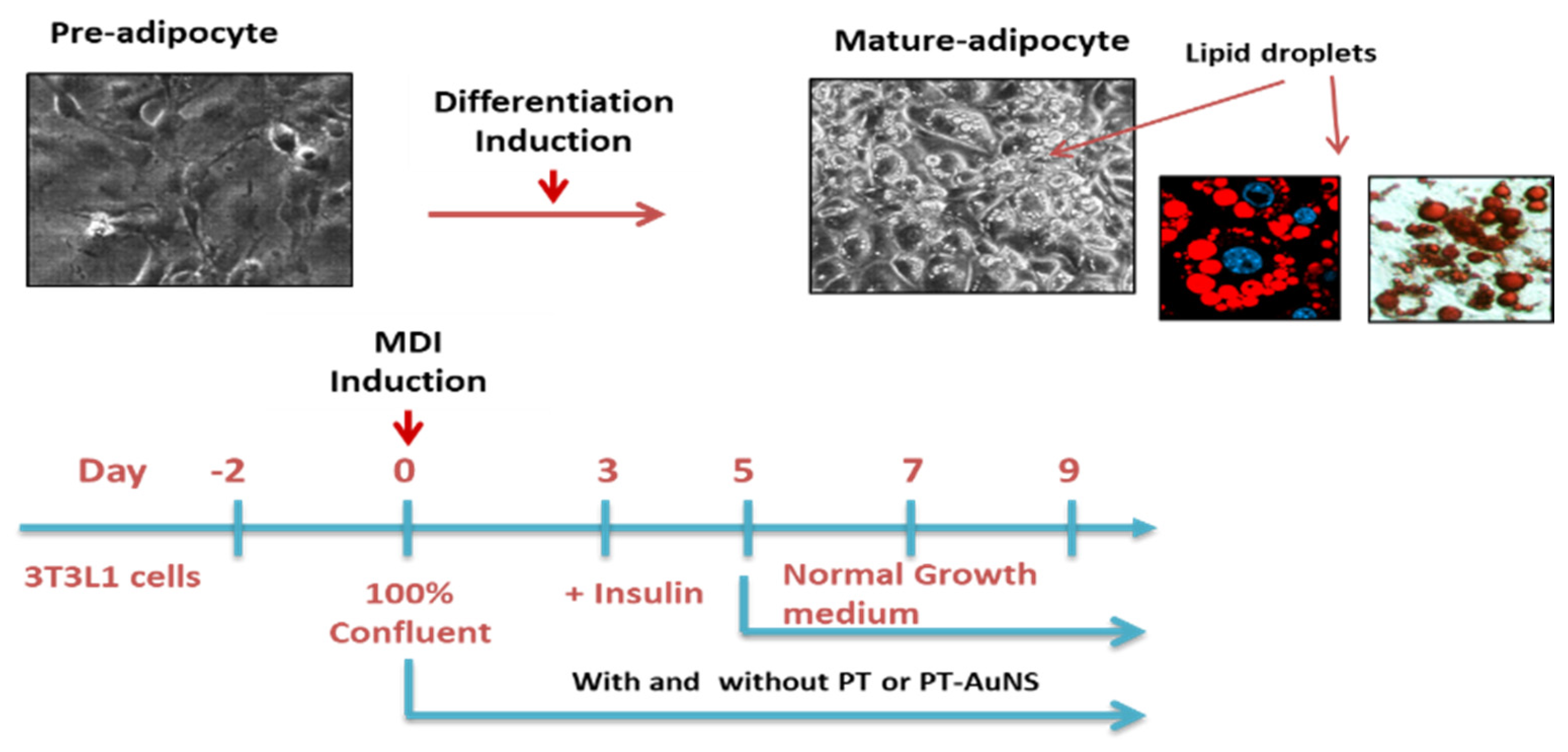
3.5. Pre- and Mature Adipocyte Culture and Treatment
3.6. Viability of Pre- and Mature-Adipocytes
3.7. Lipid Droplet Measurement Using Oil Red O and Nile Red Staining
3.8. Triglyceride Analysis
3.9. Reverse Transcription-Quantitative Polymerase Chain Reaction (RT-qPCR)
3.10. Western Blotting
3.11. Immunofluorescence Assay
3.12. PLD1 Knockdown Using siRNA Transfection
3.13. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Mir, I.A.; Soni, R.; Srivastav, S.K.; Bhavya, I.; Dar, W.Q.; Farooq, M.D.; Chawla, V.; Nadeem, M. Obesity as an Important Marker of the COVID-19 Pandemic. Cureus 2022, 14, e21403. [Google Scholar] [CrossRef] [PubMed]
- Murugan, D.D.; Balan, D.; Wong, P.F. Adipogenesis and therapeutic potentials of antiobesogenic phytochemicals: Insights from preclinical studies. Phytother. Res. 2021, 35, 5936–5960. [Google Scholar] [CrossRef] [PubMed]
- Barrea, L.; Frias-Toral, E.; Pugliese, G.; Garcia-Velasquez, E.; DE Los Angeles Carignano, M.; Savastano, S.; Colao, A.; Muscogiuri, G. Vitamin D in obesity and obesity-related diseases: An overview. Minerva Endocrinol. 2021, 46, 177–192. [Google Scholar] [CrossRef]
- Lo Re, O.; Maugeri, A.; Hruskova, J.; Jakubik, J.; Kucera, J.; Bienertova-Vasku, J.; Oben, J.A.; Kubala, L.; Dvorakova, A.; Ciz, M.; et al. Obesity-induced nucleosome release predicts poor cardio-metabolic health. Clin. Epigenetics 2019, 12, 2. [Google Scholar] [CrossRef] [Green Version]
- Jiao, Y.; Ahmed, U.; Sim, M.F.M.; Bejar, A.; Zhang, X.; Talukder, M.M.U.; Rice, R.; Flannick, J.; Podgornaia, A.I.; Reilly, D.F.; et al. Discovering metabolic disease gene interactions by correlated effects on cellular morphology. Mol. Metab. 2019, 24, 108–119. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Du, X.; Turner, N.; Brown, A.J.; Yang, H. Enhanced acyl-CoA:cholesterol acyltransferase activity increases cholesterol levels on the lipid droplet surface and impairs adipocyte function. J. Biol. Chem. 2019, 294, 19306–19321. [Google Scholar] [CrossRef]
- Hauffe, R.; Rath, M.; Agyapong, W.; Jonas, W.; Vogel, H.; Schulz, T.J.; Schwarz, M.; Kipp, A.P.; Blüher, M.; Kleinridders, A. Obesity Hinders the Protective Effect of Selenite Supplementation on Insulin Signaling. Antioxidants 2022, 11, 862. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Ma, K.; Kang, Y.; Liu, W.; Liu, X.; Long, X.; Hayashi, T.; Hattori, S.; Mizuno, K.; Fujisaki, H.; et al. Type I collagen reduces lipid accumulation during adipogenesis of preadipocytes 3T3-L1 via the YAP-mTOR-autophagy axis. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2022, 1867, 159181. [Google Scholar] [CrossRef]
- Sathyanarayana, A.R.; Lu, C.K.; Liaw, C.C.; Chang, C.C.; Han, H.Y.; Green, B.D.; Huang, W.J.; Huang, C.; He, W.D.; Lee, L.C.; et al. 1,2,3,4,6-Penta-O-galloyl-d-glucose Interrupts the Early Adipocyte Lifecycle and Attenuates Adiposity and Hepatic Steatosis in Mice with Diet-Induced Obesity. Int. J. Mol. Sci. 2022, 23, 4052. [Google Scholar] [CrossRef]
- Bae, S.J.; Kim, J.E.; Choi, Y.J.; Lee, S.J.; Gong, J.E.; Choi, Y.W.; Hwang, D.Y. Novel Function of α-Cubebenoate Derived from Schisandra chinensis as Lipogenesis Inhibitor, Lipolysis Stimulator and Inflammasome Suppressor. Molecules 2020, 25, 4995. [Google Scholar] [CrossRef]
- Park, E.; Lee, C.G.; Kim, J.; Yeo, S.; Kim, J.A.; Choi, C.W.; Jeong, S.Y. Antiobesity Effects of Gentiana lutea Extract on 3T3-L1 Preadipocytes and a High-Fat Diet-Induced Mouse Model. Molecules 2020, 25, 2453. [Google Scholar] [CrossRef] [PubMed]
- Mladenova, S.G.; Savova, M.S.; Marchev, A.S.; Ferrante, C.; Orlando, G.; Wabitsch, M.; Georgiev, M.I. Anti-adipogenic activity of maackiain and ononin is mediated via inhibition of PPARγ in human adipocytes. Biomed. Pharmacother. 2022, 149, 112908. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, T.; Saito, T.; Aoki-Saito, H.; Okada, S.; Ikeda, H.; Nakakura, T.; Fukuda, H.; Arai, S.; Fujiwara, K.; Nakajima, Y.; et al. Resolvin E3 ameliorates high-fat diet-induced insulin resistance via the phosphatidylinositol-3-kinase/Akt signaling pathway in adipocytes. FASEB J. 2022, 36, e22188. [Google Scholar] [CrossRef] [PubMed]
- Dai, H.B.; Wang, H.Y.; Wang, F.Z.; Qian, P.; Gao, Q.; Zhou, H.; Zhou, Y.B. Adrenomedullin ameliorates palmitic acid-induced insulin resistance through PI3K/Akt pathway in adipocytes. Acta Diabetol. 2022, 59, 661–673. [Google Scholar] [CrossRef]
- Cataldi, S.; Aprile, M.; Melillo, D.; Mucel, I.; Giorgetti-Peraldi, S.; Cormont, M.; Italiani, P.; Blüher, M.; Tanti, J.F.; Ciccodicola, A.; et al. TNFα Mediates Inflammation-Induced Effects on PPARG Splicing in Adipose Tissue and Mesenchymal Precursor Cells. Cells 2021, 11, 42. [Google Scholar] [CrossRef]
- Lee, K.D.; Ilavenil, S.; Karnan, M.; Yang, C.J.; Kim, D.; Choi, K.C. Novel Bacillus ginsengihumi CMRO6 Inhibits Adipogenesis via p38MAPK/Erk44/42 and Stimulates Glucose Uptake in 3T3-L1 Pre-Adipocytes through Akt/AS160 Signaling. Int. J. Mol. Sci. 2022, 23, 4727. [Google Scholar] [CrossRef]
- Abbas, M.A.; Boby, N.; Lee, E.B.; Hong, J.H.; Park, S.C. Anti-Obesity Effects of Ecklonia cava Extract in High-Fat Diet-Induced Obese Rats. Antioxidants 2022, 11, 310. [Google Scholar] [CrossRef]
- Hinds, T.D., Jr.; Kipp, Z.A.; Xu, M.; Yiannikouris, F.B.; Morris, A.J.; Stec, D.F.; Wahli, W.; Stec, D.E. Adipose-Specific PPARα Knockout Mice Have Increased Lipogenesis by PASK-SREBP1 Signaling and a Polarity Shift to Inflammatory Macrophages in White Adipose Tissue. Cells 2021, 11, 4. [Google Scholar] [CrossRef]
- Jee, W.; Lee, S.H.; Ko, H.M.; Jung, J.H.; Chung, W.S.; Jang, H.J. Anti-Obesity Effect of Polygalin C Isolated from Polygala japonica Houtt. via Suppression of the Adipogenic and Lipogenic Factors in 3T3-L1 Adipocytes. Int. J. Mol. Sci. 2021, 22, 10405. [Google Scholar] [CrossRef]
- Lim, S.H.; Lee, H.S.; Han, H.K.; Choi, C.I. Saikosaponin A and D Inhibit Adipogenesis via the AMPK and MAPK Signaling Pathways in 3T3-L1 Adipocytes. Int. J. Mol. Sci. 2021, 22, 11409. [Google Scholar] [CrossRef]
- Chater, P.I.; Wilcox, M.D.; Houghton, D.; Pearson, J.P. The role of seaweed bioactives in the control of digestion: Implications for obesity treatments. Food Funct. 2015, 6, 3420–3427. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, M.D.; Cherry, P.; Chater, P.I.; Yang, X.; Zulali, M.; Okello, E.J.; Seal, C.J.; Pearson, J.P. The effect of seaweed enriched bread on carbohydrate digestion and the release of glucose from food. J. Funct. Foods 2021, 87, 104747. [Google Scholar] [CrossRef] [PubMed]
- Attjioui, M.; Ryan, S.; Ristic, A.K.; Higgins, T.; Goñi, O.; Gibney, E.R.; Tierney, J.; O’Connell, S. Comparison of edible brown algae extracts for the inhibition of intestinal carbohydrate digestive enzymes involved in glucose release from the diet. J. Nutr. Sci. 2021, 10, e5. [Google Scholar] [CrossRef] [PubMed]
- Gabbia, D.; Dall’Acqua, S.; Di Gangi, I.M.; Bogialli, S.; Caputi, V.; Albertoni, L.; Marsilio, I.; Paccagnella, N.; Carrara, M.; Giron, M.C.; et al. The Phytocomplex from Fucus vesiculosus and Ascophyllum nodosum Controls Postprandial Plasma Glucose Levels: An In Vitro and In Vivo Study in a Mouse Model of NASH. Mar. Drugs 2017, 15, 41. [Google Scholar] [CrossRef] [PubMed]
- Gabbia, D.; Saponaro, M.; Sarcognato, S.; Guido, M.; Ferri, N.; Carrara, M.; De Martin, S. Fucus vesiculosus and Ascophyllum nodosum Ameliorate Liver Function by Reducing Diet-Induced Steatosis in Rats. Mar. Drugs 2020, 18, 62. [Google Scholar] [CrossRef] [Green Version]
- Hwang, J.; Yadav, D.; Lee, P.C.; Jin, J.O. Immunomodulatory effects of polysaccharides from marine algae for treating cancer, infectious disease, and inflammation. Phytother. Res. 2022, 36, 761–777. [Google Scholar] [CrossRef]
- Gao, Y.; Li, Y.; Niu, Y.; Ju, H.; Chen, R.; Li, B.; Song, X.; Song, L. Chemical Characterization, Antitumor, and Immune-Enhancing Activities of Polysaccharide from Sargassum pallidum. Molecules 2021, 26, 7559. [Google Scholar] [CrossRef]
- Babu, B.; Palanisamy, S.; Vinosha, M.; Anjali, R.; Kumar, P.; Pandi, B.; Tabarsa, M.; You, S.; Prabhu, N.M. Bioengineered gold nanoparticles from marine seaweed Acanthophora spicifera for pharmaceutical uses: Antioxidant, antibacterial, and anticancer activities. Bioprocess Biosyst. Eng. 2020, 43, 2231–2242. [Google Scholar] [CrossRef]
- Jeyarani, S.; Vinita, N.M.; Puja, P.; Senthamilselvi, S.; Devan, U.; Velangani, A.J.; Biruntha, M.; Pugazhendhi, A.; Kumar, P. Biomimetic gold nanoparticles for its cytotoxicity and biocompatibility evidenced by fluorescence-based assays in cancer (MDA-MB-231) and non-cancerous (HEK-293) cells. J. Photochem. Photobiol. B 2020, 202, 111715. [Google Scholar] [CrossRef]
- Khanna, P.; Kaur, A.; Goyal, D. Algae-based metallic nanoparticles: Synthesis, characterization and applications. J. Microbiol. Methods 2019, 163, 105656. [Google Scholar] [CrossRef]
- Chen, X.; Han, W.; Zhao, X.; Tang, W.; Wang, F. Epirubicin-loaded marine carrageenan oligosaccharide capped gold nanoparticle system for pH-triggered anticancer drug release. Sci. Rep. 2019, 9, 6754. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Zhao, X.; Gao, Y.; Yin, J.; Bai, M.; Wang, F. Green Synthesis of Gold Nanoparticles Using Carrageenan Oligosaccharide and Their In Vitro Antitumor Activity. Mar. Drugs 2018, 16, 277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.A.; Lee, H.G.; Li, X.; Hyun, J.M.; Kim, H.S.; Kim, T.H.; Kim, H.M.; Lee, J.J.; Kang, M.C.; Jeon, Y.J. Anti-obesity effects of red seaweed, Plocamium telfairiae, in C57BL/6 mice fed a high-fat diet. Food Funct. 2020, 11, 2299–2308. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Yoon, M.Y.; Cha, M.R.; Hwang, J.H.; Park, E.; Choi, S.U.; Park, H.R.; Hwang, Y.I. Methanolic extracts of Plocamium telfairiae induce cytotoxicity and caspase-dependent apoptosis in HT-29 human colon carcinoma cells. J. Med. Food 2007, 10, 587–593. [Google Scholar] [CrossRef]
- Huang, H.L.; Wang, B.G. Antioxidant capacity and lipophilic content of seaweeds collected from the Qingdao coastline. J. Agric. Food Chem. 2004, 52, 4993–4997. [Google Scholar] [CrossRef]
- Oruganti, L.; Reddy Sankaran, K.; Dinnupati, H.G.; Kotakadi, V.S.; Meriga, B. Anti-adipogenic and lipid-lowering activity of piperine and epigallocatechin gallate in 3T3-L1 adipocytes. Arch. Physiol. Biochem. 2021, 9, 1–8. [Google Scholar] [CrossRef]
- Park, M.; Han, J.; Lee, H.J. Anti-Adipogenic Effect of Neferine in 3T3-L1 Cells and Primary White Adipocytes. Nutrients 2020, 12, 1858. [Google Scholar] [CrossRef]
- Tung, Y.C.; Chou, R.F.; Nagabhushanam, K.; Ho, C.T.; Pan, M.H. 3’-Hydroxydaidzein Improves Obesity Through the Induced Browning of Beige Adipose and Modulation of Gut Microbiota in Mice with Obesity Induced by a High-Fat Diet. J. Agric. Food Chem. 2020, 68, 14513–14522. [Google Scholar] [CrossRef]
- Sung, Y.Y.; Son, E.; Im, G.; Kim, D.S. Herbal Combination of Phyllostachys pubescens and Scutellaria baicalensis Inhibits Adipogenesis and Promotes Browning via AMPK Activation in 3T3-L1 Adipocytes. Plants 2020, 9, 1422. [Google Scholar] [CrossRef]
- Gu, Y.; Xiao, X.; Pan, R.; Zhang, J.; Zhao, Y.; Dong, Y.; Cui, H. Lactobacillus plantarum dy-1 fermented barley extraction activates white adipocyte browning in high-fat diet-induced obese rats. J. Food Biochem. 2021, 45, e13680. [Google Scholar] [CrossRef]
- Pan, J.; Kothan, S.; Moe Moe, A.T.; Huang, K. Dysfunction of insulin-AKT-UCP1 signalling inhibits transdifferentiation of human and mouse white preadipocytes into brown-like adipocytes. Adipocyte 2022, 11, 213–226. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.P.; Schragenheim, J.; Cao, J.; Falck, J.R.; Abraham, N.G.; Bellner, L. PGC-1 alpha regulates HO-1 expression, mitochondrial dynamics and biogenesis: Role of epoxyeicosatrienoic acid. Prostaglandins Other Lipid Mediat. 2016, 125, 8–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subash-Babu, P.; Mohammed Alowaidh, H.; Al-Harbi, L.N.; Shamlan, G.; Aloud, A.A.; AlSedairy, S.A.; Alshatwi, A.A. Ocimum basilicum L. Methanol Extract Enhances Mitochondrial Efficiency and Decreases Adipokine Levels in Maturing Adipocytes Which Regulate Macrophage Systemic Inflammation. Molecules 2022, 27, 1388. [Google Scholar] [CrossRef]
- Kang, D.W.; Hwang, W.C.; Noh, Y.N.; Che, X.; Lee, S.H.; Jang, Y.; Choi, K.Y.; Choi, J.Y.; Min, D.S. Deletion of phospholipase D1 decreases bone mass and increases fat mass via modulation of Runx2, β-catenin-osteoprotegerin, PPAR-γ and C/EBPα signaling axis. Biochim. Biophys. Acta Mol. Basis Dis. 2021, 1867, 166084. [Google Scholar] [CrossRef]
- Song, H.I.; Yoon, M.S. PLD1 regulates adipogenic differentiation through mTOR-IRS-1 phosphorylation at serine 636/639. Sci. Rep. 2016, 6, 36968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Müller, G.A.; Tschöp, M.H.; Müller, T.D. Upregulated phospholipase D activity toward glycosylphosphatidylinositol-anchored proteins in micelle-like serum complexes in metabolically deranged rats and humans. Am. J. Physiol. Endocrinol. Metab. 2020, 318, E462–E479. [Google Scholar] [CrossRef] [PubMed]
- Jun, E.S.; Kim, Y.J.; Kim, H.H.; Park, S.Y. Gold Nanoparticles Using Ecklonia stolonifera Protect Human Dermal Fibroblasts from UVA-Induced Senescence through Inhibiting MMP-1 and MMP-3. Mar. Drugs 2020, 18, 433. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Forward Primer (5′→3′) | Reverse Primer (3′→5′) |
|---|---|---|
| C/EBPα | TGGTGATTTGTCCGTTGTCT | GGA AACCTGGCCTGTTGTAAG |
| PPARγ | GGTGATTTGTCCGTTGTCT | GCTTCAATCGGATGGTTCTTC |
| SREBP-1 | TAGAGCATATCCCCCAGGTG | GGTACGGGCCACAAGAAGTA |
| FAS | GCTGCGGAAACTTCAGGAAAT | AGAGACGTGTCACTCCTGGACTT |
| aP2 | GGATTTGGTCACCATCCGGT | TTCACCTTCCTGTCGTCTGC |
| UCP1 | CCTGCCTCTCTCGGAAACAA | GTAGCGGGGTTTGATCCCAT |
| PRDM16 | CAGCACGGTGAAGCCATTC | GCGTGCATCCGCTTGTG |
| PGC1α | ATGTGCAGCCAAGACTCTGTA | CGCTACACCACTTCAATCCAC |
| GAPDH | AGGTCGGTGTGAACGGATTTG | TGTAGACCATGTAGTTGAGGTCA |
| Antibody | Company | Catalog | Species | Dilution |
|---|---|---|---|---|
| C/EBPα | Cell Signaling | #8178 | Rabbit | 1:500 |
| PPARγ | Cell Signaling | #2435 | Rabbit | 1:500 |
| SREBP-1 | Santa Cruz | sc-365513 | Mouse | 1:1000 |
| FAS | Cell Signaling | #3180 | Rabbit | 1:500 |
| aP2 | Cell Signaling | #2120 | Rabbit | 1:500 |
| UCP1 | Santa Cruz | sc-293418 | Mouse | 1:1000 |
| PRDM16 | Abcam | ab106410 | Mouse | 1:500 |
| PGC1α | Santa Cruz | sc-518025 | Mouse | 1:500 |
| β-actin | Santa Cruz | sc-47778 | Goat | 1:1000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, S.Y.; Kang, H.m.; Song, W.C.; Oh, J.-W.; Park, G.; Choi, Y.-W. Characterization of Plocamium telfairiae Extract-Functionalized Au Nanostructures and Their Anti-Adipogenic Activity through PLD1. Mar. Drugs 2022, 20, 421. https://doi.org/10.3390/md20070421
Park SY, Kang Hm, Song WC, Oh J-W, Park G, Choi Y-W. Characterization of Plocamium telfairiae Extract-Functionalized Au Nanostructures and Their Anti-Adipogenic Activity through PLD1. Marine Drugs. 2022; 20(7):421. https://doi.org/10.3390/md20070421
Chicago/Turabian StylePark, Sun Young, Hye mi Kang, Woo Chang Song, Jin-Woo Oh, Geuntae Park, and Young-Whan Choi. 2022. "Characterization of Plocamium telfairiae Extract-Functionalized Au Nanostructures and Their Anti-Adipogenic Activity through PLD1" Marine Drugs 20, no. 7: 421. https://doi.org/10.3390/md20070421
APA StylePark, S. Y., Kang, H. m., Song, W. C., Oh, J. -W., Park, G., & Choi, Y. -W. (2022). Characterization of Plocamium telfairiae Extract-Functionalized Au Nanostructures and Their Anti-Adipogenic Activity through PLD1. Marine Drugs, 20(7), 421. https://doi.org/10.3390/md20070421