
Matairesinol Induces Mitochondrial Dysfunction and Exerts Synergistic Anticancer Effects with 5-Fluorouracil in Pancreatic Cancer Cells



Abstract
:1. Introduction
2. Results
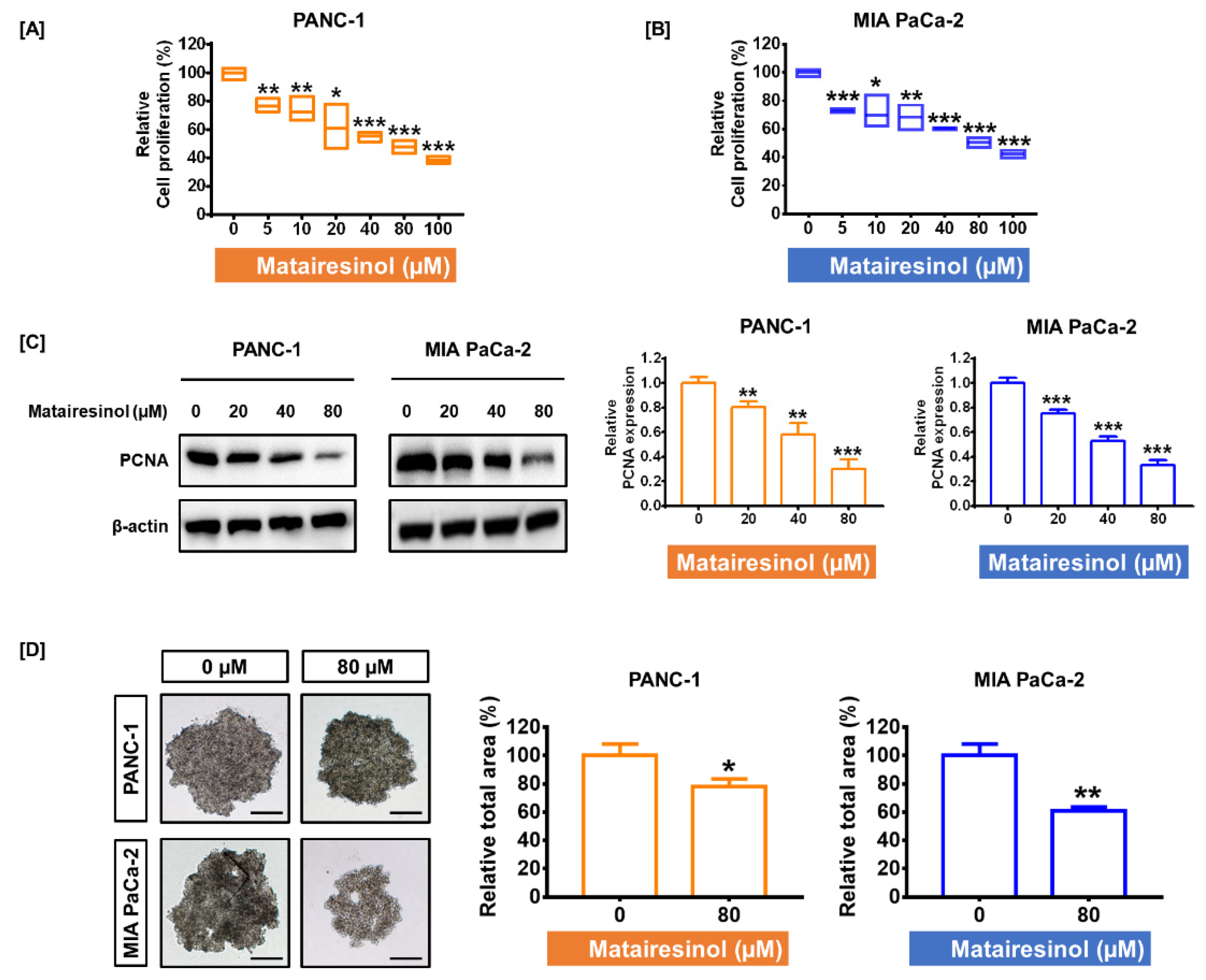
2.1. Matairesinol Inhibited Cell Proliferation and Progression in PC Cells
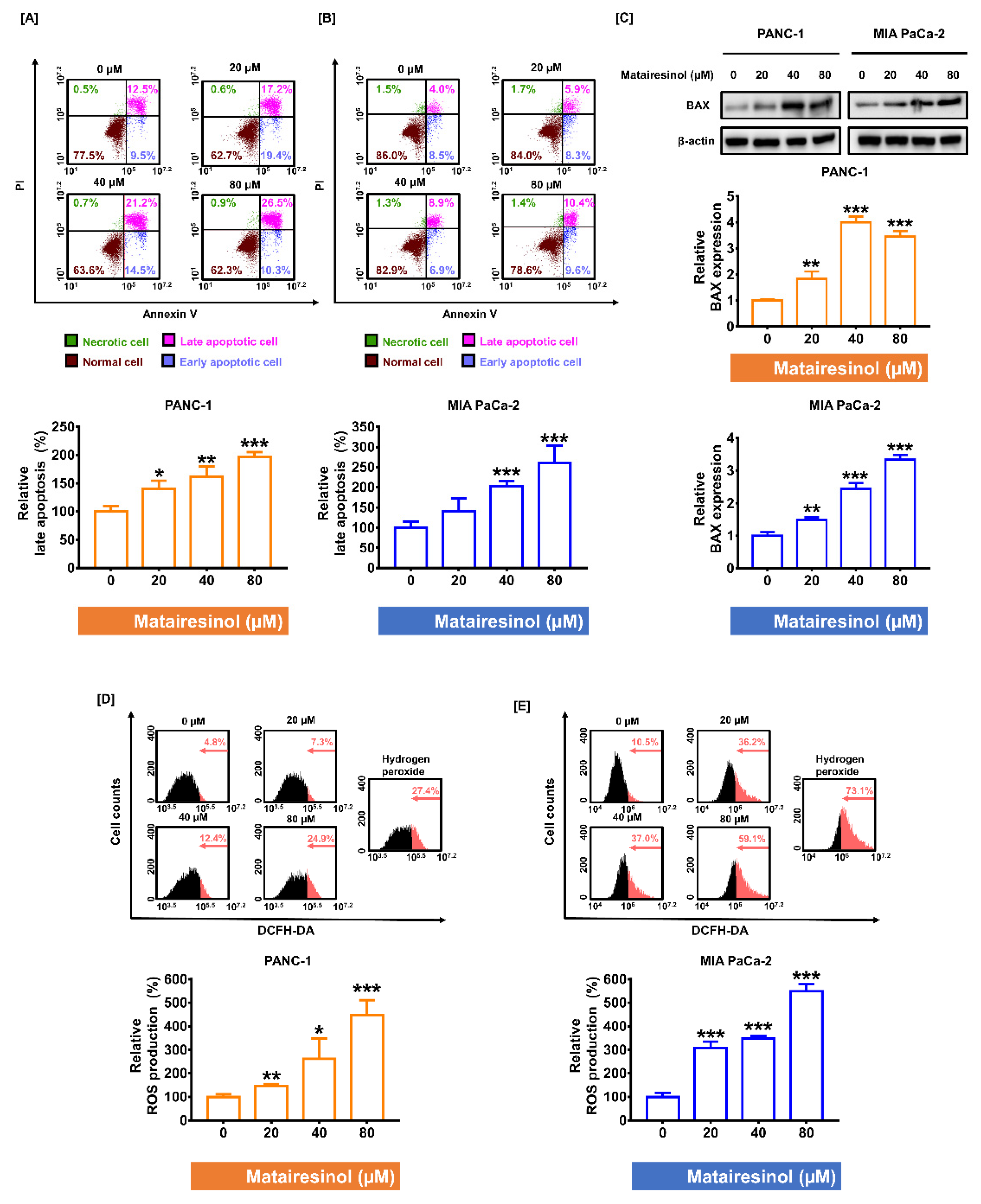
2.2. Matairesinol Intensified Apoptosis Induction and ROS Accumulation in PC Cells
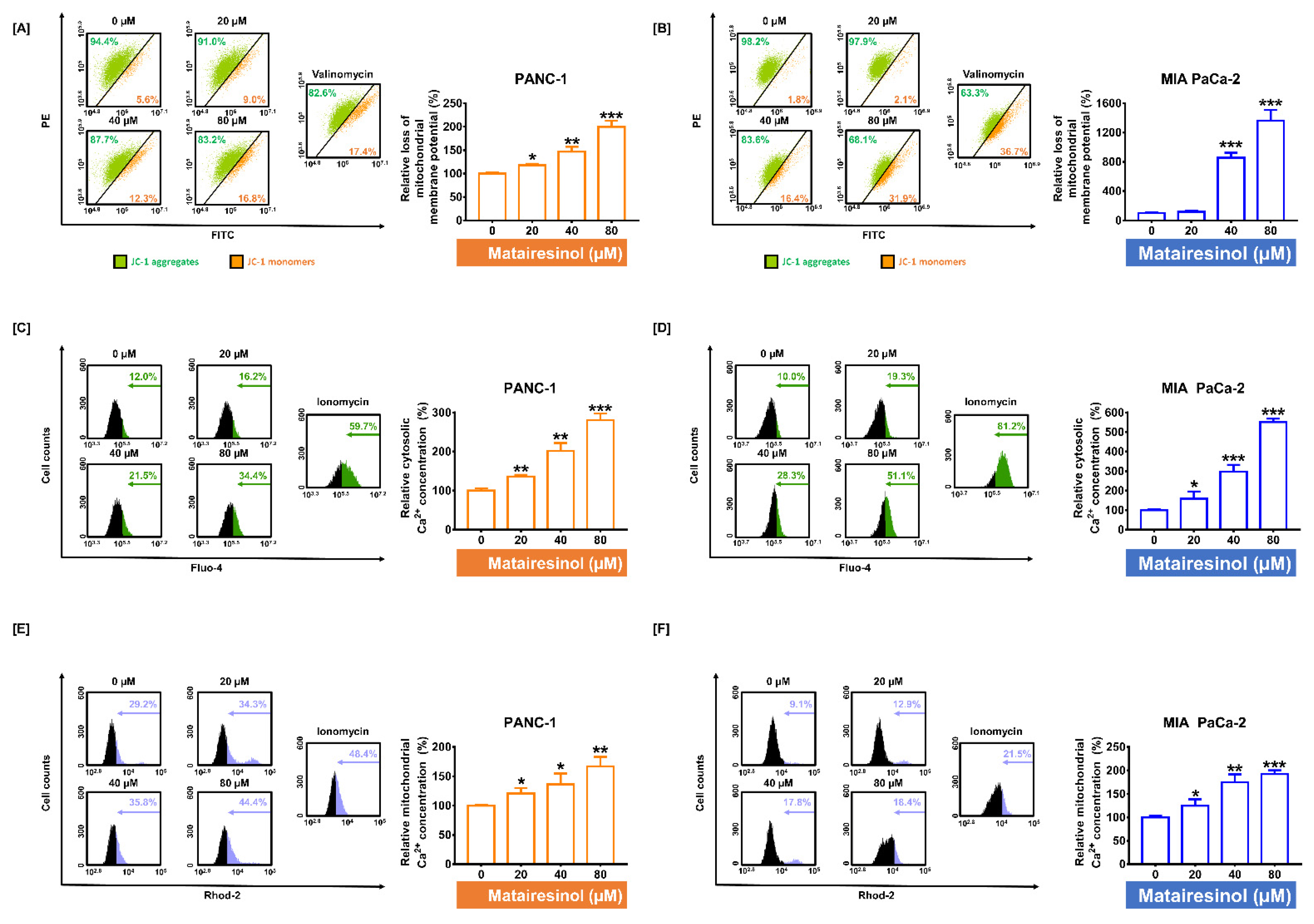
2.3. Matairesinol Provokes Mitochondrial Dysfunction through MMP Loss and Calcium Dysregulation in PC Cells
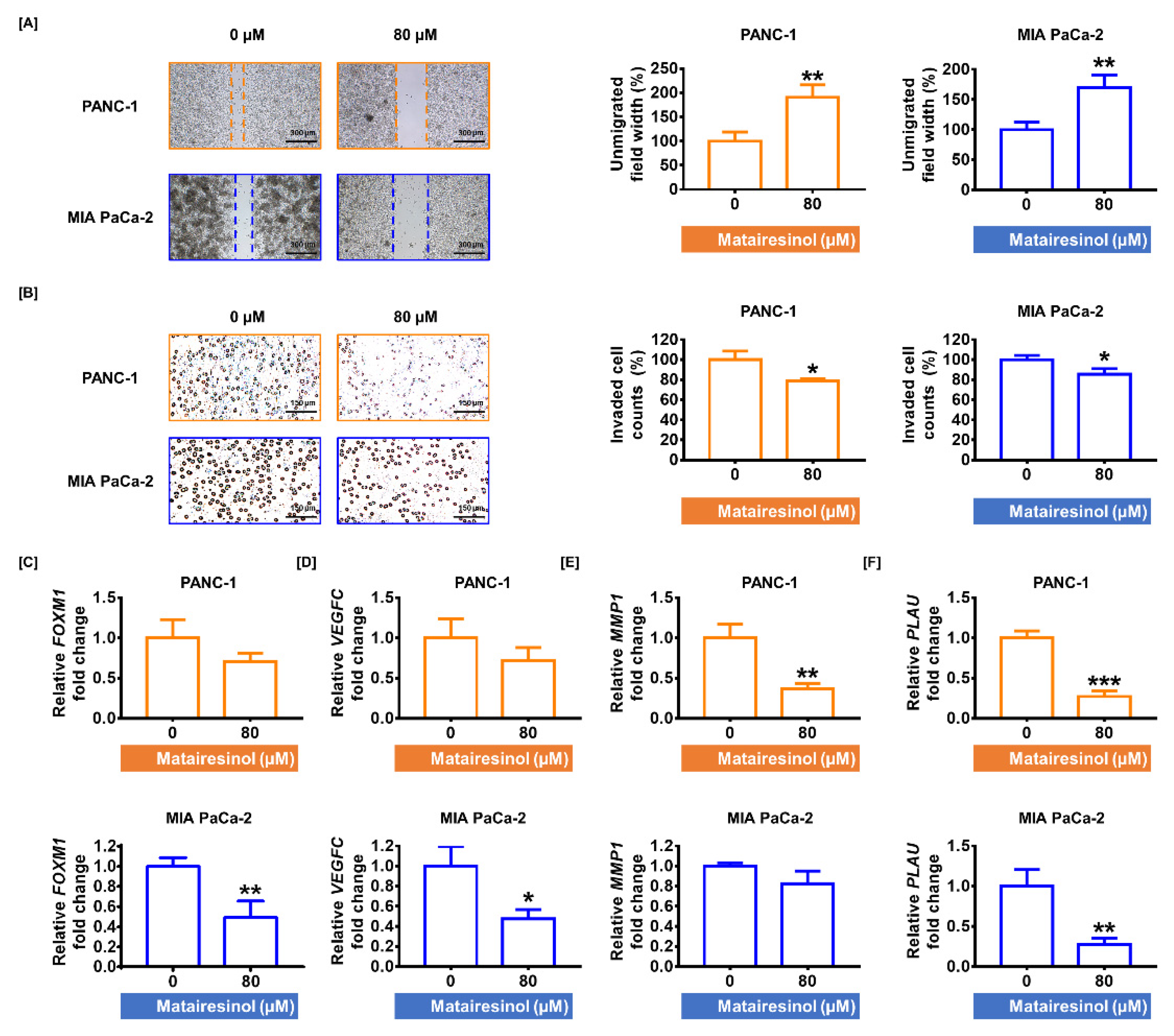
2.4. Matairesinol Mitigates Migratory Ability in PC Cells
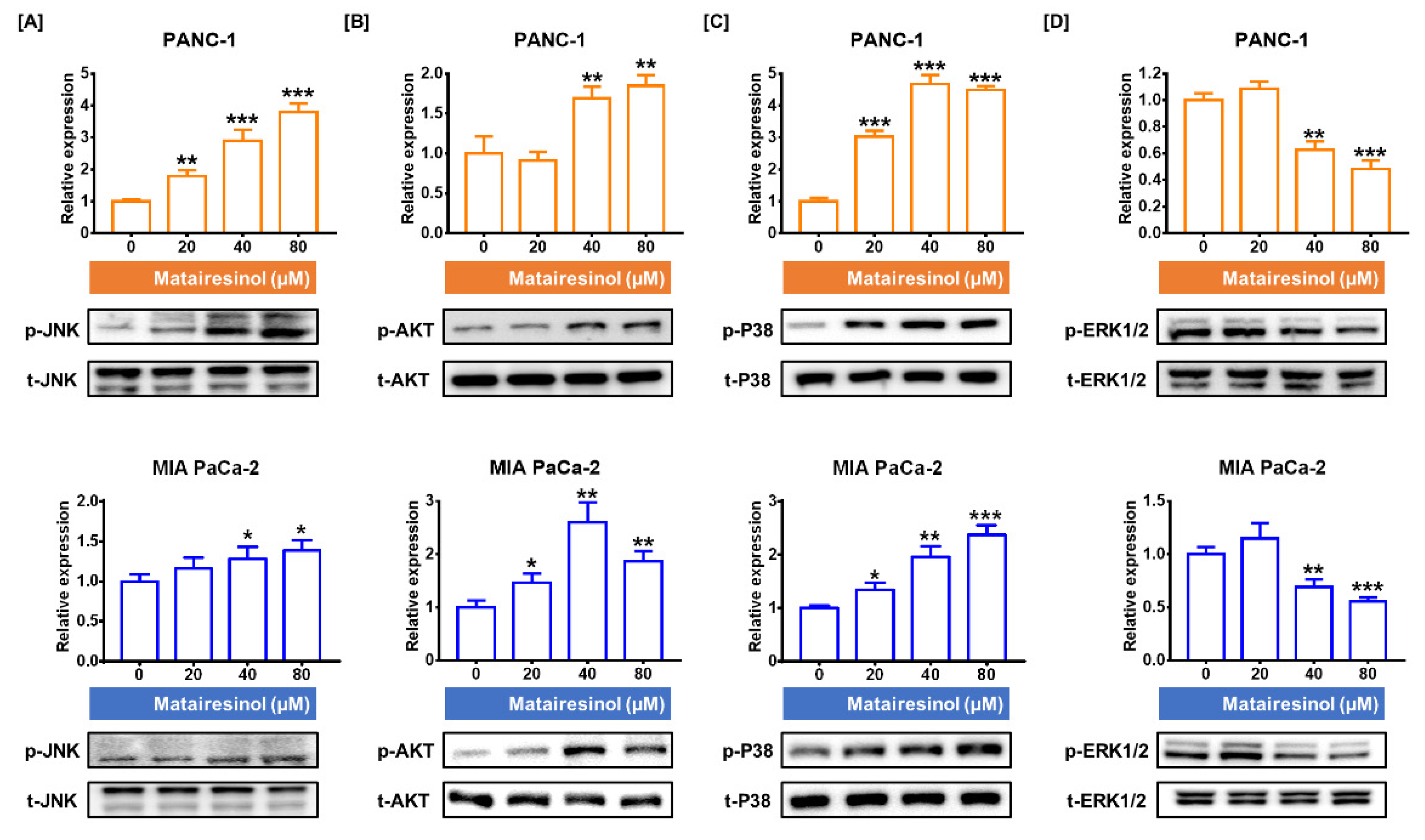
2.5. Signaling Pathways Associated with Antitumor Effects of Matairesinol in PC Cells
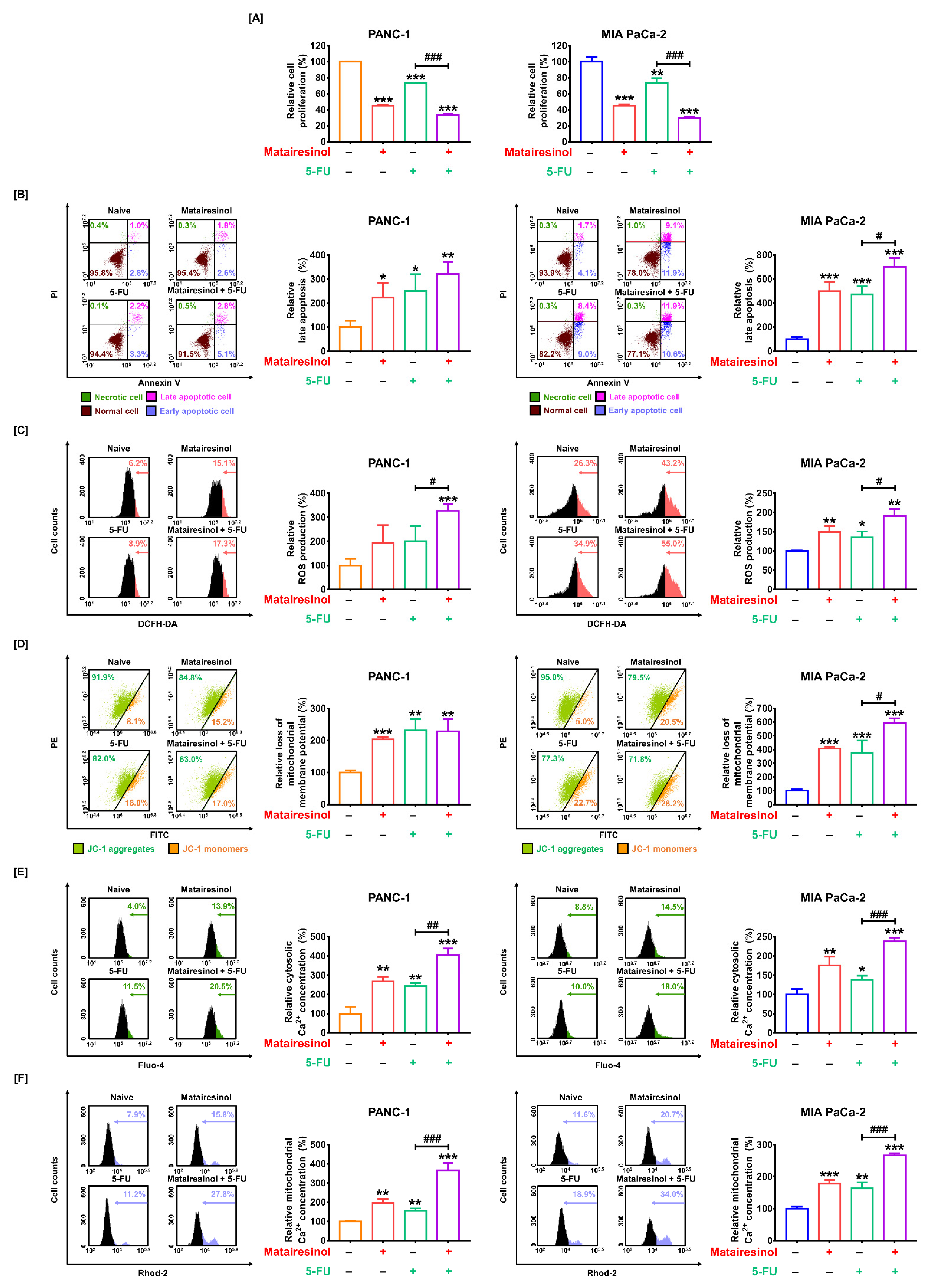
2.6. Synergistic Effects of Matairesinol with Anticancer Drugs in Respect to Mitochondrial Dysfunction
3. Discussion
4. Materials and Methods
4.1. Chemicals and Antibodies
4.2. Cell Culture
4.3. Cell Proliferation Analysis
4.4. Spheroids Formation Analysis
4.5. Reactive Oxygen Species (ROS) Assay
4.6. Mitochondrial Membrane Potential (MMP) Evaluation
4.7. Apoptosis Analysis
4.8. Mitochondrial Calcium Measurement with Rhod-2
4.9. Intracellular Calcium Measurement with Fluo-4
4.10. Cell Migration Using a Transwell Assay
4.11. Cell Invasion Analysis
4.12. Quantitative PCR (qPCR)
4.13. Western Blot
4.14. Synergistic Drug Combination Analysis
4.15. Statistical Analysis
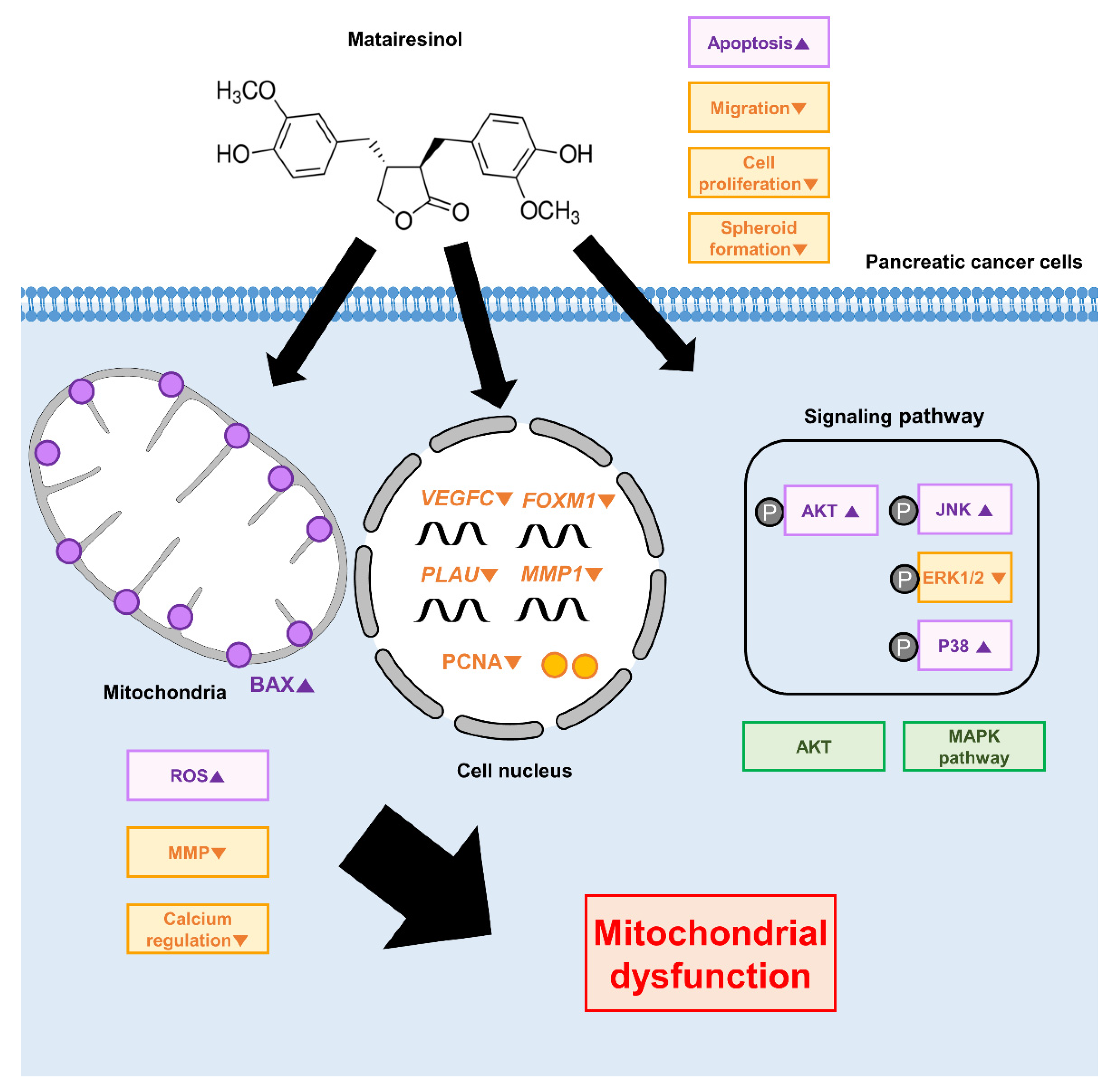
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef] [PubMed]
- Becker, A.E.; Hernandez, Y.G.; Frucht, H.; Lucas, A.L. Pancreatic ductal adenocarcinoma: Risk factors, screening, and early detection. World J. Gastroenterol. 2014, 20, 11182–11198. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, M.; Abdelrahim, M. The Latest Advancement in Pancreatic Ductal Adenocarcinoma Therapy: A Review Article for the Latest Guidelines and Novel Therapies. Biomedicines 2021, 9, 389. [Google Scholar] [CrossRef] [PubMed]
- Rahib, L.; Smith, B.D.; Aizenberg, R.; Rosenzweig, A.B.; Fleshman, J.M.; Matrisian, L.M. Projecting cancer incidence and deaths to 2030: The unexpected burden of thyroid, liver, and pancreas cancers in the United States. Cancer Res. 2014, 74, 2913–2921. [Google Scholar] [CrossRef] [Green Version]
- Waters, A.M.; Der, C.J. KRAS: The Critical Driver and Therapeutic Target for Pancreatic Cancer. Cold Spring Harb. Perspect. Med. 2018, 8, a031435. [Google Scholar] [CrossRef]
- Conroy, T.; Desseigne, F.; Ychou, M.; Bouche, O.; Guimbaud, R.; Becouarn, Y.; Adenis, A.; Raoul, J.L.; Gourgou-Bourgade, S.; de la Fouchardiere, C.; et al. FOLFIRINOX versus gemcitabine for metastatic pancreatic cancer. N. Engl. J. Med. 2011, 364, 1817–1825. [Google Scholar] [CrossRef] [Green Version]
- Mizrahi, J.D.; Rogers, J.E.; Hess, K.R.; Wolff, R.A.; Varadhachary, G.R.; Javle, M.M.; Shroff, R.T.; Ho, L.; Fogelman, D.R.; Raghav, K.P.S.; et al. Modified FOLFIRINOX in pancreatic cancer patients Age 75 or older. Pancreatology 2020, 20, 501–504. [Google Scholar] [CrossRef]
- Conroy, T.; Hammel, P.; Hebbar, M.; Ben Abdelghani, M.; Wei, A.C.; Raoul, J.L.; Chone, L.; Francois, E.; Artru, P.; Biagi, J.J.; et al. FOLFIRINOX or Gemcitabine as Adjuvant Therapy for Pancreatic Cancer. N. Engl. J. Med. 2018, 379, 2395–2406. [Google Scholar] [CrossRef]
- McCann, S.E.; Freudenheim, J.L.; Marshall, J.R.; Graham, S. Risk of human ovarian cancer is related to dietary intake of selected nutrients, phytochemicals and food groups. J. Nutr. 2003, 133, 1937–1942. [Google Scholar] [CrossRef]
- Sung, Y.Y.; Lee, A.Y.; Kim, H.K. Forsythia suspensa fruit extracts and the constituent matairesinol confer anti-allergic effects in an allergic dermatitis mouse model. J. Ethnopharmacol. 2016, 187, 49–56. [Google Scholar] [CrossRef]
- Bel Mabrouk, S.; Reis, M.; Sousa, M.L.; Ribeiro, T.; Almeida, J.R.; Pereira, S.; Antunes, J.; Rosa, F.; Vasconcelos, V.; Achour, L.; et al. The Marine Seagrass Halophila stipulacea as a Source of Bioactive Metabolites against Obesity and Biofouling. Mar. Drugs 2020, 18, 88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, P.; Huang, M.W.; Xiao, C.X.; Long, F.; Wang, Y.; Liu, S.Y.; Jia, W.W.; Wu, W.J.; Yang, D.; Hu, J.F.; et al. Matairesinol Suppresses Neuroinflammation and Migration Associated with Src and ERK1/2-NF-kappaB Pathway in Activating BV2 Microglia. Neurochem. Res. 2017, 42, 2850–2860. [Google Scholar] [CrossRef] [PubMed]
- Choi, S.W.; Park, K.I.; Yeon, J.T.; Ryu, B.J.; Kim, K.J.; Kim, S.H. Anti-osteoclastogenic activity of matairesinol via suppression of p38/ERK-NFATc1 signaling axis. BMC Complement. Altern. Med. 2014, 14, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peuhu, E.; Rivero-Muller, A.; Stykki, H.; Torvaldson, E.; Holmbom, T.; Eklund, P.; Unkila, M.; Sjoholm, R.; Eriksson, J.E. Inhibition of Akt signaling by the lignan matairesinol sensitizes prostate cancer cells to TRAIL-induced apoptosis. Oncogene 2010, 29, 898–908. [Google Scholar] [CrossRef] [Green Version]
- Lee, B.; Kim, K.H.; Jung, H.J.; Kwon, H.J. Matairesinol inhibits angiogenesis via suppression of mitochondrial reactive oxygen species. Biochem. Biophys. Res. Commun. 2012, 421, 76–80. [Google Scholar] [CrossRef]
- Mahajan, M.; Suryavanshi, S.; Bhowmick, S.; Alasmary, F.A.; Almutairi, T.M.; Islam, M.A.; Kaul-Ghanekar, R. Matairesinol, an active constituent of HC9 polyherbal formulation, exhibits HDAC8 inhibitory and anticancer activity. Biophys. Chem. 2021, 273, 106588. [Google Scholar] [CrossRef]
- Chang, H.; Wang, Y.; Gao, X.; Song, Z.; Awale, S.; Han, N.; Liu, Z.; Yin, J. Lignans from the root of Wikstroemia indica and their cytotoxic activity against PANC-1 human pancreatic cancer cells. Fitoterapia 2017, 121, 31–37. [Google Scholar] [CrossRef]
- Smith, S.J.; Li, C.M.; Lingeman, R.G.; Hickey, R.J.; Liu, Y.L.; Malkas, L.H.; Raoof, M. Molecular Targeting of Cancer-Associated PCNA Interactions in Pancreatic Ductal Adenocarcinoma Using a Cell-Penetrating Peptide. Mol. Ther. Oncolytics 2020, 17, 250–256. [Google Scholar] [CrossRef]
- Liou, G.Y.; Storz, P. Reactive oxygen species in cancer. Free Radic. Res. 2010, 44, 479–496. [Google Scholar] [CrossRef] [Green Version]
- Delierneux, C.; Kouba, S.; Shanmughapriya, S.; Potier-Cartereau, M.; Trebak, M.; Hempel, N. Mitochondrial Calcium Regulation of Redox Signaling in Cancer. Cells 2020, 9, 432. [Google Scholar] [CrossRef] [Green Version]
- Quinonero, F.; Mesas, C.; Doello, K.; Cabeza, L.; Perazzoli, G.; Jimenez-Luna, C.; Rama, A.R.; Melguizo, C.; Prados, J. The challenge of drug resistance in pancreatic ductal adenocarcinoma: A current overview. Cancer Biol. Med. 2019, 16, 688–699. [Google Scholar] [CrossRef] [PubMed]
- Wu, Q.; Wang, Y.; Li, Q. Matairesinol exerts anti-inflammatory and antioxidant effects in sepsis-mediated brain injury by repressing the MAPK and NF-kappaB pathways through up-regulating AMPK. Aging 2021, 13, 23780–23795. [Google Scholar] [CrossRef] [PubMed]
- Yatkin, E.; Polari, L.; Laajala, T.D.; Smeds, A.; Eckerman, C.; Holmbom, B.; Saarinen, N.M.; Aittokallio, T.; Makela, S.I. Novel Lignan and stilbenoid mixture shows anticarcinogenic efficacy in preclinical PC-3M-luc2 prostate cancer model. PLoS ONE 2014, 9, e93764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoo, J.H.; Lee, H.J.; Kang, K.; Jho, E.H.; Kim, C.Y.; Baturen, D.; Tunsag, J.; Nho, C.W. Lignans inhibit cell growth via regulation of Wnt/beta-catenin signaling. Food Chem. Toxicol. 2010, 48, 2247–2252. [Google Scholar] [CrossRef]
- Strzalka, W.; Ziemienowicz, A. Proliferating cell nuclear antigen (PCNA): A key factor in DNA replication and cell cycle regulation. Ann. Bot. 2011, 107, 1127–1140. [Google Scholar] [CrossRef] [Green Version]
- Bras, M.; Queenan, B.; Susin, S.A. Programmed cell death via mitochondria: Different modes of dying. Biochemistry 2005, 70, 231–239. [Google Scholar] [CrossRef]
- Pinton, P.; Giorgi, C.; Siviero, R.; Zecchini, E.; Rizzuto, R. Calcium and apoptosis: ER-mitochondria Ca2+ transfer in the control of apoptosis. Oncogene 2008, 27, 6407–6418. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Ding, Y.; Ye, N.; Wild, C.; Chen, H.; Zhou, J. Direct Activation of Bax Protein for Cancer Therapy. Med. Res. Rev. 2016, 36, 313–341. [Google Scholar] [CrossRef]
- Tait, S.W.; Green, D.R. Mitochondria and cell death: Outer membrane permeabilization and beyond. Nat. Rev. Mol. Cell Biol. 2010, 11, 621–632. [Google Scholar] [CrossRef]
- Stojak, M.; Mazur, L.; Opydo-Chanek, M.; Lukawska, M.; Oszczapowicz, I. In vitro induction of apoptosis and necrosis by new derivatives of daunorubicin. AntiCancer Res. 2013, 33, 4439–4443. [Google Scholar]
- Roderick, H.L.; Cook, S.J. Ca2+ signalling checkpoints in cancer: Remodelling Ca2+ for cancer cell proliferation and survival. Nat. Rev. Cancer 2008, 8, 361–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gorlach, A.; Bertram, K.; Hudecova, S.; Krizanova, O. Calcium and ROS: A mutual interplay. Redox Biol. 2015, 6, 260–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brookes, P.S.; Yoon, Y.; Robotham, J.L.; Anders, M.W.; Sheu, S.S. Calcium, ATP, and ROS: A mitochondrial love-hate triangle. Am. J. Physiol. Cell Physiol. 2004, 287, C817–C833. [Google Scholar] [CrossRef]
- Giorgi, C.; Baldassari, F.; Bononi, A.; Bonora, M.; De Marchi, E.; Marchi, S.; Missiroli, S.; Patergnani, S.; Rimessi, A.; Suski, J.M.; et al. Mitochondrial Ca2+ and apoptosis. Cell Calcium 2012, 52, 36–43. [Google Scholar] [CrossRef] [Green Version]
- Azimi, I.; Roberts-Thomson, S.J.; Monteith, G.R. Calcium influx pathways in breast cancer: Opportunities for pharmacological intervention. Br. J. Pharmacol. 2014, 171, 945–960. [Google Scholar] [CrossRef] [Green Version]
- Tsai, J.H.; Yang, J. Epithelial-mesenchymal plasticity in carcinoma metastasis. Genes Dev. 2013, 27, 2192–2206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Huang, S.; Sun, Y.L. Epithelial-Mesenchymal Transition in Pancreatic Cancer: A Review. Biomed Res. Int. 2017, 2017, 2646148. [Google Scholar] [CrossRef] [Green Version]
- Palamaris, K.; Felekouras, E.; Sakellariou, S. Epithelial to Mesenchymal Transition: Key Regulator of Pancreatic Ductal Adenocarcinoma Progression and Chemoresistance. Cancers 2021, 13, 5532. [Google Scholar] [CrossRef]
- Chen, Y.; Peng, S.; Cen, H.; Lin, Y.; Huang, C.; Chen, Y.; Shan, H.; Su, Y.; Zeng, L. MicroRNA hsa-miR-623 directly suppresses MMP1 and attenuates IL-8-induced metastasis in pancreatic cancer. Int. J. Oncol. 2019, 55, 142–156. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.A.; Li, C.F.; Huang, R.C.; Li, Y.H.; Liou, J.P.; Tsai, S.J. Suppression of Extracellular Vesicle VEGF-C-mediated Lymphangiogenesis and Pancreatic Cancer Early Dissemination By a Selective HDAC1/2 Inhibitor. Mol. Cancer Ther. 2021, 20, 1550–1560. [Google Scholar] [CrossRef]
- Zhao, X.; Liu, Z.; Ren, Z.; Wang, H.; Wang, Z.; Zhai, J.; Cao, D.; Lyu, S.; Li, L.; Lang, R.; et al. Triptolide inhibits pancreatic cancer cell proliferation and migration via down-regulating PLAU based on network pharmacology of Tripterygium wilfordii Hook F. Eur. J. Pharmacol. 2020, 880, 173225. [Google Scholar] [CrossRef] [PubMed]
- Buscail, L.; Bournet, B.; Cordelier, P. Role of oncogenic KRAS in the diagnosis, prognosis and treatment of pancreatic cancer. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 153–168. [Google Scholar] [CrossRef] [PubMed]
- Nevala-Plagemann, C.; Hidalgo, M.; Garrido-Laguna, I. From state-of-the-art treatments to novel therapies for advanced-stage pancreatic cancer. Nat. Rev. Clin. Oncol. 2020, 17, 108–123. [Google Scholar] [CrossRef]
- Lugowska, I.; Kosela-Paterczyk, H.; Kozak, K.; Rutkowski, P. Trametinib: A MEK inhibitor for management of metastatic melanoma. Onco Targets Ther. 2015, 8, 2251–2259. [Google Scholar] [PubMed] [Green Version]
- Asati, V.; Mahapatra, D.K.; Bharti, S.K. K-Ras and its inhibitors towards personalized cancer treatment: Pharmacological and structural perspectives. Eur. J. Med. Chem. 2017, 125, 299–314. [Google Scholar] [CrossRef]
- Hayes, T.K.; Neel, N.F.; Hu, C.; Gautam, P.; Chenard, M.; Long, B.; Aziz, M.; Kassner, M.; Bryant, K.L.; Pierobon, M.; et al. Long-Term ERK Inhibition in KRAS-Mutant Pancreatic Cancer Is Associated with MYC Degradation and Senescence-like Growth Suppression. Cancer Cell 2016, 29, 75–89. [Google Scholar] [CrossRef] [Green Version]
- Kinsey, C.G.; Camolotto, S.A.; Boespflug, A.M.; Guillen, K.P.; Foth, M.; Truong, A.; Schuman, S.S.; Shea, J.E.; Seipp, M.T.; Yap, J.T.; et al. Protective autophagy elicited by RAF-->MEK-->ERK inhibition suggests a treatment strategy for RAS-driven cancers. Nat. Med. 2019, 25, 620–627. [Google Scholar] [CrossRef]
- Bryant, K.L.; Stalnecker, C.A.; Zeitouni, D.; Klomp, J.E.; Peng, S.; Tikunov, A.P.; Gunda, V.; Pierobon, M.; Waters, A.M.; George, S.D.; et al. Author Correction: Combination of ERK and autophagy inhibition as a treatment approach for pancreatic cancer. Nat. Med. 2020, 26, 982. [Google Scholar] [CrossRef]
- Sharma, P.R.; Shanmugavel, M.; Saxena, A.K.; Qazi, G.N. Induction of apoptosis by a synergistic lignan composition from Cedrus deodara in human cancer cells. Phytother. Res. 2008, 22, 1587–1594. [Google Scholar] [CrossRef]
- Su, S.; Cheng, X.; Wink, M. Natural lignans from Arctium lappa modulate P-glycoprotein efflux function in multidrug resistant cancer cells. Phytomedicine 2015, 22, 301–307. [Google Scholar] [CrossRef]
- Caparello, C.; Meijer, L.L.; Garajova, I.; Falcone, A.; Le Large, T.Y.; Funel, N.; Kazemier, G.; Peters, G.J.; Vasile, E.; Giovannetti, E. FOLFIRINOX and translational studies: Towards personalized therapy in pancreatic cancer. World J. Gastroenterol. 2016, 22, 6987–7005. [Google Scholar] [CrossRef]
- Longley, D.B.; Harkin, D.P.; Johnston, P.G. 5-fluorouracil: Mechanisms of action and clinical strategies. Nat. Rev. Cancer 2003, 3, 330–338. [Google Scholar] [CrossRef]
- Oman, M.; Wettergren, Y.; Odin, E.; Westermark, S.; Naredi, P.; Hemmingsson, O.; Taflin, H. Pharmacokinetics of preoperative intraperitoneal 5-FU in patients with pancreatic ductal adenocarcinoma. Cancer Chemother. Pharmacol. 2021, 88, 619–631. [Google Scholar] [CrossRef]
- Bae, H.; Lee, W.; Song, J.; Hong, T.; Kim, M.H.; Ham, J.; Song, G.; Lim, W. Polydatin Counteracts 5-Fluorouracil Resistance by Enhancing Apoptosis via Calcium Influx in Colon Cancer. Antioxidants 2021, 10, 1477. [Google Scholar] [CrossRef]
- Liu, Q.; Yin, X.F.; Languino, L.R.; Altieri, D.C. Evaluation of Drug Combination Effect Using a Bliss Independence Dose-Response Surface Model. Stat. Biopharm. Res. 2018, 10, 112–122. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibody | Catalog Number | Supplier | Dilution |
|---|---|---|---|
| PCNA | 10205-2-AP | Proteintech | 1:2000 |
| BAX | 50599-2-Ig | Proteintech | 1:2000 |
| p-JNK (Thr183/Tyr185) | 4668 | Cell Signaling Technology | 1:1000 |
| JNK | 9252 | Cell Signaling Technology | 1:1000 |
| p-ERK1/2 (Thr202/Tyr204) | 9101 | Cell Signaling Technology | 1:1000 |
| ERK1/2 | 4695 | Cell Signaling Technology | 1:1000 |
| p-P38 (Thr180/Tyr182) | 4511 | Cell Signaling Technology | 1:1000 |
| P38 | 9212 | Cell Signaling Technology | 1:1000 |
| p-AKT (Ser473) | 4060 | Cell Signaling Technology | 1:1000 |
| AKT | 9272 | Cell Signaling Technology | 1:1000 |
| β-actin | sc-47778 | Santa Cruz Biotechnology | 1:1000 |
| Gene | Size (bp) | GenBank Accession No. | Primer Sequence (5′ → 3′) |
|---|---|---|---|
| forkhead box protein M1 (FOXM1) | 104 | NM_001243088.2 | F: AGTCACACCCTAGCCACTGC |
| R: ACCATTGCCTTTGTTGTTCC | |||
| matrix metallopeptidase 1 (MMP1) | 147 | NM_001145938.2 | F: GGGAGCAAACACATCTGACC |
| R: CTGCTTGACCCTCAGAGACC | |||
| plasminogen activator, urokinase (PLAU) | 139 | NM_002658.6 | F: TGTGAGATCACTGGCTTTGG |
| R: TTTTGGTGGTGACTTCAGAG | |||
| vascular endothelial growth factor C (VEGFC) | 116 | NM_005429.5 | F: AGTTCCACCACCAAACATGC |
| R: CCAATATGAAGGGACACAACG | |||
| glyceraldehyde-3-phosphate dehydrogenase (GAPDH) | 149 | NM_001256799.3 | F: GGCTCTCCAGAACATCATCC |
| R: TTTCTAGACGGCAGGTCAGG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, W.; Song, G.; Bae, H. Matairesinol Induces Mitochondrial Dysfunction and Exerts Synergistic Anticancer Effects with 5-Fluorouracil in Pancreatic Cancer Cells. Mar. Drugs 2022, 20, 473. https://doi.org/10.3390/md20080473
Lee W, Song G, Bae H. Matairesinol Induces Mitochondrial Dysfunction and Exerts Synergistic Anticancer Effects with 5-Fluorouracil in Pancreatic Cancer Cells. Marine Drugs. 2022; 20(8):473. https://doi.org/10.3390/md20080473
Chicago/Turabian StyleLee, Woonghee, Gwonhwa Song, and Hyocheol Bae. 2022. "Matairesinol Induces Mitochondrial Dysfunction and Exerts Synergistic Anticancer Effects with 5-Fluorouracil in Pancreatic Cancer Cells" Marine Drugs 20, no. 8: 473. https://doi.org/10.3390/md20080473
APA StyleLee, W., Song, G., & Bae, H. (2022). Matairesinol Induces Mitochondrial Dysfunction and Exerts Synergistic Anticancer Effects with 5-Fluorouracil in Pancreatic Cancer Cells. Marine Drugs, 20(8), 473. https://doi.org/10.3390/md20080473