
Novel Labdane Diterpenes-Based Synthetic Derivatives: Identification of a Bifunctional Vasodilator That Inhibits CaV1.2 and Stimulates KCa1.1 Channels

 ,
,  ,
,  ,
,  , , and
, , and 
Abstract
:1. Introduction
2. Results
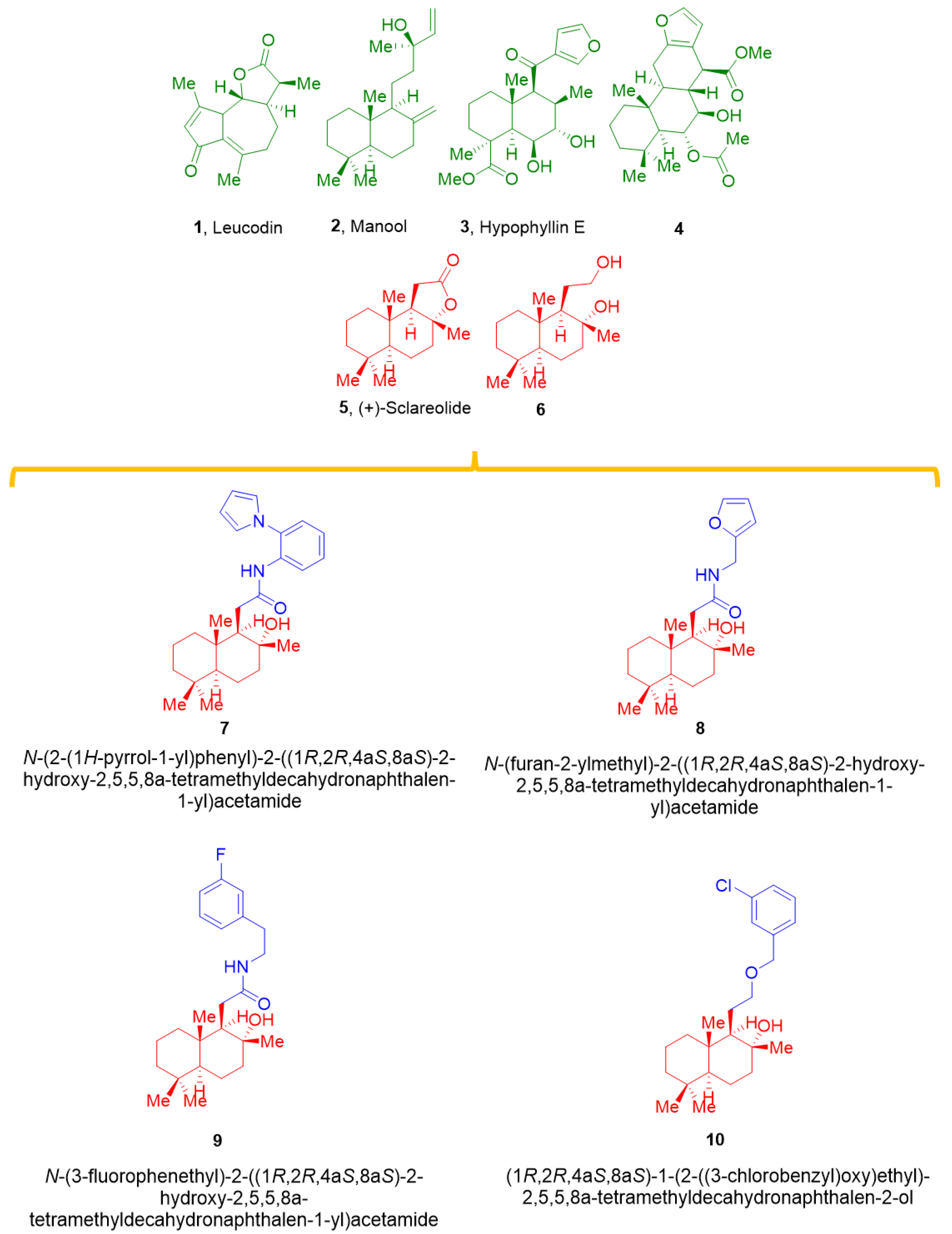
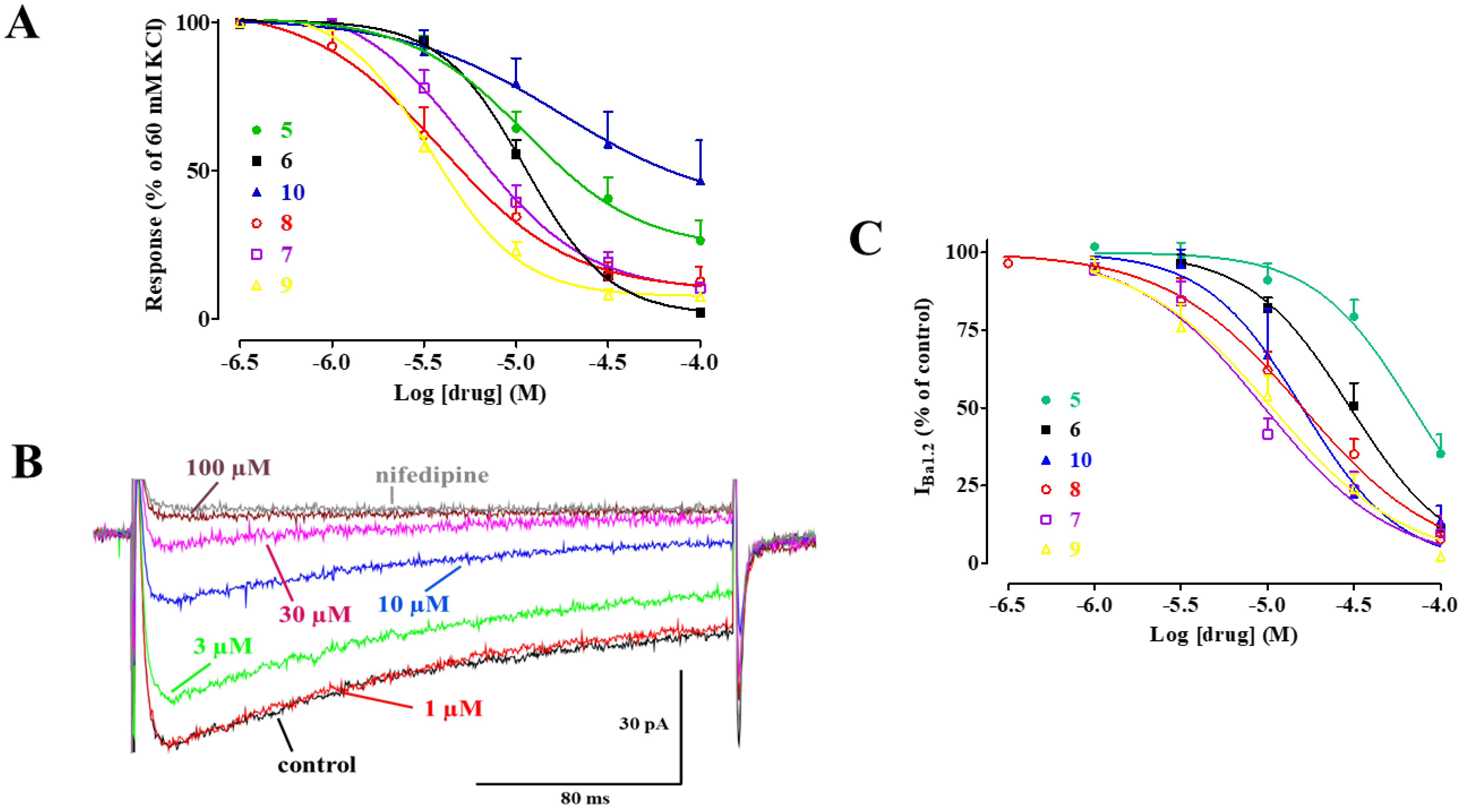
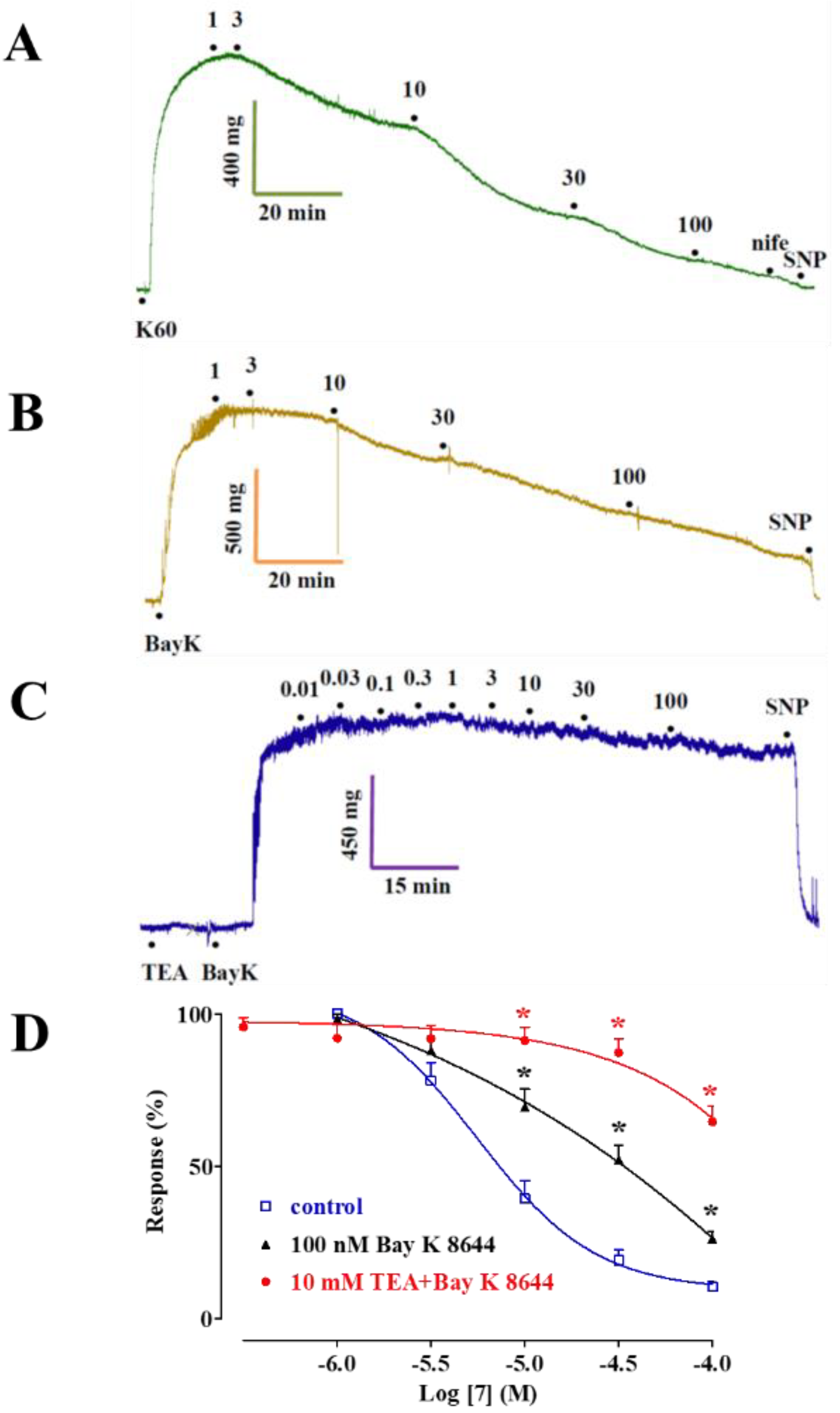
2.1. Effect of 5 and Its Derivatives on High KCl-Induced Contraction and Ba2+ Current through CaV1.2 Channels (IBa1.2)
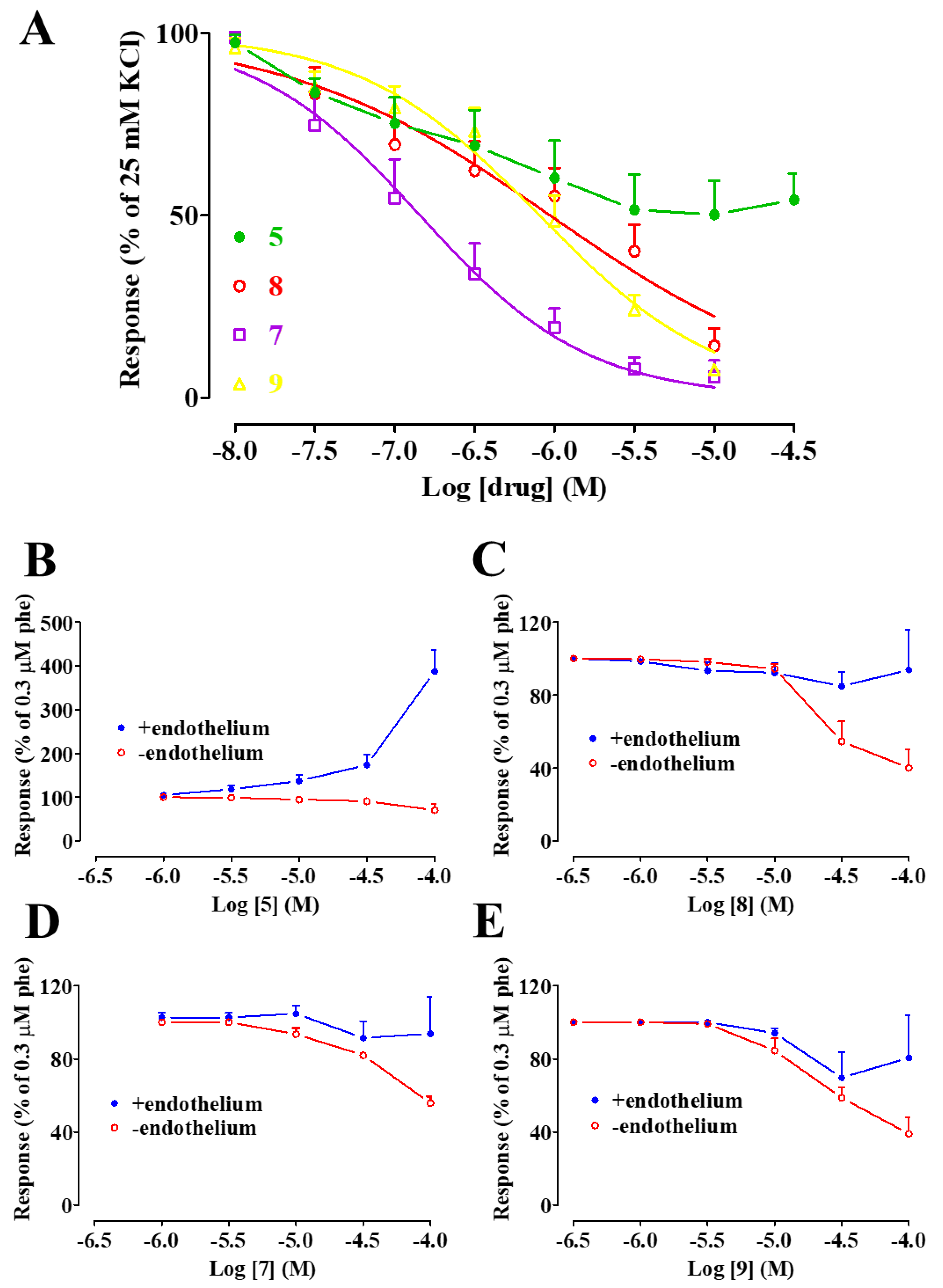
2.2. Effect of 5, 7, 8, and 9 on the Contraction Induced by Moderate KCl Concentrations
2.3. Effect of 5, 7, 8, and 9 on Phenylephrine-Induced Contraction
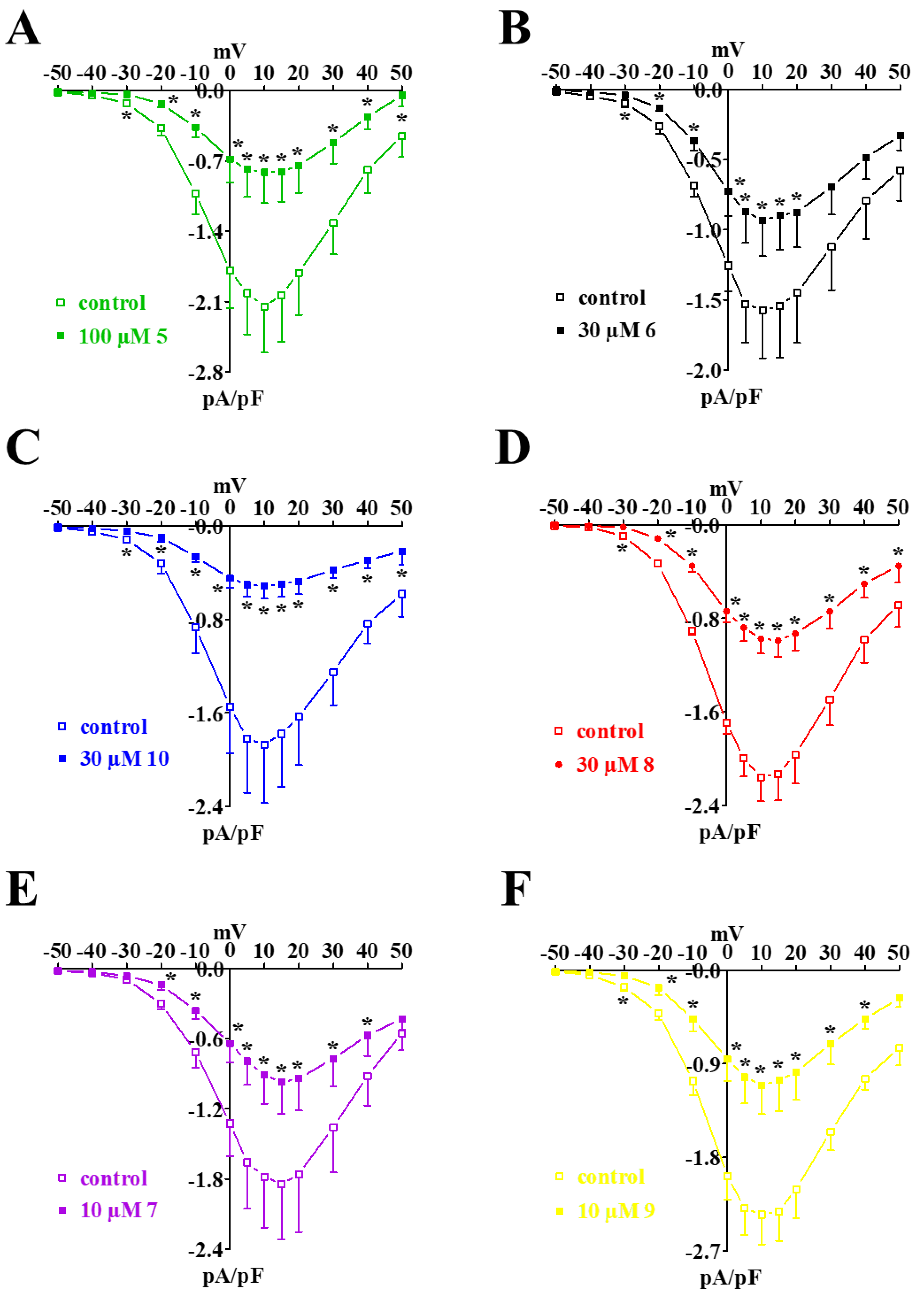
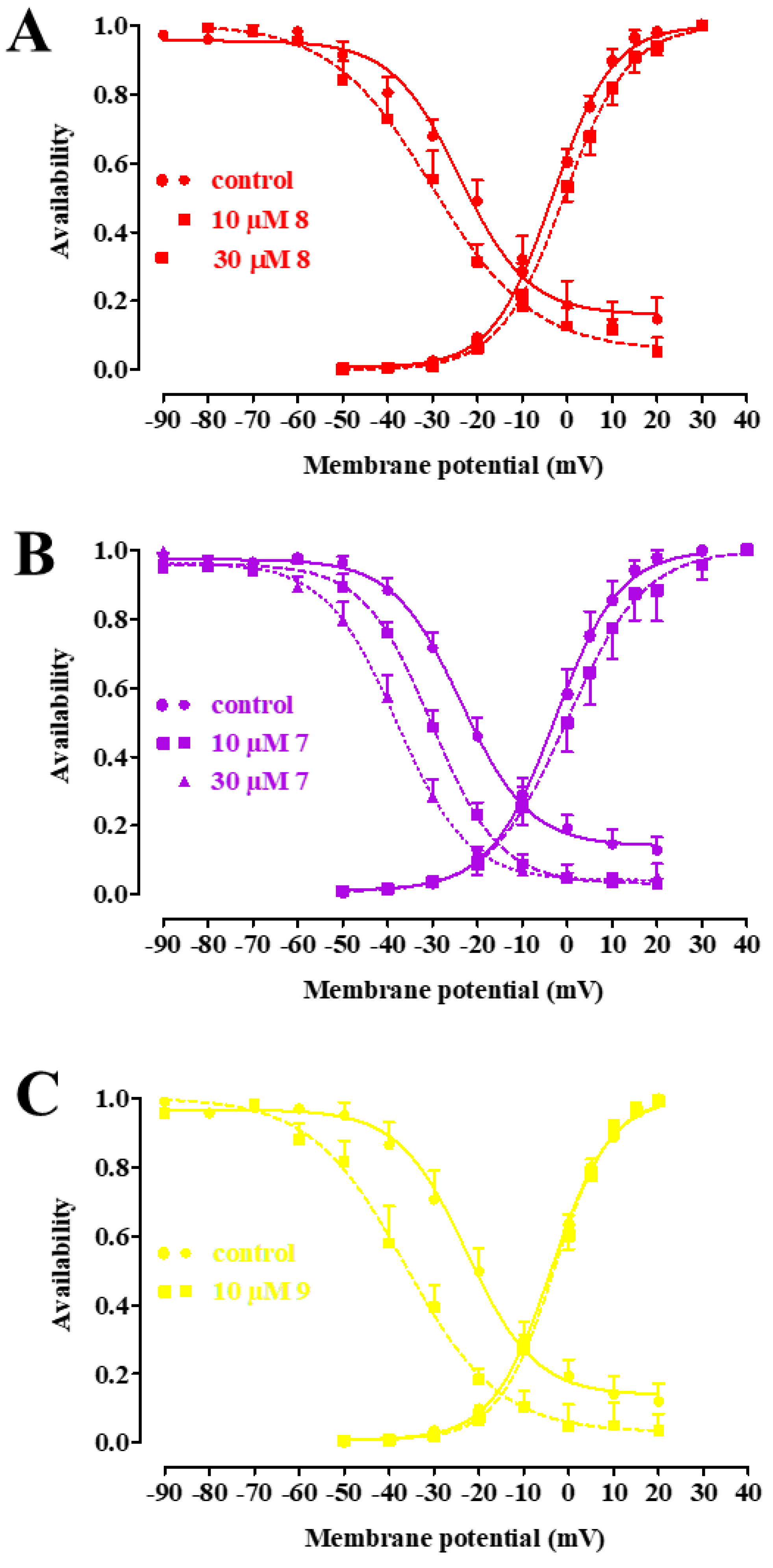
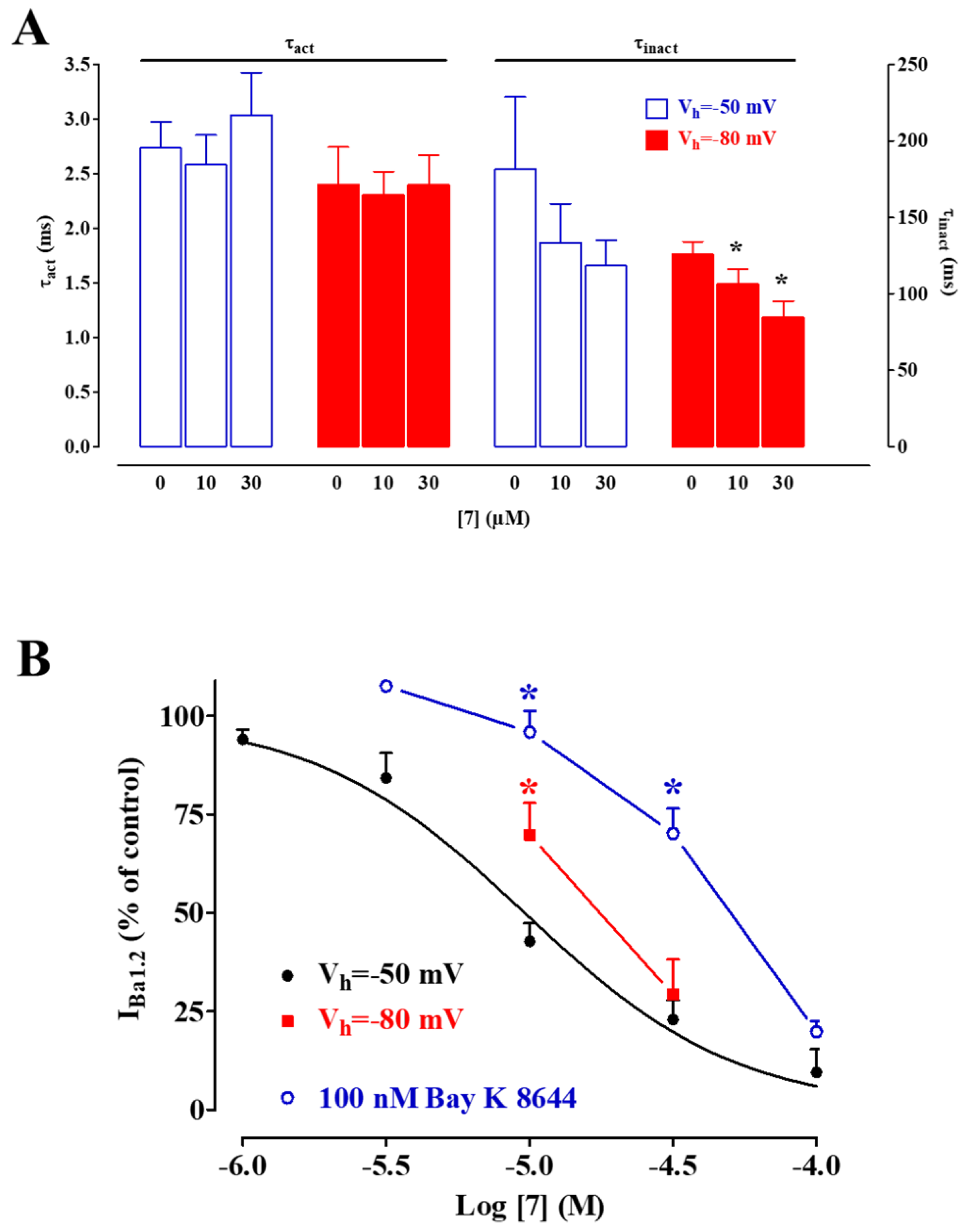
2.4. Biophysical and Pharmacological Analysis of the Effect of 7, 8, and 9 on CaV1.2 Channels
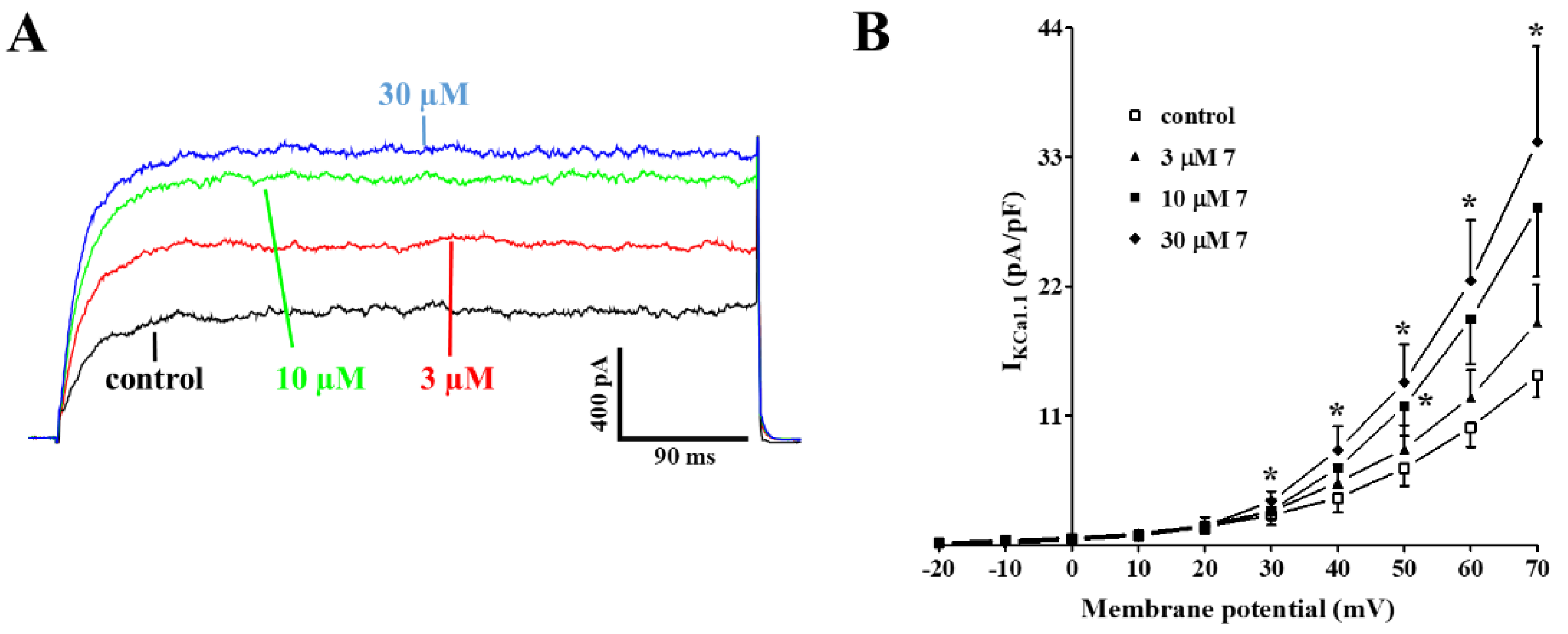
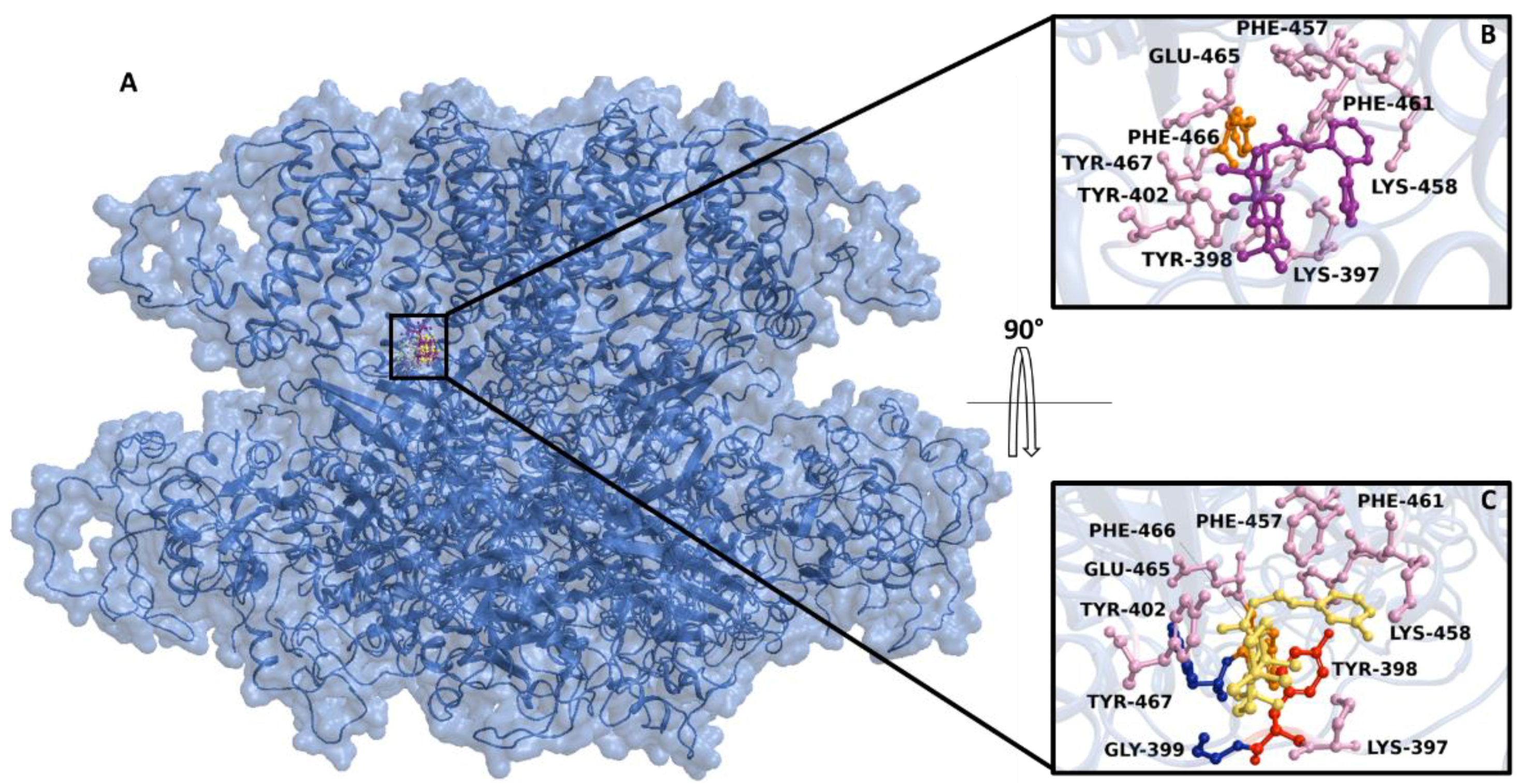
2.5. Effect of 7 on KCa1.1 Channels
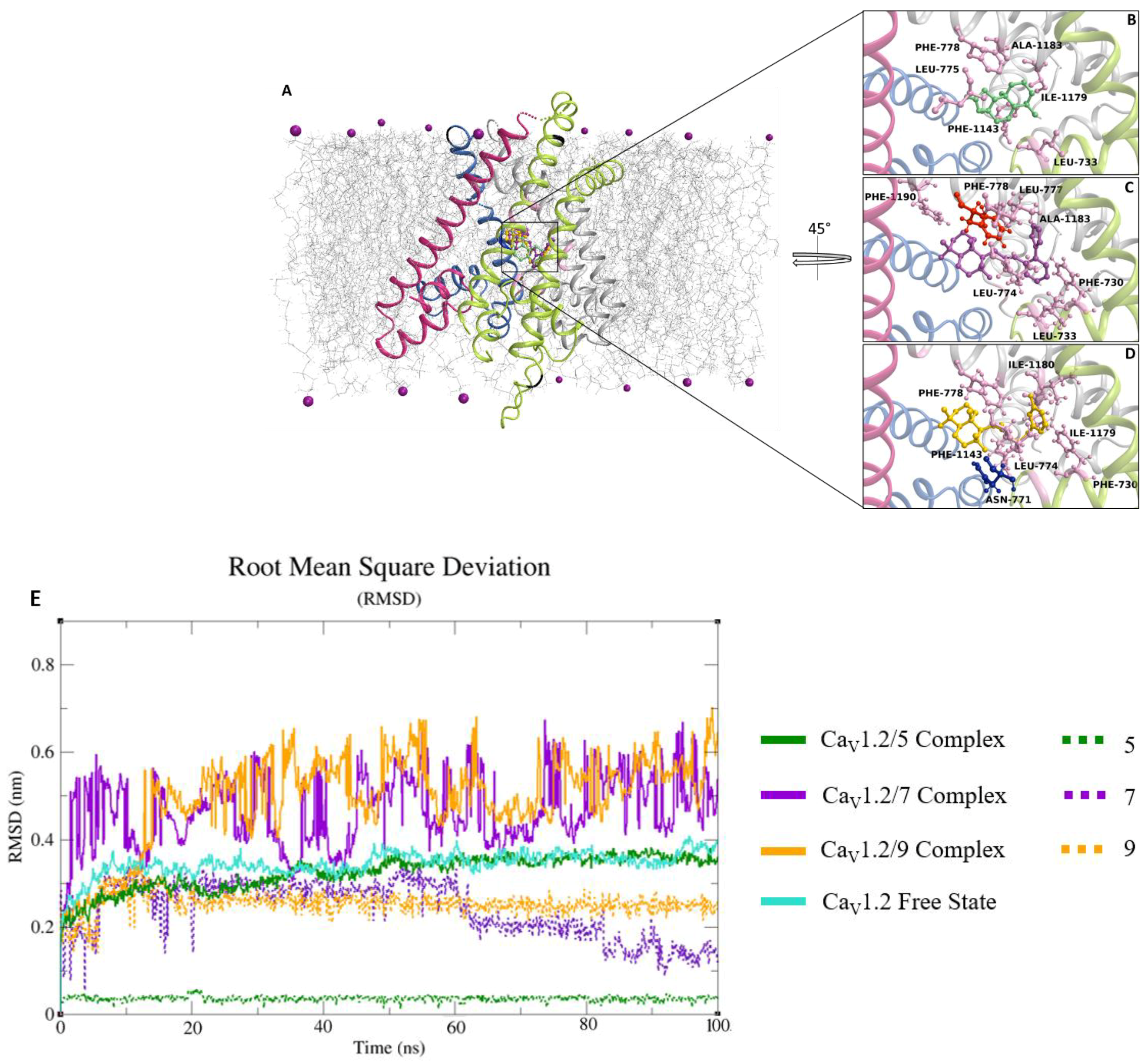
2.6. Docking and Classical Molecular Dynamics Simulations
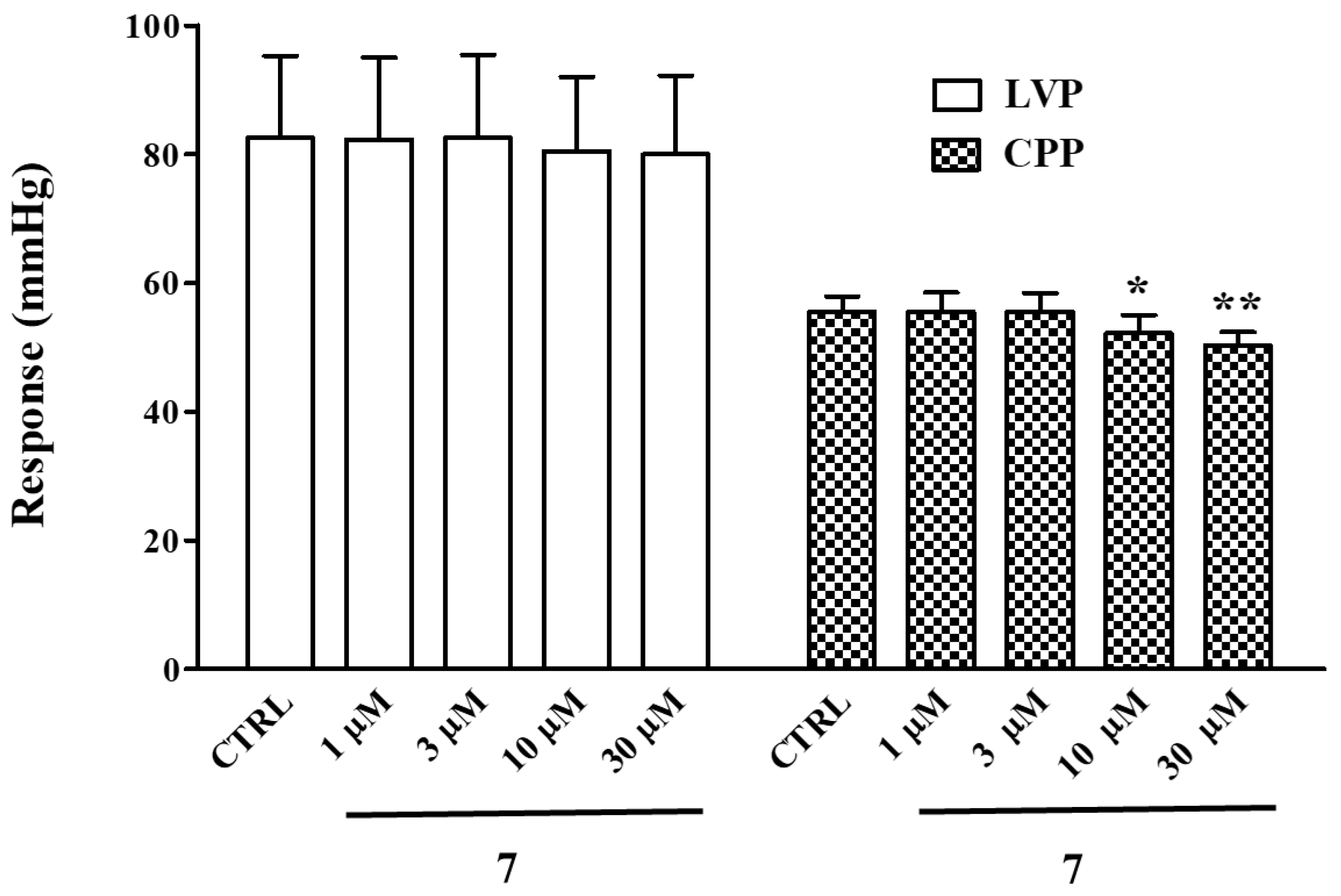
2.7. Effect of 7 on Langendorff Perfused Rat Heart
3. Discussion
- chemical modification of 5 generated effective vasodilating agents that were more potent CaV1.2 channel blockers than the parent compound;
- their Ca2+ antagonism seems to be related to the stabilization of the CaV1.2 channel inactivated state;
- a functional endothelium is not crucial for, and in some cases counteracts, their activity, though to a lesser extent as compared to 5;
- 7, besides blocking CaV1.2 channels, is also capable of stimulating the KCa1.1 channels;
- 7 is a negative chronotropic and coronary vasodilating agent.
4. Materials and Methods
4.1. Chemicals
4.2. Animal Care Statement
4.3. Aorta Ring Preparation
4.4. Effect of Sclareolide and Its Derivatives 7–10 on Phenylephrine- or High KCl-Induced Contraction
4.5. Cell Isolation Procedure
4.6. Whole-Cell Patch-Clamp Recordings
4.7. Ba2+ Current through CaV1.2 Channel (IBa1.2) Recordings
4.8. K+ Current through KCa1.1 Channel (IKCa1.1) Recordings
4.9. Isolated Rat Heart Preparation and Perfusion
4.10. Statistical Analysis
4.11. In Silico Methods: Structural Resource
4.12. Docking and Classical Molecular Dynamics Simulations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oparil, S.; Acelajado, M.C.; Bakris, G.L.; Berlowitz, D.R.; Cífková, R.; Dominiczak, A.F.; Grassi, G.; Jordan, J.; Poulter, N.R.; Rodgers, A.; et al. Hypertension. Nat. Rev. Dis. Prim. 2018, 4, 18014. [Google Scholar] [CrossRef]
- Tang, F.; Yan, H.L.; Wang, L.X.; Xu, J.F.; Peng, C.; Ao, H.; Tan, Y.Z. Review of Natural Resources With Vasodilation: Traditional Medicinal Plants, Natural Products, and Their Mechanism and Clinical Efficacy. Front. Pharmacol. 2021, 12, 1–24. [Google Scholar] [CrossRef]
- Carullo, G.; Mazzotta, S.; Koch, A.; Hartmann, K.M.; Friedrich, O.; Gilbert, D.F.; Vega-Holm, M.; Schneider-Stock, R.; Aiello, F. New oleoyl hybrids of natural antioxidants: Synthesis and in vitro evaluation as inducers of apoptosis in colorectal cancer cells. Antioxidants 2020, 9, 1077. [Google Scholar] [CrossRef] [PubMed]
- Carullo, G.; Sciubba, F.; Governa, P.; Mazzotta, S.; Frattaruolo, L.; Grillo, G.; Cappello, A.R.; Cravotto, G.; Di Cocco, M.E.; Aiello, F. Mantonico and pecorello grape seed extracts: Chemical characterization and evaluation of in vitro wound-healing and anti-inflammatory activities. Pharmaceuticals 2020, 13, 97. [Google Scholar] [CrossRef] [PubMed]
- Mazzotta, S.; Governa, P.; Borgonetti, V.; Marcolongo, P.; Nanni, C.; Gamberucci, A.; Manetti, F.; Pessina, F.; Carullo, G.; Brizzi, A.; et al. Pinocembrin and its linolenoyl ester derivative induce wound healing activity in HaCaT cell line potentially involving a GPR120/FFA4 mediated pathway. Bioorg. Chem. 2021, 108, 104657. [Google Scholar] [CrossRef] [PubMed]
- Aiello, F.; Restuccia, D.; Spizzirri, U.G.; Carullo, G.; Leporini, M.; Loizzo, M.R. Improving kefir bioactive properties by functional enrichment with plant and agro-food waste extracts. Fermentation 2020, 6, 83. [Google Scholar] [CrossRef]
- Carullo, G.; Spizzirri, U.G.; Loizzo, M.R.; Leporini, M.; Sicari, V.; Aiello, F.; Restuccia, D. Valorization of red grape (Vitis vinifera cv. Sangiovese) pomace as functional food ingredient. Ital. J. Food Sci. 2020, 32, 367–385. [Google Scholar] [CrossRef]
- Spizzirri, U.G.; Carullo, G.; De Cicco, L.; Crispini, A.; Scarpelli, F.; Restuccia, D.; Aiello, F. Synthesis and characterization of a (+)-catechin and L-(+)-ascorbic acid cocrystal as a new functional ingredient for tea drinks. Heliyon 2019, 5, e02291. [Google Scholar] [CrossRef]
- Mazzotta, S.; Berastegui-Cabrera, J.; Carullo, G.; Vega-Holm, M.; Carretero-Ledesma, M.; Mendolia, L.; Aiello, F.; Iglesias-Guerra, F.; Pachón, J.; Vega-Pérez, J.M.; et al. Serinol-Based Benzoic Acid Esters as New Scaffolds for the Development of Adenovirus Infection Inhibitors: Design, Synthesis, and in Vitro Biological Evaluation. ACS Infect. Dis. 2021, 7, 1433–1444. [Google Scholar] [CrossRef]
- Carullo, G.; Governa, P.; Leo, A.; Gallelli, L.; Citraro, R.; Cione, E.; Caroleo, M.C.; Biagi, M.; Aiello, F.; Manetti, F. Quercetin-3-Oleate Contributes to Skin Wound Healing Targeting FFA1/GPR40. ChemistrySelect 2019, 4, 8429–8433. [Google Scholar] [CrossRef]
- Pozzetti, L.; Ferrara, F.; Marotta, L.; Gemma, S.; Butini, S.; Benedusi, M.; Fusi, F.; Ahmed, A.; Pomponi, S.; Ferrari, S.; et al. Extra Virgin Olive Oil Extracts of Indigenous Southern Tuscany Cultivar Act as Anti-Inflammatory and Vasorelaxant Nutraceuticals. Antioxidants 2022, 11, 437. [Google Scholar] [CrossRef]
- Carullo, G.; Ahmed, A.; Fusi, F.; Sciubba, F.; Di Cocco, M.E.; Restuccia, D.; Spizzirri, U.G.; Saponara, S.; Aiello, F. Vasorelaxant Effects Induced by Red Wine and Pomace Extracts of Magliocco Dolce cv. Pharmaceuticals 2020, 13, 87. [Google Scholar] [CrossRef] [PubMed]
- Carullo, G.; Durante, M.; Sciubba, F.; Restuccia, D.; Spizzirri, U.G.; Ahmed, A.; Di Cocco, M.E.; Saponara, S.; Aiello, F.; Fusi, F. Vasoactivity of Mantonico and Pecorello grape pomaces on rat aorta rings: An insight into nutraceutical development. J. Funct. Foods 2019, 57, 328–334. [Google Scholar] [CrossRef]
- Fusi, F.; Cavalli, M.; Mulholland, D.; Crouch, N.; Coombes, P.; Dawson, G.; Bova, S.; Sgaragli, G.; Saponara, S. Cardamonin is a bifunctional vasodilator that inhibits Ca(v)1.2 current and stimulates K(Ca)1.1 current in rat tail artery myocytes. J. Pharmacol. Exp. Ther. 2010, 332, 531–540. [Google Scholar] [CrossRef] [PubMed]
- Tirapelli, C.R.; Ambrosio, S.R.; De Oliveira, A.M.; Tostes, R.C. Hypotensive action of naturally occurring diterpenes: A therapeutic promise for the treatment of hypertension. Fitoterapia 2010, 81, 690–702. [Google Scholar] [CrossRef] [PubMed]
- Leitolis, A.; Amoah, S.K.S.; Biavatti, M.W.; da Silva-Santos, J.E. Sesquiterpene lactones from Hedyosmum brasiliense induce in vitro relaxation of rat aorta and corpus cavernosum. Rev. Bras. Farmacogn. 2016, 26, 363–368. [Google Scholar] [CrossRef]
- Simplicio, J.A.; Pernomian, L.; Simão, M.R.; Carnio, E.C.; Batalhão, M.E.; Ambrosio, S.R.; Tirapelli, C.R. Mechanisms underlying the vascular and hypotensive actions of the labdane ent-3-acetoxy-labda-8(17),13-dien-15-oic acid. Eur. J. Pharmacol. 2014, 726, 66–76. [Google Scholar] [CrossRef]
- Ribeiro, L.A.A.; Alencar Filho, E.B.; Coelho, M.C.; Silva, B.A. The vasorelaxant effect of 8(17),12E,14-labdatrien-18-oic acid involves stimulation of adenylyl cyclase and cAMP/PKA pathway: Evidences by pharmacological and molecular docking studies. Eur. J. Pharmacol. 2015, 764, 363–371. [Google Scholar] [CrossRef]
- Arias-Durán, L.; Estrada-Soto, S.; Hernández-Morales, M.; Millán-Pacheco, C.; Navarrete-Vázquez, G.; Villalobos-Molina, R.; Ibarra-Barajas, M.; Almanza-Pérez, J.C. Antihypertensive and vasorelaxant effect of leucodin and achillin isolated from Achillea millefolium through calcium channel blockade and NO production: In vivo, functional ex vivo and in silico studies. J. Ethnopharmacol. 2021, 273, 113948. [Google Scholar] [CrossRef]
- Monteiro, A.S.E.N.; Campos, D.R.; Albuquerque, A.A.S.; Evora, P.R.B.; Ferreira, L.G.; Celotto, A.C. Effect of diterpene manool on the arterial blood pressure and vascular reactivity in normotensive and hypertensive rats. Arq. Bras. Cardiol. 2020, 115, 669–677. [Google Scholar] [CrossRef]
- Wu, X.D.; Luo, D.; Tu, W.C.; Deng, Z.T.; Chen, X.J.; Su, J.; Ji, X.; Zhao, Q.S. Hypophyllins A-D, Labdane-Type Diterpenoids with Vasorelaxant Activity from Hypoestes phyllostachya “Rosea”. Org. Lett. 2016, 18, 6484–6487. [Google Scholar] [CrossRef] [PubMed]
- De Fátima Reis, C.; De Andrade, D.M.L.; Neves, B.J.; De Almeida Ribeiro Oliveira, L.; Pinho, J.F.; Da Silva, L.P.; Cruz, J.D.S.; Bara, M.T.F.; Andrade, C.H.; Rocha, M.L. Blocking the L-type Ca2+ channel (Cav 1.2) is the key mechanism for the vascular relaxing effect of Pterodon spp. and its isolated diterpene methyl-6α-acetoxy-7β-hydroxyvouacapan-17β-oate. Pharmacol. Res. 2015, 100, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Carullo, G.; Ahmed, A.; Trezza, A.; Spiga, O.; Brizzi, A.; Saponara, S.; Fusi, F.; Aiello, F. A multitarget semi-synthetic derivative of the flavonoid morin with improved in vitro vasorelaxant activity: Role of CaV1.2 and KCa1.1 channels. Biochem. Pharmacol. 2021, 185, 114429. [Google Scholar] [CrossRef] [PubMed]
- Carullo, G.; Ahmed, A.; Trezza, A.; Spiga, O.; Brizzi, A.; Saponara, S.; Fusi, F.; Aiello, F. Design, synthesis and pharmacological evaluation of ester-based quercetin derivatives as selective vascular KCa1.1 channel stimulators. Bioorg. Chem. 2020, 105, 104404. [Google Scholar] [CrossRef] [PubMed]
- Mazzotta, S.; Carullo, G.; Schiano Moriello, A.; Amodeo, P.; Di Marzo, V.; Vega-Holm, M.; Vitale, R.M.; Aiello, F.; Brizzi, A.; De Petrocellis, L. Design, Synthesis and In Vitro Experimental Validation of Novel TRPV4 Antagonists Inspired by Labdane Diterpenes. Mar. Drugs 2020, 18, 519. [Google Scholar] [CrossRef]
- Saponara, S.; Testai, L.; Iozzi, D.; Martinotti, E.; Martelli, A.; Chericoni, S.; Sgaragli, G.; Fusi, F.; Calderone, V. (+/−)-Naringenin as large conductance Ca(2+)-activated K+ (BKCa) channel opener in vascular smooth muscle cells. Br. J. Pharmacol. 2006, 149, 1013–1021. [Google Scholar] [CrossRef]
- Tykocki, N.R.; Boerman, E.M.; Jackson, W.F. Smooth muscle ion channels and regulation of vascular tone in resistance arteries and arterioles. Compr. Physiol. 2017, 7, 485–581. [Google Scholar] [CrossRef]
- Mugnai, P.; Durante, M.; Sgaragli, G.; Saponara, S.; Paliuri, G.; Bova, S.; Fusi, F. L-type Ca(2+) channel current characteristics are preserved in rat tail artery myocytes after one-day storage. Acta Physiol. (Oxf.) 2014, 211, 334–345. [Google Scholar] [CrossRef]
- Zhao, Y.; Huang, G.; Wu, J.; Wu, Q.; Gao, S.; Yan, Z.; Lei, J.; Yan, N. Molecular Basis for Ligand Modulation of a Mammalian Voltage-Gated Ca2+ Channel. Cell 2019, 177, 1495–1506. [Google Scholar] [CrossRef]
- Gessner, G.; Cui, Y.-M.; Otani, Y.; Ohwada, T.; Soom, M.; Hoshi, T.; Heinemann, S.H. Molecular mechanism of pharmacological activation of BK channels. Proc. Natl. Acad. Sci. USA 2012, 109, 3552–3557. [Google Scholar] [CrossRef]
- Fransen, P.; Van Hove, C.E.; Leloup, A.J.A.; Martinet, W.; De Meyer, G.R.Y.; Lemmens, K.; Bult, H.; Schrijvers, D.M. Dissecting out the complex Ca2+-mediated phenylephrine-induced contractions of mouse aortic segments. PLoS ONE 2015, 10, e0121634. [Google Scholar] [CrossRef] [PubMed]
- Prystowsky, E.N. The effects of slow channel blockers and beta blockers on atrioventricular nodal conduction. J. Clin. Pharmacol. 1988, 28, 6–21. [Google Scholar] [CrossRef] [PubMed]
- Stark, G.; Stark, U.; Tritthart, H.A. Modulation of cardiac impulse generation and conduction by nifedipine and verapamil analyzed by a refined surface ECG technique in Langendorff perfused guinea pig hearts. Basic Res. Cardiol. 1988, 83, 202–212. [Google Scholar] [CrossRef] [PubMed]
- Saponara, S.; Ferrara, A.; Gorelli, B.; Shah, A.; Kawase, M.; Motohashi, N.; Molnar, J.; Sgaragli, G.; Fusi, F. 3,5-dibenzoyl-4-(3-phenoxyphenyl)-1,4-dihydro-2,6-dimethylpyridine (DP7): A new multidrug resistance inhibitor devoid of effects on Langendorff-perfused rat heart. Eur. J. Pharmacol. 2007, 563, 160–163. [Google Scholar] [CrossRef]
- Fusi, F.; Marazova, K.; Pessina, F.; Gorelli, B.; Valoti, M.; Frosini, M.; Sgaragli, G. On the mechanisms of the antispasmodic action of some hindered phenols in rat aorta rings. Eur. J. Pharmacol. 2000, 394, 109–115. [Google Scholar] [CrossRef]
- Fusi, F.; Ferrara, A.; Zalatnai, A.; Molnar, J.; Sgaragli, G.; Saponara, S. Vascular activity of two silicon compounds, ALIS 409 and ALIS 421, novel multidrug-resistance reverting agents in cancer cells. Cancer Chemother. Pharmacol. 2008, 61, 443–451. [Google Scholar] [CrossRef]
- Saponara, S.; Durante, M.; Spiga, O.; Mugnai, P.; Sgaragli, G.; Huong, T.; Khanh, P.; Son, N.; Cuong, N.; Fusi, F. Functional, electrophysiological and molecular docking analysis of the modulation of Cav1.2 channels in rat vascular myocytes by murrayafoline A. Br. J. Pharmacol. 2016, 173, 292–304. [Google Scholar] [CrossRef]
- Budriesi, R.; Cosimelli, B.; Ioan, P.; Ugenti, M.P.; Carosati, E.; Frosini, M.; Fusi, F.; Spisani, R.; Saponara, S.; Cruciani, G.; et al. L-Type calcium channel blockers: From diltiazem to 1,2,4-oxadiazol-5-ones via thiazinooxadiazol-3-one derivatives. J. Med. Chem. 2009, 52, 2352–2362. [Google Scholar] [CrossRef]
- Fusi, F.; Manetti, F.; Durante, M.; Sgaragli, G.; Saponara, S. The vasodilator papaverine stimulates L-type Ca(2+) current in rat tail artery myocytes via a PKA-dependent mechanism. Vascul. Pharmacol. 2016, 76, 53–61. [Google Scholar] [CrossRef]
- Karmažínová, M.; Lacinová, L. Measurement of cellular excitability by whole cell patch clamp technique. Physiol. Res. 2010, 59 (Suppl. S1), S1–S7. [Google Scholar] [CrossRef]
- Iozzi, D.; Schubert, R.; Kalenchuk, V.U.; Neri, A.; Sgaragli, G.; Fusi, F.; Saponara, S. Quercetin relaxes rat tail main artery partly via a PKG-mediated stimulation of KCa1.1 channels. Acta Physiol. 2013, 208, 329–339. [Google Scholar] [CrossRef] [PubMed]
- Pessina, F.; Gamberucci, A.; Chen, J.; Liu, B.; Vangheluwe, P.; Gorelli, B.; Lorenzini, S.; Spiga, O.; Trezza, A.; Sgaragli, G.; et al. Negative chronotropism, positive inotropism and lusitropism of 3,5-di-t-butyl-4-hydroxyanisole (DTBHA) on rat heart preparations occur through reduction of RyR2 Ca2+ leak. Biochem. Pharmacol. 2018, 155, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Campiani, G.; Cavella, C.; Osko, J.D.; Brindisi, M.; Relitti, N.; Brogi, S.; Saraswati, A.P.; Federico, S.; Chemi, G.; Maramai, S.; et al. Harnessing the Role of HDAC6 in Idiopathic Pulmonary Fibrosis: Design, Synthesis, Structural Analysis, and Biological Evaluation of Potent Inhibitors. J. Med. Chem. 2021, 64, 9960–9988. [Google Scholar] [CrossRef] [PubMed]
- Federico, S.; Khan, T.; Fontana, A.; Brogi, S.; Benedetti, R.; Sarno, F.; Carullo, G.; Pezzotta, A.; Saraswati, A.P.; Passaro, E.; et al. Azetidin-2-one-based small molecules as dual hHDAC6/HDAC8 inhibitors: Investigation of their mechanism of action and impact of dual inhibition profile on cell viability. Eur. J. Med. Chem. 2022, 238, 114409. [Google Scholar] [CrossRef]
- Ferrara, A.; Fusi, F.; Gorelli, B.; Sgaragli, G.; Saponara, S. Effects of freeze-dried red wine on cardiac function and ECG of the Langendorff-perfused rat heart. Can. J. Physiol. Pharmacol. 2014, 92, 171–174. [Google Scholar] [CrossRef]
- Fusi, F.; Durante, M.; Gorelli, B.; Perrone, M.G.; Colabufo, N.A.; Saponara, S. MC225, a Novel Probe for P-glycoprotein PET Imaging at the Blood–brain Barrier: In Vitro Cardiovascular Safety Evaluation. J. Cardiovasc. Pharmacol. 2017, 70, 405–410. [Google Scholar] [CrossRef]
- Trezza, A.; Spiga, O.; Mugnai, P.; Saponara, S.; Sgaragli, G.; Fusi, F. Functional, electrophysiology, and molecular dynamics analysis of quercetin-induced contraction of rat vascular musculature. Eur. J. Pharmacol. 2022, 918, 174778. [Google Scholar] [CrossRef]
- Janson, G.; Paiardini, A. PyMod 3: A complete suite for structural bioinformatics in PyMOL. Bioinformatics 2021, 37, 1471–1472. [Google Scholar] [CrossRef]
- Tao, X.; MacKinnon, R. Molecular structures of the human Slo1 K+ channel in complex with β4. eLife 2019, 8. [Google Scholar] [CrossRef] [PubMed]
- Laskowski, R.A.; MacArthur, M.W.; Moss, D.S.; Thornton, J.M. PROCHECK: A program to check the stereochemical quality of protein structures. J. Appl. Crystallogr. 1993, 26, 283–291. [Google Scholar] [CrossRef]
- Greenwood, J.R.; Calkins, D.; Sullivan, A.P.; Shelley, J.C. Towards the comprehensive, rapid, and accurate prediction of the favorable tautomeric states of drug-like molecules in aqueous solution. J. Comput. Aided. Mol. Des. 2010, 24, 591–604. [Google Scholar] [CrossRef] [PubMed]
- Friesner, R.A.; Murphy, R.B.; Repasky, M.P.; Frye, L.L.; Greenwood, J.R.; Halgren, T.A.; Sanschagrin, P.C.; Mainz, D.T. Extra Precision Glide: Docking and Scoring Incorporating a Model of Hydrophobic Enclosure for Protein−Ligand Complexes. J. Med. Chem. 2006, 49, 6177–6196. [Google Scholar] [CrossRef] [PubMed]
- Salentin, S.; Schreiber, S.; Haupt, V.J.; Adasme, M.F.; Schroeder, M. PLIP: Fully automated protein–ligand interaction profiler. Nucleic Acids Res. 2015, 43, W443–W447. [Google Scholar] [CrossRef] [PubMed]
- Berendsen, H.J.C.; van der Spoel, D.; van Drunen, R. GROMACS: A message-passing parallel molecular dynamics implementation. Comput. Phys. Commun. 1995, 91, 43–56. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cpds | Rat Aorta Rings | Rat Tail Artery Myocytes | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|
| KCl | Phenylephrine | IBa1.2 | ||||||||
| 25 mM | 60 mM | −Endothelium | +Endothelium | |||||||
| IC50 (µM) | Emax (%) | IC50 (µM) | Emax (%) | IC50 (µM) | Emax (%) | IC50 (µM) | Emax (%) | IC50 (µM) | Emax (%) | |
| 5 | N.D. | 45.9 ± 8.5 (6) | 29.2 ± 6.1 (5) | 73.5 ± 6.8 * (5) | N.D. | 29.8 ± 13.9 (5) | N.D. | +287.1 ± 48.8 * (6) | 69.0 a | 64.8 ± 6.1 (6) |
| 6 | 11.9 ± 1.3 (5) | 98.8 ± 1.3 (5) | 33.2 ± 6.8 (5) | 88.7 ± 4.0 (5) | ||||||
| 7 | 0.2 ± 0.1 (7) | 94.3 ± 4.5 (3) | 9.3 ± 2.0 * (6) | 89.7 ± 1.9 (5) | N.D | 44.2 ± 3.8 (5) | N.D. | 8.6 ± 9.3 * (6) | 13.2 ± 5.4 (6) | 90.5 ± 5.8 (6) |
| 8 | 2.1 ± 0.9 (8) | 85.8 ± 4.8 (8) | 6.1 ± 1.6 * (5) | 87.3 ± 4.9 (5) | 44.7 a | 59.9 ± 10.2 (7) | N.D. | 15.2 ± 7.9 * (6) | 16.7 ± 3.0 (7) | 92.4 ± 2.2 (7) |
| 9 | 0.9 ± 0.2 (5) | 92.3 ± 0.6 (5) | 4.5 ± 0.5 * (5) | 92.4 ± 1.3 (5) | 55.0 a | 61.0 ± 8.9 (5) | N.D. | 19.4 ± 23.2 (6) | 15.3 ± 5.3 (8) | 97.8 ± 0.8 (5) |
| 10 | 69.2 a | 53.1 ± 13.4 (5) | 20.8 ± 6.1 (6) | 86.4 ± 4.9 (6) | ||||||
| Cpds | V50 act (mV) | Slope act | n | V50 inact (mV) | Slope inact | n |
|---|---|---|---|---|---|---|
| Control | −2.64 ± 1.03 | 7.16 ± 0.21 | 5 | −21.34 ± 3.59 | −12.09 ± 1.36 | 5 |
| 8 10 µM | −30.97 ± 3.49 * | −11.51 ± 1.90 | 5 | |||
| 8 30 µM | 0.03 ± 1.35 | 7.61 ± 0.66 | 5 | |||
| Control | −1.97 ± 2.36 | 7.08 ± 0.32 | 6 | −23.59 ± 1.40 | −7.56 ± 0.36 | 6 |
| 7 10 µM | 1.99 ± 3.26 * | 8.36 ± 0.75 | 6 | −30.24 ± 1.29 # | −7.53 ± 0.24 | 6 |
| 7 30 µM | −38.29 ± 2.00 # | −7.70 ± 0.51 | 6 | |||
| Control | −2.88 ± 0.91 | 7.31 ± 0.27 | 6 | −22.90 ± 2.53 | −9.03 ± 1.32 | 5 |
| 9 10 µM | −2.37 ± 0.95 | 6.79 ± 0.35 | 6 | −36.46 ± 2.73 * | −10.25 ± 1.33 | 5 |
| (7) | HR (bpm) | RR (ms) | PQ (ms) | QRS (ms) | QTc (ms) |
|---|---|---|---|---|---|
| 0 | 261.9 ± 9.6 | 231.4 ± 7.8 | 37.8 ± 1.8 | 13.4 ± 0.7 | 72.97 ± 1.7 |
| 1 µM | 261.8 ± 9.4 | 230.8 ± 7.4 | 37.2 ± 2.0 | 13.6 ± 0.5 | 71.59 ± 0.8 |
| 3 µM | 256.9 ± 8.3 | 231.8 ± 7.1 | 37.4 ± 1.9 | 14.0 ± 0.6 | 72.65 ± 1.3 |
| 10 µM | 255.3 ± 8.0 | 236.4 ± 7.1 | 39.0 ± 1.9 | 13.9 ± 0.6 | 73.31 ± 1.2 |
| 30 µM | 242.8 ± 11.5 ** | 249.8 ± 10.7 ** | 42.2 ± 2.7 ** | 15.2 ± 0.8 | 75.13 ± 1.7 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carullo, G.; Saponara, S.; Ahmed, A.; Gorelli, B.; Mazzotta, S.; Trezza, A.; Gianibbi, B.; Campiani, G.; Fusi, F.; Aiello, F. Novel Labdane Diterpenes-Based Synthetic Derivatives: Identification of a Bifunctional Vasodilator That Inhibits CaV1.2 and Stimulates KCa1.1 Channels. Mar. Drugs 2022, 20, 515. https://doi.org/10.3390/md20080515
Carullo G, Saponara S, Ahmed A, Gorelli B, Mazzotta S, Trezza A, Gianibbi B, Campiani G, Fusi F, Aiello F. Novel Labdane Diterpenes-Based Synthetic Derivatives: Identification of a Bifunctional Vasodilator That Inhibits CaV1.2 and Stimulates KCa1.1 Channels. Marine Drugs. 2022; 20(8):515. https://doi.org/10.3390/md20080515
Chicago/Turabian StyleCarullo, Gabriele, Simona Saponara, Amer Ahmed, Beatrice Gorelli, Sarah Mazzotta, Alfonso Trezza, Beatrice Gianibbi, Giuseppe Campiani, Fabio Fusi, and Francesca Aiello. 2022. "Novel Labdane Diterpenes-Based Synthetic Derivatives: Identification of a Bifunctional Vasodilator That Inhibits CaV1.2 and Stimulates KCa1.1 Channels" Marine Drugs 20, no. 8: 515. https://doi.org/10.3390/md20080515
APA StyleCarullo, G., Saponara, S., Ahmed, A., Gorelli, B., Mazzotta, S., Trezza, A., Gianibbi, B., Campiani, G., Fusi, F., & Aiello, F. (2022). Novel Labdane Diterpenes-Based Synthetic Derivatives: Identification of a Bifunctional Vasodilator That Inhibits CaV1.2 and Stimulates KCa1.1 Channels. Marine Drugs, 20(8), 515. https://doi.org/10.3390/md20080515